
Abstract
Global warming scenarios combined with political and public awareness have led to increasing funding and research efforts on the measurement and prediction of effects of a changing world on the ecosystems. Fish parasites represent a major part of aquatic biodiversity, and consequently become affected either directly through the environment or indirectly through their respective hosts. On the basis of a conservative estimate of an average of 3–4 fish parasites in each existing fish species alone and a current number of 31,400 described fish species, we can estimate the existence of up to 120,000 fish parasite species, including both protozoans and metazoans. Combined with a number of life cycle stages that may infect all aquatic hosts and organs, this vast biodiversity represents a widely neglected tool for a variety of ecology-based applications. Studies have demonstrated that fish parasites can serve as biological indicator organisms to illustrate the ecology of their infected hosts, including feeding, migration and population structure. Parasite metrics have been connected to specific environmental conditions, and they can indicate different pollutants such as heavy metal concentrations, industrial and sewage pollution, and also eutrophication. Most recently, parasite infections have been connected to anthropogenic impact and environmental change also in marine habitats. This book chapter summarizes the use of fish parasites as biological indicators, and discusses their potential and the requirements in the utilization of fish parasites as biological indicators of climate change.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
Aquatic ecosystems along the costal zones belong to the most vulnerable systems on earth and face increasing anthropogenic stress in terms of pollution and environmental degradation. About 2.75 bn people are expected to live within 60 mi of the coastline in 2025, living from or indirectly using the coastal environments. This, however, is the region that harbours the highest aquatic biodiversity, especially in tropical coastal waters. It is obvious that extensive anthropogenic activities directly affect the species composition and diversity of the aquatic biota, possibly negatively influencing the long-term perspectives and sustainability of these ecosystems.
Fish parasite biodiversity and species composition in the aquatic realm depends on species richness of the final hosts and their ecosystem. The global fish fauna comprises more than 31,400 species (Froese and Pauly 2010), about half of them (14,970 species) live in marine waters. Because of the long-term stability of marine ecosystems, fish parasite diversity per host is higher than in freshwater. Rohde (2002) estimated 100,000 fish parasites in about 30,000 known fish species, resulting in an average of 3.3 parasite species in each fish studied. Margolis and Arthur (1979) and McDonald and Margolis (1995) recorded 925 different fish parasites on 292 marine and freshwater fish species from Canadian waters, including protozoans and metazoans (3.2 parasite species/fish species). Palm et al. (1999) reported 191 different metazoan parasite species from another northern habitat, the coastal waters of Germany. A total of 62 wild fish species from the North and Baltic Sea coast harboured an average of 3.1 metazoan parasite species per fish species. This contrasts the deep-sea, where the average number of parasites per fish species is 1.5, a value that did not increase in the last 8 years (Klimpel et al. 2001, 2009). On the basis of the existence of more than three metazoan fish parasites in each existing fish species alone, we can estimate the existence of up to 120,000 fish parasite species, including both protozoans and metazoans.
The aquatic environment can be studied either directly by a regular monitoring of water quality parameters or indirectly by using bioindicators (also see Palm and Rückert 2009), such as fish parasites (Galli et al. 2001). These organisms react on specific environmental conditions or change, leading to a wide range of applications (bioindication for water quality, MacKenzie et al. 1995; environmental stress, Landsberg et al. 1998; pollution, Khan and Thulin 1991; Yeomans et al. 1997). Vidal-Martínez et al. (2010) generally distinguished between accumulation or effect bioindicators, where organisms efficiently take up substances in the former or are used to detect environmental impact in the latter. This is done while recording a definite change in their physiology, chemical composition, behaviour, or number. Also other parasite metrics such as diversity indices or species richness can be a source of information (e.g. Rückert et al. 2009a), demonstrating a possible effect of specific environmental conditions on the fish parasite community. Among others, Lafferty (1997), Marcogliese and Cone (1997), Overstreet (1997), Williams and MacKenzie (2003), Marcogliese (2005) and most recently Vidal-Martínez et al. (2010) summarized the published literature on how to use fish parasites as bioindicators in the aquatic environment.
In the context of long-term and climate change scenarios, rising sea-level and water temperatures may have direct effects on the fish parasite composition within a respective habitat. Only very few freshwater habitats, however, are under pristine conditions, and anthropogenic species introduction connected to fisheries combined with regular migration events of neozoons alter the regular fish and parasite fauna. Most marine environments have suffered heavy fishing pressure over the last century. Anthropogenic changes have greatly altered the fish species composition, especially of large predators at high trophic levels (Hutchings and Baum 2005; Baum and Worm 2009). This has measurable effects even on life history traits, substantially changing age and size at maturation (Sharpe and Hendry 2009). Consequently, fish parasite numbers that are related to their changing host numbers may also change with a shift in environmental conditions. A conclusive description of the circumstances under which parasites can be used as indicators of environmental impact, however, still remains difficult (Vidal-Martínez et al. 2010).
The present chapter describes a simplified method of routine fish parasitological investigations, presents an overview on the available literature, and provides different examples on how to use the resulting fish parasite metrics as biological, accumulation and effect indicators in different aquatic environments. Differences of the freshwater and sea water ecosystems are discussed. A perspective on how to use natural fish parasite populations as biological indicators for climate change is presented, focusing on different problems involved in the freshwater and marine environments.
2 Applied Methodology
The application of fish parasites as bioindicators requires the routine study of fish for the presence of a variety of fish parasites. The parasitological samplings must follow standard protocols. To use fish parasites as effect indicators for environmental change, the study of both protozoans and metazoan parasites is suggested. Consequently, the external examination should be done directly after catch from the living fish when the protozoan parasites are still alive on the body surface. They disappear soon after the death of the fish. Smears for the detection of ectoparasitic trichodinid ciliates must be taken directly from the gills, surface and the inner opercula of the living fish, followed by the study of the skin, fins, eyes, gills, mouth- and gill cavity for other metazoan ectoparasites, such as monogeneans and crustaceans. The inner organs, digestive tract, liver, gall bladder, spleen, kidneys, gonads, heart and swim bladder should be subsequently separated and transferred into saline. While a stereomicroscope must be used to study the internal organs, a phase-contrast microscope or better with Nomarski (DIC = differential interference contrast) enables recognition of parasitic life cycle stages in the gall bladder and urogenital system. Belly flaps and musculature must be examined on a candling table with a light source from beneath.
Most useful fixatives are 4% formalin and 70% ethanol, or absolute ethanol for molecular analyses (see Klimpel and Palm 2011). The smears from the gills, surface and opercula must be stained by using a silver nitrate impregnation after Klein (1926, 1958), rinsed and covered with 5% silver nitrate solution and impregnated for 30 min in the dark. The AgNO3 has to be removed and the slides must be covered with distilled water, exposed to ultraviolet light for 40–50 min, and dried after exposure. Acanthocephala should be transferred to freshwater until the proboscis everts prior to fixation. For identification purposes, Nematoda and Acanthocephala should be dehydrated in a gradated ethanol series and transferred to 100% glycerine (Riemann 1988). Digenea, Monogenea and Cestoda are stained with acetic carmine, dehydrated, cleared with a clearing agent such as eugenol and mounted in Canada balsam (Palm 2000, 2004). Directly fixed nematodes in 100% ethanol can be identified by using molecular tools, such as described for the ascaridoid genus Anisakis (see Palm et al. 2008; Klimpel and Palm 2011).
The parasitological terms (prevalence, intensity and mean intensity) are standardized, and should follow Bush et al. (1997), where the prevalence (P) is the number of fish with one or more individuals of a particular parasite species (or taxonomic group) divided by the number of hosts examined (expressed as a percentage) [Prevalence (P) = No. of hosts infested / No. of hosts examined * 100]. Intensity (of infection, I) is the number of individuals of a particular parasite species in a single host (expressed as a numerical range); and mean intensity (of infection, I m ) is the total number of parasites of a particular species found in a sample divided by the number of infected hosts [Mean Intensity (I m) = Total no. of a particular parasite / No. of infected hosts]. The diversity of the parasite fauna (endoparasite fauna according to Rückert et al. 2009a) should be determined by using the Shannon–Wiener diversity index (H′) and the evenness index (E) of Pielou (Magurran 1988) [Shannon-Wiener Index (H’) = ∑P ilnP i, Evenness (E) = H′/lnS, with H′ being the diversity index, P i the proportion of the individual (ith) species to the total and S is the total number of species in the community (species richness)]. The ecto- versus endoparasite ratio (E/E) has been introduced by Palm and Rückert (2009). Any other ecological measures or parameters might also be applied to compare the different sampling sites.
