
Abstract
Conventional agricultural practices have been associated with negative effects, such as reduction of soil fertility, pollution of surface and groundwater and loss of biodiversity and ecosystem services (ES). To mitigate these effects, while sustaining high levels of crop production, innovative land-use practices are necessary. A promising land-use approach are alley cropping systems (ACS) with short rotation coppices, which are agroforestry systems, that combine the cultivation of conventional agricultural crops with fast-growing trees to produce biomass for energy purposes at the same time on the same piece of land. In the presented study, the effects of trees planted in ACS on agricultural land in Central Europe on microclimate, on soil organic matter (SOM) and on the provision of ecosystem services (ES) were elaborated, based on a review of relevant literature and results of recent research projects. The outcomes suggest that, due to their structural complexity, ACS can be more efficient regarding main microclimatic factors than either crop or tree monocultural systems. As a main factor, wind protection by the hedgerows in ACS was identified. Other microclimatic factors, such as light, temperature or evaporation, were also clearly influenced by the presence of trees; however, occurring interactions were often complex, and cause-effect relations were difficult to ascertain. A further outcome is that planting trees on agricultural sites potentially increases soil organic carbon (SOC) stocks, which can be considered as a main indicator for soil humus and soil fertility. However, it became evident that, in addition to the depth dimension (30 cm sampling depth can be considered as insufficient), the dimension of time needs to be taken more strongly into account. The authors suggest a division of the lifetime of agricultural trees in an initial (SOC stocks may decrease), a transitional (stocks approach steady state; SOC distribution pattern in the soil may change) and a steady-state phase (no major changes in stocks or distribution patterns) when interpreting effects of trees on SOC. Subsequently, in the attempt to value the effects of ACS on crop productivity and soil, suitable and transferable methods for the assessment of ES were discussed. It was demonstrated that the provision of ES from ACS was higher than from conventional agriculture and that ACS can increase productivity while sustaining high levels of SOC. Summarizing, the results suggested that ACS – if designed and managed appropriately – may function as a practical and diverse tool to mitigate negative effects of agricultural production.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others

Keywords
- Agroforestry
- Alley cropping system
- Biomass production
- Ecosystem services
- Microclimate
- Soil organic carbon
1 Introduction
Driven by the necessity to sustainably adapt agricultural production to changes in weather regimes and the growing demand for renewable energy carriers, new land-use systems and land-use strategies are put to the test. In this regard, the concept of agroforestry is gaining attraction in the temperate zone in recent years (Rigueiro-Rodríguez et al. 2009a).
A main characteristic of agroforestry systems is the large area share of trees which are combined with feedstock or agriculture on the same site (Nair 1985). The advantage of such mixed cropping compared to monocropping systems is the creation of a variety of interactions between the trees and the crops, comprising aspects of the nutrient and water cycle , microclimate and biodiversity (Nair 1993). Tree-induced interactions may sustainably stabilize or improve yield (Kort 1988) and mitigate negative effects of agricultural production on the environment, such as wind damage and soil erosion (Böhm et al. 2014), pollution or eutrophication of soil and waterbodies (Osborne and Kovacic 1993) or relative poverty of species (Greef et al. 2012). Furthermore, due to a more extensive management of agroforestry systems (at least of the areas with trees), the use of agrochemicals , such as fertilizer or pesticides , and soil cultivation measures are reduced. In result, agroforestry potentially produces ecological and economic benefits from which nature, society and land user may benefit (Dupraz et al. 2005; Quinkenstein et al. 2009a; Tsonkova et al. 2012). However, the intensity and the specific characteristics of these interactions depend mainly on the design and management of the system as well as the local site and growth conditions (Nair 1993).
In the temperate region, traditional agroforestry concepts, which focus mainly on food production and, therefore, on the optimization of nutrient and water cycles (Steppler and Nair 1987), are supplemented by complementary requirements such as production of woody biomass as energy resource, carbon (C) sequestration or provision of ecosystem services (Hall et al. 1991; Rigueiro-Rodríguez et al. 2009b; Kumar and Nair 2011). In this context, the agroforestry variant alley cropping with short rotation coppices (ACS) became relevant in Central Europe (Grünewald et al. 2007). ACS are agroforestry systems in which several sets of single or multiple rows of trees are planted in parallel tree strips with variable space between them (Nair 1985). This design creates alleys within which agricultural crops are cultivated using conventional techniques (Fig. 10.1).

Alley cropping system for biomass production in Eastern Germany, looking north-south direction along tree rows of poplar and winter wheat in the alleys (Photo: Dirk Freese, 2014)
Within ACS, fast-growing tree species such as poplar (Populus spp.), black locust (Robinia pseudoacacia) or willow (Salix spp.) are used. The trees are planted as cuttings or saplings in high densities of typically 8,000–15,000 (and more) plants per hectare wood area in single- or double-row design. This planting pattern facilitates quick growth and easy harvesting with specialized machinery, typically, at intervals of 2–6 years (Fig. 10.2).

Top, mechanized planting of poplar cuttings in an ACS on agricultural land in Eastern Germany (Photo: Michael Kanzler, 2011); bottom, mechanized harvesting of short rotational black locust trees on a reclamation site of a lignite opencast mine in Eastern Germany (Photo: Ansgar Quinkenstein, 2011)
The utilized tree species can resprout during the growing season following harvest, which allows such systems to be run for 20–30 years without replanting the trees, before yield starts to decline. The harvested wood is chipped directly at the field and, usually, used for power and heat production by combustion.
During the last years of ACS research, the influence of these agroforestry systems on microclimatic conditions (protection from erosion, moisture availability), soil humus contents (soil fertility, C sequestration) and the potential of agroforestry systems to sustainably provide ecosystem services (soil fertility, sustainability, system productivity) emerged as important ACS-related research issues (Grünewald et al. 2007; Quinkenstein et al. 2009a; Tsonkova et al. 2012).
The following text provides an overview of results of selected growth experiments in ACS systems in Central Europe. The focus is put on the influence of ACS on microclimate, the influence on soil organic matter (SOM) and the potential of ACS to provide selected ecosystem services (ES). Relevant scientific literature is reviewed and complemented with results from ongoing field studies.
2 Microclimate
The agricultural hedgerow systems in ACS increase the complexity and structural diversity of agricultural landscapes and, thus, modify microclimatic conditions in the sheltered areas in several respects (Quinkenstein et al. 2009a). By means of appropriate species selection, design and management of the hedgerows, these effects can be optimized to favour agricultural crop production. Main microclimatic factors that may be influenced by ACS are wind speed, wind turbulences, light distribution (shading by the trees), temperature, precipitation distribution, evaporation/evapotranspiration and available soil moisture (Quinkenstein et al. 2009a; Tsonkova et al. 2012). The intensity of these interacting microclimatic effects depends mainly on the height, length, structure and density of the hedgerow, the spatial layout of ACS (e.g. orientation of the hedgerows according to prevailing wind direction), the applied management (e.g. rotation interval and harvesting strategy) and the utilized tree species (Ringler et al. 1997; Brandle et al. 2004). The large number of different interactions can be summarized in two major potential microclimatic influences of ACS in temperate Europe: influences on the wind speed and influences on the water regime at the site (Tsonkova et al. 2012).
2.1 Wind Speed
If properly designed, ACS may function as an agricultural system consisting of multiple windbreaks , due to their composition of several hedgerows. Windbreaks provide shade and shelter for the crops and thus have been long used as a management technique to produce a more beneficial microclimate for agricultural crop production compared to open field (Cleugh 1998; Nuberg 1998). Such structures alter the mean wind speed, wind direction and turbulence of the airflow and provide shelter for some distance downwind with the positive effects that soil erosion by wind (Kort 1988) and the hazard of crop losses by physical damages due to strong winds are reduced (Cleugh 1998; Nuberg 1998). Accordingly, planting windbreaks is a common measure to prevent soil erosion by wind (Sudmeyer and Scott 2002; Brandle et al. 2004), especially at intensively cultivated areas with light erodible soils, such as can be found in Eastern Germany (Grimm et al. 2002; Nordstrom and Hotta 2004).
A typical wind profile around a single hedgerow can be divided into the ‘quiet zone’, a triangular zone that extends between three and eight times the height of the hedgerow leeward of the trees, and the ‘mixing zone’, a turbulent layer of air above and downwind of the ‘quite zone’, which merges into an ‘equilibrium’ zone in a leeward distance to the hedgerow of more than ten times the height of the hedgerow, where the original wind profile is re-established (Cleugh 1998). The extension and shape of these zones mainly depend on the width, height, structure and permeability or density of the hedgerow, as well as its continuity and spatial orientation (Brandle et al. 2004).
Reports of several studies regarding the effectiveness of windbreaks in wind reduction have been discussed. Sudmeyer and Scott (2002) reported that if the windbreak was perpendicular to the wind direction, wind speed reductions of greater than 20% occurred over the whole growing season for up to 3–6 tree heights from the windbreaks. Brandle et al. (2004) investigated different types of windbreaks (regarding composition and density) and reported extensions of the wind protection zone of up to 30 times the height of the windbreak leeward until the wind speed regained 100% of open-field wind speed. The authors reported also of small wind protection zones windward of the hedgerows of up to three times the height of the hedgerows (Brandle et al. 2004). Regarding ACS, Böhm et al. (2014) investigated the wind speeds in two ACS in Brandenburg, Germany, over a period of 5 years (including a partial harvest of the trees) and 1 year, respectively. The authors reported that the wind speed in crop alleys was reduced significantly within the ACS. In the middle of 24-m-wide crop alleys, the measured decrease of the annual average wind speed was more than 50% compared to the wind speeds at open field (Böhm et al. 2014). The authors concluded that ACS can represent effective windbreaks despite their comparably low hedgerows compared to ‘conventional windbreaks’ consisting of multiple rows of trees and/or shrubs of different heights (Böhm et al. 2014).
2.2 Water Regime
The water regime in ACS is influenced by a variety of microclimatic factors, and thus the water availability in the system usually depends on a complex interaction of different processes. Key processes for the water regime in ACS are distribution of precipitation, radiation, temperature, evaporation and evapotranspiration and influences of the plants on available soil moisture.
2.2.1 Distribution of Precipitation
Do to their windbreak properties, hedgerows in ACS may alter the distribution of precipitation within ACS compared to open-field conditions (Brandle et al. 2004). Typically, depending on the height and the porosity of the hedgerow, the precipitation increases slightly in the area immediately adjacent to the hedgerow on both sides. A small reduction in rain may occur on the downwind side due to wind turbulences and interception losses due to the trees and on the upwind site as the hedgerow may function as a barrier that redirects the airflow over the hedgerow (Monteith et al. 1991; Brandle et al. 2004; Kanzler and Böhm 2015). Due to the rain interception by the trees, slightly higher soil moisture values may be measured under the trees due to increased stem flow or dripping from the canopy (Monteith et al. 1991; Brandle et al. 2004). In addition to that, during the winter season, hedgerows lead to a more equal distribution of snow and to a delayed melting of snow during the spring that can help to enlarge water resources of the soil throughout the year (Scholten 1988).
2.2.2 Temperature and Radiation
Lower wind velocities on the leeward side of ACS hedgerows lead to a reduction in turbulent mixing of air masses with different temperatures compared to the conditions on open field. In result, in the wind protection zone of the hedgerows, the air temperature may be several degrees higher than in the open field (Brandle et al. 2004). An opposite effect results from the shading by the trees, which is an additional influencing factor for the microclimatic conditions in ACS. Generally, shading reduces the available radiation for the plants which leads to a reduction of photosynthesis activity. Furthermore, average air temperature as well as temperature extremes is reduced, which leads to a reduction of heat-induced plant damages, as well as reduces rates of transpiration and water evaporation from the soils, at least near the hedgerows (Feldhake 2001; Brandle et al. 2004). On grassland , in addition to an increase of agricultural production, this effect may also be useful to provide shade-, rain- and water-shading zones for feedstock. However, due to the relatively low trees in ACS (depending on the rotation interval up to about 5–6 m) and wide crop alleys (depending on the system design), the effect of shading is smaller than in types of agroforestry systems in which the trees can develop large, overhanging canopies.
