
Abstract
In percid fishes, the development of digestive structures and activities is quite similar to that of other carnivorous species (sea bass and sea bream). In most species at hatching, the digestive tract is a simple tube consisting of undifferentiated cells. The mouth and anus are closed and the esophagus is not connected with the intestine. Liver and pancreas are undifferentiated. Digestive enzymatic activities (pancreas, intestine) are detected shortly after hatching. According to histological and enzymatic studies, important changes occur around mouth opening (fifth to seventh dph). The pancreas shows exocrine activity and the liver becomes functional with adipogenic and glycogenic functions. The primary stomach develops in pikeperch and even earlier in Eurasian perch. Pancreatic (trypsin, amylase) and intestinal (leucine-alanine peptidase, Alkaline phosphatase, aminopeptidase N) enzyme activities increase at mouth opening. Leucine-alanine peptidase (cytosolic enzyme) activity declines after mouth opening concurrently with the strong increase of the brush border membrane enzymes (Alkaline phosphatase, leucine-aminopeptidase N) activity indicating the development of the brush border membrane of intestinal enterocytes. The stomach development occurs between 15th and 20th dph in pikeperch and between 21th and 35th in Eurasian perch. Pepsin activity is detected only on day 29 in pikeperch as well as in Eurasian perch larvae and is concurrent with the development of gastric glands. The development of brush border membrane of the intestinal enterocytes and the gastric activity indicate that larvae acquire an adult mode of digestion. The digestive structures and activities can be affected by the nature and the diet composition. This aspect is also discussed in this chapter allowing an approach of the nutritional requirements of percid larvae.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
1 Introduction
In the past, percids were commonly reared in fertilized ponds, as complementary carnivorous species in ponds mainly devoted to the production of cyprinids. In consequence, this fish generally fed on available zooplankton during larval stage. Protozoans, rotifers and young copepods are considered as important feed sources at this stage. Due to temperature and trophic conditions, suitable periods of larval production in ponds are, however, rather limited and usually restricted to spring. In order to secure mass production of juveniles all the year round and reduce the variability in the larval rearing success and juvenile yields obtained from pond system, more intensive percid larval culture has been developed during the last decades, first for North-American percid species (mainly walleye) and more recently for European ones (Eurasian perch and pikeperch) (see Chaps. 9 and 10 for more details). The growing of percid fish larvae in intensive indoor conditions have several advantages but, as feed availability fully depends on the quantity and type of feed supplied by the farmer, the feeding and nutritional requirements of the larvae must be well understood. Acceptance and efficiency of feed provided largely depend on the development of the digestive tract and associated organs (liver, pancreas, pyloric caeca, etc.).
Compared with most carnivorous fish, hatching size (total length = 5.4 mm, body weight = 0.7–0.8 mg) and mouth gape (0.36 mm) of percid fish like Eurasian perch or pikeperch are relatively small and thus constitute a limiting factor in initial feeding with live organisms (Kestemont et al. 1996). As many other fish species, percid larvae have a rudimentary digestive tract at first feeding and are not “equipped” with a functional stomach (Mani-Ponset et al. 1994; Vlavonou 1996; Ostaszewska 2005), which leads to a reduced ability to digest complex dietary proteins (Tonheim et al. 2004; Ronnestad et al. 2007). In many fish species including percids, the anatomy of the larval digestive system is quite different from that of juveniles and adults (Beccaria et al. 1991; Zambonino and Cahu 1994). Thus, a better knowledge of the nutrition physiology during early fish ontogeny is essential for the understanding of larval nutritional needs and for suggesting adequate feeding scheme.
2 Passage from Endotrophy to Exotrophy
At mouth opening (around 95˚ days), pikeperch larvae have a substantial vitelline vesicle under the anterior area of the gut. This vesicle includes the oil globule in front and the vitellus at the back (Mani-Ponset et al. 1994). These two parts, surrounded by periblast, are isolated from the digestive tract but connected with liver by vitelline veins. Yolk reserves cover the energetic expenses during the embryonic stage. At the start of exogenous feeding, most of the yolk sac is utilized. The resorption of vitellus begins on 2 days post-hatching (dph) in both Eurasian perch (Vlavonou 1996) and pikeperch (Hamza et al. 2007). Its total resorption occurs around 400˚ days and corresponds to the end of endo-exotrophic period.
Mani-Ponset et al. (1994) distinguished three phases in the post-embryonic development of pikeperch:
-
An endotrophic phase when the pre-larva lives on its yolk reserves (0–95˚ days);
-
An endo-exotrophic phase characterized by the depletion of yolk reserves and the beginning of feeding (95–400˚ days);
-
And an exotrophic phase when exogenous feeding is the only nutrient source (after 400˚ days)
3 Ontogeny of Digestive Organs
Development of the alimentary tract is similar in pikeperch (Sander lucioperca), perch (Perca fluviatilis), and streber (Zingel streber) (Mani-Ponset et al. 1994; Kestemont et al. 1996; Kováč 2000; Ostaszewska 2002). However, there are some differences concerning the time of differentiation of various structures, and thus, also the moment of functional ability in food utilization by the digestive tract.
In most species of percid fish at hatching, the digestive tract is a simple tube consisting of cells which are not differentiated. The mouth and anus is closed and the esophagus is not connected with the intestine (Fig. 8.1a, b).

(a) Longitudinal section of pikeperch head at hatching. The closed mouth (arrow). Scale bar 100 μm. (b) Longitudinal section of the posterior body part. Primary digestive tract (I) at hatching. Abdominal muscles (TM), anus closed (arrow). Scale bar 100 μm
The total body length of pikeperch at hatching is about 5–6 mm. The body of newly hatched pikeperch larvae is transparent and surrounded by a fin fold. The larvae display oval yolk sac divided into two sections: oil globule in the anterior part, and yolk in the posterior part. Both oil globule and yolk are enclosed within the yolk syncytial layer (YSL). Two syncytial zones are visible: one surrounding the oil globule, and the other surrounding the yolk. On the basis of ultrastructural YSL studies it was found that the spare substances of yolk sac are synthesized in periblast and overload in the circulatory (Mani-Ponset et al. 1994; Ostaszewska 2002). No blood vessel networks are observed on the yolk sac.
The mouth and pharynx of pikeperch are lined with cubic epithelial cells of irregular shape. Over the development time, these cells gradually flatten, and on the third day they transform into the multilayered squamous epithelium. The esophagus is undifferentiated. At hatching, the intestinal lumen is lined with irregular cubic cells that transform the second day into the unilayered cylindrical epithelium (Fig. 8.2a) (Ostaszewska 2005). On the first day post hatching the gut of Eurasian perch is extended and some folds are present (Kestemont et al. 1996). The liver and pancreas of pikeperch are located closed to the yolk sac, as undifferentiated cells (Fig. 8.2a, b). The undifferentiated liver and pancreas are separated from the yolk sac by syncytial layer. The cells of liver adjoin blood vessels and the syncytium (Fig. 8.2b) (Ostaszewska 2005). At hatching, the liver of Eurasian perch overhangs the anterior intestine while the pancreas is not distinguishable. The pancreas becomes visible under the stomach on day 2 post hatching (dph) (Kestemont et al. 1996).