3 Fish Parasites as Biological Indicators
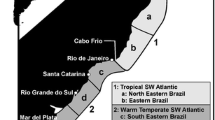
One of the first applications to utilize fish parasites as biological indicators using standard parasitological methodologies (see above) was related to fisheries, including the separation of fish stocks (e.g. Lester 1990; Khan and Tuck 1995; MacKenzie 1990, 2002; Moser 1991; Moser and Hsieh 1992; Williams et al. 1992; Pascual and Hochberg 1996). Herrington et al. (1939) was the first to use the crustacean parasite of a marine fish in the North Atlantic as a biological tag in a population study. The abundance and occurrence of the parasites directly relates to the distribution, migration patterns and population biology of their hosts (Kabata et al. 1987; MacKenzie 1985; Boje et al. 1997; Klimpel et al. 2010), and can be used for stock identification (Fig. 12.1a-b) and even risk assessment of fish food-borne zoonoses. MacKenzie (1983) listed different criteria to decide on the usefulness of parasites as biological indicators that he applied for the Atlantic herring Clupea harengus and the mackerel Scomber scombrus in the North Sea and Northeast Atlantic (MacKenzie 1985, 1987, 1990). Larval trypanorhynch cestodes (Fig. 12.2b) and anisakid nematodes were most useful for stock recognition and separation. It is important that the parasites are easy to collect and can be easily recognized, excluding systematic errors based on erroneous species identification (Table 12.1). Suitable parasites should also infect their hosts at a particular age or in a definite region and are differently abundant in different regions, allowing the original locality of infection to be traced. Anisakis simplex infection of herrings in the Baltic Sea off the coast of Poland has identified the western spring herring that migrates from the North Sea, the origin of the infection, into the Baltic Sea (Grabda 1974). Klimpel et al. (2010) found that migrating myctophids around the South Shetland Islands, Antarctica, were infected with Anisakis simplex C and A. pegreffii, having identical ITS-1, 5.8 S and ITS-2 sequences to specimens from Pacific Canada or California (A. simplex C) and the Mediterranean Sea (A. pegreffii). The authors suggested that the nematodes originated from more northern waters, and became introduced to the Southern Ocean by migrating myctophids and whales. Marine fish parasite larvae have been recorded in freshwater up to 3,000 km away from the coast (Lühe 1910; Shulman 1957). Anadromous fish species such as salmonids transport these parasites upstream, explaining the presence of trypanorhynch cestodes within the German rivers Rhine, Elbe and Weser as well as within a lake in Switzerland (Lühe 1910; Palm 2004). These records provide evidence for the extensive fish migrations from the Ocean also upstream into the freshwater environments.

(a) Scolex of the trypanorhynch cestode Grillotiella exile from Indonesia, with two bothria and armed tentacles (worm length = 0.8 mm). This species has been recorded from the tiger shark (Galeocerdo cuvier), infests the gills of the narrow-barred Spanish mackerel (Scomberomorus commerson) and has been used as a biological tag for stock analyses. (b) Tentacular armature of G. exile from Indonesia. The arrangement of the tentacular hooks is species specific and enables identification (Palm and Klimpel 2007; Palm 2008)
Gibson (1972) separated flounders (Platichtys flesus) from two estuaries and the sea within a 40 mi range. The fish had dissimilar parasite-faunas, and individual fish from each locality could be recognized by its parasites, especially by the level of certain indicator species. Similarly, “foreign” flounders moving into a flounder population could be picked out by the markedly different composition of their parasite-fauna. Using two species of myxozoans, Myxobolus arcticus in the brain and Henneguya salmonicola in the musculature, Margolis (1982) was able to distinguish three different spawning stocks of sockeye salmon, Oncorhynchus nerka, off Vancouver Island, Canada. These stocks came from three different lakes, where the parasites infect the juveniles in freshwater. Juvenile fish from the Great Central Lake were virtually free of these parasites, those from Sproat Lake had only M. arcticus and sockeye from Henderson Lake were infected with both species. These results permitted for the first time in-season management of a fishery (Moser 1991). MacKenzie and Abaunza (1998), MacKenzie (2002) and Lester and MacKenzie (2009) later summarised and discussed the procedures and methods for the stock discrimination of marine fish, and Latama (2006), Lester et al. (2001) and Charters et al. (2010) recently applied these methods for Spanish and grey mackerel (both genus Scombermorus) populations even in the widely distributed stocks off the northern Australian coast and in tropical Indonesia (Fig. 12.1). It was demonstrated that Spanish mackerel from Australia and Indonesia clearly separate, and harbour different metazoan parasites. Four different clusters of Spanish mackerels were found in Sulawesi waters alone, four stocks or populations of grey mackerels around the northern and eastern coastline of Australia, indicating the presence of different local stocks in the areas that do not entirely mix. Such results are of importance for the fishing industry in the attempt to better utilize and sustainably use these commercially important fish stocks.

(a) Diphyllidean cestode from Raja sp., Pelabuhan Ratu, Indonesia. These parasites are highly host specific and can be used for host species identification and possibly phylogeny. (b) Trypanorhynch cestode Lacistorhynchus sp. from Hemipristis elongata, Pelabuhan Ratu, Indonesia. These parasites can be used for stock identification, host abundance, migration, elasmobranch ecology and phylogeny. (c) Tetraphyllidean cestode Scolex pleuronectis from Hawaiian waters. Biological indicators for assessing spatial variation in shark distribution. (d, e) Fish parasitic Cucullanus sp. from Balistapus undulatus and Pro(spiro)camallanus sp. from Gomphosus varius, French Polynesia. These parasites might be useful as effect indicators for environmental change. (f) Opisthaptor of the monogenean Dasyonchocotyle dasyatis from Dasyatis lata, Hawaii. Potential bioindicator for the host distribution
Relevant information also results from the knowledge of parasites that might be transferred into humans via the musculature of fishes from our markets (e.g. Palm and Overstreet 2000). The prevalence and abundance of zoonotic parasites is different among most fish stocks and geographical regions, leading to a different risk resulting from fish food that is meant for human consumption. Petersen et al. (1993) indicated that based on a parasitological survey the risk of acquiring parasitoses from eating raw fish from central Philippine waters is low. Consequently, the abundance of fish parasites can be used as an indicator for the risk of fish food-borne zoonoses. This human health aspect has implications not only for the fishing but also for the tourism industry within a region (e.g. Anisakis spp., see Palm et al. 2008; Klimpel and Palm 2011).
Another interesting use of fish parasites as bioindicators concerns their potential to elucidate aspects of the biology of the host organism (Caira 1990), for example feeding ecology and behaviour. Many metazoan fish parasites are transmitted through the food chain. Because many life cycle stages are long-lived and can maintain themselves over a longer time inside the host, a parasite record integrates over time and has significant advantages to time-consuming stomach content analyses. Parasitological studies of metazoan parasites with a long life span (Table 12.1) combined with an analysis of the stomach contents are useful for ecological studies of fish that are difficult to study in vivo, because they live in inaccessible and extreme environments (Fig. 12.2c, f). For example, Klimpel et al. (2003, 2006a, b) and Palm and Klimpel (2008) analyzed the parasite fauna and feeding ecology of deep-sea fish around the Great Meteor sea mount and the Mid-Atlantic Ridge, and Palm (1999) and Palm et al. (1998, 2007) analyzed the food web in the Southern Ocean. On the other hand, the occurrence of fish parasites associated with specific prey organisms in the host can help to identify the life cycles of the parasite species involved. An analysis of the parasites and the stomach contents of the pearlside Maurolicus muelleri from the Norwegian Deep revealed the typical life cycle of the anisakid nematode Anisakis simplex (Klimpel et al. 2004), and Klöser et al. (1992) and Palm (1999) studied the life cycles of the anisakids Contracaecum spp. and Pseudoterranova decipiens in the Southern Ocean. Palm and Schröder (2001) postulated, from a study of cestode parasites, that deep-water elasmobranchs of the genus Deania can serve as food fish for larger oceanic sharks around the Great Meteor Seamount in the central North Atlantic Ocean.
Finally, fish parasites can be used as biological indicators for host identification and phylogeny (Fig. 12.2a), and its systematic position (Rokicki 1983). Palm (2007), Palm and Klimpel (2007) and Palm et al. (2009) used trypanorhynch cestodes as a model system to better understand the ecology and co-evolutionary history (cumulative evolution) of parasitic life cycles in the ocean. Olson et al. (2010) estimated the original hosts of two primary parasite lineages in the cestode order Trypanorhyncha to be alternatively rajiform batoids and carcharhiniform sharks. This fundamental split provided independent support for rejecting the notion that rays are derived sharks, and supported the most recent molecular phylogenies of the Neoselachii. Beyond the basal split between shark- and ray-inhabiting lineages, no pattern was found to suggest that the trypanorhynchs have closely tracked the evolutionary histories of these host lineages, but instead, it appears that host-switching has been common (Palm et al. 2009) and that the subsequent evolution of the parasites has been ecologically driven primarily through overlap in the niches of their shark and ray hosts. This was possible by relaxed host specificity within the group (Palm and Caira 2008), making these parasites of interest in the analysis of the feeding ecology of their hosts. A number of other fish parasites in the marine ecosystem have high host specificity, living only on one or a few closely related fish species. They have thus developed in close host–parasite co-evolution, and illustrate the evolutionary history and phylogeny of their hosts, as exemplified for anisakid nematodes of the Anisakis simplex species complexes and their whale hosts (Klimpel et al. 2011) and the phylogeny of deep-sea trematodes in fish (Bray et al. 1999).
