
Abstract
Droughts are large-scale perturbations that affect freshwater ecosystems worldwide. A water level reduction caused by drought is an important driving factor of phytoplankton dynamics. It has been suggested that a water level reduction alters the light and mixing regime and increases nutrient concentrations and phytoplankton biomass favoring cyanobacterial blooms. We took advantage of two exceptionally dry years in the Brazilian semi-arid region to investigate the effects of the water level reduction on the water quality and phytoplankton communities of two shallow man-made lakes. In both lakes, the water level was reduced by half, while the water turbidity, conductivity, and nutrient concentrations increased. In the deeper lake, the phytoplankton biomass increased and was dominated by a cyanobacteria group as expected, but it decreased in the shallower lake and was dominated by mixotrophic flagellate groups. This was because of sediment resuspension by wind and fish facilitated by a water level reduction and increased the water turbidity more strongly in the shallower than in the deeper lake. Therefore, a water level reduction caused by a drought may either increase or decrease the phytoplankton biomass and cyanobacteria dominance in tropical shallow lakes depending on the lake depth and the concentration of inorganic suspended sediments.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Droughts are generally recognized as an ecological perturbation in ecosystems and are strongly related to reduced precipitation (Lake, 2003; Mishra & Singh, 2010). Climate changes are expected to increase the frequency and intensity of droughts in semi-arid regions (IPCC, 2007; Marengo et al., 2010). Large-scale intensive droughts have been observed in all continents around the world (Ross & Lott, 2003; Bates et al., 2008). The negative water balance (i.e., evaporation higher than precipitation) during drought periods usually reduces the water level of lakes, increasing nutrient concentrations and enhancing the proliferation and dominance of cyanobacteria (Naselli-Flores et al., 2007, Paerl & Huisman, 2009). On the other hand, the water level reduction may also increase sediment resuspension and therefore inorganic turbidity, decreasing light availability and the phytoplankton biomass (Jeppesen et al., 2015) and changing the phytoplankton community structure (Allende et al., 2008).
Phytoplankton species have different strategies to survive under different trophic conditions. The investigation of environmental conditions (e.g., light and nutrient availability, mixing regime, and hydrological events) and biological factors (e.g., herbivory) are important to understand phytoplankton ecology, including relevant processes in species replacement and habitat selection (Reynolds, 2006; Zohary et al., 2010). In eutrophic systems, high nutrient concentrations and low light availability are the main factors related to species selection (Reynolds, 1998). Many genera of cyanobacteria have adaptive advantages to survive under these conditions, enabling them to dominate phytoplankton in eutrophic systems, such as the ability to fix atmospheric nitrogen (N2) and to stock phosphorous and maintain high rates of photosynthesis under low light conditions (Bonilla et al., 2012; Paerl & Otten, 2013). Previous studies on phytoplankton dynamics in semi-arid regions have shown cyanobacterial blooms during most of the year as a consequence of droughts (Bouvy et al., 1999; Naselli-Flores et al., 2007; Dantas et al., 2012).
Ecologists often group organisms with similar traits and function to simplify the complexity of the ecosystem. Phytoplankton communities can be characterized either by their taxonomic or functional group composition (Salmaso et al., 2014). Reynolds et al. (2002) proposed a functional classification of freshwater phytoplankton using morphological, physiological, and ecological traits. The phytoplankton functional groups approach currently identifies 40 assemblages by alpha-numeric codes according to their sensitivities and tolerances to environmental variability (Padisák et al., 2009). This approach allows us to classify the species into groups directly related to ecosystem function and supplies important knowledge about the dynamics of the species and habitat selection; it is currently being widely employed around the world ( Kruk et al., 2002; Huszar et al., 2003; Becker et al., 2010; Crossetti et al., 2013).
Our study aims to evaluate the effects of droughts on the dynamics of the phytoplankton biomass and functional groups composition. We took advantage of two exceptionally dry years in the semi-arid region of Northeastern Brazil to investigate the effects of a water level reduction on the water quality and phytoplankton communities of two shallow man-made lakes. We aimed to investigate whether water level reduction cause increase or decrease of the phytoplankton biomass and cyanobacteria dominance depending on the lake depth and inorganic turbidity.
Materials and methods
Study site
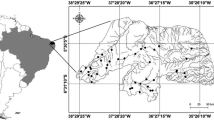
The man-made Lakes Pocinhos (6°34′42′′S; 37°19′47″W) and ESEC (6°34′49″S; 37°15´20″W) are located in Serra Negra do Norte, RN, Brazil and inserted in the same watershed of Espinharas River (Fig. 1). Lake Pocinhos is located in a private farm called Solidão, while Lake ESEC is partly located inside this farm and partly located at the Ecological Station of Seridó, a conservation unit of the Caatinga Biome. These lakes, as all lakes in the region, were constructed to provide a water supply. Both lakes are small and shallow. At the beginning of this study, Lake Pocinhos had an area of ~17 ha and a maximum depth of 6.4 m, while Lake ESEC had an area of ~12 ha and maximum depth of 4.5 m. The lakes have a poorly developed littoral zone and a homogeneous thermal and chemical regime. Lake Pocinhos is characterized by a litholic neossolo, comprised organic matter with a loam texture, and Lake ESEC is characterized by a chromic luvisol soil, a mineral soil with the presence of clay (EMBRAPA, 1971).

