
Abstract
Among the strategies that parasites have evolved to increase the probability of a successful transmission, the (targeted) alteration of the host’s phenotype is certainly the most impressive and spectacular way. A large number of publications exist that have compiled and categorised correlations of parasitic infections and the occurrence of changes of phenotypic traits (e.g. behaviour, morphology, physiology) of their hosts. In this context, studies on fish parasites have documented an impact on almost every aspect of fish behaviour alongside changes in host morphology and/or physiology. This short review provides a quick and basic overview over the forms and diversity of parasite induced phenotypic changes in marine and freshwater fish species. Most frequently reported parasites and their correlated alterations were compiled from 57 publications and presented in a table. Behavioural changes were found among seven different parasite groups: Protozoa, Myxozoa, Digenea, Monogenea, Cestoda, Nematoda and Crustacea. Changes were categorized into five major categories: foraging, habitat selection, motility, predator avoidance and reproduction, whereas the most common behavioural change was involving motility with 34 cases, followed by predator avoidance (10), reproduction (8), foraging (5), and habitat selection (3). Most species involved in behavioural changes were found among the group of Digenea, with 13 different parasite species, followed by Cestoda with 6 different species. All other groups were more or less equally represented. Parasite specific forms of host manipulation are exemplified and discussed in the text.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
9.1 Introduction
According to the concepts of evolution and adaptation, the entire biology of all animal species, whether free-living or parasitic, is a result of and geared towards its survival and reproduction. By definition, a parasite is an organism that lives on or in another organism (host) exploiting the resources necessary for their own survival and causing it harm. Although probably one of the most successful life strategies in nature (in fact, it is generally believed that parasites outnumber free-living species in food webs), limiting determinant is doubtlessly the need of the parasite to encounter an appropriate host at each stage of its life cycle (e.g. Windsor 1998; MacKenzie 1999; Poulin and Morand 2004; Rossiter 2013). The life-cycle of fish parasites can either be direct or indirect, i.e. involving one or a trophic transmission via several host species, respectively. Fish parasites with a complex life cycle are faced with a particular challenge in large volume aquatic environments where densities of hosts are generally low. Under such circumstances, the emergence of specific strategies to increase frequencies of encounters of parasites and hosts seems inevitable (Cézilly et al. 2010). Parasites have evolved strategies to increase the probability of a successful transmission, i.e. either by increasing random encounters simply by disseminating vast amounts of propagules, or by producing specified propagules that will reduce the randomness of host encounters (Combes 1991; Sasal and Thomas 2005).
Among these strategies, the (targeted) alteration of the host’s phenotype is certainly the most impressive and spectacular way of a parasite to improve its odds in the transmission process. Although it has generated a lot of attention among scientists who put massive efforts into the documentation and exploration of these phenomena, the basic mechanisms behind these alterations remain largely unknown. Those who have thoroughly studied and reviewed the facilitation of parasite transmission and demographic effects, (physiological) mechanisms, adaptive nature, population biology, and phenotypic alterations in hosts in the context of parasite induced changes in host behaviour are, to name but a few, Barber (2007), Barber et al. (2000), Combes (1991), Dobson (1988), Hurd (1990), Lafferty (1999), Moore (1984, 2002, 2013), Moore and Gotelli (1990), Poulin (1994), Poulin and Thomas (1999).
Combes (1991) analysed the elementary ethological mechanisms that may have been the object of selection in a parasite’s life cycle and coined the term “favorization” which describes an adaptive ethological process modifying the spatio-temporal positions of the parasite and its target in a way that increases the probability of their encounter. Kuris (1997) introduced the more specific term “parasite increased susceptibility to predation” to represent the manipulation by a parasite that increases predation on its intermediate hosts. Lafferty (1999) refined this later as “parasite increased trophic transmission (PITT)” under the assumption that an increased predation is not necessarily adaptive (cf. Combes 1991).
In this context, Poulin (1995) pointed out that phenotypic changes can only be considered adaptive if certain prerequisites apply and proposed four criteria to distinguish between adaptive and non-adaptive alterations: (1) complexity of alterations, (2) degrees of purposive design, (3) independent evolution among lineages, and most importantly, (4) an increase of the fitness of the host. However, the question whether phenotypic host alterations are of adaptive origin or rather side effects or “by products” of an infection, is still controversially discussed (e.g. Cézilly et al. 2010; Thomas et al. 2012).
An impressive number of publications exist that have compiled and categorised parasitic infections and the correlated occurrence of changes in phenotypic traits (e.g. behaviour, morphology, physiology) of their hosts. Among those, many studies on fish parasites have documented an impact on almost every aspect of fish behaviour (Barber et al. 2000). The aim of this short communication is to provide a quick and basic overview over the forms and diversity of parasite induced behavioural changes in marine and freshwater fish species. Most frequently reported parasites and fish behaviour alterations were compiled and presented in a table. Additionally, some parasite specific forms of behaviour are exemplified in the text. As this overview study does not claim to be exhaustive, we refer the reader to more detailed and in depth reviews by e.g. Barber (2007), Barber et al. (2000), Dobson (1988), Moore (2002).
9.2 Forms of Host Manipulation
Host manipulation, best studied and known from parasites with indirect life cycles, seems to enhance probabilities of transmission and vulnerability to predatory definitive hosts (Moore 2002; Thomas et al. 2005). Poulin (2010) defined host manipulation as any alteration in host phenotype induced by a parasite that has fitness benefits for the parasite. As such, typical phenotypic changes in hosts comprise behavioural alterations alongside changes in host morphology and/or physiology. As described by Thomas et al. (2005) there are two ways of how parasites can alter host behaviour. Direct mechanisms involve interactions with nervous system or muscle, e.g. neuroactive substances may be secreted by the parasite resulting in a behavioural change of the host. Indirect effects can occur when the presence of the parasite affects host development, intermediate metabolism and/or immunity, leading secondarily to changes in behaviour (host-mediated). By the same token it is assumed that a direct manipulation of the host’s behaviour requires the parasite to expend energy, whilst indirect manipulations do not pose additional energetic costs for the parasite. However, a differentiation of manipulative behavioural changes (direct and indirect) as well as the isolation of specific mechanisms has proven to be difficult. Another problem is that many results on parasite induced behavioural changes in hosts have been obtained in laboratory experiments. Thus, they might only be valid under controlled conditions and their relevance in the field needs to be considered carefully. In conclusion, changes in host behaviour are most likely a mix of direct and indirect effects and it is believed that the basic mechanisms behind host manipulation involve four physiological systems, which are to a large extent interconnected: neural, endocrine, neuromodulatory, and immunomodulatory (Lafferty and Shaw 2013).
Various alterations in behaviour of hosts infected by parasites are known and have been extensively described by Moore (2002). In fish hosts, they are frequently reported from host individuals carrying very heavy parasite burden (Barber et al. 2000). In this section, a general overview of the most common behavioural changes in fish hosts will be given: foraging behaviour (Fig. 9.1), implications on motility (Fig. 9.2), altered sexual behaviour (reproduction) (Fig. 9.3), anti-predator behaviour (predator avoidance) (Fig. 9.4), and habitat selection (Fig. 9.5). For a more extensive review see Barber et al. (2000).

Influence of parasites on foraging behaviour of fish host

Influence of parasites on motility of fish host

Influence of parasites on reproduction of fish host

Influence of parasites on predator avoidance of fish host

Influence of parasites on habitat selection of fish host
9.2.1 Foraging Behaviour (Fig. 9.1)
Alterations of foraging behaviour may either be directly caused by the physical presence of the parasite that restricts the capacity of the stomach (Wright et al. 2006), or a response of the host to compensate for the physiological disadvantages by the infection (e.g. Godin and Sproul 1988). One of the disadvantages is that parasites utilize the host’s energy reserves for their own growth and development. Thus, energetic drain is one of the major physiological implications of parasite infections leading to nutritional stress in the host. However, nutritional stress can also be a consequence of infections that reduce the competitive ability or act on the foraging success (Barber and Wright 2005). Hence, one way to counteract the energetic limitation is to avoid food competition through alterations in prey choice and diet composition (e.g. Milinski 1984). Another way for compensation is to increase time for foraging, which is a typical behaviour of hosts infected by large parasite species (Barber and Wright 2005; Östlund-Nilsson et al. 2005). Conversely, decreased foraging is observed in hosts infected with highly pathogenic parasites and might be caused by decreased appetite, which may be an adaptive strategy by the host to deprive energy for the parasite and consequently lower the growth rate of parasites (Bernier 2006, 2010; Chin et al. 2004).