The described microclimatic relations regarding temperature and radiation are supported by results published by Kanzler and Böhm (2015). The authors investigated microclimatic effects in an ACS with poplar and black locust hedgerows managed in short rotations and crop alley widths of 24, 48 and 96 m, established in 2011 on a former agricultural site in Brandenburg, Germany, and in an ACS with black locust established in 2007 on a lignite-mining reclamation site, about 35 km distance to the first site. They reported for the two investigated ACS slightly lower temperatures (about 1% lower) in the ACS on the agricultural site but about 5% lower temperatures for the ACS on the reclamation site. However, they found a clear reduction of extreme temperatures in both ACS compared to the reference sites (Kanzler and Böhm 2015). At the same time, the authors report a reduction of solar radiation of about 11% in vicinity to the hedgerows (about 3 m distance), but they stress that no direct effect on crop yield could be identified (Kanzler and Böhm 2015).
2.2.3 Evaporation, Evapotranspiration and Air Humidity
A reduction of wind speed and of wind turbulences by tree rows is an important influencing factor on the evapotranspiration rates within ACS, what might be a relevant feature of agricultural trees especially in dryer regions (Blenk 1953). Air turbulences are a main mechanism for exchanging heat and water vapour between land surface and atmosphere and are more effective in the direction from the surface to the atmosphere (Cleugh 1998). Accordingly, temperature and air humidity in the ‘mixing zone’, according to Cleugh (1998), may be lower by day but higher by night than in the ‘quiet zone’, whereas evaporation shows the opposite behaviour (McNaughton 1988; Cleugh 1998). However, these cause-effect relationships are not always clearly identifiable, because growing plants may adapt to the microclimatic conditions in the different zones, e.g. by modifying stomatal conductivity or leaf sizes, by what the evapotranspiration near the hedgerows might even increase (Brenner 1991). Kanzler and Böhm (2015) reported for the investigated ACS on agricultural area (see above) an average 3% higher air humidity about 3 m leeward of the hedgerow and about 1% higher humidity in the middle of a 96-m-wide crop alley compared to a reference site. However, for the ACS in the mining reclamation area, they reported the opposite trend with lower air humidity within the ACS compared to the reference site at an open field (Kanzler and Böhm 2015).
2.2.4 Soil Moisture and Groundwater Recharge
Plant-available soil water is one of the key factors for agricultural crop production. Generally, soil water depends on climatic conditions (e.g. precipitation), soil properties (e.g. texture or humus content) and management factors (e.g. ploughing) but is also influenced by the microclimatic interactions outlined above. Due to higher transpiration rates of the trees compared to agricultural crops throughout the year (Quinkenstein et al. 2009b), usually, lower soil moisture values directly under the trees than under the crops are measured. Accordingly, Kanzler and Böhm (2015) reported for the investigated ACS on the reclamation site and for the ACS on agricultural land in average lower soil moisture values in the topsoil (0–20 cm) than for the reference sites. However, for the ACS on the agricultural site, they reported an average 6% higher soil moisture in 20 cm soil depth on the downwind side of the hedgerow compared to the reference site. They explained this finding with a possibly higher dew formation due to the trees in the ACS, which is located close to a river, but stated also that the high variability of soil texture at the plot could have influenced the measurement results (Kanzler and Böhm 2015). Similar results were reported for an experimental ACS with poplar, willow and black locust trees on reclamation areas of the lignite-mining pit ‘Jänschwalde’ in the same region. The authors reported that the soil moisture from 0 to 30 cm soil depth in the middle of the 18-m-wide field strips was about 3–5% higher than on a neighbouring conventional agricultural reference site (Quinkenstein et al. 2009b).
Controversially discussed is a possible influence of agricultural trees on the recharge capacity of agricultural landscapes on groundwater , which is – at least in densely populated areas like Germany – an important and potentially scarce resource for drinking water. Within a modelling study, Quinkenstein et al. (2009b) investigated the average annual groundwater recharge, considering several water regime parameters such as interception or evapotranspiration, under poplar and black locust short rotation stands for different weather regimes and site conditions, typical for Eastern Germany, and compared the results with recharge rates under conventional agricultural crops. In result, the authors reported highest seepage rates of 35–39% of the precipitation in open area for the agricultural crops and clearly lower seepage rates of 17–20% and 11–14% for black locust and poplar, respectively (Quinkenstein et al. 2009b). Although the modelled seepage rates may be subject to some restrictions (see Quinkenstein et al. (2009b) for details), considering ACS, the seepage rates can be expected to be more similar to the rates under agricultural crops due to the comparably smaller area shares of trees.
3 Soil Organic Matter (SOM)
3.1 Functions and Importance of SOM for Soil Fertility
Soil organic matter (SOM) is defined as the sum of all biologically derived organic material in the soil or on the soil surface excluding aboveground living plant parts (Baldock and Nelson 1999). Since the direct determination of SOM contents is methodically difficult, most scientific studies measure organic C (Corg) contents and apply, if necessary, conversion factors ranging from 1.72 to 2.0 to the results to obtain SOM contents (Baldock and Nelson 1999). These conversion factors are variable, because they depend mainly on the C content of dead organic matter which ranges around about 50% but varies in dependence of the composition of the organic material and increases with increasing grade of humification (Kuntze et al. 1994). To avoid these variances in most cases, only the Corg or, in relation to the soil, the soil organic carbon (SOC) contents are given.
SOM describes a very complex mixture of different organic compounds. According to Kuntze et al. (1994), the main fractions of SOM are dead organic material (about 85%), plant roots (about 10%) and edaphic biomass (about 5%) which, in turn, consist of a large variety of different chemical and biological components. Due to this chemical complexity, SOM interacts in various ways with soil processes and influences soil properties, which have been broadly divided into biological/biochemical, chemical and physical aspects (Baldock and Nelson 1999). The first group comprises the provision of organic energy sources for biochemical soil processes by SOM and its role as a source of macro- (N, P and S) and micronutrients as well as a large variety of different organic substances, influencing pools of plant-available nutrients in different ways (Baldock and Nelson 1999). Furthermore, sufficient quantities of SOM (stabilized in terms of quantity and quality) and associated nutrients can enhance the resilience of ecosystem against external stresses (e.g. functioning as buffers for in-excess nutrients) and sustain a healthy soil life and plant growth. Regarding chemical soil properties, SOM is an important sorbent for inorganic and organic substances in the soil solution due to a large reactive surface and a high and partly pH-depending charge capacity of the organic soil compounds (Blume et al. 2016). Therefore, SOM is an important controlling factor for the cation exchange capacity of soils and may enhance retention of important soil ions such as K+, Ca2+, Mg2+, NH4 + or NO3 −. These chemical properties of SOM also lead to a comparably high buffer capacity of SOM against acids and, therefore, support a sustainably stable soil pH (Baldock and Nelson 1999). Moreover, SOM alters the biodegradability activity and persistence of pesticides in soils and may mitigate the effect of toxic metals, as well as enhance the availability of important nutrients such as phosphorus due to chelation of ions (Baldock and Nelson 1999). Regarding physical properties, SOM stabilizes soil structure through formation of stable aggregates by creation of organo-mineral compounds. Due to its influence on soil structure and pore geometry (Baldock and Nelson 1999), its charge and the presence of hydrophobic and hydrophilic areas, SOM is an important determinant for the water storage capacity of soils – it can absorb up to 20 times its mass of water (Baldock and Nelson 1999). Because of these factors combined, SOM enhances the resilience of soil particles against erosion and reduces leaching processes, and due to its dark colour, it also influences soil temperature regime to a certain extent (Baldock and Nelson 1999; Blume et al. 2016). Summing up, SOM usually accounts only for a small proportion of soil mass but, nevertheless, is of special relevance regarding soil fertility and, as the globally most important storage form of biologically sequestered C, also of high relevance regarding the global C cycle. Cultivation strategies greatly influence SOM as plants and their residues are the main source of SOC.
3.2 Formation, Decay and Composition of SOM
The major source for Corg in natural systems is the vegetation. During photosynthesis , the plants assimilate C from atmospheric carbon dioxide (CO2) and use it to build up organic compounds. A certain proportion of this fixed C is respired directly by the plants (autotrophic respiration) . The remaining C compounds are incorporated in the living biomass and, with time, gradually transferred as aboveground or belowground litter, as exudates or in other forms to the litter layer or directly into the soil. There, dead organic biomass is decomposed and mineralized by the edaphon, the fixed C is respired (heterotrophic respiration) and released back to the atmosphere as CO2. The contained nutrients are released to the soil solution from where they can be recaptured by organisms and incorporated into living biomass again (Blume et al. 2016). The total amount of SOM in the soil is the result of a dynamic equilibrium between deposition rate of organic residues and the mineralization rate, which in turn depends on the microbial activity (Sollins et al. 1996; Post and Kwon 2000). Important factors supporting an increase of Corg accumulation in the soil are large amounts of organic input of sufficient quality, the relocation of organic materials into the deeper soils by direct input (roots) or bioturbation by soil organisms and decomposition conditions that favour the development of stable C compounds. The activity of soil microbes, which, usually, is highest in the topsoil, is mainly controlled by weather conditions (temperature, moisture), nutrient supply and level of soil aggregation, which influences the physical availability of organic material, water and air to the microbes (Schinner and Sonnleitner 1996; Sollins et al. 1996). Major factors that favour the formation of bigger and more stable soil aggregates are plant remnants, root exudates and soil fungi (Blume et al. 2016). In this regard, the formation of organo-mineral compounds in the intestines of earthworms is of special relevance, due to the large resilience of organo-mineral complexes against biological, chemical or physical degradation (Post and Kwon 2000; Blume et al. 2016). Furthermore, the SOM content of soils is usually closely related to soil texture (especially the clay content) and, therefore, is also depending on abiotic site conditions (Schimel et al. 1985). Generally, soils with finer textures can store more Corg than soils with coarser textures (Blume et al. 2016).
The highest concentrations of Corg are found in the litter layers and in the topsoil. Litter horizons have an organic proportion of up to 100%, and their Corg contents usually range from 40% to 45% (Blume et al. 2016), whereas higher values of up to 55% Corg in the litter layers of forest soils in Central Europe have also been reported (Zech et al. 1992). Typical contents of Corg in agricultural soils in Central Europe range between 1% and 2% and in forest soils between 0.5% and 10% (Wessolek et al. 2008). Accordingly, Corg contents of 0.75–2% for Ah horizons of forest and agricultural soils have been reported, while in grassland higher values of up to 15% can be found (Blume et al. 2016). In contrast, the lower soil horizons of most soil types show clearly lower Corg values of 0.1–1% (Blume et al. 2016). The total stocks of organic matter stored in German arable soils in the topsoil layer were estimated to range between 100 and 200 Mg ha−1 (Blume et al. 2016).
3.3 Influence of ACS on SOM
The formation and maintenance of site-related high quantities of SOM are of great importance for the development and conservation of the soil fertility of agricultural soils. Agroforestry systems integrate tree with crop and animal production systems and are management practices that conserve and potentially enlarge Corg stocks in the biomass and the soils more than pastures or agricultural crops (Schroeder 1993). This idea assumes that integrated multispecies systems are more efficient in their resource usage (space, nutrients, light, water), produce beneficial interactions which stimulate plant growth and provide larger amounts of aboveground and belowground litter and favourable conditions for soil life, decomposition and humus formation than mono-species systems (Nair et al. 2009; Tsonkova et al. 2012). An important controlling factor seems to be the reduction of soil cultivation activities under the trees due to the limited accessibility of the forested areas (e.g. only possible after removal of the trees), by which the period of soil rest is enlarged (Rehbein et al. 2013). This, in combination with modifications of microclimatic conditions (e.g. alterations of moisture and temperature) and the comparably large litter production by the trees, leading to the formation of a persistent litter layer, protects the soil from microclimatic stress factors, stimulates the formation of more stable macroaggregates and reduces turnover and losses of Corg (Quinkenstein et al. 2009a, 2017; Rehbein et al. 2013).