Map of Lake ESEC and Lake Pocinhos, showing the ecological station in darker gray
The regional climate is tropical and semi-arid (BS’h’, Kottek et al., 2006), characterized by irregular rainfall, high evapotranspiration rates, and negative water balance during most of the year. The mean annual precipitation is 733 mm and the rainy season is concentrated in 5 months, generally from January to May (data from 1992 to 2014 provided by EMPARN, the Local Agriculture Company).
Fish removal
In Lake ESEC, the shallower one, the water level and the oxygen concentrations were drastically reduced after the 2012 drought. Therefore, fish removal was employed as a management action to avoid massive fish mortality and further degradation of water quality. A total of 5.80 tons of fish was removed from this lake in March 2013. The most abundant species removed were the benthivorous Prochilodus brevis (Steindachner) and the filter-feeding Oreochromis niloticus (Linnaeus), which accounted for 87 and 8% of the total fish biomass removed, respectively.
Sampling and analysis
Water samples were taken at monthly intervals from May 2012 to May 2014, at eight randomly sampled sites of each lake. Due the lakes being small and spatially homogeneous, samples were integrated in one composite sample for nutrient, chlorophyll-a, and phytoplankton community analysis. Water temperature and dissolved oxygen were measured at each 0.5 meter from the surface to the bottom, using an oxygen probe (Instrutherm MO-900). Electric conductivity and turbidity were measured by specific portable equipment (AP2000, HPH 1002, respectively) at each sampling point. Phytoplankton samples were fixed with acetic solution of lugol.
Total nitrogen (TN) and dissolved inorganic nitrogen (DIN) were analyzed by the combustion of unfiltered and filtered water samples, respectively, using a carbon analyzer TOC-V Shimadzu with a TN analyzer attached. Total phosphorus (TP) was analyzed by the ascorbic acid method after oxidation with potassium persulfate (Valderrama, 1981). Water samples were filtered through glass fiber filters (VWR), and the filters were used for the analysis of chlorophyll-a and suspended solids. Chlorophyll-a was extracted with ethanol 95% and measured by spectrophotometry (Jespersen & Christoffersen, 1987). Total and inorganic suspended solids were determined by gravimetry after drying the filters overnight at 100 °C and the ignition of filters at 500 °C for 3 h (APHA, 2005). The organic suspended solids were measured by the difference between total suspended solids and inorganic suspended solids (APHA, 2005). The inorganic suspended solids (ISS) were used as a proxy of inorganic turbidity and the organic suspended solids (OSS) as a proxy of organic turbidity.
Phytoplankton composition was characterized at species level whenever possible and counted according to Utermöhl (1958) using an inverted microscope at ×400 magnification. Settling units (cells, colonies, and filaments) were enumerated in random fields (Uehlinger, 1964) and at least 100 specimens of the most frequent species were counted (Lund et al., 1958). Algal biovolume (mm3 l−1) was calculated using formulae for geometric shapes (Hillebrand et al., 1999), and 20 and 30 specimens or colonies on the average of each taxon were measured and expressed the fresh-weight unit in mass, where 1 mm3 l−1 = 1 mg l−1 (Wetzel & Likens, 2000). Descriptor species were defined as those whose contribution was >5% of total algal biomass and then classified into functional groups using the criteria of Reynolds et al. (2002) and Padisák et al. (2009). Species diversity was estimated using the Shannon and Wiener index (Shannon & Wiener, 1963) based on biomass and expressed by bits per milligrams. The Shannon and Wiener index and species richness were calculated using the PC-ORD® v.6 program.
The drought was classified based on the standardized precipitation index (SPI) (McKee et al., 1993) using data from 1962 to 2014 provided by INMET (National Institute of Meteorology) at the time scale of 12 months (SPI12). The index is based on the cumulative probability of a given rainfall event occurring at a station and determines the degree of a drought at a given time scale. Temperature and dissolved oxygen depth-lines were plotted using data triangulation with linear interpolation method. The euphotic depth (zeu) was calculated as 2.7 times the Secchi depth (Cole, 1994), and the ratio euphotic depth/maximum depth (zeu/zmax) was used as a proxy of light availability in the water column (Jensen et al., 1994).
Statistical analysis
In order to explain the sources of turbidity in the studied lakes, a multiple linear regression was used to relate the inverse of the Secchi depth (1/Sd), used here as a measure of turbidity, to the concentrations of chlorophyll-a, detritus, and inorganic suspended solids (Scheffer, 1998). To estimate the concentration of detritus, we assumed that the fraction of organic suspended solids comprised by algal biomass was 70 times de concentration of chlorophyll-a and the remained fraction was assumed to be detritus (Scheffer, 1998). A non-parametric correlation (Spearman) was used with the intent to observe differences between S N group and the environmental variables in both lakes. Multiple linear regression and correlational analyses were performed in Statistica 7 software.
A detrended correspondence analysis (DCA) with biological data was used to determine the best constrained ordination method for our data (Ter Braak & Prentice, 1988). Redundancy analysis (RDA) was then used to examine the relationships between the environmental variables of the man-made lakes and phytoplankton functional groups and to select the variables that best described the distribution of groups. The significance of the ordination axes was tested with a Monte Carlo test using 999 permutations (Mc Cune & Mefford, 2011). For the ordination analysis, data were transformed to log10 (x + 1). The environmental variables included in the RDA were euphotic and maximum depth, inorganic and organic suspended solids, total phosphorous, total nitrogen, dissolved inorganic nitrogen, and phytoplankton biomass. The program used for DCA and RDA was PC-ORD® v.6. The variance inflation factor (VIF) was used for detecting the presence of multicollinearity among the predictor variables in the RDA; VIF > 10 indicates multicollinearity (Kutner et al., 2004). Multicollinearity analysis was performed in R software (R Development Core Team).
