
Abstract
Objective
The activation of NF-κB signaling and unbalance of T-helper (Th) cells have been reported to play a key role in the pathogenesis of colitis. Cortex Phellodendri Chinensis (CPC) is commonly used to treat inflammation and diarrhea. Demethyleneberberine (DMB), a component of CPC, was reported to treat alcoholic liver disease as a novel natural mitochondria-targeted antioxidant in our previous study. In this study, we investigated whether DMB could protect against dextran sulfate sodium (DSS)-induced inflammatory colitis in mice by regulation of NF-κB pathway and Th cells homeostatis.
Methods
Inflammatory colitis mice were induced by 3% DSS, and DMB were orally administered on the doses of 150 and 300 mg/kg. In vitro, DMB (10, 20, 40 μM) and N-acetyl cysteine (NAC, 5 mM) were co-cultured with RAW264.7 for 2 h prior to lipopolysaccharide (LPS) stimulation, and splenocytes from the mice were cultured ex vivo for 48 h for immune response test.
Results
In vivo, DMB significantly alleviated the weight loss and diminished myeloperoxidase (MPO) activity, while significantly reduced the production of pro-inflammatory cytokines, such as interleukin (IL)-6 and tumor necrosis factor-α (TNF-α), and inhibited the activation of NF-κB signaling pathway. Furthermore, DMB decreased interferon (IFN)-γ, increased IL-4 concentration in the mice splenocytes and the ratio of IgG1/IgG2a in the serum. In vitro, ROS production and pro-inflammation cytokines were markedly inhibited by DMB in RAW264.7 cell.
Conclusions
Our findings revealed that DMB alleviated mice colitis and inhibited the inflammatory responses by inhibiting NF-κB pathway and regulating the balance of Th cells.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Inflammatory bowel disease(IBDs), which include ulcerative colitis (UC) and Crohn’s disease, are a group of chronic, multifactorial inflammatory disorders characterized by cytokine-driven and T lymphocyte-dependent inflammation of the intestinal mucosa [1, 2]. They have a high incidence of 24.5/100,000 in western countries [3], not only impacting quality of life, but also increasing the risk of colon cancer. A lot of evidence has shown that the risk of colorectal cancer development in patients with IBD is much higher than in the general population [4]. Immunosuppressants, such as prednisone, azathioprine and methotrexate, are used to treat IBD. Because of the potential side effects of immunosuppressants, including steroid dependence [5] and serious infections [6], there is an urgent need to develop new therapeutic options and approaches for IBD.
Reactive oxygen species (ROS) plays a pivotal role in the pathogenesis of colitis and innate immune response activation [7]. Once the innate immune response is initiated, inflammatory factors derived from intestinal epithelial cells and innate immune cells such as macrophages, including increased TNF-α, IL-6 and so on, lead to exaggerated adaptive immune responses [8]. Among these signals, NF-κB is a key transcription factor responsible for inflammation and immunity of mammals and human beings [9]. Recently, it has been reported that dampening NF-κB signaling pathway attenuated immune responses and ameliorated colitis [10]. Moreover, an imbalance of Th cell type 1 (Th1) versus type 2 (Th2) appears to be a key pathogenic mechanism in colitis [11, 12]. Th1 cells produce IFN-γ and positively contribute to the humoral (antibody) response to a limited extent by supporting the production of the IgG2a antibody subclass. On the contrary, the cytokines, which include IL-4, IL-5 and IL-10, are produced in Th2 cells and involved in the stimulating IgG1 production by B cells. Furthermore, IL-4 was found to be reduced in IBD, which may cause defection of immunosuppressive and anti-inflammatory mechanism [13].
Traditional Chinese medicine Cortex Phellodendri Chinensis (CPC) has been reported with several pharmacological activities, including anti-microbial, anti-inflammatory and anti-diarrheal [14–17]. Demethyleneberberine (DMB), a component in the CPC, has been showed an obvious therapy effect to alcoholic liver disease in our previous study [18]. DMB (9, 10-dimethoxy-5, 6-dihydroisoquinolino [2, 1-b] isoquinolin-7-ium-2, 3-diol, structure was shown in Fig. 1) is an isoquinoline alkaloid, which is a natural mitochondria-targeted antioxidant with both cationic and catechol groups [18]. Research has showed that antioxidant agents ameliorate experimental colitis [19]. However, the anti-colitic effect and mechanism of action of DMB are unknown. Thus, the aim of this study was to determine the anti-oxidant and anti-inflammatory effect of DMB on the experimental colitis induced by DSS and Raw264.7 cell stimulated by lipopolysaccharide (LPS).