(a) Longitudinal section of pikeperch of differentiated pancreas (P) cells are located over the yolk sac (Y). Intestine (I). (b) Longitudinal section of pikeperch of differentiated liver cells (L) adjacent to the blood vessels (BV) and syncytium (YSL) at 2 dph. Yolk sac (Y). Scale bars 10 μm
On the third day post hatching (TL = 5 mm) the intestine length and lumen of pikeperch increase, particularly in the anterior section. The intestine is lined with slightly folded unilayered mucosa of cylindrical epithelial cells, the nuclei of which is situated in the basal region (Fig. 8.3a). The hepatocytes with centrally located nucleus and distinct nucleolus increase, resulting in liver growth (Fig. 8.3b). At this stage, no glycogen storage is observed (Ostaszewska 2005).

(a) Longitudinal section of pikeperch of the primary intestine (I) lined with single layer of columnar epithelium of variable cell height at 3 dph. Scale bar 100 μm. (b) Longitudinal section of differentiated liver cells (L) at 3 dph. Scale bar 10 μm
In pikeperch considerable changes occur between the fifth and seventh dph (TL = 6 mm). The mouth opens, and so does the esophagus which connects the anterior intestine (Fig. 8.4). The primary stomach develops between esophagus and anterior intestine (Fig. 8.4) (Ostaszewska 2005). The “stomach section” is discernible because, contrary to the entire esophagus and intestine, it lacks mucous cells producing acidic mucus. They are absent from the future stomach region, and pyloric sphincter. The stomach of Eurasian perch starts differentiating earlier. At the second dph a large part of the stomach inner surface is smooth and bordered by a mucus layer (Kestemont et al. 1996).

Longitudinal section of pikeperch. Connection between the esophagus (E) and anterior intestine (I) at 5 dph. Liver (L), pancreas (P), primary stomach (S) oil globule (OG) mucous cells (*). Scale bar 100 μm
The mucous cells develop in the esophagus of pikeperch larvae after the mouth opening, simultaneously with the onset of a mixed endo-exogenous feeding (Fig. 8.4) (Ostaszewska 2005). At the same time mucous cells appear also in Solea senegalensis (Kaup) (Sarasquete et al. 1996), and in Solea solea (L.) (Boulhic and Gabaudan 1992), while they develop later in other species such as Sparus aurata L. and Scophthalmus maximus L. (Sarasquete et al. 1995). According to Gisbert et al. (1999), esophageal mucous cells develop 2 days before the beginning of active feeding in Acipenser baeri, similarly to Aspius aspius L. (Ostaszewska and Wegiel 2002). The mucous cells of pikeperch posterior esophagus secrete mainly acidic carbohydrate compounds, while those in the pharynx and anterior esophagus secrete neutral glycoproteins. The number and size of mucous cells increase with fish age. Similar pattern of mixed secretion (of acidic and neutral mucins) was observed in the larvae of Sparus aurata (Domeneghini et al. 1998), Acipenser baeri (Gisbert et al. 1999), and Melanogrammus aeglefinus (Hamlin et al. 2000). According to Boulhic and Gabaudan (1992), the esophagus of Solea solea larvae secretes exclusively acidic glycoproteins.
In pikeperch, further increase in liver volume takes place between the fifth and seventh dph, and hepatic blood vessels filled with blood cells become visible (Ostaszewska 2005). Glycogen storage is observed (Fig. 8.5a). Similarly, in Eurasian perch at the sixth day post hatching the vascularization of the liver is secured by numerous porte vein and centro-lobular vein. This indicates a liver development in which adipogenic and glycogenic functions are evidenced (Kestemont et al. 1996). Similar observations concerning the activity of this gland were reported by Boulhic and Gabaudan (1992) in Solea solea L. organogenesis. According to Diaz and Connes (1991), glycogen appears at the same time as hepatocytes differentiation, and its storage starts when the animal still relies on maternal nutrition (mammals) or yolk nutrients (birds, fish).

Longitudinal section of pikeperch of liver (L) and pancreas at 6 dph. (a) Blood vessels (arrows) among the hepatocytes. Glycogen PAS-positive areas. (b) Exocrine part of pancreas (EPA) and Langerhans islet (IPA). Proenzyme (Z). Scale bars 10 μm
The pancreas of pikeperch is situated above the liver, and shows exocrine activity. On the fifth day post hatching, first proenzyme granules appear in the basophilic pancreatic cells. Two days later, the number of granules considerably increases (Fig. 8.6) and a large Langerhans islet appears (Fig. 8.5b). At the beginning of endo-exogenous feeding, increased pancreatic activity indicates an important role of pancreatic secretory products before the stomach development (Ribeiro et al. 1999; Zambonino Infante and Cahu 2001).

(a) Longitudinal section of pikeperch of anterior intestine (I) at 7 dph. Connection (arrow) between the bile duct with intestine (I). Liver (L). (b) Intestinal valve (arrow) between the anterior (AI) and posterior intestine (PI) at 7 dph. Scale bars 10 μm
From the very beginning of mixed, endo-exogenous feeding (six to seven dph), a bile duct is observed in pikeperch, connecting the liver with the intestine. It opens into the anterior intestine section (Fig. 8.6a). Both, the gall bladder and the bile duct mucosa consist of unilayered cubic epithelium. The gall bladder is situated between the liver and pancreas.
From the beginning of mixed feeding (six to seven dph), the intestine of pikeperch is divided into two sections: anterior and posterior, separated by a valve (Fig. 8.6b). In Eurasian perch larvae, on the first dph, the intestinal valve appears between the median and posterior intestine (Kestemont et al. 1996). According to Pedersen and Hjelmeland (1988), the intestinal valve plays an important role at early larval stage, preventing enzyme escape from the intestine.
The yolk sac of pikeperch content considerably decreases between the 7th and 15th dph. The esophagus becomes longer and the secretory activity and number of mucous cells increase. The end of endogenous feeding of pikeperch larvae reared at 20 °C takes place at the same time as macroscopic yolk sac resorption, on day 6 post hatching. The remaining traces of yolk nutrients visible under the microscope are absorbed until day 15 (Ostaszewska 2005). At the same time the yolk of Eurasian perch reared at 21 °C almost disappears (Kestemont et al. 1996). Histological observations revealed that Petenia splendida larvae retained endogenous yolk reserves until 24 dph (Treviño et al. 2011).
In enterocyte cytoplasm of pikeperch, a small number of light vacuoles are observed on the ninth dph (Fig. 8.7a). In the anterior part, they disappear after 1–2 days, while, in the posterior part, the supranuclear regions contain vacuoles with acidophilic granules (Fig. 8.7b). These supranuclear vacuoles do not disappear before stomach development. Similar supranuclear vacuoles containing acidophilic granules visible in the cytoplasm of pikeperch posterior intestine were found also in larvae of other fish species: Gadus morhua; Dicentrarchus labrax; Pleuronectes ferrunginea; Pagrus pagrus (Dabrowski and Culver 1991; Kjørsvik et al. 1991; Deplano et al. 1991; Baglole et al. 1998; Darias et al. 2007). These results suggest that the vacuoles are the result of pinocytotic absorption of protein macromolecules from the intestine. This process is typical for fish larvae showing very low secretion of digestive enzymes, and lacking the gastric protease-producing glands (Govoni et al. 1986). Watanabe (1984) suggests that acidophilic granules found in this part of epithelium indicate active intracellular digestion necessary for protein assimilation, before the stomach development. The presence of supranuclear vacuoles indicates lipid storage (De Silva and Anderson 1995; Fontagné et al. 1998; Crespo et al. 2001). Lipids are hydrolyzed in the intestine to fatty acids and monoglycerides, and then absorbed. Then they are resynthesized in the smooth endoplasmic reticulum, and stored as fat droplets in enterocytes (De Silva and Anderson 1995).