9.2.2 Implications on Motility (Fig. 9.2)
Motility is a key element of organisms and a prerequisite for flight and foraging behaviour. Changes in motility can be found in a wide variety of fish genera. Here, parasite infections mostly result in reduced, but versatile, swimming behaviour, e.g. due to muscle atrophy, a damaged nervous system or impairment of blood circulation (Barber 2007; Coleman 1993; Shirakashi and Goater 2005; Sweeting 1977). Frequent infection sites are organs necessary for orientation such as eyes, inner ear, nostrils and linea lateralis (Barber 2007). Fish motility might also be affected by infestations of the cardio-visceral system or infections with blood sucking ectoparasites compromising the endurance and swimming performance (Barber 2007; Coleman 1993). Whether ectoparasites increase the host’s energetic costs of swimming, e.g. by changing hydrodynamics, is controversially discussed (Binning et al. 2014; Östlund-Nilsson et al. 2005).
9.2.3 Reproduction (Fig. 9.3)
How parasites alter the reproductive behaviour of their hosts includes a broad research spectrum. Here, only gonad development, mate choice, courtship, and parental care will be addressed briefly. A widely distributed strategy of parasites is the interference with host reproductive effort (Barber et al. 2000). Fecundity reduction can be caused by parasites through simple nutrient theft (‘consumers’) or manipulation of energy allocation away from reproduction to growth (‘castrators’) (Baudoin 1975; Hall et al. 2007; Heins et al. 2010). Most likely as a response to the reduced fecundity, two phenomena, known as ‘gigantism’ and ‘early infection fecundity compensation’, are known from infected hosts (see Hall et al. 2007). Host castration occurs widely among taxa, but most often occurs in invertebrates such as crustaceans, echinoderms, and molluscs. This can be explained by a higher investment in reproductive tissues relative to the body mass in smaller species (Lafferty and Kuris 2009). However, a variety of fish are also affected (in Fogelman et al. 2009).
Alternatively to direct physical implications, parasites can also alter mating behaviour. Sexual selection is the preference of certain mating partners over others in order to increase fitness. Traits that are selected include benefits such as parental care, attractiveness of offspring, and resistance against pathogens and parasites (Andersson 1994). The Hamilton-Zuk (H-Z) hypothesis (1982) states that females choose their mating partners based on the evolved secondary sexual characters, which should reflect the heritable resistance to parasites. More specifically, it is hypothesized that female sticklebacks prefer brighter males because (1) colouration reflects the healthiness of the mating partner, i.e. males would be pale if they were infected with white-spot disease (2) after spawning males do parental care and healthier males should be more able to defend the eggs (Milinski and Bakker 1990), and (3) the offspring are more resistant to parasites which is especially beneficial in habitats with high parasite abundance (Barber et al. 2000). On the latter point, a study has been conducted that supported the sexual ornamentation theory (H-Z hypothesis) and showed a trade-off between growth and parasite resistance, which may reflect the maintenance of heritable variation in infection resistance and male colouration (Barber et al. 2000). In order to further study the heritability of resistance to pathogens and parasites, a variety of studies has focused on the diversity of major histocompatibility complex (MHC) alleles which is important in controlling the vertebrate immune system and plays a considerable role in mate choice (Consuegra and Garcia de Leaniz 2008; Eizaguirre et al. 2012; Reusch et al. 2001).
9.2.4 Predator Avoidance and Microhabitat Selection (Figs. 9.4 and 9.5)
Trophically transmitted parasites may facilitate their transmission by manipulating their hosts for higher vulnerability to predation (Parasite transmission hypothesis, Kuris 1997; Moore 2002). As predation has a serious fitness consequence for both hosts and parasites there should be strong selection on avoidance traits (Barber et al. 2000). Parasite-induced host vulnerability to predators should occur in a way that targets an increase in predation by a definitive host, as otherwise a non-host predation would lead to the end of the parasite life-cycle (Mouritsen and Poulin 2003). Predators themselves should avoid infected fish if parasites pose a high impact on their constitution (Barber et al. 2000). There are several ways of parasite-induced changes on predator avoidance which facilitate a transmission of parasites from intermediate to final hosts.
Protective colouring is a host adaptation to decrease the risk of being detected by a predator and can be lost by parasite-induced morphological changes such as white- and black-spot disease making parasites more visible on the host (Bakker et al. 1997; Krause and Godin 1996; Milinski 1985). Another example is the orange-yellow acanthocephalan parasite Pomphorhynchus laevis. As it is visible through the transparent cuticle of crustacean Gammerus pulex the crustacean becomes more prone to fish predators (Bakker et al. 1997).
In order to not encounter their predators, fish avoid being present at the same time and place as their predators (Barber et al. 2000). Hosts that have been parasitized, however, need to increase foraging activity and thus, seem to return to feeding more quickly after a frightening event compared to uninfected individuals (Giles 1983; Godin and Sproul 1988). A form of predator avoidance is shoaling. Fish form preferentially large shoals, especially after a predator attack, due to the effects of predator confusion, risk dilution and early predator detection (see Krause and Godin 1994a). A social adaptation of fish against parasites may be to form larger shoals in their presence in order to reduce the individual risk to acquire a parasite (Poulin and FitzGerald 1989). Conversely, parasitized fish have been shown to spend less time in shoals and occur more frequently in the more risky peripheral shoal positions (Krause and Godin 1994b).
Parasite-induced alterations in predator avoidance often overlap with habitat choice. Habitat choice is associated with predator avoidance, mating, and foraging behaviour (Barber et al. 2000). Parasites may alter aggression levels and territorial behaviour which affect vulnerability to predators due to reduced avoidance (Mikheev et al. 2010). Alterations in habitat choice can also be related to physiological restrictions. Parasites might pose high metabolic demands, e.g. ectoparasites on the gills reduce the oxygen level for host respiration or impair gas exchange efficiency (Barber and Wright 2005; Smith 1972). As a consequence fish need to compensate the respiratory deficiency through location of habitats with higher oxygen levels. Such habitats are usually near the water surface where fish are more visible to avian predators (Barber and Wright 2005; Lester 1971). Physiological restrictions such as cataracts caused by eye-infecting parasites lead to surface-swimming behaviour and consequently increase the risk of being detected (Crowden and Broom 1980). Impairments in the visual performance forces fish to sojourn at habitats with better light conditions, and reduce the ability to recognize predators (Seppälä et al. 2004).
9.3 Examples of Phenotypic Changes in Fish
Based mainly on the reviews by Moore (2002) and Barber et al. (2000), 57 studies were chosen to represent common parasite-fish behavioural interactions and covering as many different parasite species and hosts as possible. Behavioural changes were found among seven different parasite groups: Protozoa (Microsporidia, Sporozoa), Myxozoa, Digenea, Monogenea, Cestoda, Nematoda, Crustacea. Changes were categorized into five major categories (Table 9.1 and Fig. 9.6) based on former classifications used by Barber et al. (2000): foraging, habitat selection, motility, predator avoidance and reproduction. A behaviour that could not be unequivocally assigned was noted as indetermined, e.g. aggressiveness.

Occurrences of different types of host manipulation found for each parasite group
The most common behavioural change was involving motility with 34 cases, followed by predator avoidance (10), reproduction (8), foraging (5), and habitat selection (3). Most species involved in behavioural changes were found among the group of Digenea, with 13 different parasite species, followed by Cestoda with 6 different species. All other groups were more or less equally represented by 3 or 4 different parasite species. Within the Digenea Diplostomum spathaceum seems to infest different host species causing a different behavioural change in each. In contrast to this, Myxobolus cerebralis, a species belonging to the Myxozoa, occurs in many different fish species but evokes only changes in motility, known as whirling disease. In the group of Cestoda, Ligula intestinalis seems to infect at least three different fish species of which all show a different behavioural change, either in reproduction, habitat choice or foraging (Table 9.1).
Glugea anomala, a protozoan species, uses Gasterosteus aculeatus as its host and three different types of behavioural alterations have been observed, i.e. foraging, motility and predator avoidance. However, this last example, which is based on two studies (Milinski 1985; Ward et al. 2005), clearly suggests that parasite infections might not only lead to changes in one type of host behaviour (always characterized and interpreted from a human perspective) but might rather be an amalgamation of different behavioural changes possibly associated with other phenotypic changes in the host (e.g. in morphology, physiology). Altogether, these changes might enhance successful trophic transmissions of the parasite.
In the following section some examples of parasite-host interactions depicted in Table 9.1 will be presented in more detail for each parasite group.
9.3.1 Protozoa (Microsporidia, Sporozoa)
Protozoa have developed a great diversity in terrestrial as well as aquatic habitats, with over 100.000 species in more than 40 phyla, and many of them have a parasitic life strategy (O’Donoghue 2005). Some of the parasitic protozoa are suspected to cause changes in host behaviour. These include species, belonging to Mastigophora (e.g. Cryptobia salmositica) and Microsporidia (e.g. Pleistophora ladogensis, Glugea anomala). In the division of Microsporidia more than 1300 species in 150 genera are known, most of them parasitizing invertebrates, but approx. 100 species have been found in fish already. One shared characteristic is their reproduction being linked to host cells (Moodie 2005). The Microsporidia Glugea anomala uses three-spined sticklebacks, Gasterosteus aculeatus as a common host. Teleost hosts of G. anomala become infected by ingesting spores directly from the water column or by preying on infected invertebrates. In the host tissue G. anomala forms xenoma by hypertrophic grows of the host cells accumulating millions of spores within them (Weissenberg 1968). Xenoma usually form in the head region of the fish as visible small white cysts, which burst open and release the containing spores (Ward et al. 2005). Moodie (2005) speculated that migratory movements are effected by cysts in muscle tissue. Ward et al. (2005) observed weight loss of naturally infected G. aculeatus, compared to none infected. Therefore, Glugea anomala seems to influence the fish host energetic costs. It also seems to have an impact on predator avoidance and shoaling behaviour: Under experimental conditions infected specimens showed increased tendency to form shoals, presumably as a predator avoidance mechanism and to take leading positions, within it, which is probably a concession to the higher food supply as it increases their chance to prey more, compared to other positions in the shoal (Krause 1994; Milinski 1985; Ward et al. 2005).