3.3.1 Litter Production of Agricultural Trees
The annually produced litter biomass , as important raw material for SOC formation, depends in quantity and quality on the land-use system design and management (planting density, rotation interval, fertilization), the cultivated species (trees to produce biomass or high-value timber) as well as local growth conditions (weather, soil properties). Over a longer period, the trees in ACS develop a larger root system and more aboveground biomass, producing larger amounts of litter than annual agricultural crops, and, similarly important, enable the formation of a persistent litter layer. Generally, deciduous trees produce more organic material for SOM formation than coniferous trees , which produce more material than grassland , which, in turn, produce more material than agricultural areas (Kuntze et al. 1994).
Schroeder (1993) conducted a literature review and estimated the average Corg storage in the aboveground biomass by agroforestry practices to be about 63 Mg C ha−1 (about 3.9 Mg C ha−1 year−1) in the temperate ecoregions assuming a cutting cycle for the trees of 30 years. Because the shoot biomass produced by fast-growing tree species cultivated in European ACS typically is removed from the site every 4–7 years, it is usually not available for SOM formation. For these systems, a directly measurable biomass input into the soil is the mass of leaf litter and litter layer. An investigation of 18 sites, distributed all over Germany, with 2-year-old clones of poplar and willow resulted in an average produced dry matter (DM) leaf biomass of about 2.5 Mg DM ha−1 (Heyn et al. 2011). For 3- and 4-year-old poplar and black locust trees in an ACS on an agricultural site in Eastern Germany, a comparable annual leaf fall of about 2.5 Mg DM ha−1 was measured (Mirck et al. 2015). For the same site, with 4- and 5-year-old trees, a litter layer with a dry matter mass of about 4.7 and 5 Mg DM ha−1 on the forested area was measured 1 year later (Kanzler and Böhm 2015). Petzold et al. (2010) investigated the aboveground and belowground biomass of a 10-year-old poplar plantation in Saxony (Eastern Germany) and reported values of about 4.46 Mg DM ha−1 for leaf biomass and about 0.43 Mg DM ha−1 litter consisting of twigs and branches.
For the dryer conditions on a reclamation site of an opencast lignite mine in the same region of Germany, a higher litter accumulation and a litter layer mass of up to 11 Mg DM ha−1 in an 8-year-old ACS with black locust were reported (Kanzler and Böhm 2015). As opposed to aboveground biomass, leaf fall and litter layer mass of agricultural trees, the belowground biomass and its turnover are comparably hard to quantify. Ślązak et al. (2013) investigated the root biomass under different land-use systems and estimated with 3–4 Mg DM ha−1 higher biomass stocks under 4-year-old poplar plantations in Germany than under neighbouring willow plantations, fallow land or grassland . For a 10-year-old poplar site in Saxony, Petzold et al. (2010) reported a belowground coarse root biomass of about 32.6 Mg DM ha−1, and Quinkenstein et al. (2012) measured a Corg stock in the living fine root biomass of between 0.25 and 1.51 Mg C ha−1 and in the coarse root biomass of between 0.48 and 11.51 Mg C ha−1 for 1- to 12-year-old black locust stands on reclamation sites in Eastern Germany.
The amount of organic material of the accumulated biomass in the aboveground or belowground plant parts or in the litter layer that is actually transferred to the SOC per time step depends on several factors (e.g. quantity, quality of the litter and edaphic activity) and is usually expressed in a turnover rate for the considered Corg fraction. These values are difficult to quantify, are highly variable and, therefore, often must be estimated (Quinkenstein and Jochheim 2015). For many applications, it is, however, more straightforward to measure the changes of SOC stocks directly.
3.3.2 Soil Organic Carbon in ACS
Many studies give evidence that the establishment of agroforestry trees in agricultural areas enhances the Corg accumulation in the soils. However, published values of the estimated Corg sequestration potentials of agroforestry systems (in the biomass as well as in the soils) vary substantially, due to differences in scope, in methodology (e.g. sampled soil depth) as well as in assessment strategy (Nair 2011). To make the results better comparable, the effect of establishment of trees within ACS on the SOC stocks can be broadly divided into three stages which overlap under natural conditions: initial phase, transitional phase and steady-state phase (Fig. 10.3).

Hypothetical influence of the establishment of trees on agricultural areas on the SOC stock with highlighted initial, transitional and steady-state phase, whereas a higher biomass productivity of the trees after the initial phase compared to the agricultural system is assumed
3.3.2.1 Initial Phase
A change of a land-use system (e.g. from conventional agriculture to ACS) always represents in the first place a disturbance of existing soil processes. Because of increased decomposition losses due to removal of the old vegetation and/or soil cultivation measures prior to tree planting, the soil might even initially lose Corg throughout the first 1–5 years, when the growth of the newly planted trees remains comparably low, depending on the biomass productivity of the newly established land-use system (Tsonkova et al. 2012). Regarding agricultural trees, this effect was described by Walle (2007) who investigated birch and poplar plantations in Belgium and found a Corg decrease of about 42% in a soil depth of 0–30 cm after 4 years of growth, compared to the amount of Corg present in the soil before planting the trees. Stetter and Makeschin (1997) investigated two sites in Germany with afforestations of poplar and willow and reported a slight decrease of SOC stocks during the initial phase. Similarly, Paul et al. (2002), who conducted an extensive review on SOC changes in the topsoil (up to 30 cm depth) after forest afforestation for more than 200 globally distributed sites, concluded that SOC decreased during the first 5 years of tree growth but then started to increase, so that after about 30 years, there was little difference or slightly more SOC than was present in the agricultural soil before afforestation. The initial decrease in total Corg stocks might, therefore, be related with a smaller biomass production of the young trees during the first years compared to conventional crops, while at the same time, the decomposition losses for the first years remain large (Paul et al. 2002).
3.3.2.2 Transitional Phase
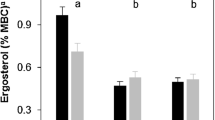
When the planted trees start to develop, the produced biomass and, therefore, the litter available for decomposition and humification processes in the soil increases for a period of several years. Moreover, in the tree rows, no ploughing takes place, which, typically, produces more or less homogeneous soil conditions and Corg contents under agricultural management throughout the ploughing horizon from about 0 to 30 cm. Because of enhanced mineralization of Corg, which mainly affects easily decomposable litter residues in the formerly ploughed horizons (Jug et al. 1999), and leaching losses originating from system establishment activities, a change in the distribution of Corg within the soil profile occurs. This leads to the formation of a depth gradient with higher Corg values in the uppermost and lower values in the deeper soil layers compared to the more homogeneously distributed Corg in agriculturally managed soils. The investigation of the soil depth distribution of Corg in a 6-year-old ACS established on an agricultural site in Eastern Germany demonstrated this change in SOC distribution induced by agricultural trees (Quinkenstein et al. 2017). Soil samples were taken in the middle of 48-m-wide crop alleys and in the middle of the framing 11-m-wide tree strips, consisting of four rows of poplar trees. The results for the Corg contents showed usual average values for agricultural sites within that region but also a trend of higher values in the topsoil and lower values in the deeper soil layers (Fig. 10.4).

Average soil contents of Corg (n = 3–4) in the middle of the agricultural site (above) and the tree strips (below) of a 6-year-old ACS in Eastern Germany (n = 3–4; black bars: median; boxes: lower and upper quartile; whisker: distribution maximum and minimum; points: outliers; Quinkenstein et al. 2017, modified)
The Corg contents over all plots ranged from 1.10 ± 0.28 (n = 8), in the uppermost soil layer (0–5 cm), to 0.24 ± 0.08 (n = 6) in the lowest soil layer (45–60 cm). Comparing the tree with the crop plots, differences between tree and crop plots for specific soil depths were not significant; however, the results indicated that under the trees, the decrease of Corg with soil depth was significant (p ≤ 0.05) – comparing the 0–5 cm and the 45–60 cm layer – and, as such, was more pronounced than in the soils under agricultural areas for which no significant difference was found for the same soil layers. The visual comparison of tree and crop plots (Fig. 10.4) supports this finding and reveals a trend of higher values in the topsoil under trees compared to the crop plots. Overall, the results indicated that the introduction of trees in the form of ACS into agricultural areas might initiate a change in the depth distribution gradient in soil regarding contents of Corg (Quinkenstein et al. 2017). A similar change of SOC distribution because of the planting of trees on agricultural lands was reported by Jug et al. (1999) who investigated changes in soil properties after afforestation of former agricultural and grassland areas with poplar and willow on three German sites. After a growing period of 7 and 10 years, respectively, they measured an increase of soil Corg in a depth of 0–10 cm, while a slight decrease of Corg for a soil depth between 10 and 30 cm occurred (Jug et al. 1999). Similar findings were reported for an afforestation of former agricultural areas with poplar and willow in Germany (Stetter and Makeschin 1997). The authors reported a slight increase of Corg in the uppermost and a slight decrease in the lowest investigated soil layer after 10 years of growth (Stetter and Makeschin 1997).
3.3.2.3 Steady-State Phase
Only when the Corg accumulation in the newly established land-use system surpasses the Corg loss via decomposition, additional Corg will be sequestered within the system (Schulze 2006), resulting in a higher steady-state SOC stock compared to the previous system. If ACS are established on agricultural land, a stable or increasing SOC content over longer time periods was reported by several studies. An investigation of the Corg contents from 0 to 30 cm in the soils under an ACS on an agricultural field (5-year-old at sampling) and an ACS on reclamation sites of a lignite opencast mining area in Eastern Germany (8-year-old at sampling) resulted in no significant changes in total Corg stocks for the first site, but a significant increase of about 1.48 Mg C ha−1 year−1 under the trees (compared to an increase of 0.68 Mg C ha−1 year−1 for the neighbouring agricultural areas) for the second site (Quinkenstein and Kanzler 2018). Medinski et al. (2014) compared agricultural areas with tree areas in three German ACS (between 1 and 4 years old) and reported in a soil depth of 0–3 cm a significant increase of Corg under the trees since planting.
If ACS are established on mining reclamation sites (with initially nearly no Corg in the soils), the C sequestration by the trees becomes even more evident and was pronounced also in deeper soil layers. Quinkenstein et al. (2011) investigated the SOC accumulation under four tree plantations of black locust established on lignite-mine reclamation areas in the Lusatian area in Eastern Germany, to a soil depth of up to 60 cm and time periods between 2 and 14 years of growth. The authors reported total Corg stocks of up to 106 Mg C ha−1 for the 14-year-old plantation and estimated an average accumulation rate of about 7 Mg C ha−1 year−1 for the considered soil depth following a pseudo-chronosequence approach. Furthermore, Nii-Annang et al. (2009) investigated an ACS established in another reclamation area in the same region and reported lower C accumulation rates in the topsoil (0–30 cm). The authors estimated a Corg accumulation of 3.4 mg C m−2 after 9 years of black locust cultivation, which corresponds to an annual accumulation rate of about 1.85 Mg C ha−1 year−1.
It is arguable whether the cited studies represent steady-state conditions within the considered ACS or if a steady state under agricultural conditions exists at all. Typically, for sites exposed to a constant land-use practice, time periods of up to 30–50 years are given until steady-state conditions for the Corg stock are reached (Robert 2001); however, even longer periods of more than 80+ years were reported (Jenkinson 1971). Due to the relative novelty of ACS, long-term studies on the development of SOC stocks are rare. Long-term studies from afforestations with forest trees could help to develop an idea of how the future development of SOC under ACS might look like. In general, from such studies, similar results were reported. For seven agricultural plots converted to tree plantations, Garten (2002) reported that SOC levels increased by 0.4–1.7 Mg Corg ha−1 (0–40 cm depth) within 10 years of establishment. Bambrick et al. (2010) investigated a 4-, an 8- and a 21-year-old tree-based intercropping site in Canada and found 77% and 21% higher SOC contents in 0–30 cm soil depth under the 8- and the 21-year-old trees, respectively, compared to nearby conventional agroecosystems, but no differences were detected at the youngest site. Post and Kwon (2000) conducted a comprehensive review of several studies on SOC sequestration after afforestation of agricultural sites and reported average values of 33.8 g C m−2 y−1 even after up to 250+ years since conversion.
