
Abstract
Until 1988, a small number of outbreaks of native pests had been recorded in the 1.84 million ha of pine plantations in Brazil. Most of these stands were planted with little species diversity, and at high density and received inadequate management. In 1988, an outbreak of Sirex noctilio was first recorded in southern Brazil and the insect currently infests 450,000 ha in the southern and southeast regions. The presence of Sirex has the potential to cause losses of up to $US30 million annually. The discovery of this insect resulted in an immediate change in silvicultural practices used in forest plantations, with the introduction of Integrated Pest Management (IPM) as a part of the Forest Management Programs. In 1989, the National Programme of Woodwasp Control (PNCVM) was established. This programme has a broad scope and includes the early detection and monitoring of the spread of Sirex through the use of trap trees (trees artificially stressed with herbicides) and ground inspections. Appropriate silvicultural practices, especially stand thinning, preventing competition and providing healthy growing conditions are also an important aspect of control programs. Curative measures used are based on biological control using the nematode parasite Deladenus (=Beddingia) siricidicola, introduced from Australia, which sterilizes the females and reaches up to 100% parasitism. The egg parasitoid Ibalia leucospoides has also been introduced and can reach about 25% parasitism. Two other parasites, Rhyssa persuasoria and Megarhyssa nortoni were introduced into Brazil between 1996 and 1998, and in 2003, from Australia. Establishment of these two parasites has not been confirmed. This combination of biological control and appropriate silvicultural practices are essential to keep the Sirex population under control in Brazil.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
16.1 Introduction
Sirex noctilio F. (Hymenoptera: Siricidae) is a minor pest in its regions of origin, Europe, Asia and North Africa (Chap. 5). In contrast, it has become the main pest of pine plantations where it has been introduced, in countries such as New Zealand, Australia, Uruguay, Argentina, Brazil, Chile and South Africa (Chaps. 13–18).
In South America, S. noctilio detected for the first time, in Uruguay in 1980 infesting several species of Pinus, with P. taeda being the most susceptible (Rebuffo 1988). Its presence in Argentina was first recorded in the province of Entre Rios, in the northwest of the country, between 1985 and 1986, where it was found infesting P. taeda and P. elliottii (Espinoza et al. 1986, Chap. 15). The first record of S. noctilio in Brazil was in February 1988, in a P. taeda plantation, in the municipal districts of Gramado, Canela and São Francisco de Paula, in Rio Grande do Sul State (Iede et al. 1988). In the following year, the insect was detected in trap trees, in a P. taeda plantation located in the municipal district of Lages, in Santa Catarina State. In Paraná State, two interceptions occurred in 1993 and 1994, but there was no evidence of pest establishment. However, in July 1996, its establishment was verified in P. taeda plantations in several municipal districts of Paraná State (Iede et al. 1998). The first record in the Southeast Region was in 2005, in São Paulo and Minas Gerais States. The woodwasp can now be found in 450,000 ha of pines throughout the states of Rio Grande do Sul, Santa Catarina, Paraná, Sao Paulo and Minas Gerais.
In Brazil there are 1.84 million ha of pine plantations. Most of the areas where S. noctilio is established have historically been planted with little species diversity and at a high density, and in some cases subject to inappropriate silvicultural practices. These conditions are ideal for pest and disease outbreaks. In the Southern and Southeastern regions, there are approximately 1.2 million ha of mainly P. taeda (loblolly pine) and P. elliottii (slash pine) established in plantations.
The presence of S. noctilio in Brazil awakened the Brazilian forest sector to the need for control strategies, prevention and monitoring actions because this exotic woodwasp has caused significant economic losses, mainly in the P. taeda and P. elliottii plantations. The National Fund for Woodwasp Control (FUNCEMA) was thus created. FUNCEMA involves public and private organizations working together to support the National Programme of Woodwasp Control (PNCVM).
Monitoring with trap trees is the most important activity of this program. In areas where the pest has not yet been detected, it is possible to carry out early detection and immediate release of biological control nematodes when necessary. It has been recommended that trap tree monitoring can cease when the number of attacked trees is 1%, and at that point to start using the sequential sampling technique described by Penteado et al. (1998, 2002a) (see discussion below). Biological control is the most efficient means to keep S. noctilio under control, especially when the nematode, which sterilizes up to 70% of the females, is used. It is also important to use forest management to maintain high levels tree vigor and health in the stands.
