
Abstract
The present study evaluates the effect of climatic events La Niña and drought on the structure of phytoplankton and hydrological variables in the Amazon estuary. Samples were collected at three sampling stations in the Taperaçu Estuary, on the Brazilian Amazon coast, in 2010 and 2011. During the rainy season of the La Niña (RS-LN) event, salinity reached significantly lower (p < 0.05) than normal values in the upper sector (10.5 ± 5.9), and significant spatial variation (p < 0.05) was recorded in the concentrations of silicate, chlorophyll-a, and phytoflagellates abundance. Significant differences were also found in salinity, turbidity, and the concentrations of dissolved nutrients and chlorophyll-a, as well as the abundance of specific phytoplankton groups, between the dry seasons of the post-drought (DS-PD) and La Niña (DS-LN) periods. During the post-drought dry season (DS-PD), high salinity (37.1 ± 1.3) and reduced precipitation contributed to the development of Psammodictyon panduriforme, a dominant marine species recorded during this period. During periods of higher rainfall (RS-LN and, to a lesser extent, DS-LN), salinity reached its lowest levels, which favored the growth of diatoms such as Cymatosira belgica, Plagiogrammopsis sp., and Skeletonema costatum. The present results indicate that the dynamics of the phytoplankton community of the Taperaçu Estuary is highly influenced by seasonality of rainfall, which is increased (more than 20%) by anomalous climate oscillations and directly affect environmental variables. This pattern would be expected in the dozens of other estuaries that have similar characteristics on the Amazon coast and in other tropical coastal zones around the world.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Estuaries are among the world’s most productive marine ecosystems, due to the availability of dissolved nutrient salts that originate primarily from inland drainage basins, together with an abundance of primary producers (Muylaert et al. 2009; Ricklefs 2010). These primary producers include phytoplankton—microscopic photosynthetic algae—which are responsible for most of the production of organic matter in estuarine environments (Fu et al. 2009; Cloern et al. 2014).
The phytoplankton community is ecologically significant as bio-indicators of the quality of the water bodies (Sathicq et al. 2017; Gao et al. 2018), given that these organisms respond rapidly to changes in environmental conditions, through shifts in their occurrence patterns and growth rates (López-Abbate et al. 2017; Barrera-Alba et al. 2019). The diatoms and dinoflagellates are typically the most dominant phytoplankton groups in estuarine environments (Matos et al. 2011; Wang et al. 2017), where their spatial and seasonal distribution patterns are influenced primarily by hydrodynamic forces, the availability of sunlight and dissolved nutrients, rainfall levels, temperature oscillations, and biological factors such as competition and herbivory, together with the intrinsic physiological characteristics of each species (Garmendia et al. 2013; Gao et al. 2018; Liu and Swart 2018).
On the Amazon coast, high rainfall levels (normally 2000–3000 mm, annually), macrotides (> 4 m), and the presence of extensive mangrove forests (Souza Filho et al. 2005; INMET 2018), all play an important role in the dynamics of the local estuarine environments, which present a set of a unique hydrological and hydrodynamic attributes, including high turbidity, nutrient-rich waters, high productivity, and strong tidal currents (Pamplona et al. 2013; Pereira et al. 2017).
But it is also important to understand how extreme climate events—droughts and major floods—affect the hydrological conditions and phytoplankton communities of the Amazon coast. During the period of the present study, two short-term events occurred. The first was in 2010, when the Amazon region was affected by the most extreme drought of the century, which caused profound social and environmental impacts throughout the Amazon region, with knock-on effects for the global climate (Marengo et al. 2011; 2013). The second event was in 2011, when a La Niña event, which began to develop in mid-2010, caused extensive flooding in northern South America, as well as Australia and Southeast Asia (Feng et al. 2013; Pereira et al. 2013; Thompson et al. 2015; Andrade et al. 2016).
Despite the considerable ecological and socioeconomic importance of the ecosystems of the Amazon coast (fisheries are one of the principal sources of income for the local populations), few studies are available on the composition of local phytoplankton communities, their spatial and temporal dynamics, or their relationships with the hydrological and climatological conditions found in the region’s estuaries. Given this, one other important aspect of the present study is that it is the first, to our knowledge, to evaluate the variation in the seasonal and spatial distribution of local phytoplankton communities in the context of the extreme fluctuations in dissolved inorganic nutrients observed during anomalous periods of rainfall (Andrade et al. 2016; Pereira et al. 2017; Costa et al. 2022).
The choice of a minor estuary (Taperaçu Estuary) as the area for the present study area was determined by the relative abundance of these features on the Amazon coast, and the fact that systems of smaller size may facilitate the understanding of the effects of natural events, such as atypical climatic conditions (Callaway et al. 2014; Pereira et al. 2017), on the oscillations in environmental variables, and their consequences for the local plankton communities. In addition, the Taperaçu Estuary is located within a Marine Extractive Reserve (‘Reserva Extrativista Marinha de Caeté-Taperacú’), a sustainable-use protected area, created by ordinance 17 on May 20, 2005, decree without number (Brasil 2005), which guarantees minimal human impact and makes the estuary an excellent study site for the understanding of climatic events, as reported by Costa et al. (2013a), Andrade et al. (2016) and Pereira et al. (2017). The present study aims to evaluate the effect of La Niña and drought climatic events on the structure and dynamics of the phytoplankton community, and on the environmental variables in an Amazonian estuary. These insights are of enormous relevance for the evaluation of similar systems at low latitudes on the Amazon coast and in other regions around the world.
Materials and methods
Study area
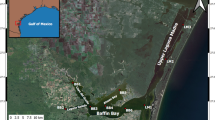
The Taperaçu Estuary is located in the municipality of Bragança, in northeastern Pará state, Brazil (46°42’–46°45’ W, 00°50’–00°57' S; Fig. 1). This estuary is part of the Bragança Coastal Plain, approximately 200 km east of the mouth of the Amazon River (Magalhães et al. 2009 and references therein).

Location of the study area in Bragança Coastal Plain, State of Pará, Amazon region, Brazil (A and B). Taperaçu Estuary (C) showing the sampling stations (upper—S1, middle—S2 and lower—S3)
The water surface of the Taperaçu has an area of 21 km2, and its catchment area is approximately 40 km2, which means that there is a negligible freshwater inflow from the adjacent area during the rainy season, although oligohaline water does flow in from the neighboring Caeté Estuary through the Taici tidal creek during the flood tide (Asp et al. 2012; Araújo & Asp, 2013; Magalhães et al. 2015; Oliveira et al. 2021). The Taperaçu is a funnel-shaped, tide-dominated estuary with relatively shallow water (mean depth of 4 m), but with deeper channels (up to 12 m in depth), located primarily at the margins of the estuary (Asp et al. 2012). In contrast with other estuaries on the Brazilian coast, the Taperaçu, together with other estuaries in northern Brazil (Asp et al. 2018), have semidiurnal tides with an amplitude of approximately 4 m during neap tides and up to 6 m on the spring tide (Asp et al. 2012; Pereira et al. 2017).
The Taperaçu can also be classified as a permanently open estuary, with highly turbid, but shallow waters (Magalhães et al. 2015). Approximately 82% of the water of the estuary flows in and out on each tide cycle, with large amounts of organic and inorganic material being imported through the resuspension of sediments from the adjacent mangrove (Asp et al. 2012).
The local climate is humid tropical, with a rainy season normally extending from December to May, when more than 85% of the annual precipitation occurs (Moraes et al. 2005). The driest months are September, October, and November, when rainfall may be negligible, leading to higher insolation and evaporation rates. Mean rainfall during the rainy season typically exceeds 2200 mm, although the data collected during the past few decades (Fig. 2) reveal major fluctuations resulting from droughts, and El Niño and La Niña events (Pereira et al. 2013, 2017; Andrade et al. 2016). Air temperatures are high and stable, ranging from 25.2 to 26.7 °C, with a mean annual temperature of approximately 25.5 °C. The wind also varies seasonally, with the strongest winds occurring during the dry season, and more moderate winds during the rainy season (INMET 2018).

Total annual precipitation (A) between 1974 and 2011, highlighting the principal deviations in rainfall levels (LN = La Niña event; EN = El Niño event; F = Flood; D = Drought); and Historical average (1974 and 2011) and Total monthly rainfall in the years of study (B) (source: INMET 2015)
Field procedures and laboratory analyses
Rainfall data were provided by the National Institute of Meteorology (INMET) and were obtained from the INMET meteorological station located in the town of Tracuateua, Pará (station 85145—47º10' W, 01º05' S), approximately 17 km from Bragança and 27 km from Taperaçu Estuary.
Samples were collected at three fixed stations (Fig. 1) distributed along the estuary, with station 1 (S1—00°56′58.4″ S, 46°46′6.9″ W) located in the upper sector, station 2 (S2—00°55′06.8″ S, 46°44′00.0″ W) in the middle sector, and station 3 (S3—00°50′30.9″ S, 46°43′2.4″ W) at the mouth of the estuary, representing the lower sector. Samples were collected on the flood tide in the rainy (February, April, and June 2011, during a La Niña event—RS-LN) and dry seasons (October 2010, following a drought event—DS-PD, and October 2011, during the La Niña event—DS-LN). The flood tide was chosen as the sampling period due to the inundation of the higher areas of mangrove, which favors the transport of organic and inorganic particles and nutrients to the center of the estuary, where they contribute to the development of the phytoplankton.
The subsurface water temperature, conductivity (salinity), turbidity, and the dissolved oxygen (DO) concentration were determined in situ using a bottom-mounted sonde to measure the electrical conductivity, temperature, pressure of seawater and dissolved oxygen concentration—CTDO (RBRmaestro) with a turbidity sensor, which was moored to the seafloor at a depth of 1.7 m. Standard 400-ml samples of the subsurface water (~ 1.0 m below the surface) were collected in Niskin oceanographic bottles for the laboratory analysis of the other environmental variables, that is, the pH and the concentrations of ammonium \({\text{NH}}_{4}^{+}\), phosphate (\({\text{PO}}_{4}^{3-}\)) and silicate (\({\text{SiO}}_{2}^{-}\)).
The Niskin bottles were also used to collected subsurface samples of the phytoplankton for qualitative and quantitative analyses, which were stored in 600-ml plastic containers and preserved immediately in Lugol’s solution (Throndsen 1978). Additional samples of the subsurface water were collected for the qualitative study, using conical plankton nets (64 μm mesh size), with 3 min horizontal subsurface hauls at a mean speed of 1.5 knots. The samples collected here were fixed in 4% formalin (final concentration) neutralized with sodium tetraborate (borax).
In the laboratory, the pH was measured with a pH meter. The analysis of dissolved nutrients was performed according to the methods described by Strickland and Parsons (1972) and Grasshoff et al. (1983), whereas chlorophyll-a concentrations were estimated spectrophotometrically, according to Parsons and Strickland (1963).
Quantitative analyses of the phytoplankton (number of cells per liter) were based on the Utermöhl method, which consists of the sedimentation of the samples in a sedimentation chamber for 12–24 h. These samples were stained previously with Rose Bengal and analyzed at 400× magnification under an inverted microscope (Carl Zeiss–Axiovert). The total area of the 7 ml cuvette was counted, considering a minimum number of 300 individuals of the most abundant species. Phytoflagellates (nanoplankton fraction \(\le\) 20 µm) were identified to the group level and counts were performed in 5 random fields. The phytoplankton species were identified based on the available taxonomic keys such as: Cupp (1943), Anagnostidis and Komárek (1988), Steindinger and Tangen (1997 apud Tomas 1997), Ross and Sims (1972), Ross et al. (1979), Round et al. (1990), Hasle and Syvertsen (1997), Hoppenrath et al. (2009), Ashworth et al. (2013) and Sims et al. (2018), among others. The classification system and the taxonomic framework of Guiry and Guiry (2015) was adopted for the present study (www.algaebase.org).