Against this background, it needs to be considered that the average expected lifetime of ACS trees is only 20–30 years what (very likely) prevents that under the trees SOC-equilibrium is reached. However, if the whole ACS system is considered (including a ‘spatial rotation’ of tree strips), nearly steady-state conditions appear possible after an appropriate amount of time. In addition, a major issue, which makes the comparison of results of different studies regarding SOC challenging, is sampling depth. Most soil studies on agricultural sites are limited to the ‘ploughing horizon’, about 20–30 cm soil depth. Regarding tree-based systems such as ACS, this sampling depth can be considered as not sufficient, mostly, because tree roots in many cases extend to deeper soil horizons (see below). However, this issue has already been stated before (Nair 2011).
4 General Competition Effects
In agroforestry systems competition for resources such as light, water, space, nutrients between the trees and the adjacent crops is in many cases inevitable and needs to be managed to optimize the system’s output (Monteith et al. 1991). Studies have shown that yields close to agricultural trees may be reduced due to allelopathy (Singh et al. 1998), nutrient deficiency, shading, temperature effects or soil moisture deficiency (Kort 1988; Jose et al. 2000a). The degree of competition varies with the utilized species, system structure, management, general system location and soil and climate conditions. In ACS, the frequent cutting of the trees ensures that the trees usually don’t excess median heights of about 5–6 m (see above) and, accordingly, don’t develop large canopies. For this reason, competition effects are limited to the immediate vicinity to the trees, and aboveground competition factors (e.g. light access) might be less important than belowground factors such as competition for nutrients or moisture (Jose et al. 2000a, b). In this context, the lateral distribution of the roots is typically more important than the vertical distribution, whereas the development of a vertical stratified root system with the roots of annual crops in the upper and persisting roots of the trees in the lower soil layers is advantageous (Schroth 1998).
Many plants naturally tend to evade competition in the rooting zone by developing separate rooting systems (Schroth 1998). In the temperate region, the maximum rooting depth of annual crops usually ranges up to a few metres, whereas for the fast-growing tree species black locust (Robinia pseudoacacia) , poplar (Populus spp.) and willow (Salix spp.), rooting depths of >7.9, >3 and 4 m, respectively, were reported (Stone and Kalisz 1991). The maximal root system diameters were of 14.0, 30.5 and 40.0 m for trees of different ages (Stone and Kalisz 1991). Accordingly, investigations in agroforestry systems in Nebraska (USA) with apple trees (Malus sp.) and corn (Zea mays) showed that the roots of the apple trees grew up to 2 m deep into the soil before they started to spread out laterally to evade water competition with the roots of the corn plants growing in the upper soil layers (Yocum 1937).
It might be possible to support the formation of layered root systems and reduce root competition to some degree by cultivating species that tend to develop vertically stratified root systems (Young 1990) or by cultivating crops with compact root systems able to deflect tree roots into greater depths (Schroth 1995, 1998). Regular harvesting, pruning or cutting lateral tree roots extending into the crop field by, for example, repeated ploughing can force the trees to relocate major parts of their root system into the deeper soil layers (Schroth 1998). However, the effectiveness of such measures depends on the system design (e.g. planting density) and species characteristics (Schroth 1998; Hou et al. 2003).
5 Productivity of ACS
All the factors mentioned above (microclimate, SOM, competition) influence the wood and crop production potential of ACS to varying degrees. In addition, it depends on many factors such as system design, which is characterized by the utilized tree species, average tree height, strip width and planting density; management regime, which is characterized by planting and harvest technology, rotation length, fertilization, quality of pest and weed control; and other general site and growing conditions such as soil water and nutrient availability (Quinkenstein et al. 2009a; Tsonkova et al. 2012). While the crops in ACS, typically, are managed conventionally (including periodical fertilizer or pesticide application), the trees are managed extensively (e.g. fertilization and pesticide application are reduced because (a) the area is not accessible to machines for several years and (b) lower application rates are sufficient for most tree species), and their development is more strongly dependent on natural growing conditions.
Fast-growing tree species typically cultivated in Central European ACS, such as poplar, willow or black locust, produce a harvestable shoot biomass in dry matter (DM) between 8 and 14 Mg DM ha−1 year−1 (Lindroth and Båth 1999; Mitchell et al. 1999) on fertile agricultural sites. For tree plantations on average fertile soils in Eastern Germany, growth increments of about 6 and 12 Mg DM ha−1 year−1 for willow and poplar have been reported (Scholz et al. 2004; Bemmann et al. 2007). In Northern Poland, even higher growth rates for a selection of willow of between 14.2 and 21.7 Mg DM ha−1 year−1 were measured (Szczukowski et al. 2002), and Aylott et al. (2008) reported values for different poplar and willow clones of about 4.9–10.7 Mg DM ha−1 year−1 using data from a 49-site short rotation coppice (SRC) yield trial network in the UK. Lower growth values were reported for more nutrient-poor soils under dryer conditions such as can be found in the opencast lignite-mining region of Lower Lusatia in Germany. Accordingly, for plantations on reclamation sites within that region and for different rotation intervals, average growth increments of 0.5–2.9 Mg DM ha−1 year−1 for willow, 2.6–4.0 Mg DM ha−1 year−1 for poplar and 3.1–9.5 Mg DM ha−1 year−1 for black locust were reported (Bungart and Hüttl 2004; Grünewald et al. 2007, 2009; Quinkenstein et al. 2012).
Compared to either crop or tree monocultures , ACS are structurally and functionally more complex and, when properly managed, more efficient regarding major growing factors such as light, water or nutrients (Buck et al. 1999; Tsonkova et al. 2012). In addition, the introduction of trees into the agricultural area enhances the C sequestration potential of a landscape and, therefore, potentially increases humus stocks in the soil (Nair et al. 2009). Many studies show that the wind protection within windbreak systems has positive effects on crop yield over a range of climate and soil conditions (Baldwin 1988; Norton 1988). Typically, crop yields decrease near the hedgerows due to competition effects but increase in a zone further away from the hedgerows (Kowalchuk and de Jong 1995). Nuberg (1998) conducted a literature review on articles on windbreak effects on crop yields and reported for many different studies from all over the world that yield increases of up to 50% due to windbreaks compared to reference sites. Similar increases and top values of up to 70% more yield in systems with windbreaks compared to reference sites were reported in the review by Kort (1988). However, to what degree the crop production might benefit from wind protection effects depends on the specific characteristics of the considered system and the management. For example, harvesting of the hedgerows in ACS should be performed in a way that a constant wind protection is ensured. Accordingly, hedgerows should not be harvested completely at the same time. If the hedgerows consist of four or more tree rows, it is more useful to cut only a half of the tree rows at one harvest and harvest the remaining half in the following seasons (Quinkenstein et al. 2009a).
The economic value of mixed land-use systems can be assessed by using the land equivalent ratio (LER) . Using this ratio, the yields from growing two or more crops in a mixed stand and the yield obtained from growing the same crops in monoculture can be compared. LER is defined as the land amount equivalent needed by one type of crop to break even in physical terms (i.e. Mg ha−1) with what is yielded of 1 ha by another (Mead and Willey 1980; Grünewald et al. 2007). The LER is calculated according to Eq. (10.1):
where mixed denotes the yield of crop i in the mixed stand and mono denotes the yield of crop i in the monocultural stand. An LER greater than 1.0 usually shows that mixed cropping is advantageous, while less than 1.0 shows that it is disadvantageous. For an experimental ACS with black locust and alfalfa (Medicago sativa) established at an experimental plot on post-mining sites in the Lusatian region, an LER of 0.98 was reported by Grünewald et al. (2007), showing that under the local growth conditions, the productivity of alfalfa for an ACS and for monocultural cropping does not differ substantially (Grünewald et al. 2007). Other authors predicted an LER between 1.0 and 1.4 for silvo-arable systems within the European climatic region, suggesting that intercropping would be a more efficient land-use option than monocultural cropping (Graves et al. 2007).
To which actual extent the productivity of agricultural mixed systems differs from that of monocultural systems depends on the site management and abiotic site conditions but also on the specific characteristics of agricultural interactions at the considered site. In this context, microclimatic factors play a decisive role, as they influence important growth factors such as water availability to the crops.
6 Assessing Ecosystem Services Focusing on the Effects of Land Use on Productivity and Soil Fertility
The previously described complexity of ACS makes a comprehensive assessment of the inherent processes challenging. Establishing extensively managed strips of fast-growing trees on agricultural land leads to extension of the overall management and thus related enhanced provision of ES, which are also known as benefits provided to human population by the ecosystems (Costanza et al. 1997; Daily 1997; Reid et al. 2005). The concept of ES can be understood as an approach aiming to improve the understanding regarding benefits provided by nature, by describing the dependence of human well-being on these benefits and valuing them in economic terms (Marzelli et al. 2014). In this context, assessing ES is necessary to fully account for the environmental, economic and social effects of cultivation strategies and ultimately adequately reward the providers of ES. According to the Common International Classification of Ecosystem Services (CICES), they have been broadly classified into (i) provisioning (all nutritional, material and energetic outputs from living systems); (ii) regulation and maintenance (include mediation of waste, toxics and other nuisances, mediation of flows and maintenance of physical, chemical and biological conditions); and (iii) cultural (obtained from physical, spiritual and other interactions with biota, ecosystem and landscape) (Haines-Young and Potschin 2013).
The type, magnitude and relative mix of services provided by the ecosystems is influenced by management decisions which cause trade-offs between them, i.e. reduced provision of one ES because of increased provision of another service (Rodríguez et al. 2006). For example, agricultural ecosystems are primarily managed to optimize provisioning ES, but in the production process, they depend on many regulating ES, such as pollination , biological pest control, soil fertility, nutrient cycling and hydrological services (Zhang et al. 2007; Power 2010). Consequently, agricultural management which focuses on provisioning ES induces a trade-off with regulating and cultural services (Raudsepp-Hearne et al. 2010; Maes et al. 2012a). On the other hand, agriculture offers an opportunity of enhanced ES provision, among others, by application of land management regimes that favour the provision of ES (Porter et al. 2009). Hence, appropriate management strategies can ameliorate many of the negative impacts of agriculture, while at the same time largely maintaining provisioning services (Power 2010). According to the literature and recent research results, diverse farming systems , like agroforestry, were found better at providing ES than conventional agriculture (Jose 2009; Quinkenstein et al. 2009a; Kremen and Miles 2012; Smith et al. 2012; Tsonkova et al. 2012). In the attempt to optimize the effects of land-use strategies, such as agroforestry, a suitable approach is to assess the ES provided by this system.
The available tools for assessing ES provided by different land-use systems range from simple spreadsheet models to complex software packages (Bagstad et al. 2013). Information based on mapping and modelling exercises has been used to analyse the spatial distribution of multiple ES at various scales (Maes et al. 2012b). Typically, planning studies are carried out on subnational levels, while studies focusing on general trends like spatial distribution of ES are carried out on continental or global level (Maes et al. 2012b). For a regional or local assessment data, sources with higher spatial resolution should be used (Marzelli et al. 2014). Moreover, the influence of site conditions and management decisions should be more precisely reflected in the ES assessments. For example, multifunctional landscape models used to map and value ES often do not adequately include the impacts of land use and management on soil properties, due to model limitations and lack of ability to account for complexity and feedbacks in the systems (Dominati et al. 2016). Considerable differences in the provision of ES, as well as economic value under the same land use (permanent pasture grazed by dairy cows) for two contrasting soil types in New Zealand, were modelled by Dominati et al. (2016).