16.2 Biology and Ecology of Sirex noctilio
In Brazil, S. noctilio infests Pinus taeda, P. elliotti, P. patula (mainly in Southern region of Brazil) and P. oocarpa, P. caribaea caribaea, P. caribaea bahamensis, P. caribaea hondurensis, P. kesiya and P. strobus chiapensis (in the Southeast Region). In Brazil, the normal life cycle of the insect usually takes 1 year; however, it can have a short cycle, also called a summer cycle, which varies from 3 to 4 months. This cycle can occur in dominated or even in co-dominant trees (Iede et al. 1988). In the South of Brazil, Carvalho et al. (1993) and Iede et al. (1993) verified that S. noctilio adults emerge between October and April, with peaks in emergence during the months of November and December.
According to Carvalho (1993), the longevity of adults in the field is of 4 and 5 days for females and males, respectively. Iede et al. (1988) obtained an average of 8 days survival for adults, in climatic chambers (25 ± 1°C; RH 70 ± 10%, and photoperiod of 12 h). The males emerge before the females and the male/female ratio varies from 1.5:1 to 32:1.
After the initial flight period, females penetrate the tree trunk with their ovipositors and lay their eggs in the sapwood; they can lay up to four galleries in each hole, with an average of 2.2 eggs per hole. Each female lays an average of 210 eggs in approximately 5 days (Carvalho et al. 1993). During this process, females introduce spores (arthrospores) of the symbiotic fungus (Amylostereum areolatum) and a mucous secretion; both the fungus and the toxin is thought to contribute to the death of trees. The pathogenic fungus, which is also a source of nutrients for the larvae, dries up the wood and causes rot. The wood quality is also affected by the larvae constructing tunnels and by the entrance of secondary agents that help damage the wood, limiting its use or ruining it for the wood market. Once trees are dead, the wood degrades rapidly and it must be used within 6 months of infestation. Penteado et al. (2000) verified that most of the larvae are distributed in the top half of the tree.
Plantations that are more susceptible to S. noctilio are generally up to 8 years-old and under stress. Stands that have not been thinned are more susceptible to infestation than those that have been thinned. The symptoms of infestation begin right after the insect population peaks, in November and December, but become more visible after March. The external symptoms of attack are a progressive yellowing of the crown, which later becomes brownish-red, wilting of the foliage, loss of needles, resin drops on the bark (due to the oviposition holes) and emergence holes. Internal symptoms are brown spots along the inner bark caused by the fungus A. areolatum and tunnels drilled by larvae, which affect wood quality.
16.3 The National Program of Woodwasp Control in Brazil
Sirex noctilio is present in 450,000 ha, in the Southern and Southeast regions of Brazil. The Brazilian forest sector went through a difficult phase due to the damage by this pest. It is estimated that the potential losses caused by this insect may reach US$ 30 million annually. Urgent and efficient strategies were required to monitor, control and delay the spread of S. noctilio. The seriousness of the situation resulted in an immediate change in the handling of forest plantations, with introduction of Integrated Pest Management (IPM) as part of the general forest management programs. Biological control associated with prevention measures is the most effective and economical method to combat damage due to this insect, because as an exotic pest, it was introduced without its complex of natural enemies (Iede et al. 1998).
In June 1989, FUNCEMA was created to manage the S. noctilio problem in Brazil. FUNCEMA is a not-for-profit civil organization formed by private and public institutions whose main objective is to generate funds for the development of the National Programme of Woodwasp Control (Iede et al. 1989). This program includes research and development activities such as: (a) monitoring for the early detection of the pest using trap trees intentionally stressed with herbicides to attract the insects; (b) adopting prevention strategies to improve the phytosanitary conditions of forest stands using silvicultural practices, especially thinning, to prevent and minimize attacks; (c) adopting quarantine strategies to control and slow- down dispersal; (d) mass rearing and releasing the nematode Deladenus siricidicola (Nematoda: Neotylenchidae); (e) mass rearing and releasing the parasites Rhyssa persuasoria and Megarhyssa nortoni (Hymenoptera: Ichneumonidae) collected in Australia; (f) field-collecting and releasing the parasite Ibalia leucospoides (Hymenoptera: Ibaliidae), to increase the range of natural enemies; (g) developing an extensive training program for technical personnel and forestry workers to improve pest management; and (h) providing information to the public (Iede et al. 1989).