Statistical analyses
Quantitative data obtained from the samples were used to calculate the relative abundance and frequency of occurrence of the phytoplankton species. The quantitative data were also used to estimate species diversity (Shannon 1948) and evenness (Pielou 1977).
The normality (Lilliefors test) and homogeneity of the variances (Levene’s test) of the data were first verified prior to the analysis of the patterns of variation in the hydrological parameters, and in the abundance and biomass of the phytoplankton by station (S1, S2, and S3) and season (dry vs. rainy). When the data were not normally distributed, they were log (x + 1) transformed to reach a near-normal distribution. The parametric Student’s t test for independent samples was used for the data with a normal distribution, and the non-parametric Mann–Whitney (U) test for those with a non-normal distribution. The interactions between sectors, seasonal periods and under climatic events were evaluated by Student–Newman–Keuls analysis performed a posteriori. A 5% significance level was considered for all analyses, which were run in the STATISTICA software, version 8.0.
Hierarchical agglomerative analyses of similarity were used to investigate similarities among the samples, based on the Bray–Curtis similarity index and the log (x + 1) transformed abundance data, and run in the PRIMER statistical package, version 6.1.6 (Clarke and Warwick 1994). A SIMPER (Similarity/distance percentages) analysis was run to determine which species contributed the most for the formation of the groups in the cluster analysis, and a similarity analysis (ANOSIM) was used to test the significance of the differences found between the groups formed in the dendrogram. Both these analyses were run in PRIMER 6.1.6. The Spearman correlation coefficient (r) was used to assess the correlation between the abiotic and biotic variables, and was calculated in the STATISTICA software, version 8.0.
A Canonical Correspondence Analysis (CCA) was used to verify the relationships between the abundance of the different species and environmental variables. Only the species with a relative abundance of over 10% in at least one sample were included in this analysis. The significance of the overall ordering and the first axis were tested using a Monte Carlo permutation test (p < 0.05) with unrestricted permutations. These analyses were run in Canoco 4.5 (Ter Braak and Verdonschot 1995; Hoppenrath et al. 2009).
Results
Influence of anomalous rainfall levels on the environmental variables and the structure of the phytoplankton community
Rainy season during a La Niña event
During the first half of 2011, which coincided with a La Niña event, precipitation increased by 40% in comparison with the long-term (37 year) average for January, and 15% in May, but in the case of the study month (February, April, and June, 2011), there was only an increase, of 23.5% in April, in comparison with the mean (Fig. 2B). The historical mean precipitation was 2622 mm (Fig. 2A). The data recorded during the three campaigns of the first half of 2011 revealed the effect of La Niña on the environmental variables (Fig. 3A–F), with the monthly temporal interactions being shown in Table 1.

Monthly average (± SD) of environmental variables: A temperature and salinity; B pH and dissolved oxygen; C turbidity and silicate; D nitrate and nitrite; E phosphate and ammonium; F chlorophyll-a
Comparing all the study months of this scenario, the lowest mean temperature (28.26 ± 0.24 ºC), pH (7.09 ± 0.25), and DO (4.76 ± 1.22 mg L−1) were recorded in February 2011, while the lowest salinity (13.9 ± 5.48) was registered in April 2011 (Figs. 3A and 2B). The mean turbidity (106.31 ± 35.13 NTU; t = 4.75, p < 0.01), and the nitrite (0.38 ± 0.19 µmol L−1; U = 0.00, p < 0.05), nitrate (6.03 ± 1.55 µmol L−1; U = 0.00, p < 0.05), phosphate (0.62 ± 0.09 µmol L−1; U = 0.00, p < 0.05), and silicate (141.65 ± 82.22 µmol L−1) concentrations were all highest in February 2011 (Fig. 3C, D and E), while the highest concentrations of ammonium (1.92 ± 0.27 µmol L−1; t = − 3.01, p < 0.05) and chlorophyll-a (28.62 ± 17.74 mg m−3) were recorded in April 2011 (Fig. 3E and F). The significant differences recorded among the months are shown in Table 1.
Comparing only the months of the rainy season (RS-LN), total phytoplankton abundance peaked in February (421.35 × 103 ± 122.86 × 103 cells L−1) and June (423.16 × 103 ± 167.76 × 103 cells L−1; Fig. 4A). During this period, diversity (t = 5.13, p < 0.01) and evenness (t = 6.46, p < 0.01) increased significantly between February (diversity = 2.24 ± 0.66; evenness = 0.44 ± 0.13) and June (diversity = 2.89 ± 0.77; evenness = 0.56 ± 0.09), accompanying the increase of salinity during this period (Figs. 3A, 4B and Table 1).

Monthly average (± SD) of total phytoplankton, microphytoplankton and phytoflagellate abundances (A), and evenness and diversity (B) in the area of study
Dry seasons: post-drought event and the effects of a La Niña event
Normally, the dry season accounts for approximately 15% of annual precipitation in the study area, based on the data for the preceding 37 years (1974–2011; see Fig. 2B). The La Niña event of mid-2010 was characterized by higher than normal rainfall (37% higher than the historical mean for the dry season), although it was preceded by an unusually dry rainy season, when rainfall was 63% lower than average, due to the drought event (DS-PD) in the first half of 2010 (Fig. 2B). In turn, the second half of 2011 (dry season) was preceded by the La Niña event in the rainy season (Fig. 2B). Annual rainfall in 2011 was 25% higher than 2010 (Fig. 2A), with 87% of the rains falling during the first half of the year.
The comparison of the two dry seasons (DS-PD vs. DS-LN) revealed significant differences in many of the study variables (Table 1). In the DS-PD dry season, the waters were slightly warmer (29.01 ± 0.33 °C), significantly more saline (37.1 ± 1.3; U = 6.00, p < 0.01), and richer in dissolved oxygen (7.82 ± 0.24 mg L−1), nitrite (0.12 ± 0.01 µmol L−1; U = 6.00, p < 0.001), nitrate (10.95 ± 4.6 µmol L−1; U = 0.00 p < 0.01), and phosphate (1.03 ± 0.46 µmol L−1; t = 2.96, p < 0.05) than the DS-LN (Fig. 3A, B, D and E). The lowest abundance of cells from 20 to 200 µm microphytoplankton (35.16 × 103 ± 5.1 × 103 cells L−1; t = − 8.09, p < 0.05) and phytoflagellates (56.78 × 103 ± 12.73 × 103 cells L−1; t = − 5.46, p < 0.001) were also observed in the DS-PD (Fig. 4A). By contrast, the highest diversity (2.50 ± 0.64) and evenness (0.51 ± 0.11) were also recorded in the DS-PD (Fig. 4B) in comparison with the DS-LN), although neither index varied significantly between the years (Table 1).
When rainfall was higher than average in the first half of the year—as it was in 2011—the dry season (DS-LN) was marked by higher turbidity (33.17 ± 4.89 NTU; U = 15.00, p < 0.05), ammonium (1.81 ± 0.22 µmol L−1; t = − 6.92, p < 0.001), silicate (144.11 ± 78.13 µmol L−1), and chlorophyll-a concentrations (28.62 ± 17.74 mg m−3; U = 15.00, p < 0.05) (Fig. 3C, E and F). The total abundance of phytoplankton (385.40 × 103 ± 100.91 × 103 cells L−1; t = − 5.02, p < 0.01) and microphytoplankton abundances (273.29 ± 127.64 × 103 cells L−1; t = − 8.09, p < 0.01) were also higher when compared with DS-PD (Fig. 4A, Table 1).
Spatial oscillations in the environmental variables during the different climatic events
Here, the spatial oscillations in the study variables will be presented first, followed by the fluctuations in these variables under the different climatic conditions. The means and standard deviations of the environmental variables and phytoplankton parameters recorded at the three sampling stations (S1, S2, and S3) are shown in Fig. 5, and their spatial interactions are presented in Table 1.

Spatial variation (average ± SD) of environmental variables: A temperature and salinity; B pH and dissolved oxygen; C turbidity and silicate; D ammonium and nitrite; E nitrate and phosphate; F chlorophyll-a and total phytoplankton abundance, G microphytoplankton and phytoflagellates abundance; H evenness and diversity
Rainy season during the La Niña event
During the rainy season of the La Niña event (RS-LN), salinity and pH reached their lowest values in the upper sector (S1) due to the increased input of freshwater from the adjacent wetlands, as well as the intrusion of less saline water from the Caeté Estuary through the Taici Creek. Given this, the mean salinity was significantly different between S1 and S3 (t = − 2.81, p < 0.05), ranging from 10.9 ± 5.9 at S1 to 24.9 ± 3.1 at S3, reflecting the greater marine influence (Fig. 5). A similar trend was observed in the pH (t = − 3.05, p < 0.05), which increased from 6.9 ± 0.2 at S1 to 7.4 ± 0.2 at S3, and also in the dissolved oxygen concentrations (t = − 3.63, p < 0.05), which varied 4.22 ± 1.18 mg L−1 at S1 to 6.85 ± 0.32 mg L−1 at S3. The silicate concentrations also decreased significantly from S1 to S3 (t = 4.61, p < 0.05), with a mean of 75.86 ± 18.25 μmol L−1 at S3 and 214.98 ± 33.33 μmol L−1 at S1. A similar pattern was recorded for chlorophyll-a (t = 6.73, p < 0.05), which varied from 13.86 ± 5.01 mg m−3 at S3 to 37.0 ± 12.14 mg m−3 at S1, and the abundance of phytoflagellates (U = 15.00, p < 0.05), which ranged from 173.79 × 103 ± 11.96 × 103 cells L−1 at S3 to 268.39 × 103 ± 122.90 × 103 cells L−1 at S1. In the rainy season of the La Niña year, diversity (2.83 ± 0.34) and evenness (0.55 ± 0.05) were highest in the lower sector (S3), although no significant differences were found among the sectors (Table 1).
Dry seasons: post-drought event and the effects of the La Niña event
The estuary was highly homogeneous during both dry season (DS-PD and DS-LN), with no significant variation being found in any of the study variables (Table 1). During the DS-PD, with the influence of less saline waters from the Caeté Estuary, there was a similar general tendency for increasing temperature (28.5–29.2 °C), salinity (36–39), pH (6.4–7.8), turbidity (11.2–26.4 NTU), and nitrite concentrations (0.11–0.13 µmol L−1) from the upper to the lower sectors (Fig. 5A, B, C and D). The opposite pattern, with values decreasing from S1 to S3 was recorded in chlorophyll-a (10 to 3.15 mg m−3), and the phosphate (1.38–0.51 µmol L−1) and silicate (146.39 to 32 µmol L−1) concentrations, as well as in the diversity (2.94–1.76 bits cell−1) and evenness (0.58–0.38) values (Fig. 5C, E, F and H). In this period (DS-PD), the waters in the middle sector were slightly richer in nitrate (17.85 µmol L−1), and had a higher phytoflagellate abundance (69.72 cells L−1) (see Fig. 5E and G), whereas during the DS-LN, high concentrations of ammonium (2.00 µmol L−1), and a greater abundance of both total phytoplankton (493.52 cells L−1) and microphytoplankton (381.43 cells L−1) were recorded in S2 (Fig. 5D, F and G). Conversely, peaks in silicate (261.30 µmol L−1) and chlorophyll-a (61.58 mg m−3) concentrations were recorded in S1 during this period (Fig. 5C and F). In all sectors, the water was more saline following the drought event, and also had slightly lower chlorophyll-a concentrations, and reduced microphytoplankton and phytoflagellate abundance, as well as the highest diversity and evenness (Fig. 5A, F, G and H), contrary to the pattern observed during the DS-LN.