The chemical structure of DMB (9, 10-dimethoxy-5, 6-dihydroisoquinolino [2, 1-b] isoquinolin-7-ium-2, 3-diol)
Methods
Materials
Synthesized DMB from berberine (BBR) with slight modification from Zuo’s method in our laboratory fulfilled the requirement for cell and animal test [18, 20]. DSS: molecular weight of 36–50 kDa (MP Biomedicals, USA). Antibodies: NF-κB kit, including antibodies against p65, p-p65, IKKβ, IκB, p-IκB, was purchased from Cell Signaling Technology, Inc (China) (Shanghai, China) and GAPDH was purchased from Santa Cruz Biotechnology, Inc.(Texas, USA). ELISA kits for murine IL-6, TNF-α, IL-4, IFN-γ, IgG1 and IgG2a were purchased from Beijing 4A Biotech (Beijing, China). MPO kit was purchased from Nanjing Jiancheng Bioengineering Institute (Nanjing, China). LPS was purchased from Sigma-Aldrich (Shanghai) Trading Co., Ltd (Shanghai, China). DMEM and FBS were purchased from Gibco. All other chemicals were analytical grade.
Animals
Female C57BL/6 mice (6–8 weeks, 18–22 g) were obtained from the Model Animal Research Center of Nanjing University (Nanjing, China) and group-housed under controlled temperature (22 ± 2 °C) and photoperiods (12:12-h light–dark cycle). Mice were acclimated to these conditions for 7 days and then matched by age and body weight for each group. All procedures were approved by the Institutional Animal Care and Use Committee at China Pharmaceutical University and adhered to the Jiangsu Provincial Guidelines for the use of experimental animals.
Induction of colitis
Acute colitis mice were induced by 3% (wt/vol) of DSS dissolved in drinking water continuously from day 0 to day 7, while mice in control group received the same drinking water without DSS (n = 7 mice in each group). DMB (150, 300 mg/kg) were orally administered once a day from day −7 to day 7 (Fig. 2a). To prepare the suspension of DMB, DMB powder was weighed, then suspended in carboxymethylcellulose sodium (CMC) to the concentration of 15 mg/ml and 30 mg/ml. Mice were observed daily for weight, morbidity, stool consistency, and gross blood in feces and at the anus. Then they were killed and rapidly dissected under anesthesia by ether inhalation. Blood sampling for serum IgG1 and IgG2a test was drawn from the posterior vena cava. Then blood samples were centrifuged 5000×g for 10 min at 4 °C, and serums were stored at −70 °C until analysis. The entire colon was quickly removed, washed with ice-cold 0.9% physiological saline then saved in the liquid nitrogen container.

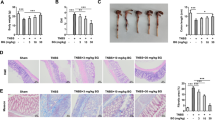
Effect of DMB on the recovery of DSS-induced colitis in mice. a The model of acute colitis in mice induced by DSS. b Body weight was measured daily throughout the experiment from day 1 to day 10. c Representative photograph showing the shortening of the colon in mice treated with DSS. d Lengths of the freshly removed colons were measured from rectum to ileocecal junction. e Colon weight was weighted and calculated by per cm of colon length. f The severity of colitis was macroscopically assessed every day. Values were shown as the mean ± SEM of seven mice. *, # <0.05, **, ## <0.01 and ***, ### <0.001. *p compared with DSS group and #p compared with the control group
Calculation of DAI
Disease activity index(DAI) is a useful composite score for following mice with colitis, which was calculated as described previously [21]. Briefly, the following parameters were used for calculation: (a) diarrhea (0 points = control, 2 points = loose stools, 4 points = watery diarrhea); (b) hematochezia (0 points = no bleeding, 2, slight bleeding, 4 points, gross bleeding).
Histopathological assay
Segments of colons were fixed in 10% formaldehyde solution, embedded in paraffin, sectioned to 4 μm thickness, and then stained with H&E for light microscopic examination. Histological scores were calculated to assess colitis by a modified combined scoring system, which includes degree of inflammation (scale of 0–2), depth of inflammation (0–3), crypt damage (0–4) and percentage of area involved by inflammation (0–4) [11]. The total score is 0 (normal) to 13 (severe colitis).
Assay for MPO and antioxidant activity
Colon tissues were homogenized in 10 mM PBS (pH 7.4) and then centrifuged at 10,000×g at 4 °C for 15 min. The supernatants were used to determine MPO level according to manufacturer’s instructions. The MDA concentration was estimated as described by Ohkawa et al. [22], while the amount of GSH was measured according to the method of Paglia and Valentine [23].
Culture and treatment of RAW 264.7 cells and splenocytes
RAW 264.7 macrophage cells were obtained from the Institute of Chinese Academy of Sciences in Shanghai, and cultured in DMEM medium containing 10% FBS, 100 U/ml penicillin and 100 μg/ml streptomycin at 37 °C with 5% CO2. DMB (10, 20, or 40 μM) were dissolved in DMSO (0.4‰) and added 2 h prior to 100 ng/ml LPS for 24 or 6 h. After incubation for 24 h, the culture mediums were collected for ELISA.