Results
Physical and chemical conditions
The Brazilian semi-arid region was marked by a severe to extreme drought from April 2012 to February 2014, according to the standardized precipitation index (Fig. 2). During the study period, the water level dropped by half in both systems, ranging from 6 to 3 meters in Lake Pocinhos (Fig. 3a), and from 4 to 2 m in Lake ESEC (Fig. 3b), reducing both the euphotic depth and the zeu/zmax ratio. With the drop in water level, the euphotic depth decreased from 40% to less than 20% the maximum depth in Lake Pocinhos (Fig. 3a). On the other hand, the euphotic depth of Lake ESEC first decreased from 30 to 10%, and then increased from 10 to 70%, following the fish removal in March 2013, and finally decreased from 70% back to 30% the maximum depth (Fig. 3b).

Monthly precipitation (dark bars) and standardized precipitation index (SPI) (dark dashed line) in the study area from January 2012 to May 2014. The months with the SPI below the gray dashed line are considered severe to extreme droughts (May/12–Feb/14) and above is a normal drought to humid. The gray box indicates the studied period and the arrow indicates fish removal

Maximum depth, Zmax (dark bars), euphotic depth, Zeu (gray bars), and the ratio Zeu/Zmax (dashed line). The arrow indicates fish removal
Isothermal conditions for Pocinhos and ESEC were observed during most of the study period (Fig. 4a, b) with micro-stratifications in ESEC during Nov/12, Apr/13, Dec/13, and May/14 (Fig. 4b). However, oxygen stratification occurred during almost the whole period in Lake Pocinhos (Fig. 4c). Lake ESEC showed a homogeneous condition for oxygen, except for the hypoxic conditions observed from March to May 2013 (Fig. 4d), when the 5.8 tons of fish was removed.

Depth-time isopleths for water temperature (°C) and for dissolved oxygen (mg/l) in Pocinhos (a, c) and ESEC (b, d). The white area is the bottom of the lake
Conductivity increased during the study period, except for a quick reduction after the 150 mm precipitation in April 2013 (Fig. 5a). A similar pattern was observed for turbidity, but the drop in turbidity after the above mentioned precipitation event, was stronger in ESEC than in Lake Pocinhos (Fig. 5b). The source of turbidity was also different in the two lakes, being mostly organic in Lake Pocinhos (Fig. 5c) and inorganic in Lake ESEC (Fig. 5d). The multiple linear regression relating the inverse of Secchi depth to chlorophyll-a, detritus and inorganic suspended solids showed a high correlation, R 2 = 0.84 and R 2 = 0.60 for Pocinhos and ESEC Lakes, respectively. In Lake Pocinhos, the concentrations of chlorophyll-a and inorganic suspended solids (ISS) were significantly related to the inverse of Secchi depth, while in Lake ESEC the only significant source of turbidity was ISS (Table 1).

Monthly variations of conductivity (a), turbidity (b), percentage of total suspended solids comprised by organic (c) and inorganic (d) fractions, total phosphorous (e), and total nitrogen (f) in Lake Pocinhos (white squares) and Lake ESEC (dark squares). The values of turbidity (NTU) are in log scale. The dashed line indicates the period of fish removal
The concentration of TP did not show any trend with the water level reduction for Lake ESEC. However, for Lake Pocinhos a tendency to increase during the study period was observed (Fig. 5e). TN (Fig. 5f) and data not shown (DIN) exhibited an increasing trend with decreasing water level.
Phytoplankton dynamic
The phytoplankton biomass, measured either as the chlorophyll-a concentration (Fig. 6a) or estimated from the total biovolume (Fig. 6b) increased in Lake Pocinhos but decreased in Lake ESEC with a water level reduction. This reduction in the phytoplankton biomass and chlorophyll-a concentration in Lake ESEC occurred few months before the fish removal. The Shannon and Wiener index (H′) (Fig. 6c) and phytoplankton richness (Fig. 6d) decreased during the studied period in Lake Pocinhos and increased in Lake ESEC.

Chlorophyll-a (a) and total phytoplankton biomass (b), Shannon’s diversity index (c), and phytoplankton richness (d). The values of chlorophyll-a and total biomass are in log scale. The dashed line indicates the period of fish removal
In Lake Pocinhos, a total of 44 species were identified and distributed into 20 functional groups. Among these species only four species were descriptors of the lake (>5% of the total biomass), representing four functional groups by Reynolds (C, F, S N , S 1 ). On the other hand, Lake ESEC had a total of 58 species identified and distributed in 14 functional groups. Twenty descriptor species were members of 11 functional groups (S N , S 1 , Lo, X 1 , X 2 , C, F, J, Y, W 1 , W2) (Table 2).