Composition of the phytoplankton
During the study period, the microphytoplankton (160 taxa) of the Taperaçu Estuary was dominated by the Bacillariophyta (88.1% of the taxa identified), followed by the Myzozoa (8.1%), Cyanobacteria (2.5%), and Chlorophyta (1.3%). The diatom Cymatosira belgica Grunow was the most abundant species (26.4% of total individuals), followed by Skeletonema costatum (Greville) Cleve (11.1%), Psammodictyon panduriforme (W. Gregory) D. G. Mann (9.5%), and Plagiogrammopsis sp. (8.3%). The other taxa contributed 44.7% of the total.
Rainy season during the La Niña event
The total abundance of phytoplankton was high during the rainy season of the La Niña event (RS-LN), when the microphytoplankton represented 40.4% of this abundance, while the phytoflagellates, which were distributed in 100% of the samples, represented 59.6%. Tychoplankton (transitory planktonic organisms) was also more abundant during this period, including Cymatosira belgica, which accounted for 66% of the abundance (one of the highest values recorded in this period) and had a frequency of 100% in February 2010 (Fig. 6A). The dominance of this species in this month was favored by the increased precipitation, which led to an increase in the resuspension of sediments and organic particles, contributing to the accumulation of phytobenthic cells in the water column (tychoplankton), which explains their abundance and the high turbidity recorded during this period (RS-LN).

Temporal (A) and spatial (B) distribution of relative abundance of the main species identified during the study
Skeletonema costatum (relative abundance = 39.2%; frequency = 77.8%) was also most abundant during the rainy season (June), followed by Trieres sinensis (Greville) Ashworth & E.C. Theriot (relative abundance = 12.5%; frequency = 44.4%) and Plagiogrammopsis sp. (relative abundance = 9.1%; frequency = 41.6%), reflecting the influence of the meso-polyhaline environment conditions (Figs. 5A and 6A) on the occurrence and dynamics of these species. Thalassiosira sp.2 was well represented in April (relative abundance = 15.4%; frequency = 61.1%), with higher values than those recorded in the dry season (Fig. 6A).
In terms of spatial variation (Fig. 6B) during the La Niña (RS-LN) event, Cymatosira belgica and Plagiogrammopsis sp. reached their highest relative abundance in the upper sector (38.7% and 6.9%, respectively) and were least abundant in the lower sector (25.7% and 1.8%), probably favored by the low salinity, high turbidity (high resuspension rates), and higher ammonium concentrations in S1. Thalassiosira sp.2, in turn, was most abundant in the upper sector (6.7%). The opposite pattern was observed in Skeletonema costatum and Trieres sinensis with high abundances in the middle (22.9% and 8.5%, respectively) and lower (20.5% and 6.7%) sectors and the lowest abundances in the upper sector (17.3% and 0.7%), favored by the high salinity and nutrient concentrations (ammonium, nitrate, and phosphate) recorded in these sectors during this period.
Dry seasons: post-drought event and the effects of the La Niña event
During the dry season, fluctuations in the abundance of the microphytoplankton community followed distinct patterns under La Niña (DS-LN; 51.8%) and post-drought (DS-PD; 38.2%) conditions, with a predominance of phytoflagellates (61.8%) in the DS-PD. The post-drought period was marked by more saline and less turbid waters, when the most abundant and frequent species (Fig. 6A) were Psammodictyon panduriforme (relative abundance = 52.9% and frequency = 100%), Cymatosira belgica (relative abundance = 9.9%; frequency = 38.9%), and Plagiogrammopsis sp. (relative abundance = 8.5%; frequency = 16.7%). The reduced abundance of the latter two species in this period (DS-PD) reflects their reduced physiological capacity for development at a salinity of above 35 (Figs. 3A and 6B). Comparing the two dry seasons (DS-PD vs. DS-LN), the lower salinity (< 35) and higher turbidity recorded during the La Niña event, resulted in a higher relative abundance (34.7%) and frequency (100%) of C. belgica, reflecting its tolerance of oscillations in salinity and the generally high salinity of the Taperaçu Estuary, as well as the importance of the resuspension processes in its hydrodynamics, even during the dry season. By contrast, the relative abundance and frequency of P. panduriforme (31.9% and 88.4%, respectively) and Plagiogrammopsis sp. (6.6% and 66.7%) decreased in this period (Fig. 6A).
Spatially, Cymatosira belgica and Plagiogrammopsis sp. were most abundant in the upper sector (Fig. 6B) probably due to the lower salinity, shallower water (high resuspension rates) and greater availability of dissolved nutrients. The highest relative abundance (64.1% for C. belgica and 20.9% for Plagiogrammopsis sp.), both with a frequency of 100%, was recorded in the dry season during the La Niña effect (DS-LN) in the upper sector (Fig. 6B). By contrast, Psammodictyon panduriforme was the most abundant species in the dry season (DS-PD), especially in the lower sector (DS-PD, 65.9%; DS-LN, 35.1%, both with 100% of frequency), as a consequence of the high salinity. In the post-drought (DS-PD), this species was also relatively abundant in the middle (52.3%) and upper (37.8%) sectors, when salinity was higher than 35 (Figs. 5A and 6B). In the comparison of the two dry seasons (DS-PD vs. DS-LN), Skeletonema costatum and Trieres sinensis were more abundant during La Niña (DS-LN), primarily in the middle sector (Fig. 6B; October 2011: S. costatum = 32.1% and T. sinensis = 10.2%) due to the polyhaline conditions. Thalassiosira sp. 2 was also more abundant in the La Niña dry season (DS-LN), principally in the lower sector (Fig. 6B).
Canonical correspondence analysis (CCA)
The first two axes of the CCA explained 53.7% of the obtained variance (Fig. 7) with the first axis (29.4%) revealing the association of the samples obtained in June (S1, S2 and S3) and those of the middle and lower sectors in October 2011, both under the influence of La Niña, reflecting the positive correlation of Skeletonema costatum and the ammonium concentration under conditions of low turbidity.

Canonical Correspondence Analysis (CCA) of the environmental variables with the most common phytoplankton species: Cymat = Cymatosira belgica; Plagi = Plagiogrammopsis sp.; Skele = Skeletonema costatum; P.laut = Planctonema lauterbornii; P.pand = Psammodictyon panduriformis; T.frauen = Thalassionema frauenfeldii; T.sine = Trieres sinensis
The second axis (24.3%) indicated a clear separation between the upper sector under La Niña (RS-LN; DS-LN), and the post-drought conditions (DS-PD; S1, S2, and S3 in October 2010), reflecting the influence of silicate concentrations and, to a lesser extent, of turbidity, on the occurrence of the diatom Cymatosira belgica in its positive portion, which may explain the increase in the chlorophyll-a concentrations on this axis. The negative portion (axis 2) included the DS-PD, DS-LN except for S3 in October 2011, and most of the data from S2 and S3 in the rainy season, with a predominance of Thalassionema frauenfeldii (Grunow) Tempère & Peragallo and Psammodictyon panduriforme, which were associated with high salinity and high concentrations of nitrate, nitrite and phosphate. In the negative portion of this axis (lower quadrants), the sectors closest to the coast were grouped, regardless of the study period, reflecting the incursion of marine waters into the estuary, as well as the low precipitation rates especially in the DS-PD (October 2010).
Cluster analysis
The cluster analysis based on the abundance of the microphytoplankton species revealed three well-defined groups with 50% similarity (Fig. 8). The separation of the groups reflected the influence of the post-drought (DS-PD) and La Niña (RS-LN; DS-LN) climate events (ANOSIM global R = 0.513, p < 0.05), with no spatial or monthly patterns being observed.

Cluster analysis (A) and relative abundance of the principal microphytoplankton species (B) identified during the study
Group 1 was composed of the samples of the post-drought dry season (DS-PD; October 10), with 61.4% similarity. This group presented the highest phytoflagellate abundance and the lowest microphytoplankton abundance, with low chlorophyll-a concentrations, with a predominance of Psammodictyon panduriforme in the lower sector, where salinity was highest (Fig. 8). According to the SIMPER, Paralia sulcata (Ehrenberg) Cleve and Nitzschia sp.1 Hassall (SIMPER (Sim/SD) = 48.5), which are marine taxa adapted to saline environments, were the principal species responsible for the formation of this clade.
The second and third groups were influenced by the La Niña event (RS-LN; DS-LN in October 2011), with the greatest abundances being related to the highest concentrations of dissolved nutrients, in particular, ammonium and silicate, which thus contributed to the high biomass (chlorophyll-a) recorded in this period. With a similarity of 51%, this group associated samples from the three sectors in February and April (RS-LN), as well as the upper sector of October 2011 (DS-LN), which presented the lowest salinity, high turbidity, as well as high silicate and ammonium concentrations (as shown in Fig. 5A, C and D). Freshwater and brackish species, such as Tryblionella granulata (Grunow) D G Mann (SIMPER (Sim/SD) = 52.6), Campylosira cymbelliformis (A W F Schmidt) Grunow ex Van Heurck (SIMPER (Yes/SD) = 21.6), and Cyclotella meneghiniana Kützing (SIMPER (Yes/SD) = 16.1) were the main taxa responsible for the formation of this group, and their presence have been probably favored by mesohaline water conditions. This group was divided into two subgroups, with all the February samples and two samples (S1 and S3) from April (RS-LN), characterized by high phosphate, nitrite and chlorophyll-a concentrations, forming subgroup 2a. Subgroup 2b, which includes the samples from the middle (April) and upper (October 2011) sectors, was characterized by high silicate concentrations and a high abundance of Cymatosira belgica, confirming the influence of the La Niña event on the October 2011 sample (DS-LN; see also abundance and the CCA), which contributed considerably to the high phytoplankton biomass (chlorophyll-a).
Nitzschia longissima (Brébisson) Ralfs (SIMPER (Sim/SD) = 16.5) was responsible for the association of the samples in the third group, which presented 67% similarity and consisted of the samples from the middle and lower sectors obtained in June. At these stations, the abundance of Skeletonema costatum prevailed due to the moderate salinity and high ammonium concentrations, which provided favorable conditions for the development of this species.
Correlations between the environmental and biological variables
The Spearman correlation coefficients calculated for the study variables are shown in Table 2. The correlation analyses showed that the high precipitation rates (especially in the rainy season—the RS-LN) recorded during the present study had a direct influence on both the hydrological and the biological variables, in particular salinity. A significantly negative correlation was recorded between these two variables (rs = − 0.89; p < 0.001).
Significant positive correlations were recorded between the microphytoplankton abundance and the pH and ammonium concentrations. The abundance of phytoflagellates was also correlated with both precipitation and turbidity. Significant correlations were also recorded between total phytoplankton and microphytoplankton abundance, as well as between the chlorophyll-a and silicate concentrations. Significant negative correlations were recorded, in turn, between microphytoplankton and nitrite concentrations, between phytoflagellates and salinity and dissolved oxygen, between total phytoplankton and phosphate concentrations, and also between chlorophyll-a concentrations and salinity.
Discussion
Natural condition of environmental variables in Amazon coast
The dynamics of estuarine ecosystems are closely related to climatic factors such as temperature, winds, and precipitation, which together with other physical and chemical variables, are crucial for the development, maintenance, and distribution of phytoplankton in these environments. In equatorial regions, the climate is defined in general by high and stable temperatures and high precipitation rates (Nittrouer and DeMaster 1996). Rainfall is one of the principal factors controlling this primary production and phytoplankton biomass by altering the salinity (Sousa et al. 2009; Pereira et al. 2010; Matos et al. 2011), dissolved nutrient concentrations (Santos et al. 2008; Pamplona et al. 2013), and transparency (turbidity) of the water (Bastos et al. 2005).