An example for decreased foraging behaviour is Cryptobia salmositica and its host, the rainbow trout (Oncorhynchus mykiss). Cryptobia salmositica is a parasitic flagellate (class Kinetoplastea) that has been categorized as haemotozoic (Woo 2011). Rainbow trout get infected either when C. salmositica multiplies in the crop of infected blood-sucking leech (Pisciola salmositica) that transmit the kinetoplast with the following blood meals (Woo 1994), or from fish to fish (Woo 2011). Clinical signs of infections are high amounts of mucus on the body surface, lethargy, and less movement of the fish. During the acute phase of the infection, the host has obvious lesions in haemopoietic tissues, and up to five weeks after the infection the fish shows anorexia causing an energetic drain (Woo 2011). On the one hand, this can lead to immune-depression, which often facilitates secondary diseases (Jones et al. 1986; Thomas and Woo 1992). On the other hand, anorexia has been shown to be beneficial because it lowers the plasma protein level and consequently reduces the severity of the infection (Li and Woo 1991). Anorexia and anaemia are most consistent clinical signs of the parasite infection (Woo 2011).
9.3.2 Myxozoa
Myxozoa are economically important spore-forming metazoan endoparasites, occurring predominantly in fish (MacKenzie and Kalavati 2014; Yokoyama et al. 2012). Hitherto more than 1350 species, belonging to 52 different genera are described, occurring in freshwater and marine habitats. For a long time it was controversially discussed whether they belong to the proto- or metazoan just as long they were assumed to perform a monoxenous life cycle, until 1986 the alternation of Myxobolus cerebralis between Oncorhynchus mykiss and Tubifex tubifex was proven (Smothers et al. 1994; Wolf et al. 1986). Today, the heteroxenous life cycle, including invertebrates and vertebrates, is common knowledge (Hedrick et al. 1998; Kent et al. 2001). The life cycle of Myxosporea alternates mostly between annelids and teleost. The life cycle includes an actinosporea-spore as infectious stadium in the invertebrate and a myxosporea-spore in the vertebrate host. Previously these spores had been classified as different species. While oligochaetes serve mostly as invertebrate hosts in freshwater waters, in marine waters only polychaetes are known (see review MacKenzie and Kalavati 2014). The vertebrate host gets infected by consuming myxosporea-spores, which anchor themselves in the host gut epithelium using polar filaments and inject an infectious sporoplasm (El-Matbouli and Hoffmann 1998). The sporoplasm then develops into a species-specific actinosporea-spore (e.g. tricatinomyxon (Myxobolus cerebralis), hexactinomyxon (Myxobolus pavlovskii), or aurantiactinomyxon (Hofferellus carassii) (El-Matbouli and Hoffmann 1998; Kent et al. 2001). These spores are released with the faeces into the water column, where they attach themselves to the teleost host skin and release again a sporoplasm into mucous cells which develop into myxosporea-spores. Later again, these are, depending on the species, released with the faeces into the water column or are otherwise set free after death of the teleost host (Kent et al. 2001). Spores, e.g. of Myxobolus cerebralis, can be set free, after digestion of predatory fish or birds with their faeces (Gilbert and Granath 2001; Hedrick et al. 1998). Myxobolus cerebralis is the known agent of the whirling disease and is mainly infecting different economically important salmonids in freshwater (Hedrick et al. 1998). Particularly young fish seem to be very susceptible to the effects of the disease and heavy infections often result in their death (Gilbert and Granath 2001). Beside morphological changes such as a black-ended tail and deformations, Myxobolus cerebralis causes specific behavioural changes in its teleost host (Gilbert and Granath 2001; Hedrick et al. 1998). Rose et al. (2000) observed three specific behaviours associated with whirling disease in Oncorhynchus mykiss, (a) a repeated rapid circular swimming, a kind of startle response, (b) lack of corrective movement while in a static position in the water, resulting in a tail-up head-down posture, (c) discontinued swimming movement and sinking to the bottom. The whirling movement is probably caused by a constriction of the spinal cord, repeatedly triggering impulses for tight turns until the signals are interrupted by competing sensory stimuli (Rose et al. 2000). It can be assumed that those changes, caused by M. cerebralis, increase the risk of predation, hence accelerating the release of the myxosporea-spores back into the water column.
9.3.3 Digenea
Digenean trematodes are endoparasites with complex life cycles that vary considerably between the taxa. These parasites commonly occur in the alimentary tract of their definitive host, which is, in most cases, a vertebrate (Cribb 2005). A typical life cycle includes the motile miracidium stage that hatches from an egg dispersed within the host’s faeces. Molluscs are the first obligate intermediate hosts in which the miracidium develops into an asexually reproducing sporocyst that may either produce further generations of sporocysts or alternatively one or more generations of rediae which themselves release infectious stages known as cercariae. Cercariae leave the mollusc and will be either encysted (metacercariae) in or on a second intermediate host (e.g. crustaceans, plants) or directly infect the final host and grow into the adult stage (Cribb 2005; Möller and Anders 1986). Additional intermediate hosts may be incorporated before infecting the definitive host. Digenea typical for fish are represented by the families Derogenidae, Hemiuridae, Lecithasteridae and Lepocreadiidae (Klimpel et al. 2009).
Most behavioural changes in marine and freshwater fish can be attributed to parasites belonging to the group of Digenea, with 13 species in at least 13 different host species. Whether these numbers are the result of a high sampling effort due to their enormous medical relevance or are simply reflecting their complex life and thus the various possibilities for host alterations to occur, is not clear. Whilst changes in foraging behaviour of infected fish hosts (Leuciscus leuciscus) as well as avoidance of predators (e.g. Galaxias vulgaris, Oncorhynchus mykiss) have been documented six times, alterations in locomotion and/or motility of fish are by far the most documented changes associated with Digenean infections (Table 9.1).
A popular and very well documented example of motility implications caused by the presence of digenean trematodes in fish is Euhaplorchis californiensis infecting killifish, Fundulus parvipinnis (Lafferty and Morris 1996). The authors provided probably the best example for a case of increased predation on infected intermediate hosts by definitive hosts. Euhaplorchis californiensis is a common trematode in salt marshes of southern California. It has a three-host life cycle, which includes a number of birds as definitive and horn snails Cerithidea californica as first intermediate hosts (Lafferty and Morris 1996). When killifish F. parvipinnis, the second intermediate host, becomes infected with cercariae, they show “conspicuous behaviour”, i.e., abrupt dashes up to the surface (surfacing), turning laterally so that one side of the body is faced up (flashing), dorsal-ventral bending (contorting) and vibrating for a few seconds (Lafferty and Morris 1996). These behavioural changes have been associated with the parasites encysting the brain case. In experiments, the authors demonstrated that infected specimens had a 10-fold increase susceptibility to be eaten by the definitive hosts than their non-infected conspecifics.
An increased likelihood of detection and attack by predators of hosts has also been associated with infections of Diplostomum spathaceum (Fig. 9.7a). After their development in lymnaeid snails (e.g. Lymnea sp., Radix sp.), the free-swimming cercariae of D. spathaceum penetrate the fish skin and migrate into the crystalline lense of the fish (Lyholt and Buchmann 1996). Adult parasites live in the intestines of piscivorously feeding birds (mainly from the family Laridae). The presence of large numbers of parasites in the eye of their host can cause the lense to become opaque which results in a complete lack of responsiveness to visual stimuli as shown for heavily infected trout (Crowden and Broom 1980; Ferguson and Hayford 1941). Generally, reduced visual acuity seems to diminish the host’s ability to locate and capture food with the consequence that the feeding efficiency is significantly reduced. Infected fish, e.g. Leuciscus leuciscus, compensate this by increasing the net amount of time devoted to feeding and also by foraging in better-illuminated surface waters where they are more prone to predation (Crowden and Broom 1980; Dobson 1988).