Splenocytes were obtained from spleens and then dispersed by tapping gently with a rubber rod on a sterile stainless sieve. The red blood cells in the suspended cells were lysed with lysis buffer (Sigma, USA) in an ice bath for 10 min. Washed twice with PBS, and then cells were cultured for 4 h in DMEM containing 10% FBS, 25 mM HEPES, 2 mM glutamine, 100 U/ml penicillin and 100 μg/ml streptomycin. Splenocytes were plated into 24-well plates at a density of 1 × 106 cells/ml, and treated with DMB (10, 20, or 40 μM) for 48 h.
Measurement of intracellular ROS
Intracellular oxidative stress was detected using DCFH-DA as described by Lemmens KJA et al. [24]. Briefly, RAW264.7 cells (2 × 105 cells/well) were inoculated in a 24-well plate and incubated to reach 80% confluence. The cells were pre-cultured for 2 h with DMB (10, 20, or 40 μM) and N-acetyl cysteine (NAC) (5 mM), and then stimulated with 100 ng/ml LPS for 24 h or 2 mM H2O2 for 2 h. Then washed twice with PBS, the cells were incubated with 10 μM DCFH-DA and 10 μg/ml Hoechst in the dark for 30 min. The harvested cells were immediately analyzed by the flow cytometry or fluorescent microscope after washed twice.
NO assay
NO productions in supernatants of RAW264.7 cell and colon tissues were determined with a colorimetric assay based on the Griess reaction [25]. The supernatants (100 μl) were reacted with 100 μl Griess reagent (6 mg/ml) at room temperature for 10 min, and then NO2 − concentration was determined by measuring absorbance at 540 nm. A standard curve was constructed using known concentrations of sodium nitrite.
ELISA of colon tissues, serum and cells
The colon homogenate and culture mediums of splenocytes and RAW264.7 cells were collected and centrifuged (12,000×g, 4 °C) for 15 min. The production of IL-1β, IL-6, TNF-a and IL-10 in colon tissues and RAW264.7 cell, IFN-γ and IL-4 in the splenocytes and IgG1 and IgG2a in the serum were determined by commercial ELISA kits.
Immunofluorescence histochemistry assay
Cryosections (4 μm) of colons were fixed in acetone, stained with anti-p65 (1:400) at room temperature for 2 h and then stained with DAPI and imaged with a confocal laser scanning microscope (Olympus, Japan).
RNA isolation and RT-PCR assay
RAW264.7 cells were seeded at 1 × 106/ml in a 6-well plate, and then treated with DMB (10, 20, or 40 μM) for 2 h prior to LPS (100 ng/ml) stimulation for an additional 6 h when reached 80% confluence. RNA isolation and RT-PCR assay were conducted according to Wen et al. [26]. The primer sequences used in RT-PCR were listed in Table 1. Gene expression levels were calculated by the △△Ct method and the values were normalized to endogenous reference cyclophilin.
Western blotting analysis
Colons were homogenized in 1 ml of ice-cold RIPA lysis buffer containing 1% protease inhibitor cocktail (Roche, Switzerland) and centrifuged (12,000×g, 4 °C) for 15 min. The protein concentration was determined with bicinchoninic acid (BCA) method. Equal amounts of total protein (100 μg) were subjected to electrophoresis on 10% SDS-PAGE and then transferred to PVDF membranes (Millipore, USA). Levels of IκB, p-IκB, IKKβ, p-p65, p65 and GAPDH were assayed as previously described [26]. ImageJ 64 software was used to analyze the relative density of each band.
Statistical analysis
Statistical significance for multiple comparisons in each study was determined by one-way ANOVA followed by Newman-Keuls analysis using Prism 5.0 (GraphPad Software). P value <0.05 was defined as statistically significant. All data presented are representative of at least five repeat experiments with means SE.
Results
Effect of DMB on the recovery of DSS-induced colitis in mice
DSS-induced mice lost weight significantly starting from the fourth day. However, the oral administration of DMB significantly alleviated weight loss and the body weight began to recover from day 7 to day 10 in the DMB group (Fig. 2b). Contrast to the obvious increase in the colon wall thickness and DAI scores in the DSS group, a remarkable attenuation of these responses was found in the colons of mice treated with either 150 or 300 mg/kg of DMB (Fig. 2f). Moreover, DMB exerted preventive effects on the colon, including the colon weight and colon length (Fig. 2c–e).
Effect of DMB on the DSS-induced colon injury in mice
The results of H&E staining indicated that DSS treatment accelerated initial damage to the mucosa, characterized by the loss of goblet cells and the occurrence of a more diffuse crypt architecture compared to the colon of control group (Fig. 3a). Additionally, the severity of inflammation seemed to be significantly lower after treated with DMB (Fig. 3b). Furthermore, increasing colonic MPO activity with the neutrophil infiltration in the colon is treated as an inflammatory marker for colitis. The MPO activity was reduced significantly in a dose-dependent manner in the mice with the treatment with DMB (Fig. 3c).