The National Programme of Woodwasp Control is an excellent example of the integration of a research and development policy nationwide, as more than 120 private companies in Southern Brazil are involved together with public organizations. The companies also offer technical assistance to small private forest owners so the control strategies will reach all plantations attacked by the woodwasp (Iede et al. 1989).
16.3.1 Prevention Strategies
Trees resistant to S. noctilio are those that remain free of injuries and continue to grow vigorously on good sites and in well-managed blocks. The level of mortality of the trees is significantly related to their diameter at breast height (DBH). Trees with small DBH are more likely to be attacked than larger individuals in the same stand. However, during the event of a broad abiotic disturbance dominant trees can also be attacked.
Effective forest management practices limit rotation lengths and encourage forest managers to consider the composition, structure, age and vigor of the plantation. According to Davis (1966), more effective pest control may be obtained through silvicultural practices in the long run, creating reasonable insect-resistance in the forest. Complete control will never be attained this way, but the loss caused by insects can be reduced.
Thinning is one of the most important silvicultural practices in pine plantation forestry. It is conducted to accelerate or modify the course of competition. The position of the crown is an important characteristic in deciding which trees to cut and which to retain. Vigorous trees that outgrow their neighbors dominate the canopy and usually have a greater chance of surviving future competition than less dominant ones, which take lower positions in the plantation. Most of the thinning practices reduce loss due to damaging agents not only because thinning is a prevention strategy but also because it increases the vigor and resistance of the remaining trees. Thinning can enhance the susceptibility of trees to insect attack only under certain circumstances, such as when it is carried out during the flight period of the pest. Pruning should also be avoided during the flight season because pruning-related injuries release some chemical compounds, like monoterpenes, that attract the female woodwasps to oviposit in the damaged trees. Even in well-managed stands, e.g., a thinned, 8 year-old, low density (650 trees/ha) stand with trees 28 cm DHB, was observed tree mortality of 8% because pruning was done during the S. noctilio flight season.
16.3.2 Early Detection and Monitoring of S. noctilio
16.3.2.1 Early Detection Using Trap Trees
The use of trap trees stressed by herbicide injection is the most appropriate and efficient technique for the early detection of the pest, and for monitoring Sirex dispersal. Detecting S. noctilio during its early stages of development and colonization helps to define locations for release of biological control agents and allows thinning practices to be carried out before the pest reaches high levels of damage. Maintaining a trap tree system can greatly increase the efficiency of the biological control of this woodwasp.
At the beginning of the IPM Sirex project in Brazil, the herbicide Dicamba was used to stress trees, but after some years, the product was no longer sold in Brazil. Some experiments were carried out with other products; Padron, which showed efficacy similar to Dicamba, has been used since that time.
The detection method, as well as the intensity with which it should be applied, must be based on a risk analysis of the introduction and dispersal of the pest in each region. Penteado et al. (2002b) recommended that the trees intended as trap trees should have a DBH of between 10 and 20 cm and be installed in groups of five trees. In areas where Sirex is already present, as well as in areas up to 10 km away from the infestation foci, the recommendation is to install groups of five trap trees every 500 m, which means, four groups of five trees in each 100 ha. At distances between 11 and 50 km from the infestation, the trap tree groups should be spaced 1,000 m apart.
The installation of trap trees in Brazil must be done from August 15 to September 30, 2 months before the population of the adult woodwasps peak, which generally occurs between November and December. During February to May, the groups of trap trees should be revisited to check for insect attack (Penteado et al. 2002b). Trap trees should be installed in susceptible plantations, close to saw mills, along the main wood transportation routes and on the borders of the area of natural dispersal of the pest. After detection, trap trees must be installed annually in areas with percentages of attack less than 1%. When more than 1% of the trees are attacked, the trap tree method is changed to the sequential sampling method.