The pattern of phytoplankton functional group replacement differed in the two lakes (Fig. 7). In Lake Pocinhos, phytoplankton composition shifted from a co-dominance of F, C, and S 1 groups (Fig. 7a), represented by the colonial green algae species Botryococcus braunii (Kützing), the small diatom Cyclotella striata (Kützing), and the cyanobacterium Planktolyngbya limnetica (Lemmermann), respectively, to a dominance of the S N group, represented by the N-fixing cyanobacterium Cylindrospermopsis raciborskii (Woloszynska).

Relative biomass of the phytoplankton functional groups in Pocinhos (a) and ESEC (b)
In Lake ESEC, we observed a shift in the phytoplankton composition from a dominance of the S N group, represented by the N-fixing cyanobacteria C. raciborskii, to a co-dominance of W 1 , W 2 , and Y, represented by the mixotrophic species Euglena spp., Trachelomonas spp., and Cryptomonas spp. Just after the fish removal, there was a recruitment of green algae groups, F and J, which were mostly represented by the colonial species B. braunii, Crucigenia quadrata (Morren), and Oocystis lacustris (Chodat) (Fig. 7b).
The correlation analysis for the S N group with environmental variables showed a negative correlation with maximum depth (r = −0.92) and euphotic zone (r = −0.91) and a positive correlation with OSS (r = 0.94) and DIN (r = 0.90) in Lake Pocinhos. For Lake ESEC, the S N group was positively related to maximum depth (r = 0.75) and euphotic zone (r = 0.49) and negatively correlated to ISS (r = −0.96) and DIN (r = −0.56).
Redundancy analysis
The variance inflation factor (VIF) was less than 10 for all variables included in the RDA (Zmax = 3.78, Zeu = 2.93, ISS = 3.36, OSS = 8.82, TP = 1.56, DIN = 3.71, TN = 1.57, Biomass = 7.40).
The first axis of RDA accounted for 41%, while the second axis accounted for just 6.1% of the species-environmental variables’ relations. The Monte Carlo test indicated that the eigenvalue of the first axis was significant (P < 0.001). This axis was positively correlated to ISS (0.40) and negatively correlated to biomass (−0.80), zeu (−0.67), zmax (−0.60), OSS (−0.40), DIN (−0.31), and TN (−0.24). The second axis was mainly positively correlated to zeu (0.67) and negatively correlated to OSS (−0.38) and TP (−0.28). Variables from Lake ESEC were positively correlated to the first axis and were mostly related to W 1, W 2 , Y, and X 2 functional groups. On the other hand, variables from Lake Pocinhos were negatively correlated with the first axis and were mostly related to S N and S 1 groups (Fig. 8).

RDA ordination of the phytoplankton functional groups and environmental variables for ESEC (dark squares) and Pocinhos (white squares). ISS inorganic turbidity, OSS organic turbidity, Zeu euphotic zone, Zmax maximum depth, TP total phosphorous, DIN dissolved inorganic nitrogen, TN total nitrogen
Discussion
The study describes contrasting dynamics of phytoplankton functional groups in two rather similar lakes during an extreme drought period. Our results show that a water level reduction may either increase or decrease the phytoplankton biomass and favor cyanobacteria or mixotrophic flagellates’ dominance in semi-arid shallow lakes depending on the water depth. In Lake Pocinhos, the negative water balance reduced the lake depth and increased the nutrient concentration and phytoplankton biomass, leading to the dominance of cyanobacteria bloom forming groups as expected (Naselli-Flores et al., 2007; Jeppesen et al., 2015). However, in the shallower Lake ESEC, the water level reduction promoted sediment resuspension and increased inorganic turbidity, decreasing light availability and the phytoplankton biomass, shifting the dominance from cyanobacteria bloom forming groups (S N ) to mixotrophic flagellate functional groups (W 1 , W 2 , Y).
The role of water level fluctuations in shallow lakes is not completely known, since they do not respond linearly to such disturbances (Coops et al., 2003). However, water level reductions have strong implications for water quality and phytoplankton community structure (Nõges & Nõges, 1999; Bouvy et al., 2003; Naselli-Flores & Barone, 2005, Medeiros et al., 2015). Water level reductions can affect the availability of light and nutrients for phytoplankton. Nutrients became more available in both lakes as they were concentrated in a smaller water volume, but in Lake Pocinhos, light became less available due to phytoplankton self-shading and due to inorganic matter resuspension in Lake ESEC. Due to its shallower depth, sediment resuspension by wind and fish facilitated turbidity more in ESEC than in Pocinhos.
Underwater light availability is a key factor regulating phytoplankton dynamics (Zohary et al., 2010). The privation of light could be lethal to a single-celled autotroph (Reynolds, 1990). The euphotic depth to maximum depth ratio was low in both lakes, except for a brief increase in Lake ESEC just after the fish removal. This may indicate that light was limiting phytoplankton growth in most of the water column of both lakes during the study period despite the high nutrient concentrations. C. raciborskii, included in the S N group is characteristic of warm, mixed, phosphorous-enriched waters, and they are tolerant to low light conditions (Reynolds et al., 2002). C. raciborskii traits, such as shape and pigment composition, are shown to enhance their ability to increase their light-harvesting capacity due a high phenotypic plasticity in response to key environmental factors (Bonilla et al., 2012). However, studies showing the importance of light on phytoplankton growth and dominance argue that representatives of group S N may not be competitively favored under lower light intensity, as in the case of Lake ESEC (Kokociński et al., 2010, Bonilla et al., 2012). Therefore, other functional groups more tolerant to this light-limiting condition should be favored. The reduced underwater light availability and the relatively high dissolved organic carbon (DOC) in the system should be an advantage for mixotrophic algae over autotrophic ones (Jones, 2000).