4 Environmental Indicators
The presence of parasites within the environment often becomes evident after a massive infestation causing clinical signs or leading to mortality of the infected hosts. Such a situation can be combined with biotic or abiotic changes in the environment (Möller 1987), in the application of fish parasites as environmental indicators. Knowledge of the biology of the parasite and its host(s), the host–parasite relationship and the environment can help to detect environmental change (Table 12.1). Particularly long-living species (some digenean trematode, cestode, nematode life cycle stages) provide information on the seasonal migration of their hosts and migration habits of different age groups (feeding area/spawning area, see above, Table 12.2). They, however, also change their abundance in the host if some of their life cycle stages become affected through the disappearance of their intermediate hosts, provoking disappearance of some parasite species under polluted conditions (Fig. 12.2d–e). Consequently, the occurrence of heteroxenous (multi host life cycle) parasites in an area affected by pollution can be related to the number of intermediate hosts at the studied sites (Xinghua 1987; Overstreet et al. 1996). The intermediate hosts may be more sensitive to environmental changes than the parasite, which in the case of endoparasites is buffered from the environment by the host physiology (Paperna and Overstreet 1981). On the other hand, Jeney et al. (2002) related an increasing experimental infection rate with the digenean Rhipidocotyle fennica cercariae in roach from a eutrophic lake contaminated with bleached paper mill effluent and an oligotrophic lake to a decreased resistance of the fish to the parasite. Short-living species combined with a direct life cycle and high reproduction rates (protozoan ectoparasites, monogenean trematodes) may react to the environmental conditions of the host (Lester 1990). Pettersen et al. (2006) demonstrated that aqueous aluminium of 200–260 μg Al/l at pH 5.8 had a negative effect on ectoparasites (elimination of gyrodactylids, decreased abundance of duck mussel glochidia, increased mortality rate in the fish louse), without any apparent negative effect on the fish hosts. Consequently, a proper choice of the parasite or the parasite metrics to be used is the prerequisite for the application of fish parasites as bioindicators for environmental change (Table 12.2).
4.1 Accumulation Bioindicators
Living inside the fish and using the host as food resource, fish parasites closely interact with the metabolisms of the host. Thus, parasite infrapopulations can be affected by changes of the host physiology and substances accumulated with the host´s food. Early studies of Read (1951) and Roberts (1961) with the adult rat tapeworm Hymenolepis diminuta demonstrated the decrease of weight, total tissue carbohydrate and reduced numbers of mature and gravid proglottids after the hosts were exposed to a low carbohydrate diet (cited in Esch et al. 1975). If the host is living within a polluted environment where some pollutants also enter the fish, the concentration of these substances in the immediate surroundings of the parasite can also increase. In such cases, some fish parasites can accumulate pollutants in a much higher concentration as their host organisms, and serve as accumulation indicators (Table 12.2). For example, some acanthocephalans specifically accumulate certain heavy metals in greater amounts than their host, and can be used as accumulation indicators of heavy metal pollution (Sures and Taraschewski 1995; Sures et al. 1994; Sures 2003; Sures and Siddall 2003). Adults of the acanthocephalans Pomphorhynchus laevis and Paratenuisentis ambiguus accumulate lead and cadmium in a greater amount than their hosts (Anguilla anguilla, Leuciscus cephalus, Perca fluviatilis). However, there is some dependence on the parasitic life cycle stage. According to Sures et al. (1994), adults of the acanthocephalan Acanthocephalus lucii accumulated Cd in higher amounts than the larvae. Sures and Reimann (2003) compared the heavy metal concentration of the acanthocephalan Aspersentis megarhynchus with the muscle of the Antarctic rock cod Notothenia coriiceps. Most of the elements were found in significantly higher concentrations in the acanthocephalan than in the muscle of its host. Levels of Ag, Co and Ni in the muscle of N. coriiceps were even below the detection limit, and were only found in the worm. Other metals commonly associated with human activities (e.g. Pb, Cd, Cu) were accumulated to a high degree in the parasite, demonstrating that pollutants of anthropogenic origin are dispersed within this remote, fairly unpolluted environment. Cestodes can also be used as accumulation indicators. For example, lead and cadmium is found in significantly higher concentrations in the tissues of the cestode Monobothrium wageneri than in their fish host tissues (Tinca tinca from Ruhr River). The marine cestode Bothriocephalus scorpii (Cestoda) from Scophthalmus maximus (Gdansk Bay) was found to accumulate these heavy metals especially in the posterior part of the proglottids, while the anterior part revealed the same amounts of the heavy metals as the fish host tissues (Sures et al. 1997). Jirsa et al. (2008) detected a higher amount of Cd, Pb and Zn in Caryophyllaeus laticeps compared with the cyprinid host Chondrostoma nasus in river sites in Austria, and according to Tekin-Özan and Kir (2005), the plerocercoids of Ligula intestinalis provided reliable information on the amount of heavy metals in a freshwater reservoir in Turkey. According to Sures (2004) and Barus et al. (2007), some fish parasitic nematodes such as Anguillicola crassus from eel and Philometra ovata from bream can also have an increased level of heavy metal concentration compared to their host organisms, and Ruus et al. (2001) recorded accumulation of another environmental contaminant, lindane, in metacercariae of the trematode Bucephaloides gracilescens in the central nervous system of Myoxocephalus scorpius.
4.2 Effect Bioindicators
One of the broadest applications of fish parasites is their use as effect bioindicators. A summary of effects combined with different environmental factors is given in Table 12.2, and was reviewed by Williams and MacKenzie (2003). Protozoan mobiline peritrichous ciliates (genus Trichodina, phylum Ciliophora) have been experimentally examined for their function as biological indicators of water quality (Voigt 1993). They were also reported as indicators for petroleum hydrocarbons (Khan and Thulin 1991). Trichodina cottidarum and T. saintjohnsi infected Myoxocephalus octodecemspinosus that had been exposed to crude oil in laboratory experiments, as well as at an oil-receiving terminal site, in a significantly higher intensity than in uncontaminated control experiments/sites. Pulp and paper mill effluents showed similar effects on these trichodinid ciliates (Khan 1990; Khan et al. 1993). Also oil-contaminated sediments caused increasing infestation with Trichodina sp. in M. octodecemspinosus after exposure over a time of 3 and 6 months. The prevalence of Trichodina spp. gill infection increased significantly on fish from inside the heavily polluted local fishing port Pelabuhan Ratu on the southern Java coast, Indonesia, compared to specimens that were caught outside the harbour (Epinephelus sp., 45.5%, n = 11 vs. 0%, n = 23). Palm and Dobberstein (1999) suggested the use of prevalence and density data of Trichodina spp. as a biological indicator for the bacterial biomass in the environment, because peritrichous ciliates are primarily filter feeders on small algae and bacteria. The first record of Trichodina rectuncinata from the gills of Melichthys vidua in Moorea, French Polynesia, might be connected to the degraded reef habitat where the fish was caught (Fig. 12.3). Ogut and Palm (2005) demonstrated the relationship between Trichodina spp. infection on Merlangius merlangus and organic pollution, measured as levels of nitrite, nitrate and phosphate in the surrounding environment. These relationships suggest the usefulness of fish parasites as biological indicators for ecosystem change (see below; Palm and Rückert 2009; Rückert et al. 2009a).

Trichodina rectuncinata from the gills of Melichthys vidua from Moorea, French Polynesia. (a-c) Silver nitrate impregnation after Klein. (d, e) Living specimens on the gills. The prevalence of infestation with trichodinid ciliates is related to the bacterial biomass in the water body, and can be used as an eutrophication effect indicator. The above specimens were found on a single fish from a degraded reef in Oponohu Bay
Also other pollutants can have effects on the parasites within or attached to the host. For example exposure to petroleum aromatic hydrocarbons (PAH) caused increasing prevalence of Ceratomyxa acadiensis (Myxosporida) in the gall bladder of the winter flounder Pseudopleuronectes americanus (Khan 1986, cited in Khan and Thulin 1991). PAHs are known to cause lesions and hyperplaisia of the secondary gill lamellae in fish. Long-time exposure of cod to aromatic hydrocarbons (max. 30 weeks) caused increasing infestation with monogenetic flatworms (prevalence as well as intensity), which was attributed to the already damaged gill tissue, and therefore to the better living conditions for monogenetic trematodes (Khan and Kiceniuk 1988). Bayoumy et al. (2008) showed conflicting evidence for the reaction of monogeneans to heavy metal concentrations in the Red Sea. While the lead concentration correlated negatively with the prevalence of five different monogenean species, nickel and also temperature were positively correlated. Gheorghiu et al. (2006) reported improved parasite Gyrodactylus turnbulli (Monogenea) population growth under water-borne zinc concentration up to 120 lg/L, while Gheorghiu et al. (2007) indicated reduced parasite survival and reduced reproduction over this range of Zn concentrations, rejecting the suggestion that water-borne Zn improves parasite growth and reproduction. These examples demonstrate the difficulties involved in finding the best host–parasite systems as effect bioindicators for pollutants in aquatic environments.