Effect of DMB on the colon damage in mice induced by DSS. Sections of the colon were collected and stained with H&E. Representative histological photo (a) and histological score (b) for mice in each group were presented. Protein from colon tissue was extracted and MPO activity was determined (c). Values were shown as the mean ± SEM of seven mice. *, # <0.05, **, ## <0.01 and ***, ### <0.001. *p compared with DSS group and #p compared with the control group
Effect of DMB on the oxidative damage in colon and ROS in the Raw264.7 cell
MDA is an end product of lipid peroxidation induced by ROS, while GSH is the most important intracellular defense against oxidative stress and is essential for both the functional and structural integrity of the gut [27]. Our current data showed that DMB treatment reduced the concentration of MDA, but increased the GSH content (Fig. 4a, b). To further investigate the antioxidant activity of DMB, we evaluated the generation of ROS in the RAW264.7 cell. When stimulated with LPS or H2O2, the intracellular ROS level in RAW264.7 cell increased with the time, but pretreatment with DMB notably prevented the LPS-induced ROS generation (Fig. 4c–e). Moreover, DMB play an important antioxidant effect in T-AOC test and LPS-induced RAW264.7 cell at the concentration of 5 mM (Fig. S1a–c). As a well-established ROS scavenger, NAC treatment also showed a significant inhibitory LPS or H2O2-induced ROS overproduction.

Effect of DMB on the oxidative damage in colon and ROS in the vitro experiment. GSH and MDA were measured in colon homogenates. The results were shown in (a) and (b) respectively. The ROS productions in RAW264.7 cell were measured by fluorescent microscope with Hoechst and DCFH-DA (c). The ROS generations induced by LPS in the RAW264.7 cell was calculated with the flow cytometry (d, e). Values were shown as the mean ± SEM of five independent tests. *, # <0.05, **, ## <0.01 and ***, ### <0.001. *p versus control group and #p versus LPS/DSS group
Effect of DMB on the NF-κB signaling of the colon
The transcription factor NF-κB is a central mediator in inflammation and several other cellular responses. Western blotting analysis was used to further confirm our results. DMB treatment caused a dose-dependent decrease in the key proteins expression of NF-κB signaling pathway in colon tissues, including IKKβ, p-p65 and p-IκB (Fig. 5a–c). In addition, the expression of IκB protein increased after DMB treatment (Fig. 5c).

Effect of DMB on the activation of NF-κB signaling in mice colon. a–c The key protein expressions of NF-κB signaling were examined by Western blotting, including IKKβ, p65 and p-p65, IκB and p-IκB. The relative densities of western blotting band were compared with the GAPDH band in each group. *, # <0.05, **, ## <0.01 and ***, ### <0.001. *p versus Control group and #p versus DSS group
Effect of DMB on the production of inflammatory cytokines in mice and RAW264.7 cell
The DSS-induced acute colitis is well-characterized by increased epithelial injury and production of inflammatory cytokines [9]. Intestinal bacterial endotoxin LPS penetrate the epithelial barrier and directly stimulate the mucosal immune system [28]. The results demonstrated elevated levels of pro-inflammatory cytokines production, including TNF-a, IL-6 and NO in the colon and TNF-a, IL-6, IL-1β and NO in the RAW264.7 cell, were reduced significantly (Fig. 6a–b). Consistent with the increased content of cytokines in the cell supernatant, stimulation with 100 ng/ml LPS for 6 h caused a significant increase in the mRNA expression of pro-inflammatory cytokines (Fig. 6c). However, Pretreatment with DMB diminished the mRNA expression of pro-inflammatory cytokines, while it remarkably increased IL-10 expression in RAW264.7 cells in dose dependent (Fig. 6c).

Effect of DMB on the production of inflammatory cytokines in DSS-induced colitis mice and LPS-stimulated RAW264.7 cell. The pro-inflammatory cytokines (TNF-α, IL-6 and IL-1β) in supernatants of colon tissues and RAW264.7 cells were collected and measured by ELISA, while NO expression measured by experiment kit (a, b). The mRNA expression of pro-inflammatory cytokines, including TNF-α, IL-6, iNOS, COX-2 and IL-10, were measured by RT- PCR (c). Values were shown as the mean ± SEM of five independent tests. *, # <0.05, **, ## <0.01 and ***, ### <0.001. *p versus Control group and #p versus LPS or DSS group
Effect of DMB on the homeostasis of Th cells in splenocytes
To investigate the function of DMB on Th cells, we analyzed the effects of DMB on the production of cytokines such as IFN-γ and IL-4 in splenocytes. Mice in DMB group showed lower production of IFN-γ and higher production of IL-4 compared with mice in control group. Furthermore, DMB regulated the production of IFN-γ, IL-4 and the ratio of IgG1/IgG2a (Fig. 7b–d). To the immune tissues, DMB treatment decreased the spleen weight, while increased the thymus weight (Fig. 7a).