During the period with more light availability and less nutrients in Lake Pocinhos, group F had a considerable proportion of total phytoplankton biomass. This group represented by large colonies of green algae requires high light availability for better cell functioning and is tolerant to low nutrients (Reynolds et al., 2002). Water bodies less eutrophic, generally have large colonies and large rounded unicellular organisms of green algae (Naselli-Flores & Barone, 2000); and the presence of groups C and S 1 during this period can be explained by their geometric forms; the small cylinder shape of Cyclotella striata (GALD < 10 μm) and the elongated and filamentous shape of Planktolyngbya limnetica can reduce significantly their sinking rate (Reynolds, 2006). However, the effect of droughts provided an ideal habitat for the dominance of the N-fixing cyanobacteria bloom forming group S N , in Lake Pocinhos. Light and temperature seem to be the most important factors related to the appearance and development of C. raciborskii, the representative species of group S N .
In contrast, Lake ESEC had an opposite pattern of total biomass and groups’ replacement. Total phytoplankton biomass declined before the fish removal, indicating that the phytoplankton biomass change was independent of the fish biomass reduction. Phytoplankton functional groups shifted from a dominance of S N to a co-dominance of mixotrophic flagellate groups W 1, W 2 , and Y. This can be explained by the water level reduction that affected the volume of the shallower man-made lake, magnifying the sediment–water interaction. Sediment resuspension by wind and benthivorous fish increased the inorganic suspended solids (ISS) more in ESEC than in Pocinhos and altered the light availability for phytoplankton functional groups. Mixotrophic species can compensate light or nutrient deficiency altering the nutrition pathway from autotrophy to heterotrophy (Katechakis et al., 2005). The flagellate groups have been found in the water layer near the bottom in different shallow lakes (Ibanez, 1998; Reynolds et al., 2002). When the water level decreased, it affected the volume and sediment–water interaction, explaining the appearance of groups W 1 and W 2 . Also, the potentially high organic matter is an important factor for the bottom-dweller groups once they adapt to mixotrophic conditions (Reynolds et al., 2002). Some representatives of the functional group Y, such as small cryptomonas, are capable of persisting under extremely poor light conditions (Sinistro et al., 2006). Medeiros et al. (2015) also found that cryptomonas favored in low waters with high inorganic turbidity during a drought period in a semi-arid region. Our results suggest that high inorganic turbidity and critical light availability are the main factors affecting the replacement of C. raciborskii (S N ) to the cryptomonas (Y), which are more competitive in these conditions.
Benthivorous fish can resuspend sediments and translocate nutrients and detritus from the bottom to the water column (Schindler et al., 1996; Vanni, 1996), affecting the biogeochemical process and trophic interactions in freshwater ecosystems (Persson & Svensson, 2006). The removal of 5.8 tons of benthivorous fish from Lake ESEC reduced the water turbidity and sediment bioturbation by the fish, increasing light availability and favoring the recruitment of green algae groups (F and J) that demand additional light and are bioindicators of improved water quality. However, groups F and J did not persist in the system, probably because of the prolonged effect of the drought or maybe because the effect of fish removal was not strong enough. Whatever the reason for the transient dominance of F and J, W 1, W 2 , Y, and Lo were the groups with greater importance in the biomass in Lake ESEC during the end of the studied period. Although the fish removal showed a tendency to improve the water quality, its effects could not persist with the extension of the extreme drought period. Experimental studies are needed to investigate the combined and isolated effects of fish removal and water level reductions on the structure and dynamics of phytoplankton communities of shallow lakes. The correlation analysis confirmed that the recruitment of the S N group in Lake Pocinhos was strongly related to the reduced water level and water transparency (Zmax and Zeu), changing phytoplankton resources and increasing the biomass and organic matter. On the other hand, in Lake ESEC, the reduced water level and water transparency leads to the collapse of the S N group, strongly related to inorganic suspended solids in the system.
The increase in phytoplankton diversity and richness in Lake ESEC was attributed to a shift in dominance from the S N (C. raciborskii) to the Y, W 1 , and W 2 groups, due water depth and turbidity change caused by drought, enhancing the number of co-dominant species and functional groups. Different behaviors were observed in Lake Pocinhos, attributed to the shift from the co-dominance of the C, S1, and F groups to the dominance of S N , decreasing the phytoplankton richness and diversity.
The response of phytoplankton organisms to environmental alterations depends on the time scale of their frequency and duration. The time scale relevant for the phytoplankton replication time varies between hours to a week (Reynolds, 1990). Several months in plankton succession correspond to decades in grassland or centuries in forest succession (Padisák, 1992). Therefore, our study comprises several generations of phytoplankton, and 2 years is an appropriate time scale to investigate the effects of a drought on phytoplankton communities. It is well documented that the association of physical, chemical, and biological factors is recognized in determining the structure of a phytoplankton community (Reynolds, 1989, Soares et al., 2009). However, studies show that physical factors, such as light and climate, may contribute to a crucial understanding of phytoplankton dynamics and species composition (Zohary et al., 2010).