16.3.2.2 Remote Sensing
The monitoring of S. noctilio attack is an essential component of an integrated pest management program. The national program originally intended to have all pine plantations in the country mapped from satellite images with the attacked trees identified on the map. Although it was possible to identify pine plantations, it was not possible to conclusively identify trees that had symptoms of Sirex attack.
Aerial sketch mapping was tested for monitoring and early detection of the pest, but because the pest initially attacks subdominant trees under the forest canopy, the method was not appropriate. According to Haugen et al. (1990), infested trees are not always visible in an aerial inspection, and this kind of evaluation is not recommended when the level of attack is lower than 5% (also see Chap. 14).
16.3.2.3 Sequential Sampling
Haugen et al. (1990) observed that methods for monitoring S. noctilio in Australia at the time (see Chap. 14 for current methods in Australia) were characterized by low precision and high operational costs, emphasizing the need for the development of more practical and economical methods. It was, therefore, necessary to establish a damage sampling technique that is practical and takes into consideration the vast stands of pines, the levels of attack, and the cost of the activity.
Sequential sampling, developed by Penteado et al. (1998, 2002a), in which the size of the sample is defined according to the results obtained during the application of the procedure, has proven to be a good alternative for evaluating the attack intensity of S. noctilio in pine plantations. This type of sampling reduces the time and cost of application and gives immediate results. The use of a sequential sampling Table (Table 16.1) reduced the size of the sample between 45% and 86%, depending on the level of attack and in comparison to the fixed-size procedure generally recommended. Sequential sampling saves time and money and increases precision when reduced sample sizes are used.
The sequential sampling technique is used as follows:
-
1.
Start by sampling 68 trees;
-
2.
Note the number of trees attacked in the sample in the second column of the table and compare it to the number of trees attacked shown in the third column of the table, in this case 34;
-
3.
If the number of trees attacked in the sample is equal to or greater than 34, the sampling procedure is complete;
-
4.
If the number of trees attacked in the sample is lower than 34 the procedure must be continued by sampling six more trees, totalling 74 sampled trees;
-
5.
If the number of trees attacked is 36 or more, the sampling should be interrupted;
-
6.
If the number of trees attacked is lower than 36, the sampling must continue until the number of attacked trees shown in the third column of the table is attained or until 272 trees have been sampled;
-
7.
Calculate the percentage of trees attacked as a function of number of trees sampled.
When establishing the sampling path in the plantation for this type of inspection, consider the distance covered and the amount of time spent, and maximize the coverage of the area. To start the sampling procedure, select a row (path) in which 40 trees will be checked. When this number is achieved in one row, skip five rows and begin again on 40 trees of the fifth row, continuing until the sampling is completed. In blocks larger than 10 ha, however, it is advisable to skip ten rows instead of five to guarantee greater coverage of the area under evaluation. This method of sampling in a plantation is similar to a step-like pathway (Fig. 16.1). We have observed that Sirex attacks are usually randomly distributed (not in groups or clusters) regardless of their intensity. This has also been noted by the National Sirex Co-ordination Committee (1991), and contributes to the generation of more precise sampling results.

Pathway to conduct sequential sampling. (a) Evaluation of S. noctilio attack on 40 trees, (b) Five or ten rows are skipped
It is recommended that sequential sampling be applied in areas up to 50 ha in size. The criteria that define the number of necessary samples in a forest block, is characterized by the soil type, relief, management practice, age and density, among other factors, can be used to define the appropriate sampling intensity for each area according to its size. The stands should, therefore, be sampled block by block, as these are considered homogeneous. If, however, there are biotic or abiotic conditions within a block that may stress the trees, at least one extra sampling unit is recommended.
The best time for sampling must be defined by its purpose. In Brazil, S. noctilio adults from the normal life cycle are found between the second half of October and the first half of January. Thus sampling done between March and April will indicate the number of trees that should be inoculated with nematodes. The nematode inoculations are done between March and August. Sampling conducted later than that, between June and July, will indicate the total mortality in the period but may interfere with the planning of the activities of inoculation with nematodes, as has also been observed by the National Sirex Co-ordination Committee (1991).