Parasitic endohelminths with a heteroxenous life cycle can be more tolerant to an increased level of water pollutants (see accumulation indicators above). However, an effect on the intermediate hosts prevents parasite transmission and reduces the observed prevalence and intensity of infection in the definite host. For example, the gastrointestinal parasites of Pseudopleuronectes americanus (Steringophorus furciger; Trematoda) and Gadus morhua (Echinorhynchus gadi; Acanthocephala) exposed to oil had a lower infection compared to the control groups. It was thought that water-soluble fractions of the crude oil were especially involved in causing this effect (Khan and Kiceniuk 1983; Khan 1987).
Another interesting biological-effect monitoring study was carried out in the context of eutrophication, in both freshwater and marine habitats. Hartmann and Nümann (1977) reported an increasing level of infection with parasites of various species, especially Diplostomum sp., with progressive eutrophication in Lake Constance. According to Lafferty (2008), parasites that increase under eutrophic conditions tend to be host generalists with local recruitment. Athanassopoulou and Ragias (1998) studied the parasites and diseases of wild fish populations from a lake in Greece over two long periods (1984–1990 and 1994–1997). The study concluded that the pollution in the lake increased together with the prevalence, intensity and pathology of most parasites. Tumours and bacterial infections were directly associated with increased pollution, especially industrial and sewage waste values. In the Baltic Sea, Reimer (1995) used parasites of piscean and other hosts as eutrophication indicators. The different infestation of gobiid fish with helminth parasites from different sampling sites in the western Baltic Sea was associated with the eutrophication level, influencing the abundance of herbivorous intermediate hosts (Zander and Kesting 1996, 1998). Kesting and Zander (2000) related the impoverishment of the metazoan parasite fauna in the Schlei Fjord over an 18-year period to increasing eutrophication. In the marine environment Moser and Cowen (1991) studied the nototheniid Trematomus bernacchi from the eutrophic east and oligotrophic west side of McMurdo Sound in Antarctica. The fish from eutrophic localities revealed high prevalences of the acanthocephalan Echinorhynchus sp., the nematode Ascarophis nototheniae, the trematode Dinosoma sp. and the cestode Phyllobothrium sp. The investigation of the abundance of arthropods at these localities demonstrated that these are more abundant at the eutrophic east side of McMurdo Sound. The higher number of available intermediate hosts of these parasites in the region triggers their prevalence in the fish as the definitive host (Moser and Cowen 1991).
Environmental monitoring programs using fish parasites in open water have not been applied, though preliminary data provide promising results. Recently, Lafferty et al. (2008) proposed that the sampling of larval cestodes in small teleosts (Fig. 12.2c) is a convenient method for assessing spatial variation in shark distribution, and that the lower parasitism at Kiritimati compared to Palmyra in the Central Pacific Ocean resulted from a simplified food web due to overfishing. Low biodiversity could impair parasite transmission by reducing the availability of hosts required by parasites with complex life cycles. This would imply that these fish parasites can be used to monitor food web composition and elasmobranch abundance in oceanic habitats. The latter is probably influenced by shark fisheries, significantly reducing the number of available oceanic top predators over time. This result supports the notion by Marcogliese (2005) that perturbations in ecosystem structure and function that affect food web topology also impacts upon parasite transmission, thus affecting parasite species abundance and composition. As such, parasite populations and communities are useful indicators of environmental stress, food web structure and biodiversity.
4.3 Ecosystem Bioindicators
In marine coastal areas, human activities directly influence living communities, which may result in a heavier parasite infestation compared to less polluted sites (Skinner 1982). On the other hand, some parasite species can disappear, and thus reduce parasite diversity within the system (see above). While it remains difficult to find the best host–parasite system that can indicate a specific parameter change, MacKenzie (1999) proposed the use of fish parasites as an early warning system for pollution and environmental change. With the identification of particular parasite species whose ecology is so delicately balanced, that environmental changes may lead to transmission failure, regular studies can monitor and indicate environmental change much earlier than using other methods. He suggested that infections with endoparasitic helminths tend to increase while those of ectoparasitic heminths tend to decrease with increasing levels of pollution. This would allow the use of entire parasite communities and their changing infection levels as environmental bioindicators (also see Marcogliese 2005).
To date, ecological aspects of infestations by parasites and their relationship to anthropogenic factors such as urban pollution, overfishing and fish-farming (mariculture) are only just beginning to be understood. Recent studies demonstrate, however, the relatedness of parasite abundance and the environmental condition. Sasal et al. (2007) utilized the entire parasite community (as larval Cestoda, adult Crustacea, larval and adult Digenea, larval Nematoda) of apogonid fish to detect anthropogenic influences (urban and industrial pollution) on two coral reef lagoons in New Caledonia. Pech et al. (2009) used fish physiological biomarkers and the parasite infracommunity characteristics to evaluate the effects of chemical pollutants on the fish host and in the environment. Rückert et al. (2009a) applied three different parasite parameters to describe the environmental situation in Segara Anakan lagoon, an extensive mangrove ecosystem in Indonesia, and Rückert et al. (2009b, 2010) compared free living Epinephelus coioides and E. fuscoguttatus with specimens from mariculture farms. The most striking result was reduced parasite diversity within the lagoon ecosystem compared with outside the lagoon, probably linked to the changing hydrographic conditions and not eutrophication. This would follow Hudson et al. (2006) who considered a healthy system to be one that is rich in parasite species. According to Hechinger et al. (2007) and Lafferty (2008), trematode diversity and prevalence in snails correlates with macro-invertebrate diversity and abundance. This may result from birds being attracted to areas with diverse invertebrate prey communities (particularly if the invertebrates present also serve as intermediate hosts for trematodes). By the same reasoning, trematodes in snails may indicate aspects of the fish community. Lafferty (1997) and Marcogliese (2005) argued that helminth infracommunity patterns are useful tools to assess the impact of anthropogenic pollution on ecosystems. Among others, Oros and Hanzelová (2009) demonstrated the re-establishment of the fish parasite fauna in a Slovakian river system after a catastrophic pollution event. Beside total abundance and prevalence of specific parasite species or their life cycle stages, parasite metrics (Diamant et al. 1999) such as the Shannon-Wiener diversity index as calculated for the endoparasites and the ratio of ecto- vs. endoparasites are suitable measures to compare the parasite community in different aquatic systems (Rückert et al. 2009a).
Palm and Rückert (2009) developed a new method to visualize environmental differences between habitats, among them a grouper mariculture facility in the Thousand Islands, Indonesia. A star graph was used to visualize the parasite composition for the different fishes, using (1) the prevalence of trichodinid ciliates, (2) the ecto-/endoparasite ratio and (3) the endoparasite diversity as bioindicators. This was found a suitable method to visualize and monitor environmental health under high parasite biodiversity conditions in tropical ecosystems, and to better communicate the scientific results to involved stake holders and decision makers. Palm et al. (2011) studied fish from floating net cages of a commercially run mariculture facility after its opening in 2001 for six consecutive years. Tetraphyllidean larvae Scolex pleuronectis (Fig. 12.2c) and the nematodes Terranova sp. and Raphidascaris sp. 1 were highly abundant in 2003/04–2005/06 (max. prevalence S. pleuronectis 40%, Terranova sp. 57%, Raphidascaris sp. 1 100%, Fig. 12.4a), and drastically reduced subsequently (up until 2008/09) (Fig. 12.4b). These parasites, especially the nematodes (e.g. Fig. 12.2d, e), together with the above parasite metrics illustrate a significant change in holding conditions over the years. This can be either referred to a definite change in management methods (such as feed use and fish treatment) or more likely to a transition of a relatively undisturbed marine environment into a more effected habitat caused by the anthropogenic activities.

(a-b) Stargraph with parasitological metrics to monitor cultured groupers (Epinephelus fuscoguttatus) from a mariculture farm. The parasite infection of the sampled groupers from the same net cages in the rainy seasons 2003/4 and 2008/9 was significantly different (note the change with black arrows), indicating different holding conditions, feed use or environmental change (see Palm et al. 2011)
5 Fish Parasites as Bioindicators for Climate Change
Global climate change with rising sea-levels and water temperatures that may result in changes in (e.g.) ocean circulation and a decrease in salinity may also cause measurable effects on fish parasite composition and biogeography. According to Harvell et al. (2002), climate warming can affect host–pathogen interactions by (i) increasing pathogen development rates, transmission and number of generation times per year (ii) raising the overwinter survival rate of the pathogen and (iii) increasing the host susceptibility to thermal stressors. More importantly, increasing temperatures alter the seasonality and biogeographical range of many species, including the hosts and the parasites. Consequently, with the potential to indicate environmental change and host migration or pollution, fish parasites as an important component of the aquatic biodiversity may well be useful as bioindicators of climate change. This application must utilize fish parasites that are widely distributed, easily monitored (Table 12.1), and can indicate long-term changes within the aquatic environment, being related either to natural or anthropogenic influenced environmental conditions.