Our study shows the effects of an extreme drought on the water quality and phytoplankton dynamic of two tropical shallow lakes. Contradicting the expectations that in future warmer climates, a reduction in water level would lead to cyanobacterial blooms (Marengo et al., 2010; Moss et al., 2011; Roland et al., 2012), our results shows that water level reductions induced by droughts may either increase or decrease the phytoplankton biomass and cyanobacterial blooms and demonstrate the importance of mixotrophic flagellate algae in low light conditions. The pressures on water resources of dry regions are strong, and the need for more ecological studies in these systems is therefore urgent (de Senerpont Domis et al., 2013; Sarmento et al., 2013).
References
Allende, L., G. Tell, H. Zagarese, A. Torremorell, G. Pérez, J. Bustingorry, R. Escaray & I. Izaguirre, 2008. Phytoplankton and primary production in clear-vegetated, inorganic-turbid, and algal-turbid shallow lakes from the pampa plain (Argentina). Hydrobiologia 624: 45–60.
APHA, 2005. Standard Methods for the Examination of Water and Wastewater, 20th ed. American Public Health Association, Washington DC.
Bates, B.C., Z.W. Kundzewicz, S. Wu & J.P. Palutikof, 2008. Climate Change and Water. Technical Paper of the Intergovernmental Panel on Climate Change, IPCC Secretariat, Geneva
Becker, V., L. Caputo, J. Ordóñez, J. Marcé, J. Armengol, L. O. Crossetti & V. L. M. Huszar, 2010. Driving factors of the phytoplankton functional groups in a deep Mediterranean reservoir. Water Research 44: 3345–3354.
Bonilla, S. L., M. C. S. Aubriot, M. Soares, A. González-Piana, V. L. M. Fabre, M. Huszar, D. Lurling, J. Padisák Antoniades & C. Kruk, 2012. What drives the distribution of the bloom-forming cyanobacteria Planktothrix agardhii and Cylindrospermopsis raciborskii? Microbiology Ecology 79: 594–607.
Bouvy, M., R. Molica, S. de Oliveira, M. Marinho & B. Beker, 1999. Dynamics of a toxic cyanobacterial bloom (Cylindrospermopsis raciborskii) in a shallow reservoir in the semi-arid region of northeast Brazil. Aquatic Microbial Ecology 20: 285–297.
Bouvy, M., S. M. Nascimento, R. J. R. Molica, A. Ferreira, V. Huszar & S. M. F. O. Azevedo, 2003. Limnological features in Tapacurá reservoir (northeast Brazil) during a severe drought. Hydrobiologia 493: 115–130.
Cole, G. A., 1994. Textbook of Limnology. Waveland Press, Illinois: 412.
Coops, H., M. Beklioglu & T. L. Crisman, 2003. The role of water-level fluctuations in shallow lake ecosystems – workshop conclusions. Hydrobiologia 506–509: 23–27.
Crossetti, L. O., V. Becker, L. S. Cardoso, L. R. Rodrigues, L. S. Costa & D. Motta-Marques, 2013. Is phytoplankton functional classification a suitable tool to investigate spatial heterogeneity in a subtropical shallow lake? Limnologica - Ecology and Management of Inland Waters 43: 157–163.
Dantas, Ê. W., M. D. C. Bittencourt-Oliveira & A. D. N. Moura, 2012. Dynamics of phytoplankton associations in three reservoirs in northeastern Brazil assessed using Reynolds’ theory. Limnologica- Ecology and Management of Inland Waters 42: 72–80.
De Senerpont Domis, L. N., J. J. Elser, A. S. Gsell, V. L. M. Huszar, B. W. Ibelings, E. Jeppesen, S. Kosten, W. M. Mooij, F. Roland, U. Sommer, E. V. Donk, M. Winder & M. Lürling, 2013. Plankton dynamics under different climatic conditions in space and time. Freshwater Biology 58: 463–482.
Empresa Brasileira de Pesquisa em Agropecuária, 1971. Levantamento exploratório – Reconhecimento de solos do Estado do Rio Grande do Norte. Available http://www.uep.cnps.embrapa.br/solos/index.php?link=rn.
Hillebrand, H., C. D. Dürselen, D. Kirschtel, U. Pollingher & T. Zohary, 1999. Biovolume calculation for pelagic and benthic microalgae. Journal of Phycology 35: 403–424.
Huszar, V., C. Kruk & N. Caraco, 2003. Steady-state assemblages of phytoplankton in four temperate lakes (NE USA). Hydrobiologia 502: 97–109.
Ibanez, M. D. S. R., 1998. Phytoplankton composition and abundance of a central Amazonian floodplain lake. Hydrobiologia 362: 79–83.
IPCC Climate Change, 2007. Impacts, Adaptation an Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernamental Painel on Climate Change. Cambridge University Press, Cambridge, UK.
Jensen, P., E. Jeppesen, K. Olrik & P. Kristensen, 1994. Impact of nutrients and physical factors on the shift from cyanobacterial to chlorophyte dominance in shallow Danish lakes. Canadian Journal of Fisheries and Aquatic Sciences 51: 1692–1699.
Jeppesen, E., S. Brucet, L. Naselli-Flores, E. Papastergiadou, K. Stefanidis, T. Nõges, P. Nõges, J. L. Attayde, T. Zohary, J. Coppens, T. Bucak, R. F. Menezes, F. R. S. Freitas, M. Kernan, M. Søndergaard & M. Beklioglu, 2015. Ecological impacts of global warming and water abstraction on lakes and reservoirs due to changes in water level and related changes in salinity. Hydrobiologia 750: 201–227.