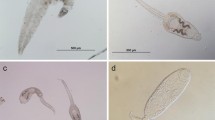
Examples of host manipulating parasites. (a) Diplostomum spathaceum (Digenea). (b) Anguillicoloides crassus (Nematoda) in eel swim bladder. (c) Lernaeocera branchiails (Crustacea) in cod gills. (d) Argulus sp. (Crustacea) (Photo (a): Emde)
In a recent experimental study, rainbow trout (Onchorhynchus mykiss) infected with D. spathaceum showed increased aggressiveness and altered territorial behaviour (Mikheev et al. 2010). After an initial decrease, the aggression rates of infected O. mykiss exceeded that of uninfected control fish, yet, quite surprisingly, they seemed to face reduced “contest ability” in territorial fights. Similarly, Seppälä et al. (2004, 2008) reported reduced escape behaviour as well as shoaling of experimentally infected, juvenile O. mykiss in the presence of artificial aerial predators. The authors argued that the reduction in vision is the definitive mechanism that causes the altered fish behaviour and increased susceptibility to predators.
Motility is another trait that can be affected by parasites infesting the cardio-visceral system and therefore compromising the endurance and swimming performance of the fish host (Barber 2007; Coleman 1993). One parasite inducing those changes is Ascocotyle pachycystis, usually parasitizing in the bulbus of the sheepshead minnow Cyprinodon variegatus. Especially during cold conditions, the negative effect of decreased oxygen transport, caused by increased blood flow resistance by the cysts of A. pachycystis, leads to faster exhaustion of parasitized specimens compared to unparasitized specimens (Coleman 1993).
9.3.4 Monogenea
The taxon Monogenea is one of the largest groups of parasitic flatworms. A vast majority is ectoparasitic on the external surfaces (fins, gills, skin) of marine and freshwater fish while some are secondarily endoparasitic in body orifices (e.g. mouth cavity, urinary bladder) (Hayward 2005). A few additional species live on aquatic invertebrates and higher vertebrates such as cephalopods, turtles and amphibians (Möller and Anders 1986). The taxon consists of two sister groups (Monoopisthocotylea, Polyopisthocotylea) that can easily be distinguished by the morphology of their posterior attachment organ (opisthaptor) (Whittington 2005). The development of the monogenean species is direct (i.e. they infect only a single host) and they usually show high degrees of host specificity. The eggs of most species have long appendages used for attachment to the gill filaments of their hosts or to bottom substrates. They can also be found on artificial substrates like e.g. the net caging of aquaculture facilities where mass infections can cause enormous ecological losses (e.g. mass mortality of aquaculture specimens) due to the serious pathological effects on their attachment sites (e.g. skin) caused by their opisthaptor (Hayward 2005). Ciliated larvae (=Oncomiracidium) hatch from eggs, infect the vertebrate host and develop into a single adult parasite.
The role of Monogenea as inducer of phenotypic changes of fish can be considered rather insignificant. Among all the literature, only four descriptions of species were found which all belong to the same genus, Gyrodactylus (Table 9.1).
Infections with viviparous Gyrodactylus bullatarudis have been associated with the occurrence of an abnormal swimming behaviour of guppys Poecilia reticulata (Scott 1985). Heavily infected guppies became lethargic and their fins often sticked together. It is suggested that the abnormal swimming behaviour attracts the attention of conspecifics which, in turn, increases the number of contacts between infected and uninfected hosts (Dobson 1988; Scott 1985).
López (1999) examined the effects on mate choice behaviour of female P. reticulata experimentally infected with Gyrodactylus turnbulli. The author observed that infected females were significantly less discriminatory than healthy ones when they were exposed to attractive (showy: higher “individual display rate”, higher “orange brightness”) and unattractive (less showy: lower “individual display rate”, lower “orange brightness”) males in a simultaneous choice test. López (1999) pointed out that although the results point towards an indirect effect of parasitic infections, where females are probably weakened by infection and, as a result, energetically constrained in their mate choice behaviour (i.e. costs for actively resisting sneaky, less attractive males or cooperation in copulation), a direct manipulation by the parasite, thus a selective advantage, could also be the case. Regardless whether a direct or indirect effect of the infection, parasite transmission and reproduction should be enhanced as infected females come into contact with male conspecifics more often than uninfected ones. In an earlier study, Houde and Torio (1992) reported that male guppies infected with G. turnbulli appear to reduce the degree of expression of carotenoid colours (cf. showiness) and that female guppies may be able to avoid becoming infected themselves by discriminating against these infected males. However, avoidance behaviour of females against infected males would result in a decreased transmission probability and thus putting selective pressure on the parasites.
9.3.5 Cestoda
Cestodes (subclass Eucestoda, true tapeworms) are exclusively endoparasitic with more than 5000 known species (Caira and Reyda 2005). Marine cestodes encompass the four orders Diphyllobothriidea, Bothriocephalidea (the latter formerly known as Pseudophyllidea), Trypanorhyncha and Tetraphyllidea (Caira and Reyda 2005; Kuchta et al. 2008), which are characterized by the morphology of the scolex (Möller and Anders 1983). The life cycles of marine cestodes are complex and require three hosts for completion. In the following section, Schistocephalus solidus will be presented as an example for an indirect life cycle. Eggs of S. solidus enter the aquatic system with the faeces of infected birds and hatch and develop to a coracidium, i.e. a motile embryo. After ingestion by cycloid copepods they develop into procercoids, which are then eaten by three-spined sticklebacks. In the fish they develop into plerocercoids. The parasite larvae penetrate through the gut wall and grow in the peritoneal cavity until they reach infectivity at a weight of 50 mg (Tierney et al. 1993). Piscivorous birds feeding on infected sticklebacks serve as final hosts (LoBue and Bell 1993).
A well-studied example for host behavioural changes is the host-parasite interaction between the pseudophyllidean cestode Schistocephalus solidus and his intermediate host, the three-spined stickleback Gasterosteus aculeatus. The rapidly growing parasite induces swelling of the abdomen (Arme and Owen 1967) and places high energy demands on the fish host during the time of parasite growth (Barber and Huntingford 1995). Similar to the effect of a gastric banding the large cestode constricts the stomach and inhibits greater distension (Wright et al. 2006). As a consequence, infected fish prefer smaller, less profitable prey species (Milinski 1984). Furthermore, it has been suggested that the competitive ability might be decreased due to the reduced streamlined shape and smaller energy reserves (Lester 1971; Milinski 1984). In an experiment, Arnott et al. (2000) showed that although energetic demands increased, infected sticklebacks grew faster and had the same or better body condition when being exposed to abundant food resources. Another alteration of behaviour is the willingness to take higher risk under threat of predation (Giles 1983; Godin and Sproul 1988; Milinski 1985). Such ‘boldness’ could be experimentally shown in parasitized G. aculeatus that returned more quickly to feeding behaviour after a stimulated predator attack (Giles 1983; Godin and Sproul 1988).
Another result from energy theft caused by S. solidus is an alteration of fecundity in three-spined sticklebacks revealed by a reduction in clutch size, egg mass, and clutch mass (Heins et al. 2010). As shown by Heins et al. (2010) infected and uninfected female sticklebacks produced egg clutches at the same body size and age, however, their egg masses and egg numbers differed.
Reduced fecundity was also found in cyprinids infected with plerocercoids of Ligula intestinalis (Arme and Owen 1968; Trubiroha et al. 2010). Although both S. solidus and Ligula intestinalis are considered to be castrators with similar life cycles, they differ in the way of altering the host reproduction. In roach (Rutilus rutilus) gonad development can be retarded in males and females at an early stage of gametogenesis, which may constitute an inhibition of host reproduction through endocrine disruption (Trubiroha et al. 2010). This seems to be a parasite strategy which allocates the host’s energy from reproduction to parasitic growth without decreasing host viability (Hurd 2001; Lafferty and Kuris 2009).
9.3.6 Nematoda
The phylum Nematoda represents one of the most species diverse phyla within the Metazoa (Anderson 2000; McClelland 2005). The general nematode life cycle includes four moults and four larval stages. Heteroxeny and paratenesis are common phenomena in the transmission pathways of parasitic species and include intermediate and definitive hosts in nearly every trophic level (McClelland 2005). Because of the dilute nature of aquatic habitats, parasitic nematodes have evolved numerous elaborate means of maintaining themselves in an environment where contact between successive hosts within a life cycle may be periodic or even rare. The use of paratenic hosts, a low host specificity, free living stages that actively search for susceptible hosts as well as the infection of short-lived organisms (e.g. small invertebrates) that occur in vast numbers are only a few examples how nematodes support their temporal and spatial dispersal and increase the likelihood to encounter a successive host (Marcogliese 2005; McClelland 2005). Despite these elaborate means, documentation on parasite induced phenotypic changes of hosts caused by nematode species in aquatic habitats is fairly scarce (Table 9.1). The physical presence of the nematode, e.g. in the musculature of the fish, often results in an impairment of the musculoskeletal system.