Longitudinal section of pikeperch of anterior (a) and posterior (b) intestine. Small vacuoles with acidophilic granules (arrowhead) in the supranuclear regions of the enterocyte at 9 dph. Mucous cells (arrows). Scale bars 10 μm
The intestine of pikeperch grows as the fish ages: the number and size of mucosal folds as well as the length of brush border increase. In the epithelium of anterior and central intestine mucous cells appear, and their number increase with fish age. In the hindmost section of the intestine, a short anal channel develops, lined with cubic epithelium without mucous cells.
Over the entire endo-exogenous feeding period of pikeperch, the liver and pancreas gradually increase. At the seventh dph, lipid storage begins in the hepatocytes. Exocrine activity of pancreas significantly decreases from the nine to tenth dph, and lower number of proenzyme granules is observed. On the 13th dph, first tooth germs and taste buds appear in the mouth (Fig. 8.8).

Longitudinal section of buccopharynx of pikeperch. Buccopharyngeal epithelium with scattered mucous cells (arrowheads), taste buds (*) and teeth (arrow). Scale bar 100 μm
In pikeperch, the stomach development occurs between the 15th and 20th dph (TL = 8–15 mm), with the appearance of gastric glands (Fig. 8.9a) and pyloric sphincter (Fig. 8.9b). Also the intestine elongates and develops first loop (Fig. 8.9b). Around day 20 post-hatching the stomach is morphologically developed (Fig. 8.10).

Longitudinal section of pikeperch at 16 dph. (a) The beginning of gastric gland (GG) development. Scale bar 10 μm. (b) Stomach (S) developed from esophagus extension. Mucous cells (*) of esophagus (E). Liver (L), pyloric sphincter (PY), pancreas (P), swim bladder (SB), kidney (K). Scale bars 100 μm

Longitudinal section of pikeperch of digestive tract. Fully morphologically developed stomach at 30 dph. Liver (L), pancreas (P), intestine (I), pyloric caeca (PC), pyloric sphincter (PY), cardia stomach (CS) and blind sac (BS). Scale bar 100 μm
Pikeperch juvenile stomach is Y-shaped and consists of the cardia, pyloric stomach, and blind sac (Fig. 8.10). These parts are identified according to the terminology developed by Harder (1975), based on the differences in histological and histochemical properties. Fish species displaying the Y-shaped stomach with the blind sac belong, among others, to the Engraulidae, Gadidae, Clupeidae, and Percidae families (Harder 1975).
The mucosa of both, cardia and blind sac consist of unilayered cylindrical epithelium, mucosal lamina propria, and muscle layer. The mucosa of cardia and blind sac contain long, tubular and divided gastric glands situated in the lamina propria and surrounded by a loose connective tissue. The gland bottoms consist of polygonal secretory cells of round nuclei and cytoplasm containing acidophilic granules visible after staining with hematoxylin and eosin. The glands open to the mucosa fold crypts. The glands of the cardiac section are considerably longer, comparing to those in the blind sac (Fig. 8.10).
The appearance of gastric glands in pikeperch larvae occurs quite early during digestive tract ontogeny comparing with other fish species such as Solea solea, Pagrus pagrus (Boulhic and Gabaudan 1992; Darias et al. 2007) but later than in Acipenser baeri (Gisbert et al. 1999). They appears in both cardia and blind sac. In Paralabrax maculatofasciatus larvae they are found only in the anterior part of the stomach (Peña et al. 2003). However, the development of gastric glands is not necessarily accompanied by the onset of stomach activity. In Coregonus lavaretus larvae, pepsin activity and acidity were observed a long time after the development of gastric glands (Mähr et al. 1983). Similarity, pikeperch gastric glands contain eosinophilic particles indicating pepsinogen presence, thus a digestive activity of the stomach 10 days after the development of the gastric glands. The presence of neutral mucins in the gastric epithelium also suggests activity of gastric glands. Neutral mucins protect the epithelium against self-digestion by hydrochloric acid, and digestive enzymes produced by the gastric glands (Gisbert et al. 1999). Secretion of neutral mucins was observed in the cardia of adult Sparus aurata (Domeneghini et al. 1998) and larvae of Acipenser baeri and Paralichthys californicus (Gisbert et al. 1999, 2004).
Between the 20th and 30th dph (TL = 15–30 mm), mouth cavity and pharynx of juveniles are lined with the multilayered squamous epithelium, with numerous taste cells. Teeth appear in the upper and lower pharyngeal region. In the endmost section of esophagus, near the stomach, the multilayered squamous epithelium is replaced by multilayered cubic cells which are present also in the stomach wall adjacent to the esophagus. Among cubic cells, numerous mucous cells secrete acidic carboxyl and sulfate mucins.
The pyloric stomach mucosa consists of unilayered cylindrical epithelium, lamina propria and muscle layer. The long mucosal folds are developed as fanlike divisions (Fig. 8.10). The apical cytoplasm of cylindrical cells of cardia, pyloric stomach and blind sac are PAS-positive which indicates the presence of neutral compounds. The gastric glands contain eosinophilic particles indicating pepsinogen secretion between the 25th and 30th dph. The intestine, including the pyloric caeca, begins with the pyloric sphincter, and its anterior section is curved downwards. The pyloric caeca of Eurasian perch are observed at the same time as those of pikeperch (Kestemont et al. 1996). Craig (1987) reported that the genus Perca have three pyloric caeca. Pyloric caeca of pikeperch are different from the intestinal ones by higher amount of submucosa, and shorter folds (Fig. 8.10). Pyloric caeca lumen cross-section is starlike. Development of pyloric caeca is the last of a series of important morphological changes occurring in the digestive tract, suggesting that fish attained the juvenile stage (Bisbal and Bengtson 1995). According to Balon (1975), metamorphosis is completed and fish reach the juvenile stage when they show all fins distinct and well developed. This definition is based on external morphological features, and does not take into consideration internal changes, which do not always occur simultaneously. According to Stroband and Dabrowski (1981), the juvenile stage begins when the gastric glands develop, stomach displays digestive activity, and pyloric caeca appear.
In the older pikeperch, from day 11 post-hatching, mucous cells appear among the enterocytes of the anterior and posterior intestines. Similar cells were found by Domeneghini et al. (1998) in Sparus aurata L., Darias et al. (2007) in Pagrus pagrus and identified as basic secretory cells. They synthesize neutral and acid glycoconjugates. According to the same authors, carbohydrate compounds are the main component of intestinal mucus in vertebrates. Grau et al. (1992) reported that neutral mucous compounds of the intestine participate in enzymatic food digestion, formation of food mass, and absorption. In fish and in mammals, intestinal mucus plays an important protective role (Domeneghini et al. 1998). In pikeperch juveniles, after metamorphosis, mucous cells are very abundant in the entire intestine, but their number is different in various sections. They are the most numerous in the posterior intestine. According to Domeneghini et al. (1998), high density of mucous cells in the colon is essential for easy defecation. The mucus produced by the fish mucous cells plays the same role as in mammals (Scocco et al. 1998), protecting the mucosa of the digestive tract.
In pikeperch juveniles the U-shaped liver is centrally located in the peritoneal cavity, ventrally from the esophagus, and anterior-ventrally from the stomach (Fig. 8.11). The hepatocytes contain numerous light vacuoles indicating lipid storage, and dark PAS-positive glycogen granules. The gall bladder is situated between the liver and anterior intestine with which it is connected with the bile duct (Fig. 8.11).