MacKenzie (1987) recorded changes in the prevalence of metazoan parasites, the trypanorhynch cestodes Grillotia angeli Dollfus, 1969 in mackerel (Scomber scombrus) and Lacistorhynchus sp. in herring (Clupea harengus) over the periods of 8 years (1978–1985) and 11 years (1974–1984), respectively (e.g. Fig. 12.1b). Both data sets showed sharp decreases in parasite prevalence from periods at relatively high levels to others at much lower levels. The changes in prevalence occurred at the same time in both host–parasite systems and coincided with the end of a hydrographic phenomenon known as the mid-70s salinity anomaly, with a salinity reduction in the upper 1,000 m of water over most parts of the eastern Atlantic. The author explained these results by invoking changes in the abundance of first intermediate and definitive hosts, host diet and variations in year class strength, and changes in hydrographic conditions. Rising sea-water temperatures and a possible change in currents and resulting hydrography must have consequences for parasite composition and abundance. Poulin (2007) emphasized the importance of latitude as a convenient surrogate measure for temperature. As a rule, fish parasite diversity increases from higher latitudes towards the tropics. Among marine fish, the mean species richness of ectoparasites per fish population sampled increases with increasing water temperature towards lower latitudes, a pattern that is not detectable for endoparasitic helminths. Among freshwater fish species, species richness of parasites seems to peak at temperate latitudes, and not in the tropics. However, as stated by Vidal-Martínez et al. (2010), the exact circumstances under which parasites can be used as indicators of environmental impact or change remain difficult, especially at the large scale of consideration. In coastal regions with a variety of different underlying factors such as river-based pollution, salinity change, industrial impact, sewage and other anthropogenic waste, or in intensive fisheries, climate change-related abnormalities in the fish parasite fauna will be difficult to assess. Consequently, such studies should be carried out in remote environments such as the Southern or Arctic Oceans, the open oceanic waters or the deep-sea, where recorded effects can be better linked to a change of the environment at a larger scale and not to local variation.
There is no doubt that climate is decisive for a variety of ecological processes, concerning the individual, populations and species biogeography and composition (e.g. Mouritsen and Poulin 2002; Mouritsen et al. 2005). Cattadori et al. (2005) concluded that specific climatic events may lead to outbreaks of infectious diseases or pests that may cause dramatic, synchronized changes in the abundance of their hosts. The size of red grouse populations in northern England either increased or decreased in synchrony to correlated climatic conditions during May and July, influencing the density-dependent transmission of the gastrointestinal nematode Trichostrongylus tenuis, a parasite that reduces grouse fecundity. This in turn forced both, the grouse and the parasite populations into synchrony. Temperature is a key factor that regulates such processes, determining species distribution and occurrence also in aquatic habitats. Because freshwater and marine environments are generally different in terms of their size, stability and physical conditions, these systems must be treated differently. While the observation of climate change-related scenarios in the large marine ecosystems may be possible (see above) and should be dealt with in future studies, it has to be questioned how small-scale parasitological data from freshwater environments can be unequivocally related to climate change by temperature increase. For example, Esch et al. (1976) related epizootic outbreaks of the facultative episymbiont Epistylis sp. and Aeromonas hydrophila in centrarchid fish in some freshwater reservoirs to a combination of eutrophication and rise of temperature. However, Kennedy et al. (2001) demonstrated that the larval cestode Ligula intestinalis (L., 1758) together with the roach Rutilus rutilus regulated the size of the other fish populations inhabiting that lake. Thus, neither temperature nor other environmental factors but this parasite–host system consisting of two generalistic, widely distributed species was the driving factor for the fish and parasite population structure within the entire ecosystem. It must be kept in mind that according to Bagge et al. (2004), the total fish population size of the crucian carp Carassius carassius alone directly influenced the mean number of parasite (monogenean) species per fish and the mean total abundance. Other factors such as the fish population density, mean fish length per pond, number of fish examined per pond, distance to the nearest lake, and several water quality measures did not influence the monogenean infection within that system.
6 Conclusions
Fish parasites can elucidate problems in a variety of applications. They can be used as biological indicators for their hosts, accumulation indicators to detect pollutants, impact indicators to describe the relationship between for example pollution or eutrophication and the parasite, and finally as systemic indicators that provide information on the health status of the environment. A proper selection of the parasites utilized and their specific life cycle stages is a prerequisite for all these applications, and requires careful planning and some background information on the parasite diversity within each habitat. Sixteen different criteria have been formulated to support selection of the best parasite indicator species for any particular purpose (Table 12.1).
The published literature demonstrates that parasites can be successfully used as biological indicators in both freshwater and marine environments. Protozoan and metazoan parasites likewise are useful, though they react differently according to their needs and characteristic life cycle biology. Trichodinid ciliates react positively to environmental pollutants while most metazoan parasites with their indirect life cycles, requiring the availability of invertebrate intermediate hosts, are negatively affected (Table 12.2). The widest range of applications has been demonstrated for the heteroxenous digeneans, cestodes, nematodes and acanthocephalans that can serve as biological, accumulation and impact indicators. Though several open questions still remain, fish parasites are ready to be used as indicator organisms for environmental change, and should be included in regular monitoring programs that will provide long-term data sets. In combination with other indicator measures, fish parasites will enable a better estimate of the causes and consequences that underlie environmental change in aquatic habitats.
While there can be no doubt that particular fish parasite species and metrics describe the environmental conditions in the aquatic realm (impact/ecosystem bioindicators), the use of fish parasites as biological indicators of climate change is more unclear. Distinction must be made between freshwater, coastal and oceanic environments, with systemic differences in biodiversity, stability and already existing anthropogenic influences. There is practically no freshwater habitat under pristine conditions (e.g. most of anthropogenic origin in the state North Rhine-Westfalia), and anthropogenic species introduction, migrations of neozoons connected to fisheries and habitat change combined with natural migration events already altered the regular fish and parasite fauna. Many freshwater parasite species are opportunistic generalists that can infect a variety of different fish hosts, use reservoir habitats, and underlie small-scale variability according to the ecological conditions in the freshwater systems. This can explain the conflicting evidence provided by Kennedy (1997) who considered eel parasites in a freshwater environment to be useful to explain observed environmental change. However, he stated that in his case the helminth communities provide no clear indication of the nature of these changes. This contrasted with his results in an earlier study (Kennedy et al. 1994). A change in species diversity and composition at a sampling site does not necessarily result from slight temperature increase over a long time period, but merely from a shift in intermediate and final host abundances at a small scale. The monogenean species richness and abundance in a freshwater lake system was directly dependent on the total fish population size (Bagge et al. 2004). Consequently it will be difficult to directly relate the observed parasite infection levels in freshwater ecosystems to changing parameters at a large scale such as under climate change, and not to local factors or naturally/anthropogenic caused species invasion events.
With high population density along the coasts and significant anthropogenic stress in terms of pollution and environmental degradation, fish parasites can describe the environmental conditions of their habitat, such as a tropical lagoon ecosystem (e.g. Rückert et al. 2009a). However, like in freshwater habitats, it will be difficult to relate parasite abundances with effects that are due to climate change and not to local factors. Marcogliese (2008) stated that though much evidence suggests that parasite and disease transmission, and possibly virulence, will increase with global warming, these effects will be superimposed on a multitude of other anthropogenic environmental changes. On the other hand, Poulin and Mouritsen (2006) developed a model to predict that rising sea surface temperatures in the Wadden Sea will lead to regular local extinction events in amphipods, affecting the broader intertidal system. Lafferty and Kuris (2005) argued that aquatic helminths vary in their optimal temperature, making it impossible to make a general prediction about the effect of warming. Single parasite species models as predictors of possible multidimensional environmental change scenarios, considering entire communities are still to be tested within the aquatic habitat.
Finally, oceanic and remote marine ecosystems such as the central Pacific, mid Atlantic, Antarctic or the Polar Sea are the best candidate localities to link an effect of a changing climate directly to aquatic parasite communities at a larger scale. These regions are less affected by factors such as anthropogenic-influenced species introduction, pollutants and for example seasonal migrations. Such studies are entirely missing, and long-term parasitological monitoring programs in these regions have not been done. Melting sea ice and increasing ice-free areas in the Southern and Arctic Oceans open up new habitats that can be newly explored for existing parasite populations. Poulin (2007) concluded that more emphasis should be placed on the geographical scale in studies of parasite communities of fish, and a little less on local scales, to assess spatial variation on a level relevant for the sort of changes ahead. It is suggested that such data sets should be collected in future studies, in order to assess the use of fish parasites as bioindicators for long-term or global climate change.