16.3.3 Biological Control
Biological control, along with prevention strategies, is the most efficient and economical method for controlling S. noctilio in Brazil. A system of control similar to that adopted in other countries (see for example Chaps. 8, 9, 13–18) where Sirex has been introduced was developed in Brazil. These biological controls include the nematode D. siricidicola and the parasites, I. leucospoides, R. persuasoria and M. nortoni.
16.3.3.1 Nematodes
The most effective biological control agent of the woodwasp is the nematode D. siricidicola, which sterilizes the females. This nematode has two life cycles: a free-living cycle, when it feeds on the same symbiotic fungus as the woodwasp and a parasitic-life cycle, which occurs inside larvae, pupae and adults of S. noctilio. Because its free-living cycle feeds on the fungus A. areolatum, the nematode is easily bred under laboratory conditions and then released in the field by application into trees attacked by S. noctilio. The 20 ml doses of nematodes contain approximately one million nematodes (Bedding 1972). The inoculation of nematodes into trees is done with a specially developed hammer that is used to make holes in the trunks of infested tree at a 30 cm spacing (see images in Chap. 14). The doses are mixed in a 10% solution of gelatine and introduced by a syringe into the holes in the wood made with the hammer (Penteado et al. 2002b). After inoculation, the nematodes penetrate the wood in search of the fungus they feed on and reproduce in free-living cycle. However, when they locate Sirex larvae, they develop into infective adults and penetrate the larvae, leaving scars in their integuments. When the host pupates, the juvenile nematodes are released into the body of the host and move to the reproductive organs. They then penetrate the eggs, sterilizing the female. The infected adult female Sirex emerge from the trees and lay their eggs in susceptible trees but these eggs are not fertile, and can contain between 100 and 200 nematodes (Bedding 1972).
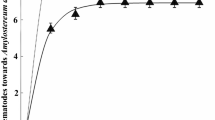
In Brazil, the first introduction of nematodes was in 1989 and 1990, from the Australian Commonwealth Scientific and Industrial Research Organization (CSIRO). The inoculations were done between March and August, when most of the larvae are inside the tree, and the nematode can be more effective. Three years after the first inoculations, it was observed that parasitism levels were very low. The average levels of parasitism obtained in Australia with the nematode was 70%. The level of parasitism verified for the nematode in attacked areas in Rio Grande do Sul and Santa Catarina states has been quite variable. It was found to be as high as 70% in a 12,000 ha P. taeda plantation, in Encruzilhada do Sul, Rio Grande do Sul, but in some places we have observed parasitism levels ranging from 10% to 90%.
It was discovered that the introduced strain of nematode became defective because it had been cultured in the free-living form for over 20 years in Australia without the intervention of the parasitic life cycle, which led to the selection of a strain that rarely formed the infective stage. Unfortunately the situation did not become apparent until this defective strain had been released for many years (Bedding and Iede 2005; also see Chap. 9). Two strategies were used to solve the problem in Brazil. These included importing (in 1994) the Kamona strain, which was re-isolated in Tasmania, from CSIRO, Australia, and releasing it in 1995; and also re-isolating the nematode from the field, in Brazil every year, and introducing the field-captured nematodes into the mass rearing programme. Every year, the nematode parasitism levels are evaluated and parasitism levels are currently above 70%. In most places where the nematode was released, it is difficult to find attacked trees because the Sirex population is under control.
16.3.3.2 Parasites
The parasite Ibalia leucospoides was recorded in Brazil for the first time in December 1990, in a pine plantation attacked by the woodwasp around the city of São Francisco de Paula, Rio Grande do Sul (Carvalho et al. 1991). It is now established in all areas where the woodwasp occurs with parasitism as high as 39%, but averaging around 25%.