Effect of DMB on the weight of thymus and spleen and disturbance in splenocytes. The test agents were orally administered once every day for 7 days prior to DSS treatment. The mice were anesthetized with ether and killed 3 days after DSS treatment. The spleen and thylm were isolated and weighed, meanwhile splenic lymphocytes were collected, and IFN-γ, IL-4, IgG2a and IgG1 levels were measured by ELISA. The results were shown in (a), (c) and (d) respectively. The ratio of IgG2a and IgG1 was calculated and shown in (b). *, # <0.05, **, ## <0.01 and ***, ### <0.001. *p versus control group and #p versus LPS
Discussion
IBD is a chronic inflammatory disorder in the gastrointestinal tract. In our research, DMB showed a significant antioxidant effect in LPS/H2O2-stimulated RAW264.7 cell and DSS-induced mice. Furthermore, DMB inhibited NF-κB signaling and helped homeostatic maintenance of Th cells in colitis mice. These findings indicate that DMB alleviates colitis through antioxidant effect and ultimately diminishing the inflammatory activity.
Most studies conducted in the last thirty years have focused on the role of adaptive immune responses in the pathogenesis of IBD. In particular, while CD has long been considered to be driven by a Th1 response, UC has been rather associated with a non-conventional Th2 response [29]. In mice, production of IgG1 versus IgG2a is widely interpreted as a reflection of differential Th2–Th1 reactivity [30]. To our surprise, we first found the imbalance of Th cells in the spleen of DSS-induced mice colitis. One of the characteristic macroscopic changes associated with DSS colitis is enlargement of the spleen. A study suggest a close correlation between spleen size and severity of DSS colitis [31]. In our results, oral administration of DMB obvious inhibited the spleen enlargement. In order to clarify whether treatment with DMB regulated the balance of Th cell, we analyzed cytokine production in the splenocyte. Contrary with previous study in the colon, our results showed that IFN-γ concentration was higher, whereas IL-4 concentration was lower in mice spleen with DSS-induced colitis than that in control mice. We demonstrated that increased IFN-γ and decreased IL-4 could be reversed by treatment with DMB. Furthermore, DMB evoked dramatically higher IgG1/IgG2a ratios relating to increased activity of the Th2-pathway. These dates may suggest that DMB suppressed the Th1 responses and enhanced Th2 responses and prevented the unbalance of Th1 and Th2 in the spleen of DSS-induced colitis.
With an approximately 17% sulfur, DSS is associated with a marked increase in the measurable ROS and the activation of an inflammatory cascade [32]. Researchers have showed agents capable of eliminating reactive oxygen species ameliorate experimental colitis [33]. To ensure the antioxidant capacity of DMB in vitro, RAW264.7 cells were induced by LPS or H2O2. Of note, we found that pro-incubated DMB reduced ROS production in dose-dependent in LPS/H2O2-stimulated RAW264.7 cells. We also observed the same anti-oxidant and anti-inflammatory effect of DMB on the LPS/H2O2-induced human colonic epithelial cell HT29 (The results were not shown). Furthermore DMB was more effective than NAC at the concentration of 40 μM in the RAW264.7 cell induced by LPS. The part reasons maybe LPS triggers the production of ROS and promotes acute inflammation while NAC is known to act by raising intracellular concentrations of cysteine and scavenging ROS [34, 35]. Based on the antioxidant effect of DMB on macrophage and anti-diarrhea of CPC, we propose a hypothesis that DMB may alleviate acute mice colitis induced by DSS by ameliorating oxidant stress. Consistent with our vitro results, the current data show DMB plays a significant in antioxidant effect in colitis mice through marked decreased MDA content and increased GSH activity. DMB not only scavenges the production of ROS, but plays a crucial role to the reduction of inflammatory cytokines in the Raw264.7 cell. To all known, production of cytokines activate transcription factors such as NF-κB [36]. It was also reported that ROS mediated activation of NF-κB in macrophage and colitis [37]. In addition, inhibited NF-κB helps the prevention of colitis [38]. Our date showed DMB reduced colon inflammation, such as neutrophil infiltration and multiple erosions, and inhibited the activation of NF-κB signaling in colon in response to DSS-induced mice colitis. As NF-κB is the down signaling of TLR4 receptor [39, 40], DMB treatment down regulated the expression of TLR4 in mice colon (Fig. S6 a, b). Although we still did not know the time course of activation of NF-κB in colitis mice, the time course of p-p65 expression and inflammatory cytokines expression in RAW264.7 cells were explored (Figs. S2, S3a, b). Consistent with the vivo results, DMB decreased the nuclear transcript of p65 (Fig. S4a, b) and the expression of inflammatory cytokines in Raw264.7 stimulated by LPS significantly, such as TNF-a, IL-6, IL-1β, NO and so on. As DMB was absorbed in cells quickly with the HPLC analysis (Table S1, Fig. S5a–e), we also researched the toxicity in mice (Table S2) and tissue distribution of DMB in rat (Tables S3, S4, Fig. S7a–d). The date indicated that DMB distributed in the brain and lung as short as 5 min, and then mainly accumulated in the liver at the time of 30 min (Fig. S7a–d). In the toxicity experiment, part of mice appeared with convulsions, shortness of breath, dark limbs and tail skin immediately and then died within 5–10 min. However, no abnormal organs were found with the naked eye observation.