On the Amazon coast, the climatic pattern is influenced directly by the seasonal displacements of the Intertropical Convergence Zone—ITCZ (Figueroa and Nobre 1990). In the first half of the year, the ITCZ shifts to the Southern Hemisphere, leading to an increase in precipitation and thereby, in fluvial discharges. Then, during the second half, the ITCZ shifts northward, leading to a reduction in rainfall levels and consequently in fluvial discharges, creating drier conditions on the Amazon coast (Marengo 1995; Pereira et al. 2013). The displacement of the ITCZ is also influenced by events such as El Niño, La Niña, and droughts, which may also change precipitation patterns significantly (Marengo et al. 2015). In 2010, a drought event related to global warming was responsible for an increase in sea surface temperatures, leading to a reduction (see Fig. 2A) in precipitation rates in the Amazon region (Marengo et al. 2013; Pereira et al. 2017). A La Niña event occurred in 2011, resulting in an increase in precipitation rates in both the rainy and dry seasons of this year. In the Taperaçu Estuary, the effects of these changes in rainfall rates are even more accentuated, given the lack of any continuous freshwater inflow from a river but only from adjacent marshes, primarily during the rainy period (Asp et al. 2012; Magalhães et al. 2015; Oliveira et al. 2021). The increased rainfall recorded in the Amazon region in first semester of 2011 (see Fig. 2B) led to an increase in the fluvial discharge of the Amazon and its tributaries, contributing to the flooding of coastal areas (Espinoza et al. 2012; Marengo et al. 2013; Pereira et al. 2013; 2017) and promoting changes in the hydrological characteristics of the local small estuaries, such as the Taperaçu, as observed in the present study.
On the Amazon coast, under normal climate conditions, the fluvial discharge of the Amazon River and its tributaries, and the strong local hydrodynamics are responsible for the availability of dissolved nutrients and organic matter in the water column. These high concentrations, which normally exceed those recorded in other, similar regions around the world (Thompson et al. 2015; Pan et al. 2016), including other parts of the Brazilian coast (Rodrigues et al. 2009; Islabão et al. 2017) have a direct effect on the biological productivity of the waters of the Amazon coast, which is also very high. The distribution of these elements is related primarily to the morphological characteristics of the environment, the residence time of the water, the tidal regime and precipitation, which together determine the nutrient input to the estuary (Dittmar and Lara 2001; Pamplona et al. 2013). These factors influence the biogeochemical processes during the transport of nutrients, thus controlling the distribution, flow, and destiny of these elements within the estuarine system, and influencing phytoplankton communities, which are considered to be a key element/group for the assessment of eutrophication in estuarine systems, due their rapid response to environmental variation (Domingues et al. 2008; Garmendia et al. 2013).
Given this dynamic, marked seasonal variation was observed in salinity over the study period, and the values recorded were invariably higher during the dry season. This seasonal pattern of variation in salinity is a characteristic of coastal environments, in particular those influenced strongly by marine waters (Lacerda et al. 2004; Magalhães et al. 2006), and has been reported previously for other Amazon estuaries (Matos et al. 2011; Pamplona et al. 2013). This was confirmed here in the case of the Taperaçu Estuary, as in previous studies in this estuary (Costa et al. 2008; Magalhães et al. 2013; Palma et al. 2013), where the restricted input of freshwater was considered one of the principal factors controlling the invariably high salinity recorded throughout the year.
Effects of climatic events on environmental variables and phytoplankton community structure
Salinity is one of the principal variables controlling the physiological processes and, consequently, the composition and distribution of the organisms that inhabit estuarine systems, as well as forming an ecological barrier for many species (Feitosa et al. 1999; Dorado et al. 2015), including some phytoplankton (Pinckney et al. 2017). During the study period, salinity was lowest during the rainy period, especially in the case of the La Niña event. Salinity varied spatially within the estuary only during the rainy season (RS-LN; see Table 1), reflecting the absence of any continuous freshwater river input, which renders the estuary homogeneous during periods of low rainfall (dry season). The increase in rainfall during the rainy season separates the upper sector completely from the lower sector, however, which retains its more saline characteristics due to its contiguity with the adjacent coastal marine waters. During the flood tide, there is also an incursion of oligohaline waters from the neighboring Caeté Estuary into the middle sector of the Taperaçu Estuary through the tidal Taici channel (Asp et al. 2012; Magalhães et al. 2015). Even so, this inflow may have only a secondary effect on salinity, given the lack of any significant effect during the dry season.
The dissolved oxygen concentration also varied among period, peaking in the La Niña dry season. A similar pattern has been observed in coastal ecosystems in other regions of the world during the dry season (Losada et al. 2003; Santos and Muniz 2010; Monteiro et al. 2015), including the Amazon coast (Sousa et al. 2009; Matos et al. 2012), where the increase in river runoff during the rainy season has a direct impact on the transparency (turbidity) of the water by increasing the amount of particulate matter in the water column, thus promoting oxygen consumption (oxidative processes) and reducing photosynthesis (Flores Montes et al. 2002; Gardner et al. 2006; Santos et al. 2008, 2012). These processes are even more accentuated during La Niña events, such as that observed in the present study, during the first semester of 2011 (see Fig. 3B). Although photosynthesis (together with respiration) also influences the pH through the uptake of dissolved carbon dioxide and the liberation of oxygen into the water column (Flores Montes et al. 2002; Sousa et al. 2009; Matos et al. 2012; Costa et al. 2013a), the alkaline water observed throughout the study period can be explained primarily by the buffer effects of the marine waters (Macêdo et al. 2000) coming into the estuarine system during the flood tide. In contrast with other Amazon estuaries, which undergo drastic spatial and seasonal variation in pH over the course of the year (Costa et al. 2009; Matos et al. 2011), the absence of any systematic freshwater input into the Taperaçu was most likely the principal factor responsible for the lack of any clear spatial and temporal variation in the pH of this estuary. Even so, rainfall did influence the spatial variation in the pH between the upper (low pH) and lower (high pH) sectors in the rainy season during La Niña (RS-LN).
Turbidity was high in the Taperaçu Estuary during the rainy season, as observed in other Amazon estuaries (Lam-Hoai et al. 2006; Costa et al. 2009; Gomes et al. 2013; Monteiro et al. 2015). The turbidity recorded during this period reflected the joint effects of high rainfall rates and strong local hydrodynamics (macrotides), which promoted the resuspension of organic and inorganic matter from the bottom and from the estuary margins into the water column, accentuating its turbidity (Costa et al. 2009). This reduces the photic zone, resulting in a decrease in the availability of light energy available for absorption by the phytoplankton, and limits photosynthesis and consequently the growth of these organisms (Azevedo et al. 2008; Gameiro et al. 2011; Lancelot et al. 2011; Matos et al. 2012). This indicates that in the Taperaçu, like other Amazon estuaries, turbidity is one of the principal factors limiting phytoplankton growth, given that most nutrients are available in the water column at high concentrations (Pamplona et al. 2013; Goes et al. 2014; Oliveira et al. 2021; Queiroz et al. 2022). While intense sunlight is a characteristic of tropical regions worldwide (Lam-Hoai et al. 2006; van der Molen & Perissinotto, 2011), the turbidity of the water may block its penetration, in particular during the rainy season, thereby limiting phytoplankton growth. This effect is expected to be even greater during La Niña events since precipitation rates and freshwater flow are even greater as observed in the present study. Previous studies in other coastal Amazon environments (Santos and Muniz 2010; Costa et al. 2013a; Goes et al. 2014) have recorded chlorophyll-a peaks—an indirect measure of phytoplankton growth—primarily during the rainy season (Sousa et al. 2009; Costa et al. 2011; Matos et al. 2012; Queiroz et al. 2022). However, these findings tend to refer primarily to data obtained at the onset of the rainy season, when there is a joint effect of increasing nutrient concentrations and sunlight availability, given that, since in the Amazon region, there is a major gap between the onset of the rainy season and the peak in river discharge (Goes et al. 2014).
In the Taperaçu Estuary, the greatest abundance of phytoplankton and the highest chlorophyll-a concentrations were recorded during the La Niña periods (RS-LN and DS-LN), when turbidity peaked, especially in the upper and middle sectors of the estuary. These findings indicate that the greater flow of marine waters into the inner portion of the estuary during the post-drought period (DS-PD), increases salinity and decreases the concentration of nutrients, in particular silicate and ammonium, creating one of the principal barriers to the growth of phytoplankton during this period. Under La Niña conditions, Cymatosira belgica was predominant, with a relative abundance of 66% in February (RS-LN; Fig. 6A), and 64.1% in the upper sector in the dry season (DS-LN; Fig. 6B). Skeletonema costatum, Trieres sinensis, and Thalassiosira sp.2 were also more abundant during the anomalous rainy season, influenced by high concentrations of dissolved nutrients, such as silicate and ammonium, reflecting the high consumption of part of the nitrate available in the water column during this period. Plagiogrammopsis sp., in turn, was most abundant during the post-drought (DS-PD) and La Niña (DS-LN) dry seasons (see Fig. 6A and B). In the present study, C. belgica was most abundant during the La Niña periods, in particular in the upper sector, when silicate concentrations and turbidity were higher, reflecting the effects of the local hydrodynamics on the increase in the contribution of benthic species to the water column. The spatial variation in the composition and biomass of phytoplankton coincided with variations in the silicate concentration, which indicates that this nutrient may become a limiting factor for the development of phytoplankton (diatoms) in the La Niña periods in the study estuary, in particular in the lower sector, and possibly also in other small Amazon estuaries with similar characteristics (see Figs. 5 and 6). On the other hand, the positive correlation between chlorophyll-a, turbidity, and ammonium (see Table 2) indicates that the resuspension of microphytobenthic species contributed to the increase in phytoplankton biomass, as observed here and in previous studies in other tropical estuaries (Matos et al. 2011; Pamplona et al. 2013; Bharathi et al. 2018), reflecting the active mixing of the water column.
Phytoplankton biomass, as indicated by chlorophyll-a, presented clear seasonal variation, with significantly higher concentrations observed during the rainy season. This same seasonal pattern has been observed in other estuarine systems along the Brazilian coast (Grego et al. 2004; Matos et al. 2012). Sassi and Kutner (1982) concluded that this pattern of seasonal variation is characteristic of coastal environments, where the high biomass observed during the rainy season is associated with the peaks in nutrient levels resulting from the increased input of these elements from the drainage basin. Increased local hydrodynamics, the availability of suspended particulate matter, and the resuspension of benthic microalgae in the water column (Lara and Dittmar 1999; Pereira-Filho 2001; Matos et al. 2012), may also have contributed to the increase in biomass.
In aquatic ecosystems, phytoplankton biomass and production are limited by the combination of the various types of nutrient salts, that is, nitrite, nitrate, ammonium, phosphate and silicate (Tundisi and Tundisi 1976), that are needed for the growth and reproduction of the microalgal community (Pannard et al. 2008; Abreu et al. 2010; Goes et al. 2014). In the estuaries of the Amazon coast, however, where dissolved nutrient concentrations tend to be high values, nutrients do not appear to be a limiting factor (Costa et al. 2011; Pamplona et al. 2013; Oliveira et al. 2021). This is also true for the adjacent marine ecosystems, and the Taperaçu, like other small Amazon estuaries with reduced freshwater input, which plays an important role in this coastal enrichment process, given that almost all the nutrient and chlorophyll-a rich waters retained in this ecosystem flow into the adjacent coastal zone during each tidal cycle (Asp et al. 2012; Costa et al. 2013a), thus replenishing the coastal waters twice a day.