Longitudinal section of pikeperch of esophagus (E), anterior stomach part (CS), and gall bladder (*) at 30 dph. Pancreas (P), intestine (I), liver (L). Scale bar 100 μm
As reported by Kestemont et al. (1996) and Ostaszewska (2005), the pancreas of Eurasian perch and pikeperch are dispersed within the mesentery of the anterior stomach section, among the pyloric caeca, and bile duct (Fig. 8.10). In pikeperch juvenile exocrine pancreatic tissue is present around the hepatic portal veins. The Langerhans islets are dispersed within entire gland. Similarly as in the other vertebrates, liver and pancreas of pikeperch are of endodermal origin. Hepatocytes proliferate very quickly during the development, and due to morphogenetic translocation, already in the fifth day of life surround the intestine.
The pikeperch digestive system is developed before the complete metamorphosis, since the alimentary tract is distinctly divided into esophagus, anterior and posterior intestine, and food particles are assimilated. Also liver and pancreas are active, and gall bladder store bile. There are no anatomical differences between larval and juvenile stages, except for the presence of gastric gland and pyloric caeca as well as intestine length. Thus, literature data (Ruuhijärvi et al. 1991; Schlumberger and Proteau 1991; Mani-Ponset et al. 1994; Ostaszewska 2002) suggest that despite such an advanced digestive system activity, feeding of pikeperch with artificial feeds is difficult before fish attain the juvenile stage.
4 Ontogeny of Digestive Enzymes
Available data on digestive enzymes at larval stage reported in this chapter concern essentially Eurasian perch and pikeperch. The ontogeny of digestive enzymes in these larvae have essentially been described by Kestemont et al. (1996), Cuvier-Péres and Kestemont (2002), and Hamza et al. (2007) and their pattern and values range are quite similar. The following description will focus essentially on pikeperch larvae. The onset of digestive functions, associated with morphological transformations, follows a sequential chronology in developing fish like that in developing mammals (Cahu and Zambonino Infante 2001). The digestive tract of fish larvae is not achieved at hatching but undergoes major developmental changes over several weeks (Cousin and Baudin Laurencin 1985; Boulhic and Gabaudan 1992). In percid fishes the development of digestive structures and activities seems similar to that of other carnivorous species. Indeed, according to Mani-Ponset et al. (1994), the development of the digestive tract of pikeperch seems chronologically similar to that of European sea bass Dicentrarchus labrax and sea bream Sparus aurata larvae.
Digestive enzyme activities, particularly pancreatic and intestinal ones, display a pattern characteristic of developing animals. Their activities are already detectable at hatching and are independent from exogenous feeding since the larvae do not feed before three (Eurasian perch) to five dph (pikeperch). Profiles of pancreatic and intestinal enzymes are depicted below for pikeperch larvae reared at 19–20 °C.
4.1 Pancreatic Enzymes
Pancreatic enzyme activities (trypsin and amylase) are detected shortly after hatching in Eurasian perch and pikeperch larvae (Cuvier-Péres and Kestemont 2002; Hamza et al. 2007). From hatching to mouth opening, trypsin and amylase activities increase (in Eurasian perch and pikeperch) concurrently with the exocrine activity of the pancreas signaled by first proenzyme granules in exocrine cells (Mani-Ponset et al. 1994; Ostaszewska 2005). Their relative decrease (from day 15 to 30) is due to protein deposition and secretion of other enzymes (Cahu and Zambonino Infante 1994). It does not reflect a decline of trypsin or amylase synthesis as segmental activity increases with larval development (Cuvier-Péres and Kestemont 2002).
-
Trypsin is a proteolytic enzyme secreted by pancreas acini. It is detected from hatching at a level of 30 mU mg protein−1 and reaches a peak for pikeperch larvae fed with live prey (50 mU mg protein−1 (Fig. 8.12a) compared with larvae fed with an artificial diet (Hamza et al. 2007).
Fig. 8.12 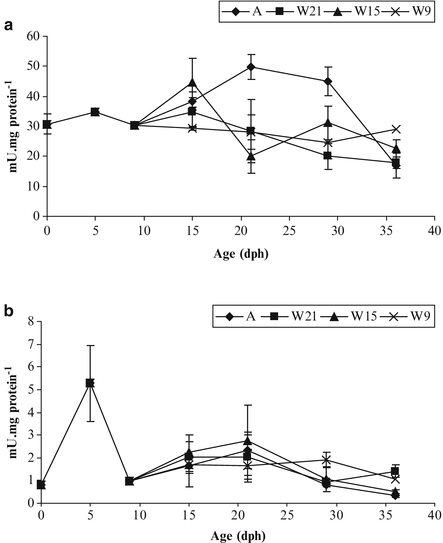
Specific activities of trypsin (a) and amylase (b) in pikeperch larvae fed Artemia (A) and weaned on day 9, 15, 21 (W9, 15, 21). Data are means ± SD (n = 4) (Modified from Hamza et al. 2007)
-
Amylase is a carbohydrase also secreted by pancreas. Also detected from hatching (0.001 U mg protein−1 in pikeperch) its activity reaches fivefolds the hatching value at mouth opening (Fig. 8.12b). Amylase is significantly affected by age but not by the nature of the diet (live prey or artificial diet) (Hamza et al. 2007). Its activity is relatively low in carnivorous species like percid fishes compared with the activity reported in omnivorous species like lipped grey mullet (Zouiten et al. 2008).

4.2 Intestinal Enzymes
-
Leucine-alanine peptidase (Leu-ala) is a peptidase present in the cytosol of the enterocytes. Its activity increases after hatching and reaches a peak at mouth opening (800 U mg protein−1 in pikeperch). This high activity reveals the development of intracellular digestion and is concurrent with the appearing of acidophilic granules observed on day 9 in the enterocytes (Fig. 8.7). A further decrease of leu-ala activity (Cuvier-Péres and Kestemont 2002; Hamza et al. 2007) indicates a lowering in intracellular digestion (Fig. 8.13a) relayed by digestion at the level of brush border membrane of the enterocytes.
Fig. 8.13 
Specific activities of intestinal enzymes leucine-alanine peptidase (a), alkaline phosphatase (b) and leucine-aminopeptidase N (c) in pikeperch larvae fed Artemia (A) and weaned on day 9, 15, 21 dph (W9, 15, 21). Data are means ± SD (n = 4) (Modified from Hamza et al. 2007)
-
Alkaline phosphatase (AP) is an esterase whose activity is induced by phosphorylated substrates such as phospholipids and phospholipoproteins. Its increase from hatching to 21 dph in pikeperch (Fig. 8.13b) traduces the development of enterocytes brush border membrane and extracellular digestion (Zambonino Infante and Cahu 2001).
-
Leucine-Aminopeptidase N (AN) is one of the intestinal brush border enzymes and hydrolyzes peptides to amino acids in the final process of protein digestion (Ugolev and De Laey 1973). It increases from hatching to mouth opening (around 40 mU mg protein−1) then decreases and finally reaches a second peak on 29 dph (35 mU mg protein−1) in pikeperch larvae (Fig. 8.13c). These two enzymes are often cited as indicator of brush border membrane development.