In summary, our data first indicated that DMB exerts therapeutic effect on DSS-induced mice colitis. The potential mechanisms may contribute to inhibiting oxidant stress and NF-κB signaling pathway, as well as partly regulating the balance of Th1 versus Th2 in spleen. These findings provide new point on DMB and its possible use in intestinal inflammatory disorder.
Abbreviations
- CMC:
-
Carboxymethylcellulose sodium
- CPC:
-
Cortex Phellodendri Chinensis
- DAI:
-
Disease activity index
- DMB:
-
Demethyleneberberine
- GSH:
-
Glutathione
- H&E:
-
Hematoxylin and eosin
- IBDs:
-
Inflammatory bowel diseases
- IFN:
-
Interferon
- IL:
-
Interleukin
- LPS:
-
Lipopolysaccharide
- MDA:
-
Malondialdehyde
- MPO:
-
Myeloperoxidase
- NAC:
-
N-acetyl cysteine
- ROS:
-
Reactive oxygen species
- Th:
-
T helper
- Th1:
-
Th cell type 1
- Th2:
-
Th cell type 2
- TNF-α:
-
Tumor necrosis factor-α
- UC:
-
Ulcerative colitis
References
Vincent B, Allen PB, Laurent PB. Update on Crohn’s disease and ulcerative colitis. Expert Rev Gastroenterol Hepatol. 2014;5:311–4.
Villablanca EJ, Cassani B, von Andrian UH, Mora JR. Blocking lymphocyte localization to the gastrointestinal mucosa as a therapeutic strategy for inflammatory bowel diseases. Gastroenterology. 2011;140:1776–84.
Lakatos L, Lakatos PL. Is the incidence and prevalence of inflammatory bowel diseases increasing in Eastern Europe? Postgrad Med J. 2006;82:332–7.
Terzić J, Grivennikov S, Karin E, Karin M. Inflammation and colon cancer. Gastroenterology. 2010;138(2101–2114):e5.
Yamamoto-Furusho JK, Rodríguez-Bores L, Granados J. HLA-DRB1 alleles are associated with the clinical course of disease and steroid dependence in Mexican patients with ulcerative colitis. Colorectal Dis. 2010;12:1231–5.
Andersen NN, Jess T. Risk of infections associated with biological treatment in inflammatory bowel disease. World J Gastroenterol. 2014;20:16014–9.
Balmus IM, Ciobica A, Trifan A, Stanciu C. The implications of oxidative stress and antioxidant therapies in inflammatory bowel disease: clinical aspects and animal models. Saudi J Gastroenterol. 2016;22:151–62.
Davies JM, Abreu MT. The innate immune system and inflammatory bowel disease. Scand J Gastroenterol. 2015;50:24–33.
Gasparini C, Feldmann M. NF-κB as a target for modulating inflammatory responses. Curr Pharm Des. 2012;18:5735–45.
Bai A, Ma AG, Yong M, Weiss CR, Ma Y, Guan Q, et al. AMPK agonist downregulates innate and adaptive immune responses in TNBS-induced murine acute and relapsing colitis. Biochem Pharmacol. 2010;80:1708–17.
Dieleman LA, Palmen MJ, Akol H, Bloemena E, Pena AS, Meuwissen SG, et al. Chronic experimental colitis induced by dextran sulphate sodium (DSS) is characterized by Th1 and Th2 cytokines. Clin Exp Immunol. 1998;114:385–91.
Yabing Z, Rui Z, Feng Z, Hong C, Bing X. Total glucosides of peony attenuates 2,4,6-trinitrobenzene sulfonic acid/ethanol-induced colitis in rats through adjustment of TH1/TH2 cytokines polarization. Cell Biochem Biophys. 2014;68:83–95.
Karttunnen R, Breese E, Walker-Smith J, MacDonald T. Decreased mucosal interleukin-4 (IL-4) production in gut inflammation. J Clin Pathol. 1994;47:1015–8.
Meng-Li C, Yan-Fang X, Siu-Po I, Sam-Hip T, Ji-Yong Y, Chun-Tao C. Chemical and biological differentiation of Cortex Phellodendri Chinensis and Cortex Phellodendri Amurensis. Planta Med. 2010;76:1530–5.
Sun H, Wang H, Zhang A, Yan G, Han Y, Li Y, et al. Chemical discrimination of Cortex Phellodendri Amurensis and Cortex Phellodendri Chinensis by multivariate analysis approach. Pharmacogn Mag. 2016;12(45):41–9.
Yan-Fang X, Qing-Qiu M, Siu-Po I, Zhi-Xiu L, Chun-Tao C. Comparison on the anti-inflammatory effect of Cortex Phellodendri Chinensis and Cortex Phellodendri Amurensis in 12-O-tetradecanoyl-phorbol-13-acetate-induced ear edema in mice. J Ethnopharmacol. 2011;137:1425–30.