Moreover, provision of simple methods or data support tools to farmers can improve the adoption of appropriate land management approaches at the farm scale (Buckwell et al. 2014). Local decisions regarding benefits of ACS in comparison with conventional agriculture on a farm scale could be supported by simple tools, such as the ecosystem services assessment tool for agroforestry (ESAT-A), which linked indicators with ES, while taking into account the spatial heterogeneity of the field (Tsonkova et al. 2014). Essential for such an approach is using simple and well-acknowledged methods which provide reliable results without lacking scientific rigour. Data required as input in ESAT-A rely temporally on average yearly values and spatially on the farm field that would be designated to ACS and were related to descriptions of soil, climate as well as the field management (Tsonkova et al. 2014). The indicator results were linked to relevant regulating ES which enabled the assessment of several ES, supplied by ACS in comparison with conventional agriculture, conducted for hypothetical scenarios representing various site conditions of agricultural fields in Germany (Tsonkova et al. 2014). Moreover, the results could be also used as input to already establish software as it was demonstrated by Tsonkova et al. (2015), who identified target scenarios for establishing ACS by using the method of partial order ranking. The authors suggested that partial order ranking was a useful tool to objectively identify fields, where provision of ES could be enhanced by planting ACS, as it ranked the scenarios, while retaining their spatial configuration and information provided by the indicator set (Tsonkova et al. 2015). This study however did not include assessment of provisioning and cultural ES.
In the context of assessing ES provided by European agroforestry systems, a recent review suggested the focus was set on traditional agroforestry systems like wood pastures in the Mediterranean and hedgerow systems in the Atlantic and Continental regions (Fagerholm et al. 2016). Most of the studies assessed regulating as well as provisioning services , such as provision of habitat and biodiversity , food, climate regulation , fibre and fuel, while the consideration of cultural services has been largely limited to aesthetic value (Fagerholm et al. 2016). Torralba et al. (2016) conducted a meta-analysis on the effects of agroforestry on ES provision and biodiversity in Europe and concluded that compared to conventional agriculture, agroforestry can enhance the provision of ES and biodiversity. Erosion control, biodiversity and soil fertility were generally enhanced by the agroforestry systems considered, while the effect on provisioning services was not clear (Torralba et al. 2016). It was suggested that a negative effect on biomass production for silvopastoral agroforestry was possible (Torralba et al. 2016). The authors, however, did not consider ACS for production of biomass which is discussed in this study.
6.1 Productivity in ACS
The extensification of production via ACS did not lead to a reduction of the yield on a hectare basis, but rather the opposite was shown by several studies calculating LER, a useful indicator for assessing the productivity of agroforestry in comparison with agricultural practices (see above). Further studies assessing this indicator are necessary to optimize production from agroforestry under different conditions, considering the distance between hedgerows and value of LER obtained (Tsonkova et al. 2012). In general, the productivity in temperate agroforestry could be predicted by tools like Yield-SAFE, a model which can describe the yields of crops and trees in agricultural, agroforestry and forestry systems (van der Werf et al. 2007). Yield-SAFE has been recently enhanced in order to predict more accurately the provision of ES by agroforestry systems relative to forestry and arable systems (Palma et al. 2016). However, yield models usually require sets of input variables linked to inter alia soil, crop and climate characteristics which are not always known (Vooren et al. 2016) or cannot be easily obtained.
Therefore, a feasible approach of assessing yield of fast-growing trees, under consideration of different pedogenic and climatic conditions, is to use statistical data, average yields or regression models (Ali 2009; Röhle et al. 2010). Furthermore, the effect of tree rows on crop yield could be assessed relatively to the height of trees and the distance of the crop to the adjacent row as demonstrated by Vooren et al. (2016). The calculations of the authors considering the effect of tree height, based on results collected from temperate ACS, showed that compared to a treeless situation, relative crop yield was 70% starting from the tree row, over a distance of 1.64 times the tree height, and 107% between 1.64 and 9.52 times the tree height (Vooren et al. 2016).
A crucial factor in yield estimation in agroforestry systems is taking into account for the interactions between crop and tree components. The advantages of tree hedgerows to improving the yield of adjacent crops, because of microclimate modifications, have been broadly investigated in the literature (see above). In this context, the design of ACS regarding the height, orientation and width of tree rows is crucial to maximize the potential benefits and optimize the productivity of the system. The magnitude of benefits to the crop is in addition dependent on the type of crop planted, as their response to microclimate modifications varies (Kort 1988). The effect on the microclimate would be nevertheless captured in the yield and could be assessed through the overall system productivity. Furthermore, productivity is closely linked with soil fertility which can be measured by the effect of trees on soil nutrients and C.
6.2 Soil Fertility
6.2.1 Soil Nutrients
Agroforestry could contribute to soil fertility by promoting a more closed nutrient cycling than agricultural systems (Young 1990). The difference between the amount of all nutrient inputs entering the system and the quantity of nutrient outputs leaving the system produces the nutrient balance, an indicator of soil fertility (Roy et al. 2003; OECD 2008). A nutrient deficit suggests declining soil fertility, while a nutrient surplus suggests a risk of air and water pollution (OECD 2008). In intensively managed systems, nutrients which are exported from the system with harvest have to be yearly compensated for by using fertilizer inputs. For example, the export of nutrients for agricultural crops like silage maize is high, if the total aboveground biomass is utilized for energy production (Böhm et al. 2012). Increased fertilizer use in agriculture incurs financial costs, and excess application induces further loss of nutrients and thus related pollution of the environment (Baligar et al. 2001; Cassman et al. 2002).
On the other hand, perennial crops combine high biomass productivity with low requirements for fertilizer inputs (Böhmel 2007). The harvest of fast-growing trees usually takes place in the winter months, and the leaves, containing a high proportion of the absorbed nutrients by the tree, remain on the field (Böhm et al. 2012). They add to the soil nutrient pool, and the available nutrients can be once again taken up by the plants during growth. Thus, no external addition of nutrients is required, as the amount of nutrients exported is low. The lower nutrient export following harvest of fast-growing trees was also related with their higher nutrient use efficiency (Quinkenstein et al. 2009a). Nutrient use efficiency (NUE) emphasizing the internal nutrient requirement of the plant was defined as the total amount of harvestable biomass produced per unit of nutrient absorbed (Adegbidi et al. 2001) and was identified as a useful indicator for assessing soil fertility (Tsonkova et al. 2014). Maximizing biomass production using species with high NUE, hence lower nutrient removal at harvest, is desirable from the perspectives of both economics and environmental sustainability (Adegbidi et al. 2001). In the context of ACS, the overall NUE of the system is improved, as the NUE of the tree strips is higher; hence the nutrient export with harvest is lower than in conventional agriculture (Quinkenstein et al. 2009a). Thereby, the nutrient pool under the tree strips of the ACS is sustained in the long term.
Calculated nitrogen (N) balances for willow and poplar trees in the temperate region were shown to be positive, suggesting a build-up of organic N, while at the same time, low amounts of N were lost to the atmosphere and groundwater (Updegraff et al. 1990; Lamersdorf and Schulte-Bisping 2010; Pugesgaard et al. 2015). The former loss refers, for example, to emissions of nitrous oxide (N2O) which were generally estimated to be lower from perennial cultures as compared to annual crops (Hellebrand et al. 2010). The latter was related to leaching of nitrate (NO3) which was reduced in agroforestry as reported by several authors summarized among others by Tsonkova et al. (2012). For the assessment of nitrate leaching besides N balance, also determining seepage rate is necessary (Feldwisch et al. 1998). The method of Wessolek et al. (2004) which differentiates between forms of land use, groundwater influence and the plant-available water threshold is a useful approach in this regard. Moreover, assessing seepage rate is important as planting fast-growing trees has been associated with a reduction in water availability, especially where precipitation was low or even when dry summers occurred in areas with otherwise adequate precipitation (Dimitriou et al. 2009). This implied that in such cases for ACS, a trade-off could be expected between water regulation and water quality (Tsonkova et al. 2014).
Moreover, surface water quality can be seen as an additional improvement due to ACS as the export of nutrients with erosion and surface runoff was reduced (Tsonkova et al. 2012, 2014). In this context, the assessment of phosphorus loss is important which is largely determined by the phosphorus concentration in soil and the amount of soil lost with erosion (Feldwisch et al. 1998). In ACS the erosion by wind and water is minimized as trees act as a barrier and provide plant residues which also influence the amount of Corg in soil (see above).
6.3 Soil Carbon
The interaction between SOC and agricultural land management can be estimated by simple tools such as humus-balancing methods (Brock et al. 2013). These methods refer both to simple models aiming to quantify SOM change in arable soils, or SOC change in particular, and models that refer to the optimization of soil productivity in arable soils by calculating demand of organic fertilizer, without quantifying the change in SOC (Brock et al. 2013). A method belonging to the latter group developed by the Association of German Agricultural Analytic and Research Institutes ‘VDLUFA’ Verband Deutscher Landwirtschaftlicher Untersuchungs- und Forschungs Anstalten (Körschens et al. 2004) has proven especially suitable in the agricultural practice due to its user friendliness (Kolbe 2010). In this method, the organic matter supply and depletion effects were considered by using Corg decomposition coefficients which describe the maximum decreasing or increasing effects of a specific long-term cultivation method or supply of organic matter on the SOC stocks (Kolbe 2010). The cultivation of crops increasing SOC, e.g. grasses and legumes, as well as addition of various types of organic manure is assumed as an increase, and positive coefficients are used (Körschens et al. 2004; Kolbe 2010). The loss of SOC results from the cultivation of humus-depleting crop species, e.g. root and tuber crops, resulting from the provided low crop residues and is depicted by specific negative coefficients (Körschens et al. 2004; Kolbe 2010). In conventional agricultural management, a reduction of SOC is an indicator of reduced soil fertility and thus related productivity, while an increase of SOC may be related with increased mineralization, loss of N and reduced NUE (Körschens et al. 2004). Hence, the method aims at optimizing crop management by preserving the typical SOC content at the field.
Perennials on the other hand demonstrate strong positive effects on the humus balance due to provided litter and root residues and the lack of tillage (Hüttl and Dominik 2008). The positive effect of trees on SOC was described in detail in the previous section. Average values for the effect of fast-growing trees over a long-term period could be calculated from long-term experiments. The average annual increase in SOC stocks over 20 years of management calculated by Tsonkova et al. (2014) was of similar magnitude to the average annual increase in SOC stocks under forest soils, measured in Germany between the first and second National Forest Inventories in 1990 and 2006, respectively (Bolte et al. 2011). The increase in SOC under trees was especially important for soils of low quality, or soils with low initial SOC content (Coleman et al. 2004). Although typical concentrations of SOC vary for different soils, it was argued that a major threshold below which potentially serious decline in soil quality would occur was 2% SOC (Loveland and Webb 2003). Accordingly, in the assessment of Tsonkova et al. (2014), soils of low quality were particularly shown to benefit if an ACS was planted, as the highest increase in soil C was predicted for scenarios with lowest initial SOC content below 1%. Moreover, even for scenarios with sandy soil texture, generally most susceptible to leaching, the nitrate concentration in groundwater was significantly reduced in ACS as compared with conventional agriculture (Tsonkova et al. 2014). Furthermore, on low-quality soils, the cultivation of fast-growing trees did not compete with crop production and could result in the provision of additional benefits (Schulze et al. 2016). Hence, the provision of both market and non-market ES is enhanced under ACS; however, only for provisioning ES, there is an established market price, while for regulating and cultural ES, no market currently exists.
6.4 Market and Non-market ES
Valuation of ES from agroforestry systems suggested that the value of market ES was surpassed by the value of non-market ES. For example, the value of market and non-market ES provided by a mixed system with fast-growing trees (hazel, Corylus spp.; willow, Salix spp.; and alder, Alnus spp.) combining food and energy production in Taastrup, Denmark, was calculated by Porter et al. (2009). After taking into account the proportional areas of the pasture (45%), cereals (45%) and biomass (10%), the value of the non-market ES considered for the system comprised 64% of the total economic value (Porter et al. 2009). The largest contributions of non-market ES came from regulation of N turnover and a contribution to landscape aesthetics (Porter et al. 2009). Similarly, the monetary values of non-market ES provided by a tree-based intercropping system in Canada amounted to 65% of the total value of the ES considered (Alam et al. 2014). Of the ES which had no market price, water-quality regulation ranked highest, followed by air quality regulation and C sequestration. The study showed that although conventional agriculture provided more private benefits than agroforestry, the value of ES provided to society in agroforestry was much higher compared to this private value (Alam et al. 2014). A comparison regarding the provision of market and non-market ES by conventional agriculture and ACS is hypothetically illustrated in Fig. 10.5.