The strong increase of brush border membrane enzymes (leucine-aminopeptidase N, alkaline phosphatase, maltase and γ-glutamyl transpeptidase) activity occurres concurrently with the decrease of cytosolic enzyme activity (Fig. 8.13). This phenomenon occurs between 15 and 30 dph in Eurasian perch (Cuvier-Péres and Kestemont 2002) and pikeperch (Hamza et al. 2007) and reflects the development of the brush border membrane of intestinal enterocytes. Then, the larvae acquire an adult mode of digestion (Cahu and Zambonino Infante 1994).
4.3 Gastric Enzyme
-
Pepsin is a proteolytic enzyme secreted by the cells of gastric glands. The stomach development occurs between 15 and 20 dph in pikeperch (Hamza et al. 2007; Ostaszewska et al. 2005) and between 21 and 35 dph in Eurasian perch (Vlavonou 1996). It is fully achieved around day 26 dph in Eurasian perch larvae according to Domeneghini et al. (2008). Pepsin activity is detected for the first time on day 29 in pikeperch larvae (Ostaszewska 2005) and reaches 55–112 mU mg protein−1 (Hamza et al. 2007). This activity is concurrent with the gastric glands development in pikeperch (Vlavonou 1996; Ostaszewska et al. 2005; Hamza et al. 2007) as well as in Eurasian perch (Cuvier-Péres and Kestemont 2002). The functionality of the stomach (pepsin secretion) is synchronized with the metamorphosis and indicates that larvae acquire an adult mode of gastric digestion (Zambonino and Cahu 1994; Ronnestad et al. 2007). This profile has been shown in several species like turbot (Psetta maxima) and European sea bass (Cousin and Baudin Laurencin 1985).
5 Effect of the Diet on Digestive Structures and Enzymes
The enzyme activities and digestive structures development can be modified by the nature (live prey versus artificial diet) and/or biochemical composition of the diet. The importance of these factors has been demonstrated in fish larvae like European sea bass (Zambonino and Cahu 1994; Cahu and Zambonino 1995; Cahu et al. 1998, 2003a) sea bream Sparus aurata (Silva et al. 2010; Gisbert et al. 2012) as well as common carp (Escaffre et al. 1997; Gisbert et al. 2012) and percid species (Kestemont et al. 1996, 2001; Ostaszewska et al. 2005; Ostaszewska and Boruta 2006; Hamza et al. 2007, 2008).
Moreover, several authors have demonstrated that the moment of weaning and dietary nutrients like protein hydrolysates, lipids or phospholipids (PL) can affect the digestive maturation process and enzyme activities (Zambonino and Cahu 1999; Buchet et al. 2000; Cahu et al. 2003a; Wold et al. 2007; Hamza et al. 2008, 2012).
5.1 Influence of Live Prey Versus Artificial Diet
A study comparing growth, digestive structures and enzymes during ontogeny of pikeperch larvae fed live (Artemia) or artificial diets at different ages showed that larvae precociously weaned (day 9) show a lower growth (9 mg) than larvae fed with Artemia (80 mg) or weaned on day 21 (100 mg on day 36). Moreover, enzymatic assays and histological observations in digestive tract evidenced that, maturation processes of enterocytes can be impaired or delayed by an inadequate diet or a precocious weaning (Hamza et al. 2007). Indeed, in larvae weaned on day 9, number and height of enterocytes are strongly reduced and the epithelium appears atrophied compared with the larvae fed live preys or weaned later (Fig. 8.14).

Sagittal section of pikeperch larva (15 dph) fed Artemia (a) and weaned on day 9 (b). GX100. AI anterior intestine, K kidney, L liver, MI median intestine, P pancreas, S stomach (here gastric area), SB swimbladder (Modified from Hamza et al. 2007)
The modulation of enzyme activity by dietary factors varies with the type of enzyme. In pikeperch larvae as in another carnivorous species, the level of amylase specific activity is weak, and is not significantly affected by the nature of the diet (live prey or artificial diet). This was shown by Hamza et al. (2007) in an experiment based on pikeperch larvae fed live preys or weaned at different ages. The general enzymatic pattern is similar to the one of other carnivorous species (Fig. 8.12b). In fish larvae, it has been shown that trypsin is modulated by the dietary protein content (Cahu et al. 2004). In percid fishes, larvae fed live preys usually exhibit a higher trypsin activity than those fed artificial diet, as shown in Fig. 8.12a (Hamza et al. 2007).
The adaptation to dietary changes was clearly observed for intestinal enzyme activities in pikeperch larvae. In an experiment comparing the enzymatic activities of pikeperch larvae fed live (Artemia) or artificial diets at different ages, leu-ala activity remains significantly higher for larvae weaned on day 9 compared with those fed live preys or weaned later. Higher AP activity is observed after weaning, especially in larvae weaned in first days. The increase of AN in all groups except in larvae precociously weaned (day 9) may reveal a perturbation in secretory process and/or a delay in the maturation of the intestine (Fig. 8.13b, c).
5.2 Influence of a Diet Containing Casein
Some studies revealed that incorporation of protein hydrolysate in diets for fish larvae stimulates secretion of pancreatic and membranous intestinal enzymes, and improves fish survival and growth (Cahu et al. 1999; Carvalho et al. 2004). Ostaszewska et al. (2005) showed that pikeperch larvae fed formulated diets based on casein or casein hydrolysates show very low growth (around 50 mg on day 35) compared with larvae fed with Artemia or commercial diets (200 mg on day 35) and anomalies in digestive system: lower and less numerous intestinal folds (Fig. 8.15a), retarded development of gastric glands (Fig. 8.15b) and smaller hepatocytes in the liver (Fig. 8.15c).

Longitudinal section of pikeperch fed diet based on casein: posterior intestine epithelium (a), the gastric glands (b) and liver (c). Dispersed groups of necrotic hepatocytes (arrows head), dilated blood vessels (arrows) in liver. The longitudinal section of pikeperch fed Artemia nauplii: posterior intestine epithelium (d), the gastric glands (e) and liver (f). Scale bars 10 μm
These morphological changes indicated that the diets did not fulfill the fish requirements during their development. On the other hand, fish fed commercial diets and live Artemia nauplii show no anomalies in intestinal folds (Fig. 8.15d), stomach (Fig. 8.15e) and liver (Fig. 8.15f) (Ostaszewska et al. 2005). Also, the digestive enzyme (amylase, trypsin and lipase) activity is higher in fish fed commercial diets comparing with fish fed casein hydrolysates diet (Kamaszewski et al. 2010).
5.3 Influence of the Phospholipids
Several studies showed that certain nutrients can affect the digestive ontogeny, including both the digestive enzymes and structures. It is well known that PL play a major role in the structure and function of cellular membranes (Tocher 2003), in the intestinal absorption and transport of long chain fatty acids (Fontagné et al. 2000) as well as in the skeletal development (Cahu et al. 2003b). In this way, several authors showed that larvae fed with high dietary levels of lipids or phospholipids (PL) display higher digestive activities and a more developed brush border membrane and hepatocytes (Fontagné et al. 1998; Zambonino Infante and Cahu 1999; Buchet et al. 2000; Cahu et al. 2003a; Gisbert et al. 2005; Wold et al. 2007).
In pikeperch larvae, an increase in dietary PL level from 1.4 % to 9.5 % led to 50 % enhancement in final weight: 160 mg to 240 mg respectively on day 34 (Hamza et al. 2008). Moreover, a dietary level of 9.5 % PL induces a higher specific activity of AP and AN than 1.4 and 4.7 % PL suggesting better development and earlier maturation of digestive structures (Fig. 8.16).