Yan-Fang X, Zhi-Xiu L, Siu-Po I, Zi-Ren S, Jian-Nan C, Xiao-Ping L. Comparison the neuropreotective effect of Cortex Phellodendri chinensis and Cortex Phellodendri amurensis against beta-amyloid-induced neurotoxicity in PC12 cells. Phytomedicine Int J Phytother Phytopharmacol. 2013;20:187–93.
Zhang P, Qiang X, Zhang M, Ma D, Zhao Z, Zhou C, et al. Demethyleneberberine, a natural mitochondria-targeted antioxidant, inhibits mitochondrial dysfunction, oxidative stress, and steatosis in alcoholic liver disease mouse model. J Pharmacol Exp Ther. 2015;352:139–47.
Arthur W, Asa VK, Van P, Catherine MM, Ida S, Joshua L, et al. Targeting mitochondria-derived reactive oxygen species to reduce epithelial barrier dysfunction and colitis. Am J Pathol. 2014;184:2516–27.
Zuo F, Nakamura N, Akao T, Hattori M. Pharmacokinetics of berberine and its main metabolites in conventional and pseudo germ-free rats determined by liquid chromatography/ion trap mass spectrometry. Drug Metab Dispos. 2006;34:2064–72.
Alex P, Zachos NC, Nguyen T, Gonzales L, Chen TE, Conklin LS, et al. Distinct cytokine patterns identified from multiplex profiles of murine DSS and TNBS-induced colitis. Inflamm Bowel Dis. 2009;15:341–52.
Ohkawa H, Ohishi N, Yagi K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biochem. 1979;95:351–8.
Paglia DE, Valentine WN. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J Lab Clin Med. 1967;70:158–69.
Lemmens KJA, Vaes N, Vijgh WJFVD, Bast A, Haenen GRMM. The antioxidant flavonoid monoHER effectively protects against oxidative stress in a cultured endothelial cell line at physiologically achievable concentrations. Free Radic Biol Med. 2012;53:S94–5.
Sherman MP, Aeberhard E, Wong VZ, Griscavage J, Ignarro L. Pyrrolidine dithiocarbamate inhibits induction of nitric oxide synthase activity in rat alveolar macrophages. Biochem Biophys Res Commun. 1993;191:1301–8.
Wen X, Wu J, Chang JS, Zhang P, Wang J, Zhang Y, et al. Effect of exercise intensity on isoform-specific expressions of NT-PGC-1α mRNA in mouse skeletal muscle. BioMed Res Int. 2014;2014:402175.
Aw TY. Intestinal glutathione: determinant of mucosal peroxide transport, metabolism, and oxidative susceptibility. Toxicol Appl Pharmacol. 2005;204:320–8.
Jarry A, Bossard C, Bou-Hanna C, Masson D, Espaze E, Denis MG, et al. Mucosal IL-10 and TGF-β play crucial roles in preventing LPS-driven, IFN-γ–mediated epithelial damage in human colon explants. J Clin Investig. 2008;118:1132–42.
Fuss IJ, Neurath M, Boirivant M, Klein JS, De la Motte C, Strong SA, et al. Disparate CD4 + lamina propria (LP) lymphokine secretion profiles in inflammatory bowel disease. Crohn’s disease LP cells manifest increased secretion of IFN-gamma, whereas ulcerative colitis LP cells manifest increased secretion of IL-5. J Immunol. 1996;157:1261–70.
Maassen CB, Boersma WJ, van Holten-Neelen C, Claassen E, Laman JD. Growth phase of orally administered Lactobacillus strains differentially affects IgG1/IgG2a ratio for soluble antigens: implications for vaccine development. Vaccine. 2003;21:2751–7.
Krieglstein CF, Cerwinka WH, Laroux FS, Grisham MB, Schürmann G, Brüwer M, et al. Role of appendix and spleen in experimental colitis. J Surg Res. 2001;101:166–75.
Bhattacharyya S, Dudeja PK, Tobacman JK. ROS, Hsp27, and IKKβ mediate dextran sodium sulfate (DSS) activation of IκBa, NFκB, and IL-8. Inflamm Bowel Dis. 2009;15:673–83.
You Y, Fu J-J, Meng J, Huang G-D, Liu Y-H. Effect of N-acetylcysteine on the murine model of colitis induced by dextran sodium sulfate through up-regulating PON1 activity. Dig Dis Sci. 2009;54:1643–50.
Bai A, Guo Y, Lu N. The effect of the cholinergic anti-inflammatory pathway on experimental colitis. Scand J Immunol. 2007;66:538–45.
Soltan-Sharifi MS, Mojtahedzadeh M, Najafi A, Khajavi MR, Rouini MR, Moradi M, et al. Improvement by N-acetylcysteine of acute respiratory distress syndrome through increasing intracellular glutathione, and extracellular thiol molecules and anti-oxidant power: evidence for underlying toxicological mechanisms. Hum Exp Toxicol. 2007;26:697–703.