References
Athanassopoulou F, Ragias V (1998) Disease investigations on wild fish from a polluted lake in northern Greece. Bull Eur Ass Fish Pathol 18:105–108
Bagge AM, Poulin R, Valtonen ET (2004) Fish population size, and not density, as the determining factor of parasite infection: a case study. Parasitology 128:305–313
Baruš V, Jarkovský J, Prokeš M (2007) Philometra ovata (Nematoda: Philometroidea): a potential sentinel species of heavy metal accumulation. Parasitol Res 100:929–933
Baum JK, Worm B (2009) Cascading top-down effects of changing oceanic predator abundances. J Anim Ecol. doi:10.1111/j.1365-2656.2009.01531.x
Bayoumy EM, Osman HAM, El-Bana LF, Hassanain MA (2008) Monogenean parasites as bioindicators for heavy metal status in some Egyptian Red Sea fishes. Global Vet 2:117–122
Boje J, Riget F, Køie M (1997) Helminth parasites as biological tags in population studies of Greenland halibut (Reinhardtius hippoglossoides (Walbaum)), in the north-west Atlantic. ICES J Mar Sci 54:886–895
Bray RA, Littlewood DTJ, Herniou EA, Williams B, Henderson RE (1999) Digenean parasites of deep-sea teleosts: a review and case studies of intrageneric phylogenies. Parasitology 119:S125–S145
Bush AO, Lafferty KH, Lotz JM, Shostak AW (1997) Parasitology meets ecology on its own terms: Margolis et al. revisited. J Parasitol 83:575–583
Caira JN (1990) Metazoan parasites as indicators of elasmobranch biology. NOAA Tech Rep NMFS 90:71–96
Cattadori IM, Haydon DT, Hudson PJ (2005) Parasites and climate synchronize red grouse populations. Nature 433:737–741
Charters RA, Lester RJG, Buckworth RC, Newman SJ, Ovenden JR, Broderick D, Kravchuk O, Ballagh A, Welch DJ (2010) The stock structure of grey mackerel Scomberomorus semifasciatus in Australia as inferred from its parasite fauna. Fisher Res 101:94–99
Diamant A, Banet A, Paperna I, von Westernhagen H, Broeg K, Kruener G, Koerting W, Zander S (1999) The use of fish metabolic, pathological and parasitological indices in pollution monitoring. II The Red Sea and Mediterranean. Helgol Mar Res 53:195–208
Esch GW, Gibbons W, Bourque JE (1975) An analysis of the relationship between stress and parasitism. Am Midl Nat 93:339–353
Esch GW, Hazen TC, Dimock RV, Gibbons W (1976) Thermal effluent and the epizootiology of the ciliate Epistylis and the bacterium Aeromonas in association with centrachid fish. Trans Am Micro Soc 95:687–693
Froese R, Pauly D (2010) FishBase. World Wide Web electronic publication. www.Fishbase.org, version (01/2010)
Galli P, Crosa G, Mariniello L, Ortis M, D’Amelio S (2001) Water quality as a determinant of the composition of fish parasite communities. Hydrobiologia 452:173–179
Gheorghiu C, Marcogliese DJ, Scott ME (2006) Concentration dependent effects of waterborne zinc on population dynamics of Gyrodactylus turnbulli (Monogenea) on isolated guppies (Poecilia reticulata). Parasitology 132:225–232
Gheorghiu C, Cable J, Marcogliese DJ, Scott ME (2007) Effects of waterborne zinc on reproduction, survival and morphometrics of Gyrodactylus turnbulli (Monogenea) on guppies (Poecilia reticulata). Int J Parasitol 37:375–381
Gibson DI (1972) Flounder parasites as biological tags. J Fish Biol 4:1–9
Grabda J (1974) The dynamics of the nematode larvae, Anisakis simplex (Rud.) invasion in the south-western Baltic herring (Clupea harengus L.). Acta Ichthyolog Piscator 4:3–21
Hartmann J, Nümann W (1977) Percids of lake constance, a lake undergoing eutrophication. J Fish Res Bd Can 34:1670–1677
Harvell CD, Mitchell CE, Ward JR, Altizer S, Dobson AP, Ostfeld RS, Samuel MD (2002) Climate warming and disease risks for terrestrial and marine biota. Science 296:2158–2162 doi:10.1126/science.1063699
Hechinger RF, Lafferty KD, Huspeni TC, Brooks A, Kuris AM (2007) Can parasites be indicators of free-living diversity? Relationships between species richness and the abundance of larval trematodes and of local fishes and benthos. Oecologia 151:82–92
Herrington WC, Bearse HM, Firth FE (1939) Observations on the life history, occurrence and distribution of the redfish parasite Sphyrion lumpi. U.S Bur Fish Spec Rep No. 5:1–18
Hudson PJ, Dobson AP, Lafferty KD (2006) Is a healthy ecosystem one that is rich in parasites? Trends Ecol Evol 21:381–385
Hutchings JA, Baum JK (2005) Measuring marine fish biodiversity: temporal changes in abundance, life history and demography. Philos Trans R Soc Lond B 360:315–338
Jeney Z, Valtonen ET, Jeney G, Jokinen EI (2002) Effect of pulp and paper mill effluent (BKME) on physiological parameters of roach (Rutilus rutilus) infected by the digenean Rhipidocotyle fennica. Folia Parasit 49:103–108
Jirsa F, Leodolter-Dvorak M, Krachler R, Frank C (2008) Heavy metals in the nase, Chondrostoma nasus (L. 1758), and its intestinal parasite Caryophyllaeus laticeps (Pallas 1781) from Austrian rivers: Bioindicative aspects. Arch Environ Contam Toxicol. doi:10.1007/s00244-008-9154-1
Kabata Z, McFarlane GA, Whitaker DJ (1987) Trematoda of sablefish, Anoplopoma fimbria (Pallas, 1811), as possible biological tags for stock identification. Can J Zool 66:195–200
Kennedy CR (1997) Long-term and seasonal changes in composition and richness of intestinal helminth communities in eels Anguilla anguilla of an isolated English river. Folia Parasit 44:267–273
Kennedy CR, Shears PC, Shears JA (2001) Long-term dynamics of Ligula intestinalis and roach Rutilus rutilus: a study of three epizootic cycles over thirty-one years. Parasitology 123:257–269
Kennedy CR, Wyatt RJ, Starr K (1994) The decline and natural recovery of an unmanaged coarse fishery in relation to changes in land use and attendant eutrophication. In: Cowes IG (ed) Rehabilitation of freshwater fisheries. Blackwell, Oxford, pp 366–375
Kesting V, Zander CD (2000) Alteration of the metazoan parasite faunas in the brackish Schlei Fjord (Northern Germany, Baltic Sea). Int Rev Hydrobiol 85:325–340
Khan RA (1987) Crude oil and parasites of fish. Parasitol Tod 3:99–100
Khan RA (1990) Parasitism in marine fish after chronic exposure to petroleum hydrocarbons in the laboratory and to the Exxon Valdez oil spill. Bull Environ Contam Toxicol 44:759–763
Khan RA, Kiceniuk J (1983) Effects of crude oils on the gastrointestinal parasites of two species of marine fish. J Wildl Dis 19:253–258
Khan RA, Kiceniuk J (1988) Effect of Petroleum Aromatic Hydrocarbons on monogeneids parasitising Atlantic cod, Gadus morhua L. Bull Environ Contam Toxicol 41:94–100
Khan RA, Thulin J (1991) Influence of pollution on parasites of aquatic animals. Adv Parasit 30:201–239
Khan RA, Tuck C (1995) Parasites as biological indicators of stocks of Atlantic cod (Gadus morhua) off Newfoundland, Canada. Can J Fish Aquat Sci 52(suppl 1):195–201
Khan RA, Baker DE, Williams-Ryan K, Hooper RG (1993) Influence of crude oil and pulp and paper mill effluent on mixed infections of Trichodina cottidarium and T. saintjohnsi (Ciliophora) parasitising Myoxocephalus octodecemspinosus and M. scorpius. Can J Zool 72:247–251
Klein B (1926) Ergebnisse mit einer Silbermethode bei Ciliaten. Arch Protistenkd 56:243–279
Klein B (1958) The “dry” silver method and its proper use. J Protozool 5:99–103
Klimpel S, Palm HW (2011) Anisakid nematode (Ascaridoidea) life cycles and distribution: Increasing zoonotic potential in the time of climate change? In: H. Mehlhorn (ed.) Progress in Parasitology, Parasitology Research Monographs, Springer Verlag
Klimpel S, Seehagen A, Palm HW, Rosenthal H (2001) Deep-water metazoan fish parasites of the world. Logos Verlag, Berlin
Klimpel S, Palm HW, Seehagen A (2003) Metazoan parasites and food composition of juvenile Etmopterus spinax (L., 1758) (Dalatiidae, Squaliformes) from the Norwegian Deep. Parasitol Res 89:245–251
Klimpel S, Palm HW, Rückert S, Piatkowski U (2004) The life cycle of Anisakis simplex in the Norwegian Deep (northern North Sea). Parasitol Res 94:1–9
Klimpel S, Palm HW, Busch MW, Kellermanns E, Rückert S (2006a) Fish parasites in the Arctic deep-sea: poor diversity in pelagial fish species vs. heavy parasite load in a demersal fish. Deep Sea Res Part I 53:1167–1181