Specific activity of alkaline phosphatase (a) and leucine-aminopeptidase N (b) during larval development of pikeperch fed with different dietary phospholipid levels (PL1, PL5, PL9 = 1.4 %, 4.7 % and 9.5 % of PL, respectively. Data are means ± SD (n = 4). Bars with different superscript letters are significantly different (P < 0.05; two-way ANOVA followed by Tukey HSD) (Modified from Hamza et al. 2008)
6 Genetic Determinism
In fish larvae, digestive enzyme synthesis is age dependent and genetically programmed (Segner et al. 1993). In this way, several studies demonstrated that pancreatic enzymes like trypsin, lipase, phospholipase or amylase are regulated at a transcriptional level during larval development (Péres et al. 1998; Cahu and Zambonino Infante 2001; Kurokawa et al. 2002; Cahu et al. 2003a). Eurasian perch and pikeperch display strong pancreatic and intestinal enzyme activities before mouth opening (Cuvier-Péres and Kestemont 2002; Hamza et al. 2007). Thus, this activity is not promoted by food intake and it can be assumed that this increase is genetically programmed.
Recent studies have investigated the effect of exogenous nutrition on the genes involved in morphogenesis. Villeneuve et al. (2006) showed that the genes involved in morphogenesis of the European sea bass can be modulated by nutrient levels like vitamin A or polyunsaturated fatty acids of phospholipids between 8 and 13 dph. In the same way, Darias et al. (2011) evidenced an effect of dietary ascorbic acid on the expression of the genes involved in acid ascorbic and calcium absorption as well as those implicated in skeletogenesis and bone mineralization. Concerning percid species, few studies treated the interactions between diet and metabolism at the cellular level. A recent study investigated the effect of nutrients on metabolic pathways in pikeperch larvae. Using proteomic, Hamza et al. (2010) evidenced that dietary PL level affected protein expression profile in the liver of 34 days old larvae. The differentially expressed proteins were involved in several cellular processes, and especially in glycolysis and gluconeogenesis. Methionine metabolism is also affected as well as structure and stress status.
7 Conclusions
This chapter aimed to resume the knowledge concerning the development and functionality of digestive structures during the ontogenesis of percid larvae (recap in Fig. 8.17). The difficulties of fish larvae to accept and digest artificial diets have often been attributed to their immature digestive system at hatching and low enzymatic capacities or to the diet composition (Lauff and Hofer 1984; Person-Le Ruyet et al. 1989; Cahu and Zambonino 2001; Ronnestad et al. 2007). As it has been demonstrated by Cahu and Zambonino (2001) for marine species, studies on digestive physiology of percid larvae showed that the digestive enzymes are present very early and their activities are high relatively to their weight (Kestemont et al. 1996; Diaz et al. 1997; Cuvier-Péres and Kestemont 2002; Hamza et al. 2007, 2008). Diaz et al. (1997) evidenced the absorption of lipids in the first stage of pikeperch development which implies suitable enzyme machinery from initiation of exogenous feeding. In the same way, Ostaszewska (2005) and Ostaszewska et al. (2005) showed an important role of pancreatic secretory products at the beginning of endo-exogenous feeding.