Grivennikov SI, Greten FR, Karin M. Immunity, inflammation, and cancer. Cell. 2010;140:883–99.
Zhang J, Dou W, Zhang E, Sun A, Ding L, Wei X, et al. Paeoniflorin abrogates DSS-induced colitis via a TLR4-dependent pathway. Am J Physiol Gastrointest Liver Physiol. 2014;306:G27–36.
Lee C, Chun J, Hwang SW, Kang SJ, Im JP, Kim JS. Enalapril inhibits nuclear factor-κB signaling in intestinal epithelial cells and peritoneal macrophages and attenuates experimental colitis in mice. Life Sci. 2014;95:29–39.
Dou W, Zhang J, Sun A, Zhang E, Ding L, Mukherjee S, et al. Protective effect of naringenin against experimental colitis via suppression of Toll-like receptor 4/NF-κB signalling. Br J Nutr. 2013;110:599–608.
Rashidian A, Muhammadnejad A, Dehpour A-R, Mehr SE, Akhavan MM, Shirkoohi R, et al. Atorvastatin attenuates TNBS-induced rat colitis: the involvement of the TLR4/NF-kB signaling pathway. Inflammopharmacology. 2016;24:109–18.
Acknowledgements
This work was supported by National Natural Science Foundation of China Grant 81573484 to Y.B.Z., Opening Project of Shanghai Key Laboratory of Complex Prescription (Shanghai University of Traditional Chinese Medicine) (14DZ2271000), A Project Funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD) and College Students Innovation Project for the R&D of Novel Drugs (J1030830).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
There are no competing interests to declare.
Additional information
Responsible Editor: Liwu Li.
Y.-Y. Chen and R.-Y. Li contributed equally to this work.
Electronic supplementary material
Below is the link to the electronic supplementary material.
11_2016_1005_MOESM2_ESM.tif
Fig.S1 DMB was determined to be a strong antioxidant in vitro and vivo. (a) Results of ROS fluorescent staining with DCFH-DA with the flow cytometry. (b) Flow cytometric histograms of A. (c) T-AOC of DMB and BBR. ***, ###< 0.001. * p versus control group and # p versus LPS group. (TIFF 1300 kb)
11_2016_1005_MOESM4_ESM.tif
Fig.S3 The time course of mRNA expression in RAW264.7 cell stimulated with 100 ng/ml or 500 ng/ml LPS for 6 h or 12 h. (a) The mRNA expression of IL-1β and TNF-α in RAW264.7 cell stimulated with 100 ng/ml or 500 ng/ml LPS for 6 h or 12 h. (TIFF 1138 kb)
11_2016_1005_MOESM5_ESM.tif
Fig.S4 The effect of DMB on the expression of p65 in the nucleus of RAW264.7 cell. (a) The western blotting result of p65 in the nucleus of RAW264.7 cell (b) The relative densities of western blotting band were compared with the Histon H3 band in each group. Values were shown as the means ± SEM of five independent tests. **<0.01 and ***, ###< 0.001. * p versus Control group and # p versus LPS group. (TIFF 1049 kb)
11_2016_1005_MOESM6_ESM.tif
Fig.S5 The HPLC analysis of DMB which move into the RAW264.7 cells. (a) The HPLC analysis of DMB standard and methanol solvent. (b) The HPLC analysis of control group in collected RAW264.7 cell which cultured without DMB. (c) The HPLC analysis of DMB and the third time washing PBS in collected RAW264.7 cell which cultured with DMB for 2 h at 2 mM concentration. (d-f) The HPLC analysis of DMB and the third time washing PBS in collected RAW264.7cell which cultured with DMB for 0.5, 1h and 2h at 2mM concentration (TIFF 1085 kb)
11_2016_1005_MOESM7_ESM.tif
Fig.S6 Effect of DMB on the expression of TLR4 in mice colon. (a) The protein expressions of TLR4 were examined by Western blotting. (b) The relative densities of western blotting band were compared with the β-actin band in each group. *<0.05 and ***, ###< 0.001. * p versus Control group and # p versus DSS group. (TIFF 743 kb)
11_2016_1005_MOESM8_ESM.tif
Fig.S7 The distribution of DMB in the rat tissues at different time. (a) Methanol HPLC results. (b - c) The DMB distribution in lung and brain at the 5 min. (d) The HPLC results of DMB distribution in the liver at the 30 min. (TIFF 608 kb)
Rights and permissions
About this article

Cite this article
Chen, YY., Li, RY., Shi, MJ. et al. Demethyleneberberine alleviates inflammatory bowel disease in mice through regulating NF-κB signaling and T-helper cell homeostasis. Inflamm. Res. 66, 187–196 (2017). https://doi.org/10.1007/s00011-016-1005-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00011-016-1005-3