The occurrence of certain nematodes (e.g. Anisakidae: Pseudoterranova, Anisakis) in the musculature or internal organs of their (paratenic) intermediate hosts is well documented and has been extensively investigated on a variety of fish, especially those with high commercial value (for recent studies see e.g. Karl et al. 2011: Anisakis; Klapper et al. 2015: Anisakis, Hysterothylacium; Kuhn et al. 2013: Pseudoterranova). Anisakid nematodes have a heteroxenous, aquatic life cycle with vertebrate hosts (pinnipeds, cetaceans, piscivorous birds) as definitive hosts. A broad spectrum of micro- and macroinvertebrates as well as teleost fish serve as intermediate/transport hosts (Mattiucci and Nascetti 2008). Möller and Klatt (1990) found that an infection of smelt Osmerus eperlanus with larvae of the sealworm Pseudoterranova decipiens reduces overall condition of the host and proposed an increasing susceptibility to predation by their definitive hosts, pinnipeds of the families Otariidae and Phocidae. Sprengel and Lüchtenberg (1991) demonstrated that a single sealworm in the musculature of a smelt reduced the swimming speed in a circular experimental chamber by 14.7 % compared to uninfected individuals. Infections with more than three nematodes reduced the average speed by up to 32.2 % and co-infections with the microsporidian Pleistophora ladogensis led to a decrease of 29.9 %, a higher reduction than that in smelt infected by two nematodes or by microsporidians only (Sprengel and Lüchtenberg 1991). The same test was performed for the swim bladder nematode Anguillicoides (Anguillicola) crassus (Fig. 9.7b) which revealed a reduction of swimming speed by 2.9 % (single nematode) to 18.6 % (more than ten nematodes). In this context, Boon et al. (1990) reported decreased quantities of plasma proteins and a lower haematocrit value in experimentally, heavily infected A. anguilla. Sanguivorous activities of the L4 and pre-adult stages decrease the number of circulating erythrocytes and therefore the O2 carrying capacities. The resulting decreased aerobic performance (energy drain) as well as the mechanical damage of the swim bladder caused by the direct invasion of the pre-adult stages impair the swimming performance of the host and increase overall energy consumption (Palstra et al. 2007).
Parasites can also influence migratory behaviour of their hosts as shown by Garnick and Margolis (1990). The authors measured the interference of three cestodes (Eubothrium salvelini, Diphyllobothrium dendriticum, Proteocephalus sp.) and the nematode Philonema oncorhynchi with the orientation of seaward migrating sockeye salmon (Onchorynchus nerka) smolts and suggested that parasite infections may account, at least to some extent, for the variability observed in migratory behaviour of smolts (Garnick and Margolis 1990).
Whilst a reduced swimming speed of the intermediate host is advantageous for the transmission of P. decipiens into the next host, there seems to be no such benefit for A. crassus and P. onchorhynchi as both are infecting their final hosts.
9.3.7 Crustacea
Crustaceans comprise a species-rich taxon of metazoan parasites that parasitize a wide range of fishes and invertebrate hosts (Fig. 9.7c, d) (Busch et al. 2012). There are seven groups of crustaceans in which parasitic forms occur: Copepoda (infecting sponges, cnidarians, echinoderms, fishes and mammals), Isopoda (mostly ectoparasitic on crustaceans and fish), Branchiura (only fish lice of the genus Argulus in the marine environment), Tantulocarida (ectoparasites of other crustaceans), Cirripedia including Thoracica (parasitic on dogfish and polychaetes) and Rhizocephala (infecting other crustaceans, mainly Decapoda), Ascothoracida (parasitizing echinoderms and cnidarians), and Amphipoda (suborder Hyperiidea on gelatinous zooplankton and Caprellidea on whales). The different groups are described in detail in Rohde (2005) and only life cycles of the main fish parasite groups, namely copepods, isopods, and branchiurans are summarized in the following section.
The life cycle of copepods, a species-rich group of metazoan ectoparasites, is mostly direct and comprises two phases, a naupliar stage with a maximum of six stages and the copepodid stage with a maximum of five stages (Boxshall 2005a). Most parasitic copepods extrude their eggs into paired egg sacs or uniseriate egg strings. Sexual dimorphism is common with females typically larger and more transformed bodies compared to dwarf males (e.g. Chondracanthidae, Lernaeopodidae) (Boxshall 2005a). Eggs of Isopoda hatch in a brood pouch called ‘marsipium’ where they moult to the ‘manca’ or ‘pullus II’ stage (Lester 2005). Mancae larvae are then released to find an intermediate fish host to take their first meal. In the isopod genus Anilocra the mancae leave the optional intermediate host, has to find a new fish host, moults, and this is repeated until they settle on the final (fish) host, while the mancae of other species e.g. gill-inhabiting forms or tong-biters and tissue dwellers attach only to one final host (Fogelman and Grutter 2008; Lester 2005). Cymothoids are protandrous hermaphrodites, i.e. the first male that infects a host changes its sex into female, whereas all following males remain males (Lester 2005). In the life-cycle of the branchiuran Argulus (Fig. 9.7d), a mature female leaves its host after taking a meal, lays eggs on any hard submerged surface, these eggs hatch, and the free-swimming larvae moult into the second stage which is parasitic as all other subsequent stages (Boxshall 2005b). They change their host at intervals throughout the development until they reach maturation at the fifth larval stage (Boxshall 2005b).
Lernaeocera branchialis (Fig. 9.7c) is a parasitic copepod that infects the gills or mantle cavity of various gadoid species in its adult phase (Kabata 1970). The two-host life cycle involves two nauplius stages, after them copepod larvae parasitize an intermediate fish host on which they mature and mate. The mated adult female leaves the host and has to find a second fish or sometimes a marine mammal as definitive host (Boxshall 2005a). Parasitisation of cod Gadus morhua with L. branchialis impacts on growth and mortality (Khan 1988). In an experiment young fish infected with L. branchialis fed less, were smaller, had a lower weight, and showed reduced fecundity. Additionally, they frequently swam close to the surface probably due to an excess of air in the swim bladder and displayed hyperactivity (Khan 1988).
Parasite-induced alterations in swimming behaviour were detected in brook trout fry Salvelinus fonatlis infected with the copepod Salmincola edwardsii (Poulin et al. 1991). In an experiment brook trout stayed more time motionless when exposed to Salmincola edwardsii, and this decreases the chance to be attacked by the parasite because the parasites reacts to stimuli such as passing shadows. After an initial parasite infection, the time spent motionless in the water is diminished and the fish increases the risk of acquiring further parasite infections.
Sticklebacks (Gasterosteus spp.) are parasitized by the branchiuran ectoparasite Argulus canadensis, which is a free-living, blood-sucking parasite (Poulin and FitzGerald 1989). Three-spined and black-spotted sticklebacks form larger shoals when the parasite is present in the water and this behaviour was attributed to be selective by lowering the risk for an individual fish to become a target of the branchiuran parasite waiting for suitable hosts to pass (Poulin and FitzGerald 1989).
Cymothoid isopods are usually ectoparasites attached to the skin, gills, or oral cavity (Bunkley-Williams and Williams 1998). Cymothoids have been found to reduce growth, motility, and reproduction (Adlard and Lester 1995; Fogelman et al. 2009; Östlund-Nilsson et al. 2005). In the Great Barrier Reef, cardinal fish Cheilodipterus quinquelineatus is regularly parasitized with Anilocra apogonae, a large, asymmetrically cymothoid isopod attached on the skin of its host. Besides the need to increase food intake, a decreased swimming speed and endurance of the fish host was noted which might be have been a consequence of the relative increase in size and asymmetric attachment of the parasite negatively impacting the host’s hydrodynamics (Östlund-Nilsson et al. 2005). The authors concluded that the energetic stress must be compensated for by increasing foraging, which may at the same time lead to decreased predator avoidance behaviour. Additionally, A. apogonae may act as a parasitic castrator parasitized fish were observed to have smaller gonads, fail to mouth-brood (males), or have fewer and smaller ova (female) (Fogelman et al. 2009).
A further example of a cymothoid isopod as parasitic castrator is Riggia paranensis. After burrowing into the flesh of freshwater fish Cyphocharax gilbert, infections lead to increased growth, but inhibition of the gonadal development through affection of the endocrine system (Azevedo et al. 2006; Lima et al. 2007).
9.4 Concluding Remarks
Regardless of whether caused directly or indirectly, and whether adaptive or a by-product, parasites have a great capability of manipulating host behaviour. Considering only fish as hosts, most examples of parasites altering fish host behaviour are known from the group Digenea and the most common manipulation involves host motility. However, a differentiation of manipulative behavioural changes (direct and indirect) as well as the isolation of specific mechanisms has proven to be difficult. Another problem is that lab studies with freshwater specimens are largely overrepresented reflecting the difficulty of undertaking research in this subject with marine and field studies clearly being much less feasible. Similarly, results might only be valid under controlled conditions and their relevance in the field needs to be considered carefully.