It is well known that nutrient enrichment has a significant effect on the phytoplankton community, altering its composition, diversity, and abundance (Piehler et al. 2004; Buyukates and Roelke 2005). In extreme cases, the input of nutrients into the aquatic environment leads to the proliferation of microalgae, often favoring the growth of some species over others. In the present study, the silicate concentrations varied significantly in space, with higher values being recorded in the upper sector, especially during the La Niña event. While a similar pattern was observed in the post-drought dry season, the concentrations were lower. The intensification of the anomalous rainfall pattern (La Niña) was probably responsible for the increase in silicate concentrations in the estuary, given that it promotes the leaching and erosion of the estuary margins and adjacent areas. The increasing silicate content in the water column in the upper sector appears to have created favorable conditions for the growth of the phytoplankton and microphytobenthos resuspended into the water column, especially the diatoms. The greatest abundance of resuspended Cymatosira belgica cells, and thus, the absorption of silicate, probably contributed to the observed increase in biomass (chlorophyll-a) in this period. During the rainy season, in addition, the greater volume of freshwater from the Taperaçu basin and oligohaline water from the Caeté Estuary probably also contributed to the greater concentration of nutrients observed in the inner sectors (S1 and S2) of the estuary. By contrast, low silicate concentrations in the lower sector (S3) were probably related to utilization by the phytoplankton and/or dilution processes, given the abundance of phytoplankton and the influence of marine waters at this station.
The composition and dynamics of the phytoplankton community of the Taperaçu Estuary was influenced considerably by climatic conditions and environmental variables. Diatoms dominated the microphytoplankton in both species’ richness and abundance, which is consistent with the findings of previous studies in estuarine and marine environments of the Amazon coast (Sousa et al. 2009; Santana et al. 2010; Costa et al. 2011, 2013b; Matos et al. 2011, 2013; Silva et al. 2010). Diatoms are the most common group in estuarine ecosystems and correspond to the principal primary producers of these environments (Procopiak et al. 2006). In these areas, the abundance of the diatoms reflects their ecophysiological adaption to oscillations in salinity (Karsten et al. 2006) and eutrophic conditions (Eskinazi-Leça et al. 2004; Silva el al. 2010; Canani et al. 2011). The results of the present study indicate that the increase in precipitation resulting from the La Niña event promoted a decrease in salinity and an increase in dissolved nutrient concentrations, thus increasing phytoplankton biomass, which was further increased by the contribution of resuspended benthic diatoms, including Cymatosira belgica, which is well adapted to low light conditions and more turbulent waters.
The high frequency of diatoms observed in the present study testifies to their excellent capacity to adapt to the oscillations in physical and chemical water characteristics (primarily salinity). The taxa of the Myzozoa, Cyanobacteria, and Chlorophyta were much less abundant, however. The most important species in terms of frequency of occurrence and relative abundance were: Cymatosira belgica, Psammodictyon panduriforme, Plagiogrammopsis sp., Skeletonema costatum, Thalassiosira sp.2, and Trieres sinensis. The latter, a marine species that was abundant during the dry season, is known to be highly adaptable to coastal environments with varying salinity levels, such as the Taperaçu Estuary, and other estuaries studied by Bergesch et al. (2009) and Eskinazi-Leca et al. (2010). The tychoplankton C. belgica peaked in abundance during both the rainy and dry seasons, in all sectors, although it was more abundant in the upper sector, primarily under the influence of La Niña. This species had a positive relationship with the concentrations of silicate and chlorophyll-a, and water turbidity, as shown by the CCA for the upper sector in February, April, and October 2011 (La Niña). Although the species of the genus Cymatosira are considered to be marine organisms, the local conditions of the Taperaçu were especially favorable for its growth, given the marine influence on this estuary and its strong local hydrodynamics, which provide optimum conditions for its development.
The abundance of phytoplankton peaked during the rainy season, reflecting the contribution of precipitation and the local drainage system to the dynamics of these organisms. In this season, microalgal growth was possibly favored by the increased availability of nutrients carried into the estuarine waters from the flushing of the local mangroves, and from the nutrient-rich waters of the Caeté Estuary. This seasonal pattern of variation has been recorded in many different environments on the Amazon coast (Paiva et al. 2006; Sousa et al. 2009; Carmona et al. 2010; Matos et al. 2012), which reflected the impact of rainfall on the lixiviation of adjacent land and the high level of connectivity among the local estuaries.
Phytoflagellates dominate in shallow coastal environments with calm and transparent waters (Smayda 1980), and may be especially common in estuarine environments during the dry season (Brandini 1982; Lacerda et al. 2004). In the present study, by contrast, the phytoflagellates dominated the local phytoplankton community during the rainy season. Even so, similar results from adjacent coastal environments corroborate our findings, by showing that these organisms can adapt to strong hydrodynamic conditions (Santos-Fernandes et al. 1998; Bastos et al. 2011; Masuda et al. 2011; Matos et al. 2011, 2012, 2016). In particular, the nanoplankton fraction (phytoflagellates) is extremely important in coastal ecosystems (Santos-Fernandes et al. 1998), given that these organisms are responsible for most of the primary production and the available chlorophyll-a in these environments. In this context, the direct relationship observed between phytoflagellate abundance and chlorophyll-a concentrations may indicate that, during the rainy season, these organisms have been the principal group responsible for the peak in the chlorophyll-a concentrations recorded in the water column during this period.
The highest phytoplankton diversity and evenness were recorded during June, and in the upper sector in October 2010 (DS-PD), while the highest species richness was recorded during La Niña, in both seasons. The abundance of Psammodictyon panduriforme peaked during the post-drought period (DS-PD), when salinity increased significantly, inhibiting the development of other organisms. The high levels of precipitation during La Niña were thus responsible for significant changes in salinity and the increasing contribution of freshwater to the estuary, which altered the chemical composition of the nutrients and together had a significant impact on the composition of the phytoplankton in the Taperaçu Estuary. In terms of the phytoplankton composition, while the upper and middle sectors behaved similarly during La Niña (RS-LN and DS-LN), the middle and lower sectors were comparable during the post-drought period (DS-PD). This suggests that changes in the physical and chemical variables of the water resulted from natural variations in the intensity of the climate are responsible for significant changes in the phytoplankton community in other small Amazonian estuaries with similar characteristics to those of the Taperaçu.
The results of SIMPER indicated that, in addition to the peak in abundance of Cymatosira belgica and Plagiogrammopsis sp. (as confirmed by the CCA—see Fig. 7), the benthic diatoms Tryblionella granulata, Campylosira cymbelliformis, Cyclotella meneghiniana, and Nitzschia longissima were the most common taxa during La Niña (RS-LN and DS-LN). This indicates that a large proportion of the identified phytoplankton taxa were derived from sediment resuspension by the intense local hydrodynamics, which is further increased by the anomalous effects of the intense precipitation of the La Niña event. By contrast, the environmental conditions recorded in post-drought period (DS-PD), when precipitation was low and salinity increased due to the greater inflow of marine water (Pereira et al. 2017), resulted in changes in the abundance of the phytoplankton community, which was composed primarily of marine species. This was confirmed by the greater contributions of Paralia sulcata and Nitzschia sp.1 in the SIMPER analysis. These results emphasize the importance of the high levels of precipitation during the La Niña period, which contributed to the abundance and representativeness of the species, which consisted primarily of benthic diatoms. During these events, the low salinity and high turbidity left the waters of the Amazon coast rich in dissolved nutrients and highly oxygenated, with a high biomass, as observed in the studies of Andrade el al. (2016) and Pereira et al. (2017). Similar patterns have been observed in other tropical regions of the world by Thompson et al. (2015) and Gorgues et al. (2010).
The attributes of an ecosystem can be used to determine the factors that influence the dynamics of the phytoplankton community. The results of the CCA indicated clearly the influence of the climatic events (drought and La Niña) on environmental variables, which affect the composition and abundance of the phytoplankton community. Phytoplankton abundance peaked during the La Niña rainy season (see Fig. 7) in the upper and middle sectors of the Taperaçu Estuary in February, April, and June, when the water was more turbid, and had higher concentrations of chlorophyll-a, silicate and ammonium. The dominance of the benthic diatom Cymatosira belgica, and the abundance of Plagiogrammopsis sp. and Skeletonema costatum during this period reflected the strong mixing of the water column. The occurrence of S. costatum, based on the CCA, was similar at all the three stations sampled in June, and in the middle station (S2) in October 2011 (DS-LN), indicating a strong correlation between this taxon and the concentrations of silicate and ammonium. This indicates that the consumption of these easily assimilated nutrients is advantageous for the development of this diatom, which was recorded during both La Niña seasons, being abundant at different salinities and co-occurring with Trieres sinensis. Both these species are generally considered to be marine and cosmopolitan (Kooistra et al. 2008; Bergesch and Odebrecht 2009), although the results of the present study highlight the euryhaline character and adaptability of these taxa to variations in salinity. In October 2010 (DS-PD), samples from all sectors (see the CCA) had the highest abundances of Psammodictyon panduriforme, showing that this species prefers more saline waters, but may also tolerate varying levels of salinity (Matos et al. 2016; Costa et al. 2016).
Overall, the Taperaçu Estuary is a shallow and highly dynamic environment in which significant spatial and seasonal variation was observed in many of the abiotic and biotic variables analyzed. This variation was observed in relation to the effects of climate events such as La Niña and the drought “of the century”, of 2010. The variation in the rainfall cycle, the influence of the tides (hydrodynamics), together with the high nutrient concentrations and the marine incursions (primarily in the dry season), all appear to be the principal factors controlling the dynamics of the phytoplankton community. The occurrence of many species of this community in both seasons, and in both years, reflects their capacity to adapt to major fluctuations in salinity. During the post-drought dry season, high salinity and lower precipitation rates favored the development of Psammodictyon panduriforme—a typical marine species—which became the most abundant taxon during this period. The dominance of this species, primarily in the dry season, was in part responsible for the low diversity indices and evenness observed in the Taperaçu Estuary, which has no major input of freshwater, and can be considered to be a typically marine environment during this period, favoring P. panduriforme. During the periods of high rainfall, that is, the rainy season (for this year RS-LN) and, to a lesser extent, DS-LN, and increased freshwater input from adjacent areas, salinity decreased to its lowest level, which permitted the development of other species of diatom, such as Cymatosira belgica, Plagiogrammopsis sp. and Skeletonema costatum. The input of oligohaline waters from the Caeté Estuary through the Taici creek may also have transported other phytoplankton species into the Taperaçu Estuary, contributing to the diversity of its phytoplankton community. Transport between these two estuaries has already been reported in the literature for sediments (Araújo and Asp 2013), but has yet to be confirmed for phytoplankton in future studies.
Conclusions
The results of the present study indicate that the dynamics of the phytoplankton community of the Taperaçu Estuary is highly influenced by seasonality of rainfall, which is intensely increased by anomalous climate oscillations (La Niña) and directly affect environmental variables (mainly salinity, turbidity and dissolved nutrient concentrations). Diatoms were essentially the dominant group year around and benthic species (tychoplankton) such as Cymatosira belgica, Plagiogrammopsis sp., Tryblionella granulata, Campylosira cymbelliformis, Cyclotella meneghiniana, and Nitzschia longissima were the most common microphytoplankton taxa during La Niña (RS-LN and DS-LN), thus indicating that many phytoplankton taxa were derived from sediment resuspension by the intense local hydrodynamics, which is further increased by intense precipitation observed during this event. Phytoflagellates numerically dominated the local phytoplankton community during the rainy season, showing the great adaptability of these organisms to strong local hydrodynamic conditions. This indicates that these organisms may be one of the main responsible for the highest chlorophyll-a concentrations recorded in the water column during the rainy season. Similar patterns would be expected in the dozens of other estuaries that have similar hydrological, hydrodynamic, and geomorphological characteristics, not only on the Amazon coast, but in many other tropical regions around the world.