Compared provision of market and non-market ecosystem services by conventional agriculture (colour, brown) and alley cropping (colour, green). The value of market ES represents approximately one-third of the total ES provided by alley cropping (Porter et al. 2009; Alam et al. 2014). The crop yield in alley cropping may be lower, due to the area planted with trees, but in addition trees provide yield and shelter from the wind, maintain soil fertility and filter nutrients which otherwise would cause water pollution and improve biodiversity and system aesthetics
The provision of ES from ACS is much higher than ES from conventional agriculture, but only approximately one-third of this value is reflected in the market. Due to the increase in SOC by plant residues, soil fertility is maintained, and the losses of nutrients from the soil are minimized which results in a favourable N balance with no losses to the atmosphere and groundwater. Therefore, ACS enhance the provision of market services, i.e. crop and tree production, as well as the provision of non-market benefits, which is greatly influenced by the improvement in SOC and nutrient balance. In ACS, the provision of crop yield could be somewhat reduced as a small proportion of the land would be dedicated to planting trees. Nonetheless, in addition to obtaining tree yield, the provision of regulating and cultural services is enhanced; as in addition to maintaining soil fertility, trees provide shelter from the wind and improve water quality, biodiversity and system aesthetics (Tsonkova et al. 2012). However, planting trees requires somewhat higher initial investment, and the revenues are not yearly obtained as in the case of cultivating conventional crops. Therefore, efficient programs to support farmers are crucial for the establishment of agroforestry, thereby optimizing the effects of agricultural production. In this context, improving the financial support for agroforestry, e.g. by providing payments for provision of non-market benefits, is essential to promote this sustainable land-use system and enhance its relevance in the practice.
7 Conclusions
The discussed findings demonstrate that through site-adapted ACS systems, in many cases, negative effects of microclimatic stress factors for crop production, such as excessive wind or temperature extremes, can be mitigated. In addition, the discussed effects of ACS on SOM underline the important role that agricultural trees can play regarding the build-up and maintenance of substantial stocks of SOC in agricultural soils which is essential for sustaining and enhancing productivity of agricultural land in the long term. In this regard, a promising management strategy for the sustainable preservation of SOC seems to be the establishment of a rotation system, within which the cultivation of trees and conventional crops alternates on the same piece of land. Such a rotation would make soil areas, improved by trees regarding SOC, soil structure or soil life, available for agricultural crops, while conventionally used areas would be planted with trees in the subsequent rotation. In addition, the assessment of ES in ACS, focusing on site productivity and soil fertility, revealed that the provision of ES for which currently no market exists, typically, is higher than the provision of market goods. Hence, providing payments to farmers reflecting the total value of provided benefits is expected to enhance the practical relevance and application of ACS as a tool to optimize the effects of agricultural production.
Summarizing, the discussed characteristics of ACS suggest that the combination of agricultural trees and annual crops in ACS can function as a practical tool for improving the growing conditions for crops on agricultural sites regarding microclimatic conditions, SOC stocks and provided ES. However, the exact expression and characteristics of the discussed effects depend on the specific site, management, weather and other local growing conditions. In this regard, the overall advantage of ACS (and other agroforestry systems) is its flexibility, so that many relevant stress factors for agricultural production can be influenced and (to some degree) be modified in a way to mitigate negative effects and to sustain crop production. Furthermore, many potential disadvantages for crop production in ACS, which may arise due to competition effects between trees and crops (e.g. water stress in the rooting zone or shading by the trees), can be managed by appropriate measures (e.g. species selection, modifying planting density, root pruning by ploughing or regular cutting of the tree canopies). As complex multifunctional land-use systems, ACS are suitable to handle different usage scenarios and different objectives. If, for example, wind protection is the major issue, the crop alley widths in ACS can be optimized to maximize the wind protection effect. If woody biomass production needs to be increased, the area share of trees can be increased, the planting density and rotation length can be adjusted or, for example, if the protection of waterbodies is the main issue, the hedgerows can be aligned along riversides.
However, the interplay between management, plants, microclimate and soil in ACS is very complex and requires a careful planning of the system and land management. For this reason, more research is needed to better understand the interplay of identified interactions in ACS. An increased knowledge in this field could function as a basis to develop a more comprehensive support strategy for farmers who choose to establish ACS on their land. First steps could include the development of a subsidy system for the provision of non-market ES by ACS and the preparation of application-oriented guidelines of how to adapt ACS to different practical usage scenarios.
References
Adegbidi HG, Volk TA, White EH, Abrahamson LP, Briggs RD, Bickelhaupt DH (2001) Biomass and nutrient removal by willow clones in experimental bioenergy plantations in New York state. Biomass Bioenergy 20(6):399–411
Alam M, Olivier A, Paquette A, Dupras J, Revéret J-P, Messier C (2014) A general framework for the quantification and valuation of ecosystem services of tree-based intercropping systems. Agrofor Syst 88(4):679–691
Ali W (2009) Modelling of biomass production potential of poplar in short rotation plantations on agricultural lands of Saxony, Germany. PhD Thesis. Technical University Dresden, Dresden, Germany, p 130
Aylott MJ, Casella E, Tubby I, Street NR, Smith P, Taylor G (2008) Yield and spatial supply of bioenergy poplar and willow short-rotation coppice in the UK. New Phytol 178(2):358–370
Bagstad KJ, Semmens DJ, Waage S, Winthrop R (2013) A comparative assessment of decision-support tools for ecosystem services quantification and valuation. Ecosyst Serv 5:27–39
Baldock JA, Nelson PN (1999) Soil organic matter. In: Summer M (ed) Handbook of soil science. CRC Press, Boca Raton/London/New York, pp 25–84
Baldwin CS (1988) The influence of field windbreaks on vegetable and specialty crops. Agric Ecosyst Environ 22:191–203
Baligar VC, Fageria NK, He ZL (2001) Nutrient use efficiency in plants. Commun Soil Sci Plant Anal 32:921–950
Bambrick AD, Whalen JK, Bradley RL, Cogliastro A, Gordon AM, Olivier A, Thevathasan NV (2010) Spatial heterogeneity of soil organic carbon in tree-based intercropping systems in Quebec and Ontario, Canada. Agrofor Syst 79(3):343–353
Bemmann A, Feger K-H, Gerold D, Große W, Hartmann K-U, Petzold R, Röhle H, Schweinle J, Steinke C (2007) Kurzumtriebsplantagen auf landwirtschaftlichen Flächen in der Region Großenhain im Freistaat Sachsen. Forstarchiv 78:95–101
Blenk H (1953) Strömungstechnische Beiträge zum Windschutzproblem. Landtechnische Forschung 3(3):87–95
Blume H-P, Brümmer GW, Fleige H, Horn R, Kandeler E, Kögel-Knabner I, Kretzschmar R, Stahr K, Wilke B-M (2016) Scheffer/Schachtschabel: Soil Science. Springer, Berlin, p 618
Böhm C, Quinkenstein A, Freese D (2012) Vergleichende Betrachtung des Agrarholz- und Energiemaisanbaus aus Sicht des Bodenschutzes. Bodenschutz 2:36–43
Böhm C, Kanzler M, Freese D (2014) Wind speed reductions as influenced by woody hedgerows grown for biomass in short rotation alley cropping systems in Germany. Agrofor Syst 88:579–591
Böhmel C (2007) Comparative performance of annual and perennial energy cropping systems under different management regimes. PhD thesis. University of Hohenheim, Hohenheim, Germany, p 139
Bolte A, Wellbrock N, Dunger K (2011) Welche Maßnahmen sind umsetzbar? AFZ-Der Wald 2:27–29
Brandle J, Hodges L, Zhou X (2004) Windbreaks in North American agricultural systems. Agrofor Syst 61–62(1):65–78
Brenner AJ (1991) Tree-crop interactions within a Sahelian windbreak system. PhD thesis. University of Edinburgh, Edinburgh, UK, p 284
Brock C, Franko U, Oberholzer H-R, Kuka K, Leithold G, Kolbe H, Reinhold J (2013) Humus balancing in Central Europe – concepts, state of the art, and further challenges. J Plant Nutr Soil Sci 176(1):3–11
Buck LE, Lassoie JP, Fernandes EC (1999) Agroforestry in sustainable agricultural systems. CRC Press, Boca Raton/London/New York, p 432
Buckwell A, Uhre AN, Williams A, Polakova J, Blum WEH, Schiefer J, Lair GJ, Heissenhuber A, Schieβl P, Kramer C, Haber W (2014) The sustainable intensification of European agriculture. The RISE Foundation, Brussels, p 98
Bungart R, Hüttl RF (2004) Growth dynamics and biomass accumulation of 8-year-old hybrid poplar clones in a short-rotation plantation on a clayey-sandy mining substrate with respect to plant nutrition and water budget. Eur J For Res 123(2):105–115
Cassman KG, Dobermann A, Walters DT (2002) Agroecosystems, nitrogen-use efficiency, and nitrogen management. Ambio 31(2):132–140
Cleugh HA (1998) Effects of windbreaks on airflow, microclimates and crop yields. Agrofor Syst 41:55–84
Coleman MD, Isebrands JG, Tolsted DN, Tolbert VR (2004) Comparing soil carbon of short rotation poplar plantations with agricultural crops and woodlots in north Central United States. Environ Manag 33(1):299–308
Costanza R, d’Arge R, de Groot R, Farber S, Grasso M, Hannon B, Limburg K, Naeem S, O’Neill RV, Paruelo J, Raskin RG, Sutton P, van den Belt M (1997) The value of the world’s ecosystem services and natural capital. Nature 387:253–260
Daily GC (ed) (1997) Nature’s services: societal dependence on natural ecosystems. Island Press, Washington, DC, p 412
Dimitriou I, Busch G, Jacobs S, Schmidt-Walter P, Lamersdorf N (2009) A review of the impacts of short rotation coppice cultivation on water issues. Agric For Res 59(3):197–206
Dominati EJ, Mackay AD, Green JBS (2016) An ecosystems approach to quantify soil performance for multiple outcomes: the future of land evaluation? Soil Sci Soc Am J 80:438–449
Dupraz C, Burgess P, Gavaland A, Graves A, Herzog F, Incoll LD, Jackson N, Keesman K, Lawson G, Lecomte I, Liagre F, Mantzanas K, Mayus M, Moreno G, Palma J, Papanastasis V, Paris P, Pilbeam DJ, Reisner Y, van Noordwijk M, Vincent G, van der Werf W (2005) SAFE final report – synthesis of the silvoarable agroforestry for Europe project. European Union, INRA-UMR System editions, p 254