References
Adlard R, Lester R (1995) The life-cycle and biology of Anilocra pomacentri (Isopoda, Cymothoidae), an ectoparasitic isopod of the coral-reef fish, Chromis nitida (Perciformes, Pomacentridae). Aust J Zool 43:271–281
Anderson RC (2000) Nematode parasites of vertebrates: their development and transmission. CABI Publishing, Wallingford, 676 pp
Andersson M (1994) Sexual selection. Princeton University Press, Princeton, 624 pp
Arme C, Owen RW (1967) Infections of the three-spined stickleback, Gasterosteus aculeatus L., with the plerocercoid larvae of Schistocephalus solidus (Muller, 1776), special reference to pathological effects. Parasitology 57:301–314
Arme C, Owen RW (1968) Occurrence and pathology of Ligula intestinalis infections in British fishes. J Parasitol 54:272–280
Arnott SA, Barber I, Huntingford FA (2000) Parasite–associated growth enhancement in a fish–cestode system. Proc R Soc Lond B Biol Sci 267:657–663
Bakker TCM, Mazzi D, Zala S (1997) Parasite-induced changes in behavior and color make Gammarus pulex more prone to fish predation. Ecology 78:1098
Barber I (2007) Parasites, behaviour and welfare in fish. Appl Anim Behav Sci 104:251–264
Barber I, Huntingford FA (1995) The effect of Schistocephalus solidus (Cestoda: Pseudophyllidea) on the foraging and shoaling behaviour of three-spined sticklebacks, Gasterosteus aculeatus. Behaviour 132:1223–1240
Barber I, Wright HA (2005) Effects of parasites on fish behaviour: interactions with host physiology. In: Wilson RW, Balshine S, Sloman KA (eds) Fish physiology: behaviour and physiology of fish. Elsevier Academic Press, Amsterdam/London, pp 109–149
Barber I, Hoare D, Krause J (2000) Effects of parasites on fish behaviour: a review and evolutionary perspective. Rev Fish Biol Fish 10:131–165
Baudoin M (1975) Host castration as a parasitic strategy. Evolution 29:335–352
Beaver PC (1939) The morphology and life history of Psilostomum ondatrae Price, 1931 (Trematoda: Psilostomidae). J Parasitol 25:383–393
Bernier NJ (2006) The corticotropin-releasing factor system as a mediator of the appetite-suppressing effects of stress in fish. Gen Comp Endocrinol 146:45–55
Bernier NJ (2010) Food intake regulation and disorders. In: Leatherland JF, Woo PTK (eds) Fish diseases and disorders, vol 2, Non-infectious disorders. CAB International, Wallingford/Cambridge, pp 238–266
Binning SA, Barnes JI, Davies JN, Backwell PRY, Keogh JS, Roche DG (2014) Ectoparasites modify escape behaviour, but not performance, in a coral reef fish. Anim Behav 93:1–7
Boon JH, Cannaerts VMH, Augustijn H, Machiels MAM, De Charleroy D, Ollevier F (1990) The effect of different infection levels with infective larvae of Anguillicola crassus on haematological parameters of European eel (Anguilla anguilla). Aquaculture 87:243–253
Boxshall GA (2005a) Copepoda (copepods). In: Rohde K (ed) Marine parasitology. CABI Publishing, Wallingford, pp 123–138
Boxshall GA (2005b) Branchiura (branchurians). In: Rohde K (ed) Marine parasitology. CABI Publishing, Wallingford, pp 145–147
Boyce NP (1979) Effects of Eubothrium salvelini (Cestoda: Pseudophyllidea) on the growth and vitality of sockeye salmon, Oncorhynchus nerka. Can J Zool 57:597–602
Brassard P, Curtis MA, Rau ME (1982) Seasonality of Diplostomum spathaceum (Trematoda: Strigeidae) transmission to brook trout (Salvelinus fontinalis) in northern Quebec, Canada. Can J Zool 60:2258–2263
Bunkley-Williams L, Williams JEH (1998) Isopoda associated with fishes: a synopsis and corrections. J Parasitol 84:893–896
Busch MW, Kuhn T, Münster J, Klimpel S (2012) Marine crustaceans as potential hosts and vectors for metazoan parasites. In: Mehlhorn H (ed) Arthropods as vectors of emerging diseases. Springer, Berlin Heidelberg, pp 329–360
Butler JA, Millemann RE (1971) Effect of the “salmon poisoning” trematode, Nanophyetus salmincola, on the swimming ability of juvenile salmonid fishes. J Parasitol 57:860–865
Caira JN, Reyda FB (2005) Eucestoda (true tapeworms). In: Rohde K (ed) Marine parasitology. CABI Publishing, Wallingford, pp 92–104
Cézilly F, Thomas F, Médoc V, Perrot-Minnot M-J (2010) Host-manipulation by parasites with complex life cycles: adaptive or not? Trends Parasitol 26:311–317
Chin A, Guo FC, Bernier NJ, Woo PT (2004) Effect of Cryptobia salmositica-induced anorexia on feeding behavior and immune response in juvenile rainbow trout Oncorhynchus mykiss. Dis Aquat Organ 58:17–26
Coleman FC (1993) Morphological and physiological consequences of parasites encysted in the bulbus arteriosus of an estuarine fish, the sheepshead minnow, Cyprinodon variegatus. J Parasitol 79:247
Combes C (1991) Ethological aspects of parasite transmission. Am Nat 138:866–880
Consuegra S, Garcia de Leaniz C (2008) MHC-mediated mate choice increases parasite resistance in salmon. Proc R Soc B Biol Sci 275:1397–1403
Cribb TH (2005) Digenea (endoparasitic flukes). In: Rohde K (ed) Marine parasitology. CABI Publishing, Wallingford, pp 76–87
Crowden AE, Broom DM (1980) Effects of the eyefluke, Diplostomum spathaceum, on the behaviour of dace (Leuciscus leuciscus). Anim Behav 28:287–294
Dence WA (1958) Studies on Ligula-infected common shiners (Notropis cornutus frontalis Agassiz) in the Adirondacks. J Parasitol 44:334–338
de Souza Azevedo J, da Silva LG, Bizerri CRSF, Dansa-Petretski MA, Lima NRW (2006) Infestation pattern and parasitic castration of the crustacean Riggia paranensis (Crustacea: Cymothoidea) on the fresh water fish Cyphocharax gilbert (Teleostei: Curimatidae). Neotropical Ichthyol 4:363–369
Dobson AP (1988) The population biology of parasite-induced changes in host behavior. Q Rev Biol 63:139–165
Eizaguirre C, Lenz TL, Kalbe M, Milinski M (2012) Rapid and adaptive evolution of MHC genes under parasite selection in experimental vertebrate populations. Nat Commun 3:621
El-Matbouli M, Hoffmann RW (1998) Light and electron microscopic studies on the chronological development of Myxobolus cerebralis to the actinosporean stage in Tubifex tubifex. Int J Parasitol 28:195–217
El-Matbouli M, Hoffmann RW, Schoel H et al (1999) Whirling disease: host specificity and interaction between the actinosporean stage of Myxobolus cerebralis and rainbow trout Oncorhynchus mykiss. Dis Aquat Org 35:1–12
Ferguson MS, Hayford RA (1941) The life history and control of an eye fluke: an account of a serious hatchery disease caused by a parasitic worm. Progress Fish-Cult 8:1–13
Fogelman RM, Grutter AS (2008) Mancae of the parasitic cymothoid isopod, Anilocra apogonae: early life history, host-specificity, and effect on growth and survival of preferred young cardinal fishes. Coral Reefs 27:685–693
Fogelman RM, Kuris AM, Grutter AS (2009) Parasitic castration of a vertebrate: effect of the cymothoid isopod, Anilocra apogonae, on the five-lined cardinalfish, Cheilodipterus quinquelineatus. Int J Parasitol 39:577–583
Garnick E, Margolis L (1990) Influence of four species of helminth parasites on orientation of seaward migrating sockeye salmon (Oncorhynchus nerka) smolts. Can J Fish Aquat Sci 47:2380–2389
Gilbert MA, Granath WO (2001) Persistent infection of Myxobolus cerebralis, the causative agent of salmonid whirling disease, in Tubifex tubifex. J Parasitol 87:101–107
Gilbert MA, Granath WO (2003) Whirling disease of salmonid fish: life cycle, biology, and disease. J Parasitol 89:658–667
Giles N (1983) Behavioural effects of the parasite Schistocephalus solidus (Cestoda) on an intermediate host, the three-spined stickleback, Gasterosteus aculeatus L. Anim Behav 31:1192–1194
Godin J-GJ, Sproul CD (1988) Risk taking in parasitized sticklebacks under threat of predation: effects of energetic need and food availability. Can J Zool 66:2360–2367
Guthrie JF, Kroger RL (1974) Schooling habits of injured and parasitized menhaden. Ecology 55:208–210
Hall SR, Becker C, Caceres CE (2007) Parasitic castration: a perspective from a model of dynamic energy budgets. Integr Comp Biol 47:295–309
Hamilton WD, Zuk M (1982) Heritable true fitness and bright birds: a role for parasites? Science 218:384–387