Data availability
Data will be available on reasonable request.
References
Abreu PC, Bergesch M, Proença L, Garcia C, Odebrecht C (2010) Short- and long-term chlorophyll a variability in the shallow microtidal Patos Lagoon estuary, Southern Brazil. Estuaries Coasts 33:554–569. https://doi.org/10.1007/s12237-009-9181-9
Anagnostidis K, Komérek J (1988) Modern approach to the classification system of Cyanophytes. 3—Oscillatoriales. Archiv Hydrol Suppl Algol Stud 50(53):327–472
Andrade MP, Magalhães A, Pereira LCC, Flores-Montes MJ, Pardal EC, Andrade TP, Costa RM (2016) Effects of a La Niña event on hydrological patterns and copepod community structure in a shallow tropical estuary (Taperaçu, Northern Brazil). J Mar Syst 164:128–143. https://doi.org/10.1016/j.jmarsys.2016.07.006
Araújo WP Jr, Asp NA (2013) Hydrodynamic connectivity between two macrotidal Amazonian estuaries. J Plankton Res SI 65(10065):1086–1091. https://doi.org/10.2112/SI65-184.1
Ashworth MP, Nakov T, Theriot EC (2013) Revisiting Ross and Sims 1971: toward a molecular phylogeny of the Biddulphiaceae and Eupodiscaceae Bacillariophyceae. J Phycol 49(6):1207–1222. https://doi.org/10.1111/jpy.12131
Asp NE, Schettini CAF, Siegle E, Silva MS, Brito RNR (2012) The dynamics of a frictionally-dominated Amazonian Estuary. Braz J Oceanogr 60(3):391–403. https://doi.org/10.1590/S1679-87592012000300011
Asp NE, Gomes VJC, Schettini CAF, Souza-Filho PWM, Siegle E, Ogston AS, Nittrouer CA, Silva JNS, Nascimento WR Jr, Souza SR, Pereira LCC, Queiroz MC (2018) Sediment dynamics of a tropical tide-dominated estuary: turbidity maximum, mangroves and the role of the Amazon River sediment load. Estuar Coast Shelf Sci 214:10–24. https://doi.org/10.1016/j.ecss.2018.09.004
Azevedo ACG, Feitosa FAN, Koening ML (2008) Distribuição espacial e temporal da biomassa fitoplanctônica e variáveis ambientais no Golfão Maranhense, Brasil. Acta Botanica Brasilica 22(3):870–877. https://doi.org/10.1590/S0102-33062008000300022
Barrera-Alba JJ, Abreu PC, Tenenbaum DR (2019) Seasonal and inter-annual variability in phytoplankton over a 22-year period in a tropical coastal region in the southwestern Atlantic Ocean. Cont Shelf Res 176:51–63. https://doi.org/10.1016/j.csr.2019.02.011
Bastos RB, Feitosa FAN, Muniz K (2005) Variabilidade espaço-temporal da biomassa fitoplanctônica e hidrologia no estuário do rio Una (Pernambuco—Brasil). Trop Oceanogr 33(1):1–18. https://doi.org/10.5914/tropocean.v33i1.5066
Bastos RB, Feitosa FAN, Koening ML, Machado RCA, Muniz K (2011) Caracterização de uma zona costeira tropical (Ipojuca, Pernambuco-Brasil): produtividade fitoplanctônica e outras variáveis ambientais. Braz J Aquatic Sci Technol 15(1):01–10. https://doi.org/10.14210/bjast.v15n1.p1-10
Bergesch M, Garcia M, Odebrecht C (2009) Diversity and morphology of Skeletonema species in southern Brazil, southwestern Atlantic Ocean. J Phycol 45:1348–1352. https://doi.org/10.1111/j.1529-8817.2009.00743.x
Bharathi MD, Sarma VVSS, Ramaneswari K (2018) Intra-annual variations in phytoplankton biomass and its composition in the tropical estuary: influence of river discharge. Mar Pollut Bull 129(1):14–25. https://doi.org/10.1016/j.marpolbul.2018.02.007
Braak CJF, Verdonschot PFM (1995) Canonical correspondence analysis and related multivariate methods in aquatic ecology. Aquatic Sci 57:255–289. https://doi.org/10.1007/BF00877430
Brandini FP (1982) Variação nictemeral de alguns fatores ecológicos na região de Cananéia (SP). Arq Biol Tecnol 25:313–327
Brasil (2005) Decreto de 20 de maio de 2005. Dispõe sobre a criação da Reserva Extrativista Marinha de Caeté-Taperacú, no município de Bragança, no Estado do Pará, e dá outras providencias. Senado Federal, Brasília-DF
Buyukates Y, Roelke D (2005) Influence of pulsed inflows and nutrient loading on zooplankton and phytoplankton community structure and biomass in microcosm experiments using estuarine assemblages. Hydrobiologia 548:233–249. https://doi.org/10.1007/s10750-005-5195-x
Callaway R, Grenfell S, Lønborg C (2014) Small estuaries: ecology, environmental drivers and management challenges. Estuar Coast Shelf Sci 150(5):193–195. https://doi.org/10.1016/j.ecss.2014.06.009
Canani LGC, Menezes M, Torgan LC (2011) Diatomáceas epilíticas de águas oligotrófi cas e ácidas do Sudeste do Brasil. Acta Botanica Brasilica 25(1):130–140. https://doi.org/10.1590/S0102-33062011000100016
Carmona PA, Pereira LCC, Pinheiro SCC, Monteiro VF, da Costa RM (2010) Variação espaço-temporal do Microfitoplâncton dos furos da Ostra e Grande (canais de maré do estuário do Caeté), Pará-Brasil. Revista Da Gestão Costeira Integrada 8:1–8
Clarke KR, Warwick RM (1994) Change in marine communities: an approach to statistical analysis and interpretation. Plymouth Marine Laboratory, Plymouth. Online publishing
Cloern JE, Foster SQ, Kleckner AE (2014) Phytoplankton primary production in the world’s estuarine-coastal ecosystems. Biogeosciences 11:2477–2501. https://doi.org/10.5194/bg-11-2477-2014
Costa KG da, Pereira LCC, Costa RM da (2008) Short and long-term temporal variation of the zooplankton in a tropical estuary (Amazon region, Brazil). Boletim do Museu Paraense Emílio Goeldi, Série Ciências Naturais 3(2):127–141
Costa RM, Leite NR, Pereira LCC (2009) Mesozooplankton of the Curucá estuary (Amazon coast, Brazil). J Coast Res 56:400–404
Costa VB, Sousa EB, Pinheiro SCC, Pereira LCC, da Costa RM (2011) Effects of a high energy coastal environment on the structure and dynamics of phytoplankton communities (Brazilian Amazon littoral). J Coast Res 64:354–358
Costa AKR, Pereira LCC, Costa RM, Monteiro MC, Flores-Montes M (2013a) Oceanographic processes in an Amazon estuary during an atypical rainy season. J Coast Res SI 65:1104–1109. https://doi.org/10.2112/SI65-187.1
Costa RM da, Matos JB, Pinto KST, Pereira LCC (2013b) Phytoplankton of a dynamic Amazon sandy beach. J Coast Res 65:1751–1756
Costa MMS, Pereira SMB, Silva-Cunha MGG, Arruda PC, Eskinazi-Leca E (2016) Community structure of epiphytic diatoms on seaweeds in Northeastern Brazil. Bot Mar 59(4):231–240. https://doi.org/10.1515/bot-2015-0014
Costa AKR, Pereira LCC, Jiménez JA, Oliveira ARG, Montes MJF, Costa RM (2022) Effects of extreme climatic events on the hydrological parameters of the estuarine waters of the Amazon coast. Estuaries Coasts 1:1–17. https://doi.org/10.1007/s12237-022-01056-y
Cupp EE (1943) Marine plankton diatoms of the west coast of North America. Bull Scripps Institut Oceanogr Univ Calif 5:1–238
Demaster DJ, Pope RH (1996) Nutrient dynamics in Amazon shelf waters: results from Amassed. Cont Shelf Res 16(3):263–289. https://doi.org/10.1016/0278-4343(95)00008-O
Dittmar T, Lara RJ (2001) Driving forces behind nutrient and organic matter dynamics in a mangrove tidal creek in North Brazil. Estuar Coast Shelf Sci 52(2):249–259. https://doi.org/10.1006/ecss.2000.0743
Domingues RB, Barbosa A, Galvão H (2008) Constraints on the use of phytoplankton as a biological quality element within the Water Framework Directive in Portuguese waters. Mar Pollut Bull 56(8):1389–1395. https://doi.org/10.1016/j.marpolbul.2008.05.006
Dorado S, Booe T, Steichen J, McInnes AS, Windham R, Shepard A, Lucchese AEB, Preischel H, Pinckney JL, Davis SE, Roelke DL, Quigg A (2015) Towards an understanding of the Interactions between Freshwater Inflows and Phytoplankton Communities in a Subtropical Estuary in the Gulf of Mexico. PLoS ONE 10(7):1–23. https://doi.org/10.1371/journal.pone.0130931
Eskinazi-Leça E, Koening ML, Silva-Cunha MGG (2004) Estrutura e dinâmica da comunidade fitoplanctônica. In: Eskinazi-Leça E, Newmann-Leitão S, Costa MF (Eds) Oceanografia um cenário tropical (1ª ed., Pp. 353–373). Edições Bagaço.
Eskinazi-Leça E, Silva Cunha MG, Santiago MF, Palmeira Borges GC, Cabral De Lima JM, Silva MH, Paula Lima J, Menezes M (2010) Bacillariophyceae. In: Forzza RC (Eds) Catálogo de Plantas e fungos do Brasil (1ª ed., Pp. 262–309). Instituto de Pesquisas Jardim Botânico do Rio de Janeiro.