Fagerholm N, Torralba M, Burgess PJ, Plieninger T (2016) A systematic map of ecosystem services assessments around European agroforestry. Ecol Indic 62:47–65
Feldhake CM (2001) Microclimate of a natural pasture under planted Robinia pseudoacacia in central Appalachia, West Virginia. Agrofor Syst 53:297–303
Feldwisch N, Frede H, Hecker F (1998) Verfahren zum Abschätzen der Erosions und Auswaschungsgefahr. In: und S. Dabbert H-GF (eds) Handbuch zum Gewässerschutz in der Landwirtschaft. Ecomed, Landsberg, pp 22–57
Garten CTJ (2002) Soil carbon storage beneath recently established tree plantations in Tennessee and South Carolina, USA. Biomass Bioenergy 23:93–102
Graves A, Burgess P, Palma J, Herzog F, Moreno G, Bertomeu M, Dupraz C, Liagre F, Keesman K, van der Werf W, de Nooy AK, van den Briel J (2007) Development and application of bio-economic modelling to compare silvoarable, arable, and forestry systems in three European countries. Ecol Eng 29(4):434–449
Greef JM, Schwarz K-U, Hoffmann J, Langhof M, Lamerre J, Grünewald H, Pfennig K, von Wühlisch G, Schmidt C (2012) Ökonomische und ökologische Bewertung von Agroforstsystemen in der landwirtschaftlichen Praxis (Verbundvorhaben AgroForstEnergie, Abschlussbericht Teilvorhaben 3: Grünland- und Ackerflächen in Niedersachsen). Brandenburgische Technische Universität Cottbus, p 87
Grimm M, Jones R, Montanarella L (2002) Soil erosion risk in Europe. European Soil Bureau – Institute for Environment and Sustainability, JRC, Ispra, p 40
Grünewald H, Brandt BKV, Schneider BU, Bens O, Kendzia G, Hüttl RF (2007) Agroforestry systems for the production of woody biomass for energy transformation purposes. Ecol Eng 29(4):319–328
Grünewald H, Böhm C, Quinkenstein A, Grundmann P, Eberts J, von Wühlisch G (2009) Robinia pseudoacacia L.: a lesser known tree species for biomass production. Bioenergy Res 2(3):123–133
Haines-Young R, Potschin M (2013) Consultation on Version 4, August–December 2012 – Report to the European Environment Agency (Revised January 2013). Common International Classification of Ecosystem Services (CICES), EEA Framework Contract No. EEA/IEA/09/003, p 32
Hall DO, Mynick HE, Williams RH (1991) Cooling the greenhouse with bioenergy. Nature 353:11–12
Hellebrand HJ, Strähle M, Scholz V, Kern J (2010) Soil carbon, soil nitrate, and soil emissions of nitrous oxide during cultivation of energy crops. Nutr Cycl Agroecosyst 87(2):175–186
Heyn N, Jörgensen RG, Amthauer-Gallardo D, Wachendorf C (2011) Streufall und Streuumsatz in Böden von Kurzumtriebsplantagen. In: DBG (eds) Tagungsband der Jahrestagung der Deutschen Bodenkundlichen Gesellschaft (DBG) 2011: Böden verstehen – Böden nutzen – Böden fit machen, abgehalten vom 03. bis zum 09. September 2011, Berlin, Deutsche Bodenkundliche Gesellschaft (DBG), pp 1–4
Hou Q, Brandle J, Hubbard K, Schoeneberger M, Nieto C, Francis C (2003) Alteration of soil water content consequent to root-pruning at a windbreak/crop interface in Nebraska, USA. Agrofor Syst 57(2):137–147
Hüttl RF, Dominik P (2008) Anbau nachwachsender Rohstoffe: Auswirkungen auf die Humusentwicklung an den Produktionsstandorten. In: Hüttl RF, Gerwin W, Bens O (eds) Zum Stand der Humusversorgung der Böden in Deutschland. Brandenburgische Technische Universität, pp 207–213
Jenkinson DS (1971) The accumulation of organic matter in soil left uncultivated – Rothamsted Experimental Station report for 1970. Rothamsted Experimental Station, pp 113–137
Jose S (2009) Agroforestry for ecosystem services and environmental benefits: an overview. Agrofor Syst 76(1):1–10
Jose S, Gillespie AR, Seifert JR, Biehle DJ (2000a) Defining competition vectors in a temperate alley cropping system in the midwestern USA: 2. Competition for water. Agrofor Syst 48(1):41–59
Jose S, Gillespie AR, Seifert JR, Mengel DB, Pope PE (2000b) Defining competition vectors in a temperate alley cropping system in the midwestern USA: 3. Competition for nitrogen and litter decomposition dynamics. Agrofor Syst 48(1):61–77
Jug A, Makeschin F, Rehfuess KE, Hofmann-Schielle C (1999) Short-rotation plantations of balsam poplars, aspen and willows on former arable land in the Federal Republic of Germany. III. Soil ecological effects. For Ecol Manag 121(1–2):85–99
Kanzler M, Böhm C (2015) Nachhaltige Erzeugung von Energieholz in Agroforstsystemen (AgroForstEnergie II) – Abschlussbericht Teilvorhaben 2: Bodenschutz und Bodenfruchtbarkeit, Wasserhaushalt und Mikroklima. Brandenburgische Technische Universität Cottbus–Senftenberg, p 161
Kolbe H (2010) Site-adjusted organic matter–balance method for use in arable farming systems. J Plant Nutr Soil Sci 173(5):678–691
Körschens M, Rogasik J, Schulz E, Böning H, Eich D, Ellerbrock R, Franko U, Hülsbergen K-J, Köppen D, Kolbe H, Leithold G, Merbach I, Peschke H, Prystav W, Reinhold J, Zimmer J (2004) VDLUFA Standpunkt: Humusbilanzierung – Methode zur Beurteilung und Bemessung der Humusversorgung von Ackerland. Verband Untersuchungs- Deutscher und Forschungsanstalten Landwirtschaftlicher (VDLUFA), p 12
Kort J (1988) Benefits of windbreaks to field and forage crops. Agric Ecosyst Environ 22–23:165–190
Kowalchuk TE, de Jong E (1995) Shelterbelts and their effect on crop yield. Can J Soil Sci 75(4):543–550
Kremen C, Miles A (2012) Ecosystem services in biologically diversified versus conventional farming systems: benefits, externalities, and trade-offs. Ecol Soc 17(4)
Kumar BM, Nair PKR (eds) (2011) Carbon sequestration potential of agroforestry systems – opportunities and challenges. Springer, New York, p 530
Kuntze H, Roeschmann G, Schwerdtfeger G (1994) Bodenkunde – 5. neubearbeitete und erweiterte Auflage. Eugen Ulmer, Stuttgart, p 424
Lamersdorf N, Schulte-Bisping H (2010) Impact of short rotation forestry on soil ecological services. In: Proceedings of the 19th World Congress of soil science: soil solutions for a changing world held 1–6 August 2010 in Brisbane (Australia), pp 48–51
Lindroth A, Båth A (1999) Assessment of regional willow coppice yield in Sweden on basis of water availability. For Ecol Manag 121(1–2):57–65
Loveland P, Webb J (2003) Is there a critical level of organic matter in the agricultural soils of temperate regions: a review. Soil Tillage Res 70(1):1–18
Maes J, Egoh B, Willemen L, Liquete C, Vihervaara P, Schägner JP, Grizzetti B, Drakou EG, Notte AL, Zulian G, Bouraoui F, Paracchini ML, Braat L, Bidoglio G (2012a) Mapping ecosystem services for policy support and decision making in the European Union. Ecosyst Serv 1(1):31–39
Maes J, Paracchini ML, Zulian G, Dunbar MB, Alkemade R (2012b) Synergies and trade-offs between ecosystem service supply, biodiversity, and habitat conservation status in Europe. Biol Conserv 155:1–12
Marzelli S, Grêt-Regamey A, Moning C, Rabe S-E, Koellner T, Daube S (2014) Die Erfassung von Ökosystemleistungen – Erste Schritte für eine Nutzung des Konzepts auf nationaler Ebene für Deutschland. Nat Landsch 89:66–73
McNaughton K (1988) 1. Effects of windbreaks on turbulent transport and microclimate. Agric Ecosyst Environ 22:17–39
Mead R, Willey RW (1980) The concept of a ‘land equivalent ratio’ and advantages in yields from intercropping. Exp Agric 16(3):217–228
Medinski TV, Freese D, Böhm C, Slazak A (2014) Soil carbon fractions in short rotation poplar and black locust coppices, Germany. Agrofor Syst 88(3):505–515
Mirck J, Böhm C, Kanzler M, Freese D (2015) Blattstreumengen in Gehölzstreifen und angrenzenden Ackerbereichen innerhalb eines Agroforstsystems. In: Kage H, Sieling K, Francke-Weltmann L (eds) Multifunktionale Agrarlandschaften: Pflanzenbaulicher Anspruch, Biodiversität, Ökosystemdienstleistungen, Tagungsband der 58. Tagung der Gesellschaft für Pflanzenbauwissenschaften e. V., 22.–24. September 2015 in Braunschweig, pp 83–84
Mitchell CP, Stevens EA, Watters MP (1999) Short-rotation forestry – operations, productivity and costs based on experience gained in the UK. For Ecol Manag 121(1–2):123–136
Monteith JL, Ong CK, Corlett JE (1991) Microclimatic interactions in agroforestry systems. For Ecol Manag 45(1–4):31–44
Nair PKR (1985) Classification of agroforestry systems. Agrofor Syst 3(2):97–128
Nair PKR (1993) An introduction to agroforestry. Kluwer Academic Publishers, Dordrecht, p 499
Nair PKR (2011) Methodological challenges in estimating carbon sequestration potential of agroforestry systems. In: Kumar BM, Nair PKR (eds) Carbon sequestration potential of agroforestry systems – opportunities and challenges. Springer, New York, pp 3–16
Nair PKR, Kumar BM, Nair DV (2009) Agroforestry as a strategy for carbon sequestration. J Plant Nutr Soil Sci 172(1):10–23
Nii-Annang S, Grünewald H, Freese D, Hüttl R, Dilly O (2009) Microbial activity, organic C accumulation and 13C abundance in soils under alley cropping systems after 9 years of recultivation of quaternary deposits. Biol Fertil Soils 45(5):531–538
Nordstrom KF, Hotta S (2004) Wind erosion from cropland in the USA: a review of problems, solutions and prospects. Geoderma 121(3–4):157–167
Norton RL (1988) Windbreaks: benefits to orchard and vineyard crops. Agric Ecosyst Environ 22:205–213
Nuberg I (1998) Effect of shelter on temperate crops: a review to define research for Australian conditions. Agrofor Syst 41:3–34
OECD (2008) Environmental performance of agriculture in OECD countries since 1990. Organisation for Economic Co-operation and Development (OECD), p 208
Osborne LL, Kovacic DA (1993) Riparian vegetated buffer strips in water-quality restoration and stream management. Freshw Biol 29(2):243–258
Palma JHN, Graves AR, Crous-Duran J, Upson M, Paulo JA, Oliveira TS, de Jalón SSG, Burgess PJ (2016) Yield-SAFE model improvements. Milestone report 29 (6.4) for EU FP7 research project: AGFORWARD 613520. AGFORWARD, p 30
Paul KI, Polglase PJ, Nyakuengama JG, Khanna PK (2002) Change in soil carbon following afforestation. For Ecol Manag 168:241–257
Petzold R, Schubert B, Feger K-H (2010) Biomasseproduktion, Nährstoffallokation und bodenökologische Veränderungen einer Pappel-Kurzumtriebsplantage in Sachsen (Deutschland). Die Bodenkultur 61(3):23–35
Porter J, Costanza R, Sandhu H, Sigsgaard L, Wratten S (2009) The value of producing food, energy and ecosystem services within an agro-ecosystem. Ambio 38(4):186–193
Post WM, Kwon KC (2000) Soil carbon sequestration and land-use change: processes and potential. Glob Chang Biol 6:317–327
Power AG (2010) Ecosystem services and agriculture: tradeoffs and synergies. Philos Trans R Soc Lond B 365:2959–2971
Pugesgaard S, Schelde K, Larsen SU, Lærke PE, Jørgensen U (2015) Comparing annual and perennial crops for bioenergy production – influence on nitrate leaching and energy balance. GCB Bioenerg 7(5):1136–1149