Megarhyssa nortoni and Rhyssa persuasoria were introduced to Brazil from Tasmania in 1996, 1997 and 1998, in a cooperative project between Embrapa Florestas, International Institute of Biological Control (CABI-Bioscience) and the USDA (United States Department of Agriculture) Forest Service. The first introduction of M. nortoni was done in 1996 with 133 individuals (88 females and 45 males). In October 1997, an experimental release of M. nortoni was conducted in Correia Pinto, in Santa Catarina State. In this place 18 previously mated females were released next to trap trees. In October 1998, to verify the establishment of the parasite, traps were installed on 10 trees near the points of release and pine logs were collected from ten trees located near the point. However, the presence of the insect was not recorded. In 1998, five field releases were carried out between October and December, with a total of 97 females and 134 males of M. nortoni and ten females and two males of R. persuasoria. Areas selected for release had been attacked by S. noctilio, were close to native forests, and had not been thinned to allow the establishment of parasites. Also in 1998 parasites were introduced from Tasmania, however, due to problems during the collection of insects in the field, the number of insects was very low, with only 15 females of M. nortoni and one female of R. persuasoria being released.
In 2000, a total of 86 males and 67 females of M. nortoni and 35 males and 70 females of R. persuasoria were used to determine longevity at different temperatures. Average longevity for M. nortoni was 17.9 and 68.87 days, at 23°C and 12°C, respectively, and for R. persuasoria, average longevity was 20.11 and 50.28 days at 23°C and 12°C, respectively. In 2005, M. nortoni adults were released in one area located in Rio Branco do Ivaí, Paraná, but its establishment has not yet been confirmed.
16.3.4 Quarantine Strategies
Sirex noctilio can spread naturally between 30 and 50 km/year. However, the transportation of infested wood to uninfested areas increases the possibility of dispersal. Probably this is the way the pest was introduced into Brazil from Uruguay. For that reason, monitoring affected areas and a quarantine system that prohibit the transportation of infested wood outside defined quarantine areas are needed.
In Brazil, moving wood from a Sirex-infested area to a Sirex-free area is permitted with an Origin Phytosanitary Certificate provided by the National Plant Protection Organization. In areas near the border of areas known to contain Sirex, trap tree monitoring is required each year. Fumigation of wood or heat treatment (minimum of 56°C throughout the profile of the wood for a minimum of 30 min) to control insects inside the wood is also required.
16.4 General Recommendations
Sirex noctilio is essentially a secondary and an opportunistic pest. The use of prevention measures in pine plantations is the most appropriate way to avoid economic losses. These preventive measures can be achieved by monitoring the plantations and by the use of appropriate silvicultural practices, such as thinning. After the initial damage by S. noctilio in Brazil, it was estimated that most plantations showed low levels of mortality and only a small portion severely infested. However, this status will change if monitoring, prevention and control strategies are not carried out.
Further research is needed to understand the genetics of the nematode D. siricidicola and its symbiotic fungus A. areolatum in different regions Brazil. This is especially because differences in levels of parasitism between different locations and also differences in the development of different isolates of the nematode and fungus in the laboratory have been observed. Cryopreservation of isolates of the fungus and nematode is needed to prevent loss of nematode infectivity and to be able to preserve isolates selected as promising for biological control.