Jespersen, A. M. & K. Christoffersen, 1987. Measurements of chlorophyll-a from phytoplankton using ethanol as extraction solvent. Hydrobiologia 109: 445–454.
Jones, R. I., 2000. Mixotrophy in planktonic protists: an overview. Freshwater Biology 45: 219–226.
Katechakis, A., T. Haseneder, R. Kling & H. Stibor, 2005. Mixotrophic versus photoautotrophic specialist algae as food for zooplankton: The light: nutrient hypothesis might not hold for mixotrophs. Limnology and Oceanography 50: 1290–1299.
Kottek, M., J. Grieser, C. Beck, B. Rudolf & F. Rubel, 2006. World map of the Koppen-Geiger climate classification updated. Meteorologische Zeitschrift 15: 259–263.
Kruk, C., N. Mazzeo, G. Lacerot & C. S. Reynolds, 2002. Classification schemes for phytoplankton: a local validation of a functional approach to the analysis of species temporal replacement. Journal of Plankton Research 24: 901–912.
Kokociński, M., K. Stefaniak, J. Mankiewicz-Boczek, K. Izydorczyk & J. Soininen, 2010. The ecology of the invasive cyanobacterium Cylindrospermopsis raciborskii (Nostocales, Cyanophyta) in two hypereutrophic lakes dominated by Planktothrix agardhii (Oscillatoriales, Cyanophyta). European Journal of Phycology 45: 365–374.
Kutner, M. H.; C. J. Nachtsheim & J. Neter, 2004. Applied Linear Regression Models (4th ed.). McGraw-Hill Irwin.
Lake, P. S., 2003. Ecological effects of perturbation by drought in flowing waters. Freshwater Biology 48: 1161–1172.
Lund, J. W. G., C. Kipling & E. D. Le Cren, 1958. The inverted microscope method of estimating algal num-ber and the statistical basis of estimating by counting. Hydrobiologia 11: 143–170.
Marengo, J. A., T. Ambrizzi, R. P. da Rocha, L. M. Alves, S. V. Cuadra, M. C. Valverde, R. R. Torres, D. C. Santos & S. E. T. Ferraz, 2010. Future change of climate in South America in the late twenty-first century: intercomparison of scenarios from three regional climate models. Climate Dynamics 35: 1073–1097.
Mc Cune, B. & M.J. Mefford, 2011. PC-ORD. Multivariate analysis of ecological data. version 6.0. MjM Software Design, Oregon.
McKee, T.B., N.J. Doesken & J. Kleist, 1993. The relationship of drought frequency and duration to time scales. In Proceedings of the 8th Conference on Applied Climatology. American Meteorological Society, Boston, MA: 179–183.
Medeiros, L. C., A. Mattos, M. Lurling & V. Becker, 2015. Is the future blue-green or brown? The effects of extreme events on phytoplankton dynamics in a semi-arid man-made lake. Aquatic Ecology 49: 293–307.
Mishra, A. K. & V. P. Singh, 2010. A review of drought concepts. Journal of Hydrology 391: 202–216.
Moss, B., S. Kosten, M. Meerhoff, R. W. Battarbee, E. Jeppesen, N. Mazzeo, K. Havens, G. Lacerot, Z. Liu, L. de Meester, H. Paerl & M. Sheffer, 2011. Allied attack: climate change and eutrophication. Inland Waters 1: 101–105.
Naselli-Flores, L. & R. Barone, 2000. Phytoplankton dynamics and structure: a comparative analysis in natural and man-made water bodies of different trophic state. Hydrobiologia 438: 65–74.
Naselli-Flores, L. & R. Barone, 2005. Water-level fluctuations in mediterranean reservoirs: setting a dewatering threshold as a management tool to improve water quality. Hydrobiologia 548: 85–99.
Naselli-Flores, L., R. Barone, I. Chorus & R. Kurmayer, 2007. Toxic cyanobacterial blooms in reservoirs under a semiarid Mediterranean climate: the magnification of a problem. Environmental Toxicology 22: 399–404.
Nõges, T. & P. Nõges, 1999. The effect of extreme water level decrease on hydrochemistry and phytoplankton in a shallow eutrophic lake. Hydrobiologia 143: 277–283.
Padisák, J., 1992. Spatial and temporal scales in phytoplankton ecology. Abstracta Botanica 16: 15–23.
Padisák, J., L. O. Crossetti & L. Naselli-Flores, 2009. Use and misuse in the application of the phytoplankton functional classification: a critical review with updates. Hydrobiologia 621: 1–19.
Paerl, H. W. & J. Huisman, 2009. Climate change: a catalyst for global expansion of harmful cyanobacterial blooms. Environmental Microbiology Reports 1: 27–37.
Paerl, H. W. & T. G. Otten, 2013. Harmful cyanobacterial blooms: causes, consequences, and controls. Microbial Ecology 65: 995–1010.
Persson, A. & J. S. M. Svensson, 2006. Effects of benthivorous fish on biogeochemical processes in lake sediments. Freshwater Biology 51: 1298–1309.
Reynolds, C. S., 1989. Physical determinants of phytoplankton succession. In Sommer, U. (ed.), Plankton Ecology: Succession in Plankton Communities. Springer Verlag, Berlin: 9–56.