Main events in morphological and functional development of digestive tract during pikeperch ontogenesis
So, as it has been shown in recent studies, enzymatic capacities of the larvae do not seem to be a limiting factor (Lazo et al. 2000; Cahu and Zambonino Infante 2001; Kolkovski 2001) to digest microparticulate diets and to ensure proper growth. In this way, Hamza et al. (2007, 2008) showed that digestive enzyme activities in pikeperch larvae can be modulated by the biochemical composition of the diet. Thus, the success of feeding and better performances seems to depend more of the diet suitability to the nutritional requirements of the larvae. A better knowledge about larval digestive capacities and better approach of nutritional requirements in recent years allowed the improvement of formulation and manufacturing of artificial diets as well as weaning success (Ostaszewska et al. 2005; Hamza et al. 2007, 2008, 2012; Kestemont et al. 2007).
References
Baglole CJ, Goff GP, Wright GM (1998) Distribution and ontogeny of digestive enzymes in larval yellowtail and winter flounder. J Fish Biol 53:767–784
Balon EK (1975) Terminology of intervals in fish development. J Fish Res Board Can 32:1663–1670
Beccaria C, Diaz JP, Connes R, Chatain B (1991) Organogenesis of the exocrine pancreas in the sea bass, Dicentrarchus labrax L., reared extensively and intensively. Aquaculture 99:339–354
Bisbal GA, Bengtson DA (1995) Development of digestive tract in larval summer flounder. J Fish Biol 47:277–291
Boulhic M, Gabaudan J (1992) Histological study of the organogenesis of the digestive system and swim bladder of the dover sole, Solea solea (Linnaeus 1758). Aquaculture 102:373–396
Buchet V, Zambonino Infante JL, Cahu CL (2000) Effect of lipid level in a compound diet on the development of red drum (Sciaenops ocellatus) larvae. Aquaculture 184:339–347
Cahu CL, Zambonino JL (1995) Maturation of the pancreatic and intestinal digestive functions in sea bass (Dicentrarchus labrax): effect of weaning with different protein sources. Fish Physiol Biochem 14:431–437
Cahu CL, Zambonino Infante JL (1994) Early weaning of sea bass (Dicentrarchus labrax) larvae with a compound diet: effect on digestive enzymes. Comp Biochem Physiol A 109:213–222
Cahu C, Zambonino Infante JL (2001) Substitution of live food by formulated diets in marine fish larvae. Aquaculture 200:161–180
Cahu C, Zambonino Infante J, Escaffre AM, Bergot P, Kaushik S (1998) Preliminary results on sea bass (Dicentrarchus labrax) larvae rearing with compound diet from first feeding. Comparison with carp (Cyprinus carpio) larvae. Aquaculture 169:1–7
Cahu C, Zambonino Infante J, Quazuguel P, Le Gall MM (1999) Protein hydrolysate vs fish meal in compound diets for 10-day old sea bass Dicentrarchus labrax larvae. Aquaculture 171:109–119
Cahu CL, Zambonino Infante JL, Barbosa V (2003a) Effect of dietary phospholipid level and phospholipid: neutral lipid value on the development of sea bass (Dicentrarchus labrax) larvae fed a compound diet. Br J Nutr 90:21–28
Cahu C, Zambonino Infante J, Takeuchi T (2003b) Nutritional components affecting skeletal development in fish larvae. Aquaculture 227:245–258
Cahu C, Ronnestad I, Grangier V, Zambonino Infante JL (2004) Expression and activities of pancreatic enzymes in developing sea bass larvae (Dicentrarchus labrax) in relation to intact and hydrolyzed dietary protein; involvement of cholecystokinin. Aquaculture 238:295–308
Carvalho AP, Sa R, Oliva-Teles A, Bergot P (2004) Solubility and peptide profile affect the utilization of dietary protein by common carp (Cyprinus carpio) during early larval stages. Aquaculture 234:319–333
Cousin JCB, Baudin Laurencin F (1985) Morphogenèse de l’appareil digestif et de la vessie gazeuse du turbot, Scophtalmus maximus. Aquaculture 47:305–319
Craig JF (1987) The biology of perch and related fish. Croom Helm, Beckenham, 333 pp
Crespo S, Marín de Mateo M, Santamaria CA, Sala R, Grau A, Pastor E (2001) Histopathological observations during larval rearing of common dentex Dentex dentex L. (Sparidae). Aquaculture 192:121–132
Cuvier-Péres A, Kestemont P (2002) Development of some digestive enzymes in Eurasian perch larvae Perca fluviatilis. Fish Physiol Biochem 24:279–285
Dabrowski K, Culver D (1991) The physiology of larval fish; digestive tract and formulation of starter diets. Aquac Mag Mar/Apr:49–61
Darias MJ, Ortiz-Delgado JB, Sarasquete C, Martínez-Rodríguez G, Yúfera M (2007) Larval organogenesis of Pagrus pagrus L., 1758 with special attention to the digestive system development. Histol Histopathol 22:753–768
Darias MJ, Mazurais D, Koumoundouros G, Le Gall MM, Huelvan C, Desbruyeres E, Quazuguel P, Cahu CL, Zambonino-Infante JL (2011) Imbalanced dietary ascorbic acid alters molecular pathways involved in skeletogenesis of developing European sea bass (Dicentrarchus labrax). Comp Biochem Physiol A 159:46–55
De Silva SS, Anderson TA (1995) Fish nutrition in aquaculture. Chapman and Hall, London, 340 pp
Deplano M, Diaz JP, Connes R, Kentouri-Divanach M, Cavalier F (1991) Appearance of lipid absorption capacities in larvae of the sea bass Dicentrarchus labrax during transition to the exotrophic phase. Mar Biol 108:361–371
Diaz JP, Connes R (1991) Development of liver of the sea bass, Dicentrarchus labrax L. (Teleost, fish): II. Hepatocyte differentiation. Biol Struct Morphol 3:57–65
Diaz JP, Mani-Ponset L, Guyot E, Connes R (1997) Biliary lipid secretion during early post-embryonic development in three fiches of aquacultural interest: sea bass, Dicentrarchus labrax L., sea bream, Sparus aurata L., and pike-perch, Stizostedion lucioperca (L). J Exp Zool 277:365–370
Domeneghini C, Pannelli Straini R, Veggetti A (1998) Gut glycoconjugates in Sparus aurata L. (Pisces, Teleostei). A comparative histochemical study in larval and adult ages. Histol Histopathol 13:359–372
Domeneghini C, Vasconi M, Di Giancamillo A, Bosi G, Saroglia M (2008) Gut development in Perca fluviatilis: a micro-anatomical study. Vet Res Commun 32:S143–S145
Escaffre AM, Zambonino Infante JL, Cahu CL, Mambrini M, Bergot P, Kaushik SJ (1997) Nutritional value of soy protein concentrate for larvae of common carp (Cyprinus carpio) based on growth performance and digestive enzyme activities. Aquaculture 153:63–80
Fontagné S, Geurden I, Escaffre AM, Bergot P (1998) Histological changes induced by dietary phospholipids in intestine and liver of common carp (Cyprinus carpio L.) larvae. Aquaculture 161:213–223
Fontagné S, Burtaire L, Corraze G, Bergot P (2000) Effects of dietary medium-chain triacylglycerols (tricaprylin and tricaproin) and phospholipid supply on survival, growth and lipid metabolism in common carp (Cyprinus carpio L.) larvae. Aquaculture 190:289–303
Gisbert E, Sarasquete MC, Williot P, Castelló-Orvay F (1999) Histochemistry of the development of the digestive system of Siberian sturgeon during early ontogeny. J Fish Biol 55:596–616
Gisbert E, Piedrahita RH, Conklin DE (2004) Ontogenetic development of the digestive system in California halibut (Paralichthys californicus) with notes on feeding practices. Aquaculture 232:455–470
Gisbert E, Villeneuve L, Zambonino-Infante JL, Quazuguel P, Cahu CL (2005) Dietary phospholipids are more efficient than neutral lipids for long-chain polyunsaturated fatty acid supply in European sea bass Dicentrarchus labrax larval development. Lipids 40:1–11
Gisbert E, Skalli A, Fernández I, Kotzamanis Y, Zambonino-Infante JL, Fabregat R (2012) Protein hydrolysates from yeast and pig blood as alternative raw materials in microdiets for gilthead sea bream (Sparus aurata) larvae. Aquaculture 338–341:96–104
Govoni JJ, Boehlert GW, Watanabe Y (1986) The physiology of digestion in fish larvae. Environ Biol Fish 16:59–77
Grau A, Crespo S, Sarasquete MC, Gonzalez de Canales ML (1992) The digestive tract of the amberjack Seriola dumerili, Risso: a light and scanning electron microscopy study. J Fish Biol 41:287–303
Hamlin HJ, Hunt von Herbing I, Kling LJ (2000) Histological and morphological evaluations of digestive tract and associated organs of haddock throughout post-hatching ontogeny. J Fish Biol 57:716–732
Hamza N, Mhetli M, Kestemont P (2007) Effects of weaning age and diets on ontogeny of digestive activities and structures of pikeperch (Sander lucioperca) larvae. Fish Physiol Biochem 33:121–133
Hamza N, Mhetli M, Ben Khemis I, Cahu C, Kestemont P (2008) Effect of dietary phospholipid levels on performance, enzyme activities and fatty acid composition of pikeperch (Sander lucioperca) larvae. Aquaculture 275:274–282
Hamza N, Silvestre F, Mhetli M, Ben Khemis I, Dieu M, Raes M, Cahu C, Kestemont P (2010) Differential protein expression profile in the liver of pikeperch (Sander lucioperca) larvae fed with increasing levels of phospholipids. Comp Biochem Physiol D 5:130–137