Quinkenstein A, Jochheim H (2015) Assessing the carbon sequestration potential of poplar and black locust short rotation coppices on mine reclamation sites in Eastern Germany – model development and application. J Environ Manag 168:53–66
Quinkenstein A, Kanzler M (2018) Wirkungen von Agrargehölzen auf den Bodenstoffhaushalt. In: Böhm C, Veste M (eds) Agrarholz – Schnellwachsende Bäume für die Energieholzgewinnung. Springer. (in press)
Quinkenstein A, Schultze B, Grünewald H, Wöllecke J, Schneider BU, Jochheim H, Hüttl RF (2009a) Landschaftsökologische Aspekte der Dendromasseproduktion – Analyse und Bewertung von Risiken und Vorteilswirkungen. In: Murach D, Knur L, Schultze M (eds) DENDROM - Zukunftsrohstoff Dendromasse: Systemische Analyse, Leitbilder und Szenarien für die nachhaltige energetische und stoffliche Verwertung von Dendromasse aus Wald- und Agrarholz. Verlag Kessel, pp 317–344
Quinkenstein A, Wöllecke J, Böhm C, Grünewald H, Freese D, Schneider BU, Hüttl RF (2009b) Ecological benefits of the alley cropping agroforestry system in sensitive regions of Europe. Environ Sci Pol 12:1112–1121
Quinkenstein A, Böhm C, Matos E, Freese D, Hüttl RF (2011) Assessing the carbon sequestration in short rotation coppice systems of Robinia pseudoacacia L. on marginal sites in NE-Germany. In: Kumar BM, Nair PKR (eds) Carbon sequestration potential of agroforestry systems – opportunities and challenges. Springer, New York, pp 201–216
Quinkenstein A, Pape D, Freese D, Schneider BU, Hüttl RF (2012) Biomass, carbon and nitrogen distribution in living woody plant parts of Robinia pseudoacacia L. growing on reclamation sites in the mining region of lower Lusatia (Northeast Germany). Int J For Res 2012:1–10
Quinkenstein A, Janus T, Freese D (2017) Depth gradient of soil C, N and S contents in an alley cropping system for biomass production. In: Böhm C (ed) Bäume in der Land(wirt)schaft, von der Theorie in die Praxis – Tagunsgband des 5. Forums Agroforstsysteme, abgehalten vom 30.11.2016 bis zum 01.12.2016 in Senftenberg (Deutschland)
Raudsepp-Hearne C, Peterson GD, Bennett EM (2010) Ecosystem service bundles for analyzing tradeoffs in diverse landscapes. Proc Natl Acad Sci USA 107(11):5242–5247
Rehbein K, Sandhage-Hofmann A, Amelung W (2013) Langfristiger Kohlenstoffumsatz und C-Sequestrierung. In: Wagener F, Böhmer J, Kuhn D, Sutterer N (eds) Entwicklung extensiver Landnutzungskonzepte für die Produktion nachwachsender Rohstoffe als mögliche Ausgleichs- und Ersatzmaßnahmen (ELKE): Phase III – Umsetzung praxisbasierter Feldmodellprojekte (Endbericht), Hochschule Trier – Umwelt-Campus Birkenfeld, Institut für angewandtes Stoffstrommanagement (IfaS), pp 149–162
Reid WV, Mooney HA, Cropper A, Capistrano D, Carpenter SR, Chopra K, Dasgupta P, Dietz T, Duraiappah AK, Hassan R, Kasperson R, Leemans R, May RM, McMichael TA, Pingali P, Samper C, Scholes R, Watson RT, Zakri A, Shidong Z, Ash NJ, Bennett E, Kumar P, Lee MJ, Raudsepp-Hearne C, Simons H, Thonell J, Zurek MB (2005) Millennium ecosystem assessment: ecosystems and human well-being – synthesis. Island Press, Washington DC, p 155
Rigueiro-Rodríguez A, Fernández-Núñez E, González-Hernández P, McAdam J, Mosquera-Losada M (2009a) Agroforestry systems in Europe: productive, ecological and social perspectives. In: Rigueiro-Rodríguez A, McAdam J, Mosquera-Losada MR (eds) Agroforestry in Europe. Springer, Dordrecht, pp 43–65
Rigueiro-Rodríguez A, McAdam J, Mosquera-Losada MR (eds) (2009b) Agroforestry in Europe – current status and future prospects. Springer, Dordrecht, p 450
Ringler A, Roßmann D, Steidl I (1997) Landschaftspflegekonzept Bayern – Lebensraum Hecken und Feldgehölze. Bayrisches Staatsministerium für Landesentwicklung und Umweltfragen, Bayerische Akademie für Naturschutz und Landschaftspflege, München, Deutschland, p 523
Robert M (2001) Soil carbon sequestration for improved land management. FAO, Rome
Rodríguez JP, Beard TD Jr, Bennett EM, Cumming GS, Cork SJ, Agard J, Dobson AP, Peterson GD (2006) Trade-offs across space, time, and ecosystem services. Ecol Soc 11(1):1–28
Röhle H, Hartmann KU, Steinke C (2010) Ertragskunde. In: Skodawessely C, Pretzsch J, Bemmann A (eds) Eigenverlag Technische Universität Dresden, pp 53–59
Roy RN, Misra RV, Lesschen JP, Smaling EM (2003) Assessment of soil nutrient balance-approaches and methodologies FAO (Food Agricultural Organization of the United Nations), p 101
Schimel D, Coleman D, Horton K (1985) Soil organic matter dynamics in paired rangeland and cropland toposequences in North Dakota. Geoderma 36(3):201–214
Schinner F, Sonnleitner R (1996) Bodenökologie: Mikrobiologie und Bodenenzymatik Band I Grundlagen, Klima, Vegetation und Bodentyp. Springer, Berlin/Heidelberg/Germany, p 450
Scholten H (1988) Snow distribution on crop fields. Agric Ecosyst Environ 22–23:363–380
Scholz V, Hellebrand HJ, Grundmann P (2004) Produktion von nachwachsenden Energierohstoffen auf landwirtschaftlichen Flächen. KTBL, Kuratorium für Technik und Bauwesen in der Landwirtschaft e.V., pp 176–181
Schroeder P (1993) Agroforestry systems: integrated land use to store and conserve carbon. Clim Res 3(1–2):53–60
Schroth G (1995) Tree root characteristics as criteria for species selection and systems design in agroforestry. Agrofor Syst 30(1–2):125–143
Schroth G (1998) A review of belowground interactions in agroforestry, focussing on mechanisms and management options. Agrofor Syst 43(1–3):5–34
Schulze E-D (2006) Biological control of the terrestrial carbon sink. Biogeosciences 3(2):147–166
Schulze J, Frank K, Priess JA, Meyer MA (2016) Assessing regional-scale impacts of short rotation coppices on ecosystem services by modeling land-use decisions. PLoS One 11(4):1–21
Singh HP, Batish DR, Kohli RK (1998) Effect of poplar (Populus deltoides) shelterbelt on the growth and yield of wheat in Punjab, India. Agrofor Syst 40(2):207–213
Ślązak A, Böhm C, Veste M (2013) Kohlenstoffspeicherung, Nährstoff- und Wasserverfügbarkeit. In: Wagener F, Böhmer J, Kuhn D, Sutterer N (eds) Entwicklung extensiver Landnutzungskonzepte für die Produktion nachwachsender Rohstoffe als mögliche Ausgleichs- und Ersatzmaßnahmen (ELKE): Phase III – Umsetzung praxisbasierter Feldmodellprojekte (Endbericht), Hochschule Trier – Umwelt-Campus Birkenfeld, Institut für angewandtes Stoffstrommanagement (IfaS), pp 130–149
Smith J, Pearce BD, Wolfe MS (2012) Reconciling productivity with protection of the environment: is temperate agroforestry the answer? Renewable Agric Food Syst 28:80–92
Sollins P, Homann P, Caldwell BA (1996) Stabilization and destabilization of soil organic matter: mechanisms and controls. Geoderma 74(1):65–105
Steppler HA, Nair PR (eds) (1987) Agroforestry – a decade of development. International Council for Research in Agroforestry (ICRAF), Nairobi, p 336
Stetter U, Makeschin F (1997) Kohlenstoff- und Stickstoffdynamik vormals landwirtschaftlich genutzter Böden nach Erstaufforstung mit schnellwachsenden Baumarten. Mitteilgn Dtsch Bodenkundl Gesellsch 85(2):1047–1050
Stone E, Kalisz P (1991) On the maximum extent of tree roots. For Ecol Manag 46(1):59–102
Sudmeyer RA, Scott PR (2002) Characterisation of a windbreak system on the south coast of Western Australia. 1. Microclimate and wind erosion. Aust J Exp Agric 42(6):703–715
Szczukowski S, Tworkowski J, Klasa A, Stolarski M (2002) Productivity and chemical composition of wood tissues of short rotation willow coppice cultivated on arable land. Rostlinná Výroba 48(9):413–417
Torralba M, Fagerholm N, Burgess PJ, Moreno G, Plieninger T (2016) Do European agroforestry systems enhance biodiversity and ecosystem services? A meta-analysis. Agric Ecosyst Environ 230:150–161
Tsonkova P, Böhm C, Quinkenstein A, Freese D (2012) Ecological benefits provided by alley cropping systems for production of woody biomass in the temperate region: a review. Agrofor Syst 85:133–152
Tsonkova P, Quinkenstein A, Böhm C, Freese D, Schaller E (2014) Ecosystem services assessment tool for agroforestry (ESAT-A): an approach to assess selected ecosystem services provided by alley cropping systems. Ecol Indic 45:285–299
Tsonkova P, Böhm C, Quinkenstein A, Freese D (2015) Application of partial order ranking to identify enhancement potentials for the provision of selected ecosystem services by different land use strategies. Agric Syst 135:112–121
Updegraff KL, Zak DR, Grigal DF (1990) The nitrogen budget of a hybrid poplar plantation in Minnesota. Can J For Res 20(11):1818–1822
van der Werf W, Keesman K, Burgess P, Graves A, Pilbeam D, Incoll L, Metselaar K, Mayus M, Stappers R, van Keulen H, Palma J, Dupraz C (2007) Yield-SAFE: a parameter-sparse process-based dynamic model for predicting resource capture, growth and production in agroforestry systems. Ecol Eng 29(4):419–433
Vooren LV, Reubens B, Broekx S, Pardon P, Reheul D, van Winsen F, Verheyen K, Wauters E, Lauwers L (2016) Greening and producing: an economic assessment framework for integrating trees in cropping systems. Agric Syst 148:44–57
Walle IV (2007) Carbon sequestration in short-rotation forestry plantations and in Belgian forest ecosystems. PhD Thesis. Ghent University, Ghent, Belgium, p 244
Wessolek G, Duijnisveld WHM, Trinks S (2004) Ein neues Verfahren zur Berechnung der Sickerwasserrate aus dem Boden: das TUB-BGR-Verfahren. In: Bronstert A, Thieken A, Merz B, Rohde M, Menzel L (eds) Wasser- und Stofftransport in heterogenen Einzugsgebieten: Beiträge zum Tag der Hydrologie am 22./23. März 2004 in Potsdam (Germany), Hydrologische Wissenschaften – Fachgemeinschaft in der ATV-DVWKKleeberg, pp 135–145
Wessolek G, Kaupenjohann M, Dominik P, Ilg K, Schmitt A, Zeitz J, Gahre F, Schulz E, Ellerbrock R, Utermann J, Düwel O, Siebner C (2008) Ermittlung von Optimalgehalten an organischer Substanz landwirtschaftlich genutzter Böden nach § 17(2) Nr. 7 BBodSchG. Umweltbundesamt, p 211
Yocum WW (1937) Root development of young delicious apple trees as affected by soils and by cultural treatments. Univ Nebraska Agric Exp stat. Res Bull 95:1–55
Young A (1990) Agroforestry for soil conservation. CAB International, Wallingford, p 317
Zech W, Ziegler F, Kögel-Knabner I, Haumaier L (1992) Humic substances distribution and transformation in forest soils. Sci Total Environ 117:155–174
Zhang W, Ricketts TH, Kremen C, Carney K, Swinton SM (2007) Ecosystem services and dis-services to agriculture. Ecol Econ 64:253–260
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer Nature Singapore Pte Ltd.
About this chapter

Cite this chapter
Quinkenstein, A., Tsonkova, P., Freese, D. (2017). Alley Cropping with Short Rotation Coppices in the Temperate Region: A Land-use Strategy for Optimizing Microclimate, Soil Organic Carbon and Ecosystem Service Provision of Agricultural Landscapes. In: Dagar, J., Tewari, V. (eds) Agroforestry. Springer, Singapore. https://doi.org/10.1007/978-981-10-7650-3_10
Download citation
DOI: https://doi.org/10.1007/978-981-10-7650-3_10
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-10-7649-7
Online ISBN: 978-981-10-7650-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)