Hayward C (2005) Monogenea Polyopisthocotylea (ectoparasitic flukes). In: Rohde K (ed) Marine parasitology. CABI Publishing, Wallingford, pp 55–63
Hedrick RP, Adkison MA, El-Matbouli M, MacConnell E (1998) Whirling disease: re-emergence among wild trout. Immunol Rev 166:365–376
Heins DC, Baker JA, Toups MA, Birden EL (2010) Evolutionary significance of fecundity reduction in threespine stickleback infected by the diphyllobothriidean cestode Schistocephalus solidus. Biol J Linn Soc 100:835–846
Herting GE, Witt A (1967) The role of physical fitness of forage fishes in relation to their vulnerability to predation by Bowfin (Amia calva). T Am Fish Soc 96:427–430
Houde AE, Torio AJ (1992) Effect of parasitic infection on male color pattern and female choice in guppies. Behav Ecol 3:346–351
Hurd H (1990) Physiological and behavioural interactions between parasites and invertebrate hosts. Adv Parasitol 29:271–318
Hurd H (2001) Host fecundity reduction: a strategy for damage limitation? Trends Parasitol 17:363–368
Jones SRM, Woo PTK, Stevenson RMW (1986) Immunosuppression in rainbow trout, Salmo gairdneri Richardson, caused by the haemoflagellate Cryptobia salmositica Katz, 1951. J Fish Dis 9:431–438
Kabata Z (1970) Diseases of fishes: Crustacea as enemies of fishes. T.F.H. Publishers, Jersey City, 171 pp
Karl H, Baumann F, Ostermeyer U, Kuhn T, Klimpel S (2011) Anisakis simplex (ss) larvae in wild Alaska salmon: no indication of post-mortem migration from viscera into flesh. Dis Aquat Org 94:201
Kent ML, Andree KB, Bartholomew JL, El-Matbouli M, Desser SS, Devlin RH, Feist SW, Hedrick RP, Hoffmann RW, Khattra J, Hallett SL, Lester RJG, Longshaw M, Palenzeula O, Siddall ME, Xiao C (2001) Recent advances in our knowledge of the Myxozoa. J Eukaryot Microbiol 48:395–413
Khan RA (1988) Experimental transmission, development, and effects of a parasitic copepod, Lernaeocera branchialis, on Atlantic cod, Gadus morhua. J Parasitol 74:586
Klapper R, Kuhn T, Münster J, Levsen A, Karl H, Klimpel S (2015) Anisakid nematodes in beaked redfish (Sebastes mentella) from three fishing grounds in the North Atlantic, with special notes on distribution in the fish musculature. Vet Parasitol 207:72–80
Klimpel S, Busch MW, Kellermanns E, Kleinertz S, Palm HW (2009) Metazoan deep-sea fish parasites. Natur u Wissenschaft, Solingen, 384 pp
Krause J (1994) The influence of food competition and predation risk on size-assortative shoaling in juvenile chub (Leuciscus cephalus). Ethology 96:105–116
Krause J, Godin J-GJ (1994a) Shoal choice in the banded killifish (Fundulus diaphanus, Teleostei, Cyprinodontidae): effects of predation risk, fish size, species composition and size of shoals. Ethology 98:128–136
Krause J, Godin J-GJ (1994b) Influence of parasitism on the shoaling behaviour of banded killifish, Fundulus diaphanus. Can J Zool 72:1775–1779
Krause J, Godin J-GJ (1996) Influence of parasitism on shoal choice in the banded killifish (Fundulus diaphanus, Teleostei, Cyprinodontidae). Ethology 102:40–49
Kuchta R, Scholz T, Brabec J, Bray RA (2008) Suppression of the tapeworm order Pseudophyllidea (Platyhelminthes: Eucestoda) and the proposal of two new orders, Bothriocephalidea and Diphyllobothriidea. Int J Parasitol 38:49–55
Kuhn T, Benninghoff T, Karl H, Landry T, Klimpel S (2013) Sealworm Pseudoterranova decipiens ss infection of European smelt Osmerus eperlanus in German coastal waters: ecological implications. Dis Aquat Organ 102:217–224
Kuris AM (1997) Host behavior modification: an evolutionary perspective. In: Beckage NE (ed) Parasites and pathogens. Chapman & Hall, New York, pp 293–315
Lafferty KD (1997) The ecology of parasites in a salt marsh ecosystem. In: Beckage NE (ed) Parasites and Pathogens. Springer, US, pp 316–332
Lafferty KD (1999) The evolution of trophic transmission. Parasitol Today 15:111–115
Lafferty KD, Kuris AM (2009) Parasitic castration: the evolution and ecology of body snatchers. Trends Parasitol 25:564–572
Lafferty KD, Morris AK (1996) Altered behavior of parasitized killifish increases susceptibility to predation by bird final hosts. Ecology 77:1390–1397
Lafferty KD, Shaw JC (2013) Comparing mechanisms of host manipulation across host and parasite taxa. J Exp Biol 216:56–66
Lester RJG (1971) The influence of Schistocephalus plerocercoids on the respiration of Gasterosteus and a possible resulting effect on the behavior of the fish. Can J Zool 49:361–366
Lester RJG (2005) Isopoda (isopods). In: Rohde K (ed) Marine parasitology. CABI Publishing, Wallingford, pp 138–144
Li S, Woo PT (1991) Anorexia reduces the severity of cryptobiosis in Oncorhynchus mykiss. J Parasitol 77:467–471
Lima NR, de Souza Azevedo J, da Silva LG, Dansa-Petretski M (2007) Parasitic castration, growth, and sex steroids in the freshwater bonefish Cyphocharax gilbert (Curimatidae) infested by Riggia paranensis (Cymothoidea). Neotropical Ichthyol 5:471–478
LoBue CP, Bell MA (1993) Phenotypic manipulation by the cestode parasite Schistocephalus solidus of its intermediate host, Gasterosteus aculeatus, the threespine stickleback. Am Nat 142:725–735
López S (1999) Parasitized female guppies do not prefer showy males. Anim Behav 57:1129–1134
Lyholt HCK, Buchmann K (1996) Diplostomum spathaceum: effects of temperature and light on cercarial shedding and infection of rainbow trout. Dis Aquat Organ 25:169–173
MacKenzie K (1999) Parasites as pollution indicators in marine ecosystems: a proposed early warning system. Mar Pollut Bull 38:955–959
Macnab V, Barber I (2012) Some (worms) like it hot: fish parasites grow faster in warmer water, and alter host thermal preferences. Glob Change Biol 18:1540–1548
MacKenzie K, Kalavati C (2014) Myxosporean parasites of marine fishes: their distribution in the world’s oceans. Parasitology 141:1709–1717
Marcogliese DJ (2005) Transmission of marine parasites. In: Rohde K (ed) Marine parasitology. CABI Publishing, Wallingford, pp 280–286
Mattiucci S, Nascetti G (2008) Advances and trends in the molecular systematics of anisakid nematodes, with implications for their evolutionary ecology and host—parasite co-evolutionary processes. Adv Parasitol 66:47–148
McClelland G (2005) Nematoda (roundworms). In: Rohde K (ed) Marine parasitology. CABI Publishing, Wallingford, pp 104–115
Mikheev VN, Pasternak AF, Taskinen J, Valtonen ET (2010) Parasite-induced aggression and impaired contest ability in a fish host. Parasit Vectors 3:17
Milinski M (1984) Parasites determine a predator’s optimal feeding strategy. Behav Ecol Sociobiol 15:35–37
Milinski M (1985) Risk of predation of parasitized sticklebacks (Gasterosteus aculeatus L.) under competition for food. Behaviour 93:203–216
Milinski M, Bakker TCM (1990) Female sticklebacks use male coloration in mate choice and hence avoid parasitized males. Nature 344:330–333
Moles A, Heifetz J (1998) Effects of the brain parasite Myxobolus arcticus on sockeye salmon. J Fish Biol 52:146–151
Möller H, Anders K (1983) Krankheiten und Parasiten der Meeresfische. Verlag Heino Möller, Kiel, 258 pp
Möller H, Anders K (1986) Diseases and parasites of marine fishes. Verlag Heino Möller, Kiel, 365 pp
Möller H, Klatt S (1990) Smelt as host of the sealworm (Pseudoterranova decipiens) in the Elbe estuary. Can Bull Fish Aquat Sci 222:129–138
Moodie E (2005) Microsporidia (microsporans). In: Rohde K (ed) Marine parasitology. CABI Publishing, Wallingford, pp 30–34
Moore J (1984) Altered behavioral responses in intermediate hosts - an Acanthoceptalan parasite strategy. Am Nat 123:572–577
Moore J (2002) Parasites and the behavior of animals. Oxford University Press, New York, 338 pp
Moore J (2013) An overview of parasite-induced behavioral alterations - and some lessons from bats. J Exp Biol 216:11–17
Moore J, Gotelli NJ (1990) A phylogenetic perspective on the evolution of altered host behaviours: A critical look at the manipulation hypothesis. In: Barnard CJ, Behnke JM (eds) Parasitism and host behaviour. Taylor & Francis, London, pp 193–233
Mouritsen KN, Poulin R (2003) Parasite-induced trophic facilitation exploited by a non-host predator: a manipulator’s nightmare. Int J Parasitol 33:1043–1050