Klimpel S, Rückert S, Piatkowski U, Palm HW, Hanel R (2006b) Diet and metazoan parasites of silver scabbard fish, Lepidopus caudatus, from the Great Meteor Seamount (North Atlantic). Mar Ecol Prog Ser 315:249–257
Klimpel S, Busch MW, Kellermanns E, Kleinertz S, Palm HW (2009) Metazoan deep-sea fish parasites. Acta Biologica Benrodis, Suppl. 11, Verlag Natur & Wissenschaft, Solingen
Klimpel S, Busch MW, Kuhn T, Rohde A, Palm HW (2010) The Anisakis simplex complex off the South Shetland Islands (Antarctica): endemic populations versus introduction through migratory hosts. Mar Ecol Prog Ser 403:1–11
Klöser H, Plötz J, Palm HW, Bartsch A, Hubold G (1992) Adjustment of anisakid nematode life cycles to the high Antarctic food web as shown by Contracaecum radiatum and C. osculatum in the Weddell Sea. Ant Sci 4:171–178
Lafferty KD (1997) Environmental parasitology: what can parasites tell us about human impacts on the environment? Parasitol Today 13:251–255
Lafferty KD (2008) Ecosystem consequences of fish parasites. J Fish Biol 73:2083–2093
Lafferty KD, Kuris AM (2005) Parasitism and environmental disturbances. In: Thomas F, Renaud F, Guégan J-F (eds) Parasitism & ecosystems. Oxford University Press, Oxford, pp 113–123
Lafferty KD, Shaw JC, Kuris AM (2008) Reef fishes have higher parasite richness at unfished Palmyra Atoll compared to fished Kiritimati Island. Ecohealth 5:338–345
Landsberg JH, Blakesley BA, Reese RO, McRae G, Forstchen PR (1998) Parasites of fish as indicators of environmental stress. Env Monit Assess 51:211–232
Latama G (2006) Metazoan parasites from the narrow-barred Spanish mackerel, Scomberomorus commerson (Lacepède, 1800) around Sulawesi waters. Dissertation, Bogor Agricultural University, Bogor (In Indonesian)
Lester RJG (1990) Reappraisal of the use of parasites for stock identification. Aust J Mar Freshw Res 41:855–864
Lester RJG, MacKenzie K (2009) The use and abuse of parasites as stock markers for fish. Fisher Res 97:1–2
Lester RJG, Thompson C, Moss H, Barker SC (2001) Movement and stock structure of the narrow-barred Spanish mackerel as indicated by parasites. J Fish Biol 59:833–842
Lühe M (1910) Parasitische Plattwuermer. II: Cestodes. Die Süsswasserfauna Deutschlands. Eine Exkursionsfauna, Jena, pp 1–153
MacKenzie K (1983) Parasites as biological tags in fish population studies. Adv Appl Biol 7:251–331
MacKenzie K (1985) The use of parasites as biological tags in population studies of herring (Clupea harengus L.) in the North Sea and to north and west of Scotland. J Int Counc Explor Sea 42:33–64
MacKenzie K (1987) Long-term changes in the prevalence of two helminth parasites (Cestoda: Trypanorhyncha) infecting marine fish. J Fish Biol 31:83–87
MacKenzie K (1990) Cestode parasites as biological tags for mackerel (Scomber scombrus L.) in the northeast Atlantic. J Int Counc Explor Sea 46:155–166
MacKenzie K (1999) Parasites as pollution indicators in marine ecosystems: a proposed early warning system. Mar Poll Bull 38:955–959
MacKenzie K (2002) Parasites as biological tags in fish population studies. An update. Parasitology 124:S153–S163
MacKenzie K, Abaunza P (1998) Parasites as biological tags for stock discimination of marine fish: a guide to procedures and methods. Fish Res 38:45–56
MacKenzie K, Williams HH, Williams B, McVicar AH, Siddall RI (1995) Parasites as indicators of water quality and the potential use of helminth transmission in marine pollution studies. Adv Parasitol 35:86–245
Magurran AE (1988) Ecological diversity and its measurement. Croom Helm, London
Marcogliese DJ (2005) Parasites of the superorganism: Are they indicators of ecosystem health? Int J Parasitol 35:705–716
Marcogliese DJ (2008) The impact of climate change on the parasites and infectious diseases of aquatic animals. Rev Sci Tech 27:467–484
Marcogliese DJ, Cone DK (1997) Parasite communities as indicators of ecosystem stress. Parassitologia 39:27–232
Margolis L (1982) Pacific salmon and their parasites. A century of study. Bull Can Soc Zool 13:7–11
Margolis L, Arthur JR (1979) Synopsis of the parasites of fishes of Canada. Bull Fish Res Bd Can 199:1–269
McDonald TE, Margolis L (1995) Synopsis of the parasites of fishes of Canada: Supplement (1978–1993). Can Spec Publ Fish Aquat Sci 122:1–265
Möller H (1987) Pollution and parasitism in the aquatic environment. Int J Parasitol 17:353–361
Moser M (1991) Parasites as biological tags. Parasitol Tod 7:182–185
Moser M, Cowen K (1991) The effects of periodic eutrophication on parasitism and stock identification of Trematomus bernacchii (Pisces: Nototheniidae) in McMurdo Sound, Antarctica. J Parasitol 77:551–556
Moser M, Hsieh J (1992) Biological tags for stock separation in Pacific herring Clupea harengus pallasi in California. J Parasitol 78:54–60
Mouritsen KN, Poulin R (2002) Parasitism, climate oscillations and the structure of natural communities. Oikos 97:462–468
Mouritsen KN, Tompkins DM, Poulin R (2005) Climate warming may cause a parasite-induced collapse in coastal amphipod populations. Oecologia 146:476–483
Ogut H, Palm HW (2005) Seasonal dynamics of Trichodina spp. on whiting (Merlangius merlangus) in relation to organic pollution on the eastern Black Sea coast of Turkey. Parasitol Res 96:149–153
Olson PD, Caira JN, Jensen K, Overstreet RM, Palm HW, Beveridge I (2010) Evolution of the trypanorhynch tapeworms: Parasite phylogeny supports independent lineages of sharks and rays. Int J Parasitol 40:223–242
Oros M, Hanzelová V (2009) Re-establishment of the fish parasite fauna in the Tisa River system (Slovakia) after a catastrophic pollution event. Parasitol Res 104:1497–1506
Overstreet RM (1997) Parasitological data as monitors of environmental health. Parassitologia 39:169–175
Overstreet RM, Hawkins WE, Deardorff TL (1996) The western mosquitofish as an environmental sentinel: parasites and histological lesions. In: Servos MR, Munkttrick KR, Carey JH, van der Kraak GJ (eds) Environmental fate and effects on pulp and paper mill effluents. St. Lucie Press, Delray Beach, Florida, pp 495–509
Palm HW (1999) Ecology of Pseudoterranova decipiens (Krabbe, 1878) (Nematoda: Anisakidae) from Antarctic waters. Parasitol Res 85:638–646
Palm HW (2000) Trypanorhynch cestodes from Indonesian coastal waters (East Indian Ocean). Folia Parasitol 47:123–134
Palm HW (2004) The Trypanorhyncha Diesing, 1863. PKSPL-IPB, Bogor
Palm HW (2007) The concept of cummulative evolution in marine fish parasites. Proceedings of the IVth international workshop on theoretical and marine parasitology, AtlantNIRO, Kaliningrad, 21–26 May 2007, pp 164–170
Palm HW (2008) Surface ultrastructure of the elasmobranchia parasitizing Grillotiella exilis and Pseudonybelinia odontacantha (Trypanorhyncha, Cestoda). Zoomorphology 127:249–258
Palm HW, Caira J (2008) Host specificity of adult versus larval cestodes of the elasmobranch tapeworm order Trypanorhyncha. Int J Parasitol 38:381–388
Palm HW, Dobberstein RC (1999) Occurrence of trichodinid ciliates (Peritricha: Urceolariidae) in the Kiel Fjord, Baltic Sea, and its possible use as a biological indicator. Parasitol Res 85:726–732
Palm HW, Klimpel S (2007) Evolution of the parasitic life in the ocean. Trends Parasitol 23:10–12
Palm HW, Klimpel S (2008) Metazoan fish parasites of Macrourus berglax Lacepède, 1801 and other rattails in the North Atlantic: Exploration of the deep-sea from the continental shelf. Deep Sea Res Part II 55:236–242
Palm HW, Overstreet RM (2000) Otobothrium cysticum (Cestoda: Trypanorhyncha) from the muscle of butterfishes (Stromateidae). Parasitol Res 86:41–53
Palm HW, Rückert S (2009) A new approach to visualize ecosystem health by using parasites. Parasitol Res 105:539–553
Palm HW, Schröder P (2001) Cestode parasites from the elasmobranchs Heptranchias perlo (Bonnaterre, 1788) and Deania spp. Jordan et Snyder, 1902 from the Great Meteorbank, central East-Atlantic. Aquat Liv Res 14:137–144
Palm HW, Reimann N, Spindler M, Plötz J (1998) The role of the rock cod Notothenia coriiceps in the life cycle of Antarctic parasites. Pol Biol 19:399–406