Espinoza JC, Ronchail J, Guyot JL, Junquas C, Drapeau G, Martinez JM, Santini W, Vauchel P, Lavado W, Ordoñez J, Espinoza R (2012) From drought to flooding: understanding the abrupt 2010–11 hydrological annual cycle in the Amazonas River and tributaries. Environ Res Lett. https://doi.org/10.1088/1748-9326/7/2/024008
Feitosa FAN, Nascimento FCR, Costa KMP (1999) Distribuição espacial e temporal da biomassa fitoplanctônica relacionada com parâmetros hidrológicos na bacia do Pina (Recife–PE). Trop Oceanogr 27(2):1–13. https://doi.org/10.5914/tropocean.v27i2.2813
Feng M, McPhaden MJ, Xie S, Hafner J (2013) La Niña forces unprecedented Leeuwin Current warming in 2011. Sci Rep 3:1277. https://doi.org/10.1038/srep01277
Figueroa SN, Nobre CA (1990) Precipitations distribution over central and western tropical South America. Climanálise 5(6):36–45
Flores Montes MJ, Macêdo SJ, Koening ML (2002) N:Si: P Atomic Ratio in the Santa Cruz Channel, Itamaracá-PE (Northeast Brazil): a Nyctemeral Variation. Braz Arch Biol Technol 45(2):115–124. https://doi.org/10.1590/S1516-89132002000200002
Fu M, Wang Z, Li Y, Li R, Sun P, Wei X, Lin X, Guo J (2009) Phytoplankton biomass size structure and its regulation in the Southern Yellow Sea (China): seasonal variability. Cont Shelf Res 29(18):2178–2194. https://doi.org/10.1016/j.csr.2009.08.010
Gameiro C, Zwolinski J, Brotas V (2011) Light control on phytoplankton production in a shallow and turbid estuarine system. Hydrobiologia 669:249–263. https://doi.org/10.1007/s10750-011-0695-3
Gao Y, Sun L, Wu C, Chen Y, Xu H, Chen C, Lin G (2018) Inter-annual and seasonal variations of phytoplankton community and its relation to water pollution in Futian Mangrove of Shenzhen, China. Continental Shelf Res 166(15):138–147. https://doi.org/10.1016/j.csr.2018.07.010
Gardner LR, Kjerfve B, Petrecca DM (2006) Tidal fluxes of dissolved oxygen at the North Inlet-Winyah Bay National Estuarine Research Reserve. Estuar Coast Shelf Sci 67(3):450–460. https://doi.org/10.1016/j.ecss.2005.12.002
Garmendia M, Ángel B, Franco J, Revilla M (2013) Phytoplankton composition indicators for the assessment of eutrophication in marine waters: Present state and challenges within the European directives. Mar Pollut Bull 66(1–2):7–16. https://doi.org/10.1016/j.marpolbul.2012.10.005
Goes JI, Gomes HR, Chekalyuk AM, Carpenter EJ, Montoya JP, Coles VJ, Yager PL, Berelson WM, Capone DG, Foster RA, Steinberg DK, Subramaniam A, Hafez MA (2014) Influence of the Amazon River discharge on the biogeography of phytoplankton communities in the western tropical north Atlantic. Prog Oceanogr 120:29–40. https://doi.org/10.1016/j.pocean.2013.07.010
Gomes VJC, Freitas PTA, Asp NE (2013) Dynamics and seasonality of the middle sector of a macrotidal estuary. J Coast Res 65(2):1140–1145. https://doi.org/10.2112/SI65-193.1
Gorgues T, Menkes C, Slemons L, Aumont O, Dandonneau Y, Radenac MH, Alvain S, Moulin C (2010) Revisiting the La Niña 1998 phytoplankton blooms in the equatorial Pacific. Deep Sea Res Part I 57(4):567–576. https://doi.org/10.1016/j.dsr.2009.12.008
Grasshoff K, Ehrhardt M, Kremling K (1983) Methods of seawater analysis. Verlag Chemie, Germany
Grego CKS, Feitosa FAN, Horonato da Silva M, Flores Montes MJ (2004) Distribuição espacial e sazonal da clorofila a fitoplanctônica e hidrologia do estuário do rio Timbó (Paulista-PE). Trop Oceanogr 32(2):181–199. https://doi.org/10.5914/tropocean.v32i2.8025
Guiry MD, Guiry GM (2015) AlgaeBase. world-wide electronic publication, National University of Ireland, Galway. From http://www.algaebase.org. Accessed 10 Jan 2016
Hasle GR, Syvertsen EE (1997) Marine diatoms. In: Tomas CR (ed) Identifying marine diatoms and dinoflagellates. Academic Press, An Imprint of Elsevier, Printed in the United States of America, pp 5–385
Hoppenrath M, Elbrachter M, Drebes G (2009) Marine phytoplankton. Selected microphytoplankton species from the North Sea around Helgoland and Sylt. Ed. Schweizerbart’sche Verlagsbuchhandlung, Stuttgart.
INMET (2011) Instituto Nacional de Meteorologia. Normas climatológicas (1961–1990). Brasília, Distrito Federal. Retrieved 19 June 2018, from https://portal.inmet.gov.br/
INMET (2015) Instituto Nacional de Meteorologia. Normas climatológicas (1961–1990). Brasília, Distrito Federal. Retrieved 15 Aug 2015, from https://portal.inmet.gov.br/
INMET (2018) Instituto Nacional de Meteorologia. Normas climatológicas (1961–1990). Brasília, Distrito Federal. Retrieved 19 June 2018, from https://portal.inmet.gov.br/
Islabão CA, Mendes CRB, Detoni AMS, Odebrecht C (2017) Phytoplankton community structure in relation to hydrographic features along a coast-to-offshore transect on the SW Atlantic Continental Shelf. Cont Shelf Res 151(1):30–39. https://doi.org/10.1016/j.csr.2017.10.003
Karsten U, Schumann R, Jung SRI, Medlin L (2006) Temperature and light requirements for growth of two diatom species (Bacillariophyceae) isolated from an Arctic macroalga. Polar Biol 29:476–486. https://doi.org/10.1007/s00300-005-0078-1
Kooistra WHCF, Sarno D, Balzano S, Gu H, Andersen RA, Zingone A (2008) Global diversity and biogeography of Skeletonema species (Bacillariophyta). Protist 159(3):177–193. https://doi.org/10.1016/j.protis.2007.09.004
Lacerda SR, Koening ML, Neumann-Leitão S, Flores-Montes MJ (2004) Phytoplankton nyctemeral variation at a tropical river estuary (Itamaracá–Pernambuco-Brazil). Braz J Biol 64(1):81–94. https://doi.org/10.1590/S1519-69842004000100010
Lam-Hoai T, Guiral D, Rougier C (2006) Seasonal change of community structure and size spectra of zooplankton in the Kaw River estuary (French Guiana). Estuar Coast Shelf Sci 68(1–2):47–61. https://doi.org/10.1016/j.ecss.2006.01.009
Lancelot C, Muylaert K (2011) Trends in estuarine phytoplankton ecology. In: Wolanski E, McLusky D (Eds) Treatise on Estuarine and Coastal Science. Elsevier Inc., pp. 5–15.
Lara RJ, Dittmar T (1999) Nutrient dynamics in a mangrove creek (North Brazil) during the dry season. Mangrove Salt Marshes 3:185–195. https://doi.org/10.1023/A:1009903824243
Liu B, Swart HED (2018) Quantifying the effect of salinity stratification on phytoplankton density patterns in estuaries. Estuaries Coasts 41:453–470. https://doi.org/10.1007/s12237-017-0276-4
López-Abbate MCL, Molinero JC, Guinder VA, Perillo GM, Freije RH, Sommer U, Spetter C, Marcovecchio JE (2017) Time-varying environmental control of phytoplankton in a changing estuarine system. Sci Total Environ 609(31):1390–1400. https://doi.org/10.1016/j.scitotenv.2017.08.002
Losada APM, Feitosa FAN, Lins IC (2003) Variação sazonal e espacial da biomassa fitoplanctônica nos estuários dos rios Ilhetas e Mamucaba (Tamandaré-PE). Trop Oceanogr 31(1):1–29. https://doi.org/10.5914/tropocean.v31i1.4987
Macêdo SJ, Flores-Montes MJ, Lins IC (2000) Características Abióticas da Área. In: Barros HM, Eskinazileça E, Macêdo SJ, Lima T (Eds) Gerenciamento Participativo de Estuários e Manguezais (1 ed., Pp 7–25). Editora Universitária da UFPE.
Magalhães ALP, Costa RM, Liang TH, Pereira LCC, Ribeiro MJS (2006) Spatial and temporal distribution in density and biomass Pseudodiaptomus species (Copepoda: Calanoida) in the Caeté river estuary (Amazon North of Brazil). Braz J Biol 66(2):421–430. https://doi.org/10.1590/S1519-69842006000300006
Magalhães ALP, Bessa RSC, Pereira LCC, da Costa RM (2009) Variação temporal da composição, ocorrência e distribuição dos Copepoda (Crustacea) do estuário do Taperaçu, Pará, Brasil. Boletim Museu Paraense Emílio Goeldi, Ciências Naturais 4(2):133–148
Magalhães A, Nobre DSB, Bessa RSC, Pereira LCC, Costa RM (2013) Diel variation in the productivity of Acartia lilljeborgii and Acartia tonsa (Copepoda: Calanoida) in a tropical estuary (Taperaçu, Northern Brazil). J Coast Res 65(2):1164–1169. https://doi.org/10.2112/SI65-197.1
Magalhães A, Pereira LCC, da Costa RM (2015) Relationships between copepod community structure, rainfall regimes, and hydrological variables in a tropical mangrove estuary (Amazon coast, Brazil). Helgol Mar Res 69:123–136. https://doi.org/10.1007/s10152-014-0421-4
Marengo JA (1995) Interannual variability of deep convection over the tropical South American sector as deduced from ISCCP C2 data. Int J Climatol 15(9):995–1010. https://doi.org/10.1002/joc.3370150906
Marengo JA, Tomasella J, Alves LM, Soares WR, Rodriguez DA (2011) The drought of 2010 in the context of historical droughts in the Amazon region. Geophys Res Lett 38(12):1–5. https://doi.org/10.1029/2011GL047436
Marengo JA, Borma LS, Rodriguez DA, Pinho P, Soares WR, Alves LM (2013) Recent extremes of drought and flooding in Amazonia: vulnerabilities and human adaptation. Am J Clim Chang 2(2):87–96. https://doi.org/10.4236/ajcc.2013.22009
Marengo JA, Nobre CA, Tomasella J, Cardoso MF, Oyama MD (2015) Hydro-climatic and ecological behaviour of the drought of Amazonia in 2005. Philos Trans R Soc B 363:1773–1778. https://doi.org/10.1098/rstb.2007.0015
Masuda LSM, Moser GAO, Barrera-Alba JJ (2011) Variação temporal do fitoplâncton no canal estuarino de Santos (SP). Braz J Aquatic Sci Technol 15(1):79–93. https://doi.org/10.1421/bjast.v15n1.p79-93
Matos JB, Sodré DKL, Costa KG, Pereira LCC, Costa RM da (2011) Spatial and temporal variation in the composition and biomass of phytoplankton in an Amazonian estuary. J Coast Res 64:1525–1529
Matos JB, Silva NIS, Pereira LCC, Costa RM (2012) Caracterização quali-quantitativa do fitoplâncton da zona de arrebentação de uma praia amazônica. Acta Botanica Brasilica 26(4):979–990. https://doi.org/10.1590/S0102-33062012000400028
Matos JB, Cardoso EHN, Pereira LCC, Costa RM da (2013) Diatomáceas cêntricas da zona de arrebentação de uma Ilha Amazônica. Trop Oceanogr 41(1–2):54–66. https://doi.org/10.5914/tropocean.v41i1-2.5414
Matos JB, Oliveira ARG, Trindade WN, Leite NR, Koening ML, Pereira LCC, Costa RM da (2016) Phytoplankton Dynamics in Three Metropolitan Beaches of the Amazon Littoral (São Luís-Maranhão). J Coast Res 75(1):413–417. https://doi.org/10.2112/SI75-083.1
Monteiro SM, El-Robrini M, Alves ICC (2015) Dinâmica sazonal de nutrientes em Estuário Amazônico. Mercator 14(1):151–162. https://doi.org/10.4215/RM2015.1401.0010
Moraes BC, Costa JMN, Costa ACL, Costa MH (2005) Variação espacial e temporal da precipitação no estado do Pará. Acta Amazon 35(2):207–214. https://doi.org/10.1590/S0044-59672005000200010