Hamza N, Kestemont P, Ben Khemis I, Mhetli M, Cahu C (2012) Effect of different sources and levels of dietary phospholipids on performances and fatty acid composition of pikeperch (Sander lucioperca) larvae. Aquac Nutr 18:249–257
Harder W (1975) Anatomy of fishes. E. Schweizerbart’sche Verlagsbuchhandlung (Nägele u. Obermiller) Stuttgart. 612 pp
Kamaszewski M, Napora-Rutkowski Ł, Ostaszewska T (2010) The effect of feeding on activity of digestive enzymes and morphological changes in pike-perch (Sander lucioperca) liver and pancreas. Isr J Aquac 62:225–236
Kestemont P, Mélard C, Fiogbé E, Vlavonou R, Masson G (1996) Nutritional and animal husbandry aspects of rearing early life stages of Eurasian perch Perca fluviatilis. J Appl Ichthyol 12:157–165
Kestemont P, Vandeloise E, Mélard C, Fontaine P, Brown PB (2001) Growth and nutritional status of Eurasian perch Perca fluviatilis fed graded levels of dietary lipids with or without ethoxyquin. Aquaculture 203:85–99
Kestemont P, Xueliang X, Hamza N, Maboudou J, Imorou Toko I (2007) Effect of weaning age and diet on pikeperch larviculture. Aquaculture 264:197–204
Kjørsvik E, van der Meeren T, Kryvi H, Arnfinnson J, Kvenseth PG (1991) Early development of the digestive tract of cod larvae, Gadus morhua L., during start-feeding and starvation. J Fish Biol 38:1–15
Kolkovski S (2001) Digestive enzymes in fish larvae and juveniles- implications and applications to formulated diets. Aquaculture 200:181–201
Kováč V (2000) Early development of Zingel streber. J Fish Biol 57:1381–1403
Kurokawa T, Suzuki T, Ohta H, Kagawa H, Tanaka H, Unuma T (2002) Expression of pancreatic enzyme genes during the early larval stage of Japanese eel Anguilla japonica. Fish Sci 68:736–744
Lauff M, Hofer R (1984) Proteolytic enzymes in fish development and the importance of dietary enzymes. Aquaculture 37:335–346
Lazo JP, Holt GJ, Arnold CR (2000) Ontogeny of pancreatic enzymes in larval red drum Sciaenops ocellatus. Aquac Nutr 6:183–192
Mähr K, Grabner M, Hofer R, Moser H (1983) Histological and physiological development of the stomach of Coregonus sp. Arch Hydrobiol 98:344–353
Mani-Ponset L, Diaz JP, Schlumberger O, Connes R (1994) Development of yolk complex, liver and anterior intestine in pike-perch larvae, Stizostedion lucioperca (Percidae), according to the first diet during rearing. Aquat Living Resour 7:191–202
Ostaszewska T (2002) The morphological and histological development of digestive tract and swim bladder in early organogenesis of pike-perch larvae (Stizostedion lucioperca L.) in different rearing environments. Treatises and monographs. SGGW, Warsaw (in Polish)
Ostaszewska T (2005) Developmental changes of digestive system structures in pike-perch (Sander lucioperca L.). Electron J Ichthyol 2:65–78
Ostaszewska T, Boruta A (2006) The effect of diet on fatty acid composition and liver histology of pikeperch (Sander lucioperca (L.)) larvae. Arch Pol Fish 14:53–66
Ostaszewska T, Wegiel M (2002) Differentiation of alimentary tract during organogenesis in larval asp (Aspius aspius L.). Acta Sci Pol Piscaria 1:23–33
Ostaszewska T, Dabrowski K, Czuminska K, Olech W, Olejniczak M (2005) Rearing of pike-perch larvae using formulated diets- first success with starter feeds. Aquac Res 36:1167–1176
Pedersen BH, Hjelmeland K (1988) Fate of trypsin and assimilation efficiency in larval herring (Clupea harengus) following digestion of copepods. Mar Biol 97:467–476
Peña R, Dumas S, Villalejo-Fuerte M, Ortiz-Galindo JL (2003) Ontogenetic development of the digestive tract in reared spotted sand bass Paralabrax maculatofasciatus larvae. Aquaculture 219:633–644
Péres A, Zambonino Infante JL, Cahu C (1998) Dietary regulation of activities and mRNA levels of trypsin and amylase in sea bass (Dicentrarchus labrax) larvae. Fish Physiol Biochem 19:145–152
Person-Le Ruyet J, Samain JF, Daniel JY (1989) Evolution de l’activité de la trypsine et de l’amylase au cours du développement chez la larve de bar (Dicentrarchus labrax). Effet de l’âge au sevrage. Océanis 15:465–480
Ribeiro L, Zambonino Infante JL, Cahu C, Dinis MT (1999) Development of digestive enzymes in larvae Solea senegalensis, Kaup 1858. Aquaculture 179:465–473
Rønnestad I, Kamisaka Y, Conceição LEC, Morais S, Tonheim SK (2007) Digestive physiology of marine fish larvae: hormonal control and processing capacity for proteins, peptides and amino acids. Aquaculture 268:82–97
Ruuhijärvi J, Virtanen E, Salminen M, Muynda M (1991) The growth and survival of pike – perch, Stizostedion lucioperca L., larvae fed on formulated feeds. In: Lavens P et al (eds) Larvi “91. European Aquaculture Society, Special Publication No. 15, Gent, pp 154–156
Sarasquete MC, Polo A, Yúfera M (1995) Histology and histochemistry of the development of digestive system of larval gilthead seabream, Sparus aurata L. Aquaculture 130:79–92
Sarasquete MC, González Canales ML, Arellano JM, Muñoz-Cueto JA, Riberio L, Dinis MT (1996) Histochemical aspects of the yolk sac and digestive tract of larvae of Senegal sole, Solea senegalensis (Kaup, 1858). Histol Histopathol 11:881–888
Schlumberger O, Proteau JP (1991) Production de juvéniles de sandre (Stizostedion lucioperca). Aqua Rev 36:25–28
Scocco P, Accili D, Menghi G, Ceccarelli P (1998) Unusual glycoconjugates in the oesophagus of a tilapine polyhybrid. J Fish Biol 53:39–48
Segner H, Rösh R, Verreth J, Witt U (1993) Larval nutritional physiology: studies with Clarias gariepinus, Coregonus lavaretus and Scophtalmus maximus. J World Aquac Soc 24:121–134
Silva FCP, Nicoli JR, Zambonino-Infante JL, Le Gall MM, Kaushik S, Gatesoupe FJ (2010) Influence of partial substitution of dietary fish meal on the activity of digestive enzymes in the intestinal brush border membrane of gilthead sea bream, Sparus aurata and goldfish, Carassius auratus. Aquaculture 306:233–237
Stroband HWJ, Dabrowski KR (1981) Morphological and physiological aspects of digestive system and feeding in fresh-water fish larvae. In: Fontain M (ed) Nutrition des Poissons. Centre National de la Recherche Scientifique, Paris, pp 355–374
Tocher DR (2003) Metabolism and functions of lipids and fatty acids in teleost fish. Rev Fish Sci 11:107–184
Tonheim SK, Espe M, Raae AJ, Darias MJ, Rønnestad I (2004) In vivo incorporation of [U]- 14C-amino acids: an alternative protein labelling procedure for use in examining larval digestive physiology. Aquaculture 235:553–567
Treviño L, Alvarez-González CA, Perales-García N, Arévalo-Galán L, Uscanga-Martínez A, Márquez-Couturier G, Fernández I, Gisbert EA (2011) A histological study of the organogenesis of the digestive system in bay snook Petenia splendida Günther, 1862 from hatching to the juvenile stage. J Appl Ichthyol 27:73–82
Ugolev AM, De Laey P (1973) Membrane digestion. A concept of enzymic hydrolysis on cell membranes. Biochem Biophys Acta 300:105–128
Villeneuve LAN, Gisbert E, Moriceau J, Cahu CL, Zambonino Infante JL (2006) Intake of high levels of vitamin A and polyunsaturated fatty acids during different developmental periods modifies the expression of morphogenesis genes in European sea bass (Dicentrarchus labrax). Br J Nutr 95:677–687
Vlavonou RS (1996) Elevage expérimental de la perche Perca fluviatilis L., Développement larvaire et croissance. Dissertation, University of Metz
Watanabe Y (1984) Morphological and functional changes in rectal epithelial cells of pond smelt during postembryonic development. Bull Jpn Soc Sci Fish 50:805–814
Wold P-A, Hoehne-Reitan K, Cahu C, Zambonino Infante J, Rainuzzo J, Kjørsvik E (2007) Phospholipids vs. neutral lipids: effects on digestive enzymes in Atlantic cod (Gadus morhua) larvae. Aquaculture 272:502–513
Zambonino Infante JL, Cahu CL (1994) Influence of diet on pepsin and some pancreatic enzymes in sea bass (Dicentrarchus labrax) larvae. Comp Biochem Physiol A 109:209–212
Zambonino Infante JL, Cahu L (1999) High dietary lipid levels enhance digestive tract maturation and improve Dicentrarchus labrax larval development. J Nutr 129:1195–1200
Zambonino Infante JL, Cahu CL (2001) Ontogeny of the gastrointestinal tract of marine fish larvae. Comp Biochem Physiol C 130:477–487
Zouiten D, Ben Khemis I, Besbes R, Cahu C (2008) Ontogeny of the digestive tract of thick lipped grey mullet (Chelon labrosus) larvae reared in “mesocosms”. Aquaculture 279:166–172
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2015 Springer Science+Business Media Dordrecht
About this chapter
Cite this chapter
Hamza, N., Ostaszewska, T., Kestemont, P. (2015). Development and Functionality of the Digestive System in Percid Fishes Early Life Stages. In: Kestemont, P., Dabrowski, K., Summerfelt, R. (eds) Biology and Culture of Percid Fishes. Springer, Dordrecht. https://doi.org/10.1007/978-94-017-7227-3_8
Download citation
DOI: https://doi.org/10.1007/978-94-017-7227-3_8
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-017-7226-6
Online ISBN: 978-94-017-7227-3
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)