Palm HW, Klimpel S, Bucher C (1999) Checklist of metazoan fish parasites of German coastal waters. Ber Inst Meereskd Nr 307:1–148
Palm HW, Klimpel S, Walter T (2007) Demersal fish parasite fauna around the South Shetland Islands: High species richness and low host specificity in deep Antarctic waters. Pol Biol 30:1513–1522
Palm HW, Damriyasa IM, Linda L, Oka IBM (2008) Molecular genotyping of Anisakis Dujardin, 1845 (Nematoda: Ascaridoidea: Anisakidae) larvae from marine fish of Balinese and Javanese waters, Indonesia. Helminthologia 45:3–12
Palm HW, Waeschenbach A, Olson P, Littlewood T (2009) Molecular phylogeny and evolution of the Trypanorhyncha Diesing, 1863. Mol Phylogenet Evol 52:351–367
Palm HW, Kleinertz S, Rückert S (2011) Parasite diversity as an indicator of environmental change? An example from tropical grouper (Epinephelus fuscoguttatus) mariculture in Indonesia. Parasitology. (Suppl.) doi:10.1017/50031182011000011
Paperna I, Overstreet RM (1981) Parasites and diseases of mullets (Mugilidae). In: Oren OH (ed) Agriculture of grey mullets. Cambridge University Press, Great Britain, pp 411–493
Pascual S, Hochberg FG (1996) Marine parasites as biological tags of cephalopods hosts. Parasitol. Today 12:324–327
Pech D, Vidal-Martínez VM, Aguirre-Macedo ML, Gold-Bouchot G, Herrera-Silveira J, Zapata-Pérez O, Marcogliese DJ (2009) The checkered puffer (Spheroides testudineus) and its helminths as bioindicators of chemical pollution in Yucatan coastal lagoons. Sci Total Environ 407:2315–2324
Petersen F, Palm HW, Möller H, Cuzi MA (1993) Flesh parasites of fish from central Philippine waters. Dis Aquat Org 15:81–86
Pettersen RA, Vøllestad LA, Flodmark LEW, Poléo ABS (2006) Effects of aqueous aluminium on four fish ectoparasites. Sci Total Environ 369:129–138
Poulin R (2007) The structure of parasite communities in fish hosts: ecology meets geography and climate. Parassitologia 49:169–172
Poulin R, Mouritsen KN (2006) Climate change, parasitism and the structure of intertidal ecosystems. J Helminthol 80:183–191
Reimer LW (1995) Parasites especially of piscean hosts as indicators of the eutrophication in the Baltic Sea. Appl Parasitol 36:124–135
Riemann F (1988) Nematoda. In: Higgins RP, Thiel H (eds) Introduction to the study of meiofauna. Smithsonian, Washington, DC, pp 293–301
Rohde K (2002) Ecology and biogeography of marine parasites. Adv Mar Biol 43:1–86
Rokicki J (1983) Ectoparasites of the hakes Merluccius merluccius capensis (Castelnau) and Merluccius merluccius paradoxus (Frank) as an aid to host systematics. ICSEAF 1983 (Part I). Coll Sci Pap Int Comm SE Atl Fish, 10:167–170, doi:10.1017/S0031182011000011
Rückert S, Hagen W, Yuniar AT, Palm HW (2009a) Metazoan parasites of fishes and their potential use as biological indicators in the Segara Anakan Lagoon, Indonesia. Reg Environ Change 9:315–328
Rückert S, Klimpel S, Al-Quraishy S, Mehlhorn H, Palm HW (2009b) Transmission of fish parasites into grouper mariculture (Serranidae: Epinephelus coioides (Hamilton, 1822)) in Lampung Bay, Indonesia. Parasitol Res 104:523–532
Rückert S, Klimpel S, Palm HW (2010) Parasites of cultured and wild grouper (Epinephelus fuscoguttatus) in Lampung Bay, Indonesia. Aquaculture Research 41:1158–1169, doi:10.1111/j.1365-2109.2009.02403.x
Ruus A, Skaare JU, Ingebrigtsen K (2001) Accumulation of the lipophilic environmental contaminant lindane in metacercariae of Bucephaloides gracilescens (Trematoda, Bucephalidae) in the central nervous system of bullrout Myoxocephalus scorpius. Dis Aquat Organ 48:75–77
Sasal P, Mouillot D, Fichez R, Chifflet S, Kulbicki M (2007) The use of fish parasites as biological indicators of anthropogenic influences in coral-reef lagoons: A case study of Apogonidae parasites in New-Caledonia. Mar Poll Bull 54:1697–1706
Sharpe DMT, Hendry AP (2009) Life history change in commercially exploited fish stocks: an analysis of trends across studies. Evol Appl 2:260–275
Shulman SS (1957) Material on the parasitofauna of lampreys from the basins of the Baltic and the White Seas. Isvestiya gosud nauchnovatel´skogo issled Inst Ozern Techn rybnogo khozyajstva 42:287–303, Translated from the Russian by the Israel program for Scientific Tanslations, No. 105, 1961
Skinner RH (1982) The interrelation of water quality, gill parasites, and gill pathology of some fishes from South Biscayne Bay, Florida. Fisher Bull 80:269–280
Sures B (2003) Accumulation of heavy metals by intestinal helminths in fish: an overview and perspective. Parasitol 126:53–60
Sures B (2004) Environmental parasitology: relevancy of parasites in monitoring environmental pollution. Trends Parasitol 20:170–177
Sures B, Reimann N (2003) Analysis of trace metals in the Antarctic host-parasite system Notothenia coriiceps and Aspersentis megarhynchus (Acanthocephala) caught at King George Island, South Shetland Islands. Pol Biol 26:680–686
Sures B, Siddall R (2003) Pomphorhynchus laevis (Palaeacanthocephala) in the intestine of chub (Leuciscus cephalus) as an indicator of metal pollution. Int J Parasitol 33:65–70
Sures B, Taraschewski H (1995) Cadmium concentrations in two adult acanthocephalans, Pomphorhynchus laevis and Acanthocephalus lucii, as compared with their fish hosts and cadmium and lead levels in larvae of A. lucii as compared with their crustacean host. Parasitol Res 81:494–497
Sures B, Taraschewski H, Jackwerth E (1994) Lead accumulation in Pomphorhynchus laevis and its host. J Parasitol 80:355–357
Sures B, Taraschewski H, Rokicki J (1997) Lead and cadmium content of two cestodes, Monobothrium wageneri and Bothriocephalus scorpii, and their fish hosts. Parasitol Res 83:618–623
Tekin-Özan S, Kir I (2005) Comparative study on the accumulation of heavy metals in different organs of tench (Tinca tinca L. 1758) and plerocercoids of its parasite Ligula intestinalis. Parasitol Res 97:156–159
Vidal-Martínez VM, Pech D, Sures B, Purucker ST, Poulin R (2010) Can parasites really reveal environmental impact? Trends Parasitol 26:44–51
Voigt MOC (1993) Protozoan ectocommensals of toadfish and soldier crab as indicators of organic pollution in the Brisbane River estuary, Queensland, Australia. Ph.D. thesis, University of Queensland, Brisbane
Williams HH, MacKenzie K (2003) Marine parasites as pollution indicators: an update. Parasitology 126:S27–S41
Williams HH, MacKenzie K, McCarthy AM (1992) Parasites as biological indicators of the population biology, migrations, diet, and phylogenetics of fish. Rev Fish Biol Fish 2:144–176
Xinghua L (1987) Factors regulating trematodes and cestodes in fish. Arch Hydrobiol 28:381–387
Yeomans WE, Chubb JC, Sweeting RA (1997) Use of protozoan communities for pollution monitoring. Parassitologia 39:201–212
Zander CD, Kesting V (1996) The indicator properties of parasite communities (Teleostei, Gobiidae) from Kiel and Lübeck Bight. Appl Parasitol 37:186–204
Zander CD, Kesting V (1998) Colonization and seasonality of goby (Gobiidae, Teleostei) parasites from the southwestern Baltic Sea. Parasitol Res 84:459–466
Acknowledgements
I am thankful to the German Research Council (DFG PA 664/4–1 and 2), the German Academic Exchange Service (DAAD) and the German Federal Ministry for Education and Science (BMBF Grant Nos. 03F0391(471)A within the framework of the joint Indonesian-German research programme SPICE (Science for the Protection of Indonesian Coastal Marine Ecosystems)) who supported parts of the funding for the presented results. Dr. R. Bray, The Natural History Museum London, provided helpful comments on an earlier version of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2011 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Palm, H.W. (2011). Fish Parasites as Biological Indicators in a Changing World: Can We Monitor Environmental Impact and Climate Change?. In: Mehlhorn, H. (eds) Progress in Parasitology. Parasitology Research Monographs, vol 2. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-21396-0_12
Download citation
DOI: https://doi.org/10.1007/978-3-642-21396-0_12
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-21395-3
Online ISBN: 978-3-642-21396-0
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)