Muylaert K, Sabbe K, Vyverman W (2009) Changes in phytoplankton diversity and community composition along a salinity gradient of the Schelde estuary (Belgium/ The Netherlands). Estuar Coast Shelf Sci 82(2):335–340. https://doi.org/10.1016/j.ecss.2009.01.024
Nittrouer CA, Demaster DJ (1996) The Amazon shelf setting: tropical, energetic, and influenced by a large river. Cont Shelf Res 16(5–6):553–574. https://doi.org/10.1016/0278-4343(95)00069-0
Oliveira ARG de, Odebrecht C, Pereira LCCP, Costa RM da (2021) Phytoplankton variation in an Amazon estuary with emphasis on the diatoms of the Order Eupodiscales. Ecohydrol Hydrobiol 22(1):55–74. https://doi.org/10.1016/j.ecohyd.2021.12.001
Paerl HW (2006) Assessing and managing nutrient-enhanced eutrophication in estuarine and coastal waters: interactive effects of human and climatic perturbations. Ecol Eng 26(1):40–54. https://doi.org/10.1016/j.ecoleng.2005.09.006
Paiva RS, Eskinazi-Leça E, Passavante JZ de O, Silva-Cunha M da GG, Melo NFC (2006) Considerações ecológicas sobre o fitoplâncton da Baía do Guajará e foz do rio Guamá, Pará, Brasil. Boletim do Museu Paraense Emilio Goeldi, Ciências Naturais 1(2):133-146
Palma MB, da Costa KG, Flores-Montes MJ, Pereira LCC, Costa RM da (2013) Zooplankton spatial and temporal distribution in an equatorial estuary (Amazon littoral, Brazil). J Coast Res 65(2):1182–1188. https://doi.org/10.2112/SI65-200.1
Pamplona FC, Paes ET, Nepomuceno A (2013) Nutrient fluctuations in the Quatipuru river: a macrotidal estuarine mangrove system in the Brazilian Amazonian basin. Estuar Coast Shelf Sci 133:273–284. https://doi.org/10.1016/j.ecss.2013.09.010
Pan CW, Chuang YL, Chou LS, Chen MH, Lin HJ (2016) Factors governing phytoplankton biomass and production in tropical estuaries of western Taiwan. Cont Shelf Res 118(15):88–99. https://doi.org/10.1016/j.csr.2016.02.015
Pannard A, Claquin P, Klein C, Roy BL, Véron B (2008) Short-term variability of the phytoplankton community in coastal ecosystem in response to physical and chemical conditions’ changes. Estuar Coast Shelf Sci 80(2):212–224. https://doi.org/10.1016/j.ecss.2008.08.008
Parsons TR, Strickland JDH (1963) Discussion of spectrophotometric determination of marine-plant pigments, with revised equations for ascertaining chlorophylls and carotenoids. J Mar Res 21:155–163
Pereira-Filho J, Schettini CAF, Rörig L, Siegle E (2001) Intratidal variation and net transport of dissolved inorganic nutrients, POC and chlorophyll a in the Camboriú River Estuary, Brazil. Estuar Coast Shelf Sci 53(2):249–257. https://doi.org/10.1006/ecss.2001.0782
Pereira LCC, Monteiro MC, Guimarães DO, Matos JB, Costa RM da (2010) Seasonal effects of wastewater to water quality of the Caeté River estuary, Brazilian Amazon. An Acad Bras Ciênc 82(2):467–478. https://doi.org/10.1590/S0001-37652010000200022
Pereira LCC, Oliveira SMO, Costa RM da, Costa KG, Vila-Concejo A (2013) What happens on an equatorial beach on the Amazon coast when La Niña occurs during the rainy season? Estuar Coast Shelf Sci 135:116–127. https://doi.org/10.1016/j.ecss.2013.07.017
Pereira LCC, Costa ÁKR, Costa RM da, Magalhães A, Flores-Montes MJ, Jiménez JA (2017) Influence of a drought event on hydrological characteristics of a small estuary on the Amazon Mangrove Coast. Estuaries Coasts 41:676–689. https://doi.org/10.1007/s12237-017-0310-6
Piehler MF, Twomey LJ, Hall NS, Paerl HW (2004) Impacts of inorganic nutrient enrichment on the phytoplankton community structure and function in Pamlico Sound, NC USA. Estuar Coast Shelf Sci 61(2):197–207. https://doi.org/10.1016/j.ecss.2004.05.001
Pielou EC (1977) Mathematical ecology. Wiley, New York
Pinckney JL, Quigg AS, Roelke DL (2017) Interannual and seasonal patterns of estuarine phytoplankton Diversity in Galveston Bay, Texas, USA. Estuaries Coasts 40:310–316. https://doi.org/10.1007/s12237-016-0135-8
Procopiak LK, Fernandes LF, Moreira Filho H (2006) Diatomáceas (Bacillariophyta) marinhas e estuarinas do Paraná, Sul do Brasil: lista de espécies com ênfase em espécies nocivas. Biota Neotrop 6(3):1–28. https://doi.org/10.1590/S1676-06032006000300013
Queiroz JBM, Oliveira ARG de, Brito EP, Fernandes FD dos S, Nunes ZMP, Koening ML, Pereira LCCP, Costa RM da (2022) Phytoplankton of the shipping sector of São Marcos Bay (Amazon Coast): a potential risk area for the establishment of non-indigenous species. Regional Stud Marine Sci. https://doi.org/10.1016/j.rsma.2021.102121
Ricklefs REA (2010) A Economia da Natureza. Guanabara Koogan.
Rodrigues LC, Train S, Bovo-Scomparin VM, Jati S, Borsalli CCJ, Marengoni E (2009) Interannual variability of phytoplankton in the main rivers of the Upper Paraná River floodplain, Brazil: influence of upstream reservoirs. Braz J Biol 69(2):501–516. https://doi.org/10.1590/S1519-69842009000300006
Ross R, Sims PA (1972) The fine structure of the frustules in centric diatoms: a suggested terminology. Brit Phycol J 7(2):139–163. https://doi.org/10.1080/00071617200650171
Ross R, Cox EJ, Karayeva NI, Mann DG, Paddock TBB, Simonsen R, Sims PA (1979) A suggested terminology for the siliceous components of the diatom cell. Nova Hedwig Beih 64:513–533
Round FE, Crawford RM, Mann DG (1990) The diatoms: biology and morphology of the genera. Cambrigde Universit Press.
Santana DS, Paiva RS, Pereira LCC, Costa RM (2010) Microphytoplankton of the Marapanim estuary (Pará, Northern Brazil). Trop Oceanogr 38(2):161–172. https://doi.org/10.5914/tropocean.v38i2.5168
Santos MLS, Muniz K (2010) Variação nictemeral de parâmetros abióticos e clorofila-a em uma estação oceanográfica na Plataforma Continental do Amazonas. Atlântica 32(2):237–246. https://doi.org/10.5088/atlântica.v32i2.2463
Santos MLS, Muniz K, Barros-Neto B, Araújo M (2008) Nutrient and phytoplankton biomass in the Amazon River shelf waters. An Acad Bras Ciênc 80(4):703–717. https://doi.org/10.1590/S0001-37652008000400011
Santos MLS, Eschirique SA, Pereira JAR, Melo NFAC, Palheta GDA, Souza RAL, Saraiva ALL (2012) Estudo Nictemeral de Parâmetros Físicos e Químicos na Plataforma Continental do Amazonas. Trop J Fisheries Aquatic Sci 12(1):25–33. https://doi.org/10.32519/tjfas.v12i1.874
Santos-Fernandes TL, Passavante JZO, Koening ML, Macedo SJ (1998) Fitoplâncton do estuário do rio Jaguaribe (Itamaracá, Pernambuco, Brasil): Biomassa. Trop Oceanogr 26(2):1–18. https://doi.org/10.5914/tropocean.v26i2.2761
Sarma VVSS, Gupta SNM, Babu PVR, Acharya T, Harikrishnachari N, Vishnuvardhan K, Rao NS, Reddy NPC, Sarma VV, Sadhuram Y, Murty TVR, Kumar MD (2009) Influence of river discharge on plankton metabolic rates in the tropical monsoon driven Godavari estuary, India. Estuar Coast Shelf Sci 85(4):515–524. https://doi.org/10.1016/j.ecss.2009.09.003
Sassi R, Kutner MBB (1982) Variação sazonal do fitoplâncton da região de Saco Ribeira (Lat. 2330’ S, Long. 4507’W), Ubatuba, Brasil. Boletim do Instituto Oceanográfico, 31(2), 29–42. https://doi.org/10.1590/S0373-55241982000200005
Sathicq MB, Gómez N, Bauer DE, Donadelli J (2017) Use of phytoplankton assemblages to assess the quality of coastal waters of a transitional ecosystem: Río de la Plata estuary. Cont Shelf Res 150(1):10–17. https://doi.org/10.1016/j.csr.2016.08.009
Shannon CE (1948) A mathematical theory of communication. Bell Syst Tech J 27(3):379–423
Silva JG da, Torgan LC, Cardoso LS (2010) Diatomáceas (Bacillariophyceae) em marismas no sul do Brasil. Acta Botanica Brasilica 24(4):935–947. https://doi.org/10.1590/S0102-33062010000400008
Sims PA, Williams DM, Ashworth M (2018) Examination of type specimens for the genera Odontella and Zygoceros (Bacillariophyceae) with evidence for the new family Odontellaceae and a description of three new genera. Phytotaxa 382(1):001–056. https://doi.org/10.11646/phytotaxa.382.1.1
Smayda TJ (1980) Phytoplankton species succession. University of California Press, Berkeley
Sousa EB, Costa VB, Pereira LCC, Costa RM da (2009) Variação temporal do fitoplâncton e dos parâmetros hidrológicos da zona de arrebentação da Ilha Canela (Bragança, PA, Brasil). Acta Botanica Brasilica 23(4):1084–1095. https://doi.org/10.1590/S0102-33062009000400018
Souza Filho PWM, Gonçalves FD, Beisl CH, Miranda FP, Almeida EF, Cunha ER (2005) Sistema de observação costeira e o papel dos sensores remotos no monitoramento da costa Norte Brasileira, Amazônia. Rev Bras Cartogr 57(2):79–86
Steindinger KA, Tangen K (1997) Dinoflagellates. In: Tomas CR (ed) Identifying marine diatoms and dinoflagellates. Academic Press, San Diego, pp 387–598
Strickland JDH, Parsons TRA (1972) The Practical handbook of seawater analysis. Fisheries Research Board of Canada.
Thompson PA, Bonham P, Thomson P, Rochester W, Doblin MA, Waite AM, Richardson A, Rousseaux CS (2015) Climate variability drives plankton community composition changes: the 2010–2011 El Niño to La Niña transition around Australia. J Plankton Res 37(5):1–19. https://doi.org/10.1093/plankt/fbv069
Throndsen J (1978) Preservation and storage: monographs on oceanographic methodology. In: Sournia A (ed) Phytoplankton manual. Norwich, UESCO, pp 69–74
Tundisi JG, Tundisi TM (1976) Produção orgânica em ecossistemas aquáticos. Ciência e Cultura 38(8):864–867
van der Molen JS, Perissinotto R (2011) Microalgal productivity in an estuarine lake during a drought cycle: the St Lucia Estuary, South Africa. Estuarine Coast Shelf Sci 92(1):1–9. https://doi.org/10.1016/j.ecss.2010.12.002
Wang L, Chen Q, Han R, Wang B, Tang X (2017) Responses of the phytoplankton community in the Yangtze River estuary and adjacent sea areas to the impoundment of the Three Gorges Reservoir. Annales De Limnologie Int J Limnol 53:1–10. https://doi.org/10.1051/limn/2016027
Funding
This research work was funded by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, 001, Antonio Rafael Gomes de Oliveira, Conselho Nacional de Desenvolvimento Científico e Tecnológico, 309491/2018, Luci Cajueiro Carneiro Pereira, and 306061/2011-2, 311782/2017-5, Rauquírio Marinho da Costa.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
de Oliveira, A.R.G., Queiroz, J.B.M., Pardal, E.C. et al. How does the phytoplankton community respond to the effects of La Niña and post-drought events in a tide-dominated Amazon estuary?. Aquat Sci 85, 9 (2023). https://doi.org/10.1007/s00027-022-00904-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00027-022-00904-0