References
Bedding RA (1972) Biology of Deladenus siricidicola (Neotylenchidae) an entomophagous-mycethophagous nematode parasitic woodwasps. Nematology 18:482–493
Bedding RA, Iede ET (2005) Application of Beddingia siricidicola for sirex woodwasp control. In: Grewel PS, Ehlers R, Shapiro-Ilan DI (eds) Nematodes as biocontrol agents. CAB International, Wallingford
Carvalho AG (1993) Parasitismo de Ibalia sp. (Hymenoptera: Ibaliidae) em Sirex noctilio Fabricius, 1793 (Hymenoptera: Siricidae) em São Francisco de Paula, RS. Boletim de Pesquisa Florestal, Colombo, no. 26/27
Carvalho AG, Berti Filho E, Vadas J, Maia SJLS (1991) Ocorrência de Ibalia sp.(Hymenoptera, Ibaliidae) em Sirex noctilio F. (Hymenoptera, Siricidae) no Rio Grande do Sul. Revista de Agricultura da USP, vol 66. USP (Escola Superior de Agricultura Luiz de Queiroz), Piracicaba – São Paulo
Carvalho AG, Pedrosa-Macedo JH, Santos HR (1993) Bioecologia de Sirex noctilio F. 1793 (Hymenoptera: Siricidae) em povoamentos de Pinus taeda L. In: Conferência Regional da Vespa-da-Madeira, Sirex noctilio, na América do Sul, Florianópolis, 1992. Anais. Embrapa-CNPF/FAO-ONU/USDA – Forest Service, Colombo
Davis KM (1966) Forest management: regulation and valuation, 2nd edn. McGraw-Hill, New York
Espinoza ZH, Lavenderos A, Lobos C (1986) Reconocimiento de la plaga Sirex noctilio en plantaciones de pinos de Uruguay y Argentina. Corporación Nacional Forestal, Santiago
Haugen DA, Bedding RA, Underdown MG, Neumann FG (1990) National strategy for control of Sirex noctilio in Australia. Aust For Grow 13:1–8, special liftout section
Iede ET, Penteado SRC, Bisol JC (1988) Primeiro registro de ataque de Sirex noctilio em Pinus taeda no Brasil. EMBRAPA-CNPF, Circular Técnica, Colombo
Iede ET, Bedding RA, Penteado SRC, Machado DC (1989) Programa Nacional de Controle da vespa-da-madeira-PNCVM. EMBRAPA-CNPF, Curitiba
Iede ET, Penteado SRC, Gaiad DCM, Silva SMS (1993) Panorama a nível mundial da ocorrência de Sirex noctilio F., (Hymenoptera:Siricidae). In: Conferência Regional da Vespa-da-Madeira, Sirex noctilio, na América do Sul, 1992, Florianópolis. Anais. EMBRAPA-CNPF, Colombo
Iede ET, Penteado SRC, Schaitza EG (1998) Sirex noctilio problem in Brazil - detection, evaluation and control. In: Iede ET, Schaitza EG, Penteado SRC, Reardon R, Murphy T (eds) Training in the control of Sirex noctilio by use of natural enemies. USDA Forest Service, Morgantown
National Sirex Co-ordination Committee (1991) National Sirex control strategy - operations worksheets. Canberra, Australia
Penteado SRC, Oliveira EB, Iede ET (1998) Sampling methods for evaluating Sirex noctilio attack levels in Pinus taeda stands and for monitoring the efficiency of its natural enemies. In: Iede ET, Schaitza EG, Penteado SRC, Reardon R, Murphy T (eds) Training in the control of Sirex noctilio by use of natural enemies. USDA Forest Service, Morgantown
Penteado SRC, Oliveira EB, Iede ET (2000) Distribuição da vespa-da-madeira e de seus inimigos naturais ao longo do tronco de Pinus. Boletim de Pesquisa Florestal, Colombo
Penteado SRC, Oliveira EB, Iede ET (2002a) Aplicação da amostragem sequencial para monitoramento dos níveis de ataque de Sirex noctilio em povoamentos de Pinus taeda. Embrapa Florestas, Boletim de Pesquisa e Desenvolvimento, Colombo
Penteado SRC, Iede ET, Reis Filho W (2002b) Manual de controle da vespa-da-madeira (Sirex noctilio) em plantios de Pinus spp. Embrapa Florestas, Colombo
Rebuffo S (1988) La “avispa de la madera” Sirex noctilio F, en el Uruguay. Montevideo: Ministerio de Ganaderia, Agricultura y Pesca, Direccion Forestal. Serie Proteccion Forestal 1:1–14
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2012 Springer Science+Business Media B.V.
About this chapter
Cite this chapter
Iede, E.T., Penteado, S.R.C., Filho, W.R. (2012). The Woodwasp Sirex noctilio in Brazil: Monitoring and Control. In: Slippers, B., de Groot, P., Wingfield, M. (eds) The Sirex Woodwasp and its Fungal Symbiont:. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-1960-6_16
Download citation
DOI: https://doi.org/10.1007/978-94-007-1960-6_16
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-1959-0
Online ISBN: 978-94-007-1960-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)