Reynolds, C. S., 1990. Temporal scales of variability in pelagic environments and the response of phytoplankton. Freshwater Biology 23: 25–53.
Reynolds, C. S., 1998. What factors influence the species composition of phytoplankton in lakes of different trophic status? Hydrobiologia 369(370): 11–26.
Reynolds, C. S., 2006. The Ecology of Phytoplankton (Ecology, Biodiversity and Conservation). Cambridge University Press, Cambridge.
Reynolds, C. S., V. L. M. Huszar, C. Kruk, L. Nasseli-Flores & S. Melo, 2002. Towards a functional classification of the freshwater phytoplankton. Journal of Plankton Research 24: 417–428.
Roland, F., V. L. M. Huszar, V. F. Farjalla, A. Enrich-Prast, A. M. Amado & J. P. H. B. Ometto, 2012. Climate change in Brazil: perspective on the biogeochemistry of inland waters. Brazilian Journal of Biology 72: 709–722.
Ross, T. & N. Lott, 2003. A climatology of 1980–2003 extreme weather and climate events. US. National Environmental Satellite Data and Information Service. National Climatic Data Center, Asheville, North Carolina
Salmaso, N., L. Naselli-Flores & J. Padisák, 2014. Functional classifications and their application in phytoplankton ecology. Freshwater Biology 60: 603–619.
Sarmento, H., A. M. Amado & J. P. Descy, 2013. Climate change in tropical fresh waters (comment on the paper ‘Plankton dynamics under different climatic conditions in space and time’by de Senerpont Domis et al., 2013). Freshwater Biology 58: 2208–2210.
Scheffer, M., 1998. Ecology of Shallow Lakes. Chapman & Hall, London.
Schindler, D. E., S. R. Carpenter, K. L. Cottingham, X. He, J. R. Hodgson, J. F. Kitchell & P. A. Soranno, 1996. Foodweb structure and littoral zone coupling to pelagic trophic cascades. In Polis, G. A. & K. O. Winemiller (eds), Food Webs: Integration of Pattern and Dynamics. Chapman & Hall, New York: 96–105.
Shannon, C. E. & W. Weaver, 1963. The mathematical theory of communication. University Press, Urbana, Illinois.
Sinistro, R., I. Izaguirre & V. Asikian, 2006. Experimental study on the microbial plankton community in a South American wetland (Lower Parana River Basin) and the effect of the light deficiency due to the floating macrophytes. Journal of Plankton Research 28: 753–768.
Soares, M. C. S., M. I. A. Rocha, M. M. Marinho, S. M. F. O. Azevedo, C. W. C. Branco & V. L. M. Huszar, 2009. Changes in species composition during annual cyanobacterial dominance in a tropical reservoir: physical factors, nutrients and grazing effects. Aquatic Microbial Ecology 57: 137–149.
Ter Braak, C. J. F. & I. C. Prentice, 1988. A theory of gradient analysis. Advances in Ecological Research 18: 271–317.
Uehlinger, V., 1964. Étude statistique des méthodes de dénobrement planctonique. Archive Science 17: 121–123.
Utermöhl, H., 1958. Zur vervollkommung der quantitativen phytoplankton methodik. Mitteilungen der international. Vereinigung für Theoretische und Angewandte Limnologie 9: 1–38.
Valderrama, J. C., 1981. The simultaneous analysis of total nitrogen and phosphorus in natural waters. Marine Chemistry 10: 109–122.
Vanni, M. J., 1996. Nutrient transport and recycling by consumers in lake food webs: implications for algal communities. In Polis, G. A. & K. O. Winemiller (eds), Food Webs: Integration of Pattern and Dynamics. Chapman & Hall, NY: 81–95.
Wetzel, R. G. & G. E. Likens, 2000. Limnological Analyses. WB Saunders Co., Philadelphia.
Zohary, T., J. Padisák & L. Naselli-Flores, 2010. Phytoplankton in the physical environment: beyond nutrients, at the end, there is some light. Hydrobiologia 639: 261–269.
Acknowledgments
This study was financially supported by CNPq (Brazilian National Research Council) and ICMBio (Institute Chico Mendes of Biodiversity Conservation) process No. 372170/2014-5. Thanks go to CAPES (Coordination of Improvement of Higher Education Personnel) for granting a master’s scholarship. We wish to thank the employees of the Ecological Station of Seridó (ESEC-RN) for their structure and field support; Leonardo Rosa for creating the localization map; Pablo Rubim, Gabriela Moura, Jurandir Mendonça-Júnior, Fabiana Araújo, Maria Marcolina Cardoso, Leonardo Teixeira, Danyhelton Dantas, Edson Santana, and Viviane Medeiros for their fieldwork, sampling, and laboratory analysis and Rosemberg Menezes for helping with the multicollinearity statistics analysis.
Author information
Authors and Affiliations
Corresponding author
Additional information
Guest editors: M. Beklioğlu, M. Meerhoff, T. A. Davidson, K. A. Ger, K. E. Havens & B. Moss / Shallow Lakes in a Fast Changing World
Rights and permissions
About this article

Cite this article
da Costa, M.R.A., Attayde, J.L. & Becker, V. Effects of water level reduction on the dynamics of phytoplankton functional groups in tropical semi-arid shallow lakes. Hydrobiologia 778, 75–89 (2016). https://doi.org/10.1007/s10750-015-2593-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-015-2593-6