Ness JH, Foster SA (1999) Parasite-associated phenotype modifications in threespine stickleback. Oikos 85:127
O’Donoghue P (2005) Protistan biodiversity. In: Rohde K (ed) Marine parasitology. CABI Publishing, Wallingford, pp 11–17
Orr TSC (1966) Spawning behaviour of rudd, Scardinius erythrophthalmus infested with plerocercoids of Ligula intestinalis. Nature 212:736–736. 11
Östlund-Nilsson S, Curtis L, Nilsson G, Grutter A (2005) Parasitic isopod Anilocra apogonae, a drag for the cardinal fish Cheilodipterus quinquelineatus. Mar Ecol Prog Ser 287:209–216
Palstra AP, Heppener DFM, Van Ginneken VJT, Székely C, Van den Thillart G (2007) Swimming performance of silver eels is severely impaired by the swim-bladder parasite Anguillicola crassus. J Exp Mar Biol Ecol 352:244–256
Poulin R (1993) Age-dependent effects of parasites on anti-predator responses in two New Zealand freshwater fish. Oecologia 96:431–438
Poulin R (1994) Meta-analysis of parasite-induced behavioural changes. Anim Behav 48:137–146
Poulin R (1995) “Adaptive” changes in the behaviour of parasitized animals: a critical review. Int J Parasitol 25:1371–1383
Poulin R (2010) Parasite manipulation of host behavior: an update and frequently asked questions. In: Brockmann HJ (ed) Advances in the study of behavior. Elsevier, Amsterdam, pp 151–186
Poulin R, FitzGerald GJ (1989) Shoaling as an anti-ectoparasite mechanism in juvenile sticklebacks (Gasterosteus spp.). Behav Ecol Sociobiol 24:251–255
Poulin R, Morand S (2004) Parasite biodiversity. Smithsonian Institution Scholarly Press, Washington, DC, 216 pp
Poulin R, Thomas F (1999) Phenotypic variability induced by parasites: extent and evolutionary implications. Parasitol Today 15:28–32
Poulin R, Curtis MA, Rau ME (1991) Size, behaviour, and acquisition of ectoparasitic copepods by brook trout, Salvelinus fontinalis. Oikos 61:169
Radabaugh DC (1980a) Changes in minnow, Pimephales promelas Rafinesque, schooling behaviour associated with infections of brainencysted larvae of the fluke, Ornithodiplostomum ptychocheilus. J Fish Biol 16:621–628
Radabaugh DC (1980b) Encystment site selection in the brain-inhabiting metacercariae of Ornithodiplostomum ptychocheilus (Trematoda: Strigeoidea). J Parasitol 66:183–184
Rees G (1957) Cercaria diplostomi phoxini (Faust), a furcocercaria which develops into Diplostomulum phoxini in the brain of the minnow. Parasitology 47:126–137
Reusch TB, Häberli MA, Aeschlimann PB, Milinski M (2001) Female sticklebacks count alleles in a strategy of sexual selection explaining MHC polymorphism. Nature 414:300–302
Rohde K (2005) Marine parasitology. CABI Publishing, Wallingford, p 565
Rose JD, Marrs GS, Lewis C, Schisler G (2000) Whirling disease behavior and its relation to pathology of brain stem and spinal cord in rainbow trout. J Aquat Anim Health 12:107–118
Rossiter W (2013) Current opinions: zeros in host–parasite food webs: are they real? Int J Parasitol Parasites Wildl 2:228–234
Santos EGN, Cunha RA, Santos Portes C (2011) Behavioral responses of Poecilia vivipara (Osteichthyies: Cyprinodontiformes) to experimental infections of Acanthocollaritrema umbilicatum (Digenea: Cryptogonimidae). Exp Parasitol 127:522–526
Santos EGN, Santos Portes C (2013) Parasite-induced and parasite development-dependent alteration of the swimming behavior of fish hosts. Acta Tropica 127:56–62
Sasal P, Thomas F (2005) Parasite induced changes in host behaviour and morphology. In: Rohde K (ed) Marine parasitology. CABI Publishing, Wallingford, pp 259–263
Scott M (1985) Experimental epidemiology of Gyrodactylus bullatarudis (Monogenea) on guppies (Poecilia reticulata): short- and long-term studies. In: Rollinson E, Anderson RM (eds) Ecology and genetics of host-parasite interactions. Academic, London, pp 21–38
Seppälä O, Karvonen A, Tellervo Valtonen E (2004) Parasite-induced change in host behaviour and susceptibility to predation in an eye fluke–fish interaction. Anim Behav 68:257–263
Seppälä O, Valtonen ET, Benesh DP (2008) Host manipulation by parasites in the world of dead-end predators: adaptation to enhance transmission? Proc R Soc B Biol Sci 275:1611–1615
Shirakashi S, Goater CP (2002) Intensity-dependent alteration of minnow (Pimephales promelas) behavior by a brain- encysting trematode. J Parasitol 88:1071–1074
Shirakashi S, Goater CP (2005) Chronology of parasite-induced alteration of fish behaviour: effects of parasite maturation and host experience. Parasitology 130:177–183
Smith JW (1972) The blood flukes (Digenea: Sanguinicolidae and Spirorchidae) of cold-blooded vertebrates and some comparison with the schistosomes. Helminthol Abstr 41:161–204
Smith RS, Kramer DL (1987) Effects of a cestode (Schistocephalus sp.) on the response of ninespine sticklebacks (Pungitius pungitius) to aquatic hypoxia. Can J Zool 65:1862–1865
Smothers JF, von Dohlen C, Smith LH, Spall RD (1994) Molecular evidence that the myxozoan protists are metazoans. Science 265:1719–1721
Sprengel G, Lüchtenberg H (1991) Infection by endoparasites reduces maximum swimming speed of European smelt Osmerus eperlanus and European eel Anguilla anguilla. Dis Aquat Organ 11:31–35
Steinbach Elweell LC, Eagle Stromberg K, Ryce EKN, Bartholomew JL (2009) Whirling disease in the United States. A summary of progress in research and management. Montana Water Center. Bozeman
Swearer SE, Robertson DR (1999) Life history, pathology, and description of Kudoa ovivora n. sp. (Myxozoa, Myxosporea): an ovarian parasite of Caribbean labroid fishes. J Parasitol 85:337–353
Sweeting RA (1976) Studies on Ligula intestinalis (L.) effects on a roach population in a gravel pit. J Fish Biol 9:515–522
Sweeting RA (1977) Studies on Ligula intestinalis Some aspects of the pathology in the second intermediate host. J Fish Biol 10:43–50
Szidat L (1969) Structure, development, and behaviour of new Strigeatoid metacercariae from subtropical fishes of South America. J Fish Res Bd Can 26:753–786
Thomas PT, Woo PTK (1992) Anorexia in rainbow trout, Oncorhynchus mykiss (Walbaum), infected with Cryptobia salmositica (Sarcomastigophora: Kinetoplastida): its onset and contribution to the immunodepression. J Fish Dis 15:443–447
Thomas F, Adamo S, Moore J (2005) Parasitic manipulation: where are we and where should we go? Behav Processes 68:185–199
Thomas F, Rigaud T, Brodeur J (2012) Evolutionary routes leading to host manipulation by parasites. In: Hughes DP, Brodeur J, Thomas F (eds) Host manipulation by parasites. Oxford University Press, Oxford, pp 16–33
Tierney JF, Huntingford FA, Crompton DWT (1993) The relationship between infectivity of Schistocephalus solidus (Cestoda) and anti-predator behaviour of its intermediate host, the three-spined stickleback, Gasterosteus aculeatus. Anim Behav 46:603–605
Trubiroha A, Kroupova H, Wuertz S, Frank SN, Sures B, Kloas W (2010) Naturally-induced endocrine disruption by the parasite Ligula intestinalis (Cestoda) in roach (Rutilus rutilus). Gen Comp Endocrinol 166:234–240
Ward AJ, Duff AJ, Krause J, Barber I (2005) Shoaling behaviour of sticklebacks infected with the microsporidian parasite, Glugea anomala. Environ Biol Fishes 72:155–160
Weissenberg R (1968) Intracellular development of the microsporidan Glugea anomala Moniez in hypertrophying migratory cells of the fish Gasterosteus aculeatus L., an example of the formation of “xenoma” tumors. J Protozool 15:44–57
Whittington I (2005) Monogenea Monopisthocotylea (ectoparasitic flukes). In: Rohde K (ed) Marine parasitology. CABI Publishing, Wallingford, pp 63–72
Windsor DA (1998) Most of the species on Earth are parasites. Int J Parasitol 28:1939–1941
Wolf K, Markiw ME, Hiltunen JK (1986) Salmonid whirling disease: Tubifex tubifex (Müller) identified as the essential oligochaete in the protozoan life cycle. J Fish Dis 9:83–85
Woo PTK (1994) Flagellate parasites of fish. In: Kreier JP (ed) Parasitic protozoa, vol 8, 2nd edn. Academic, London, pp 1–80
Woo PTK (2011) Cryptobia (Trypanoplasma) salmositica. In: Woo PTK, Buchmann K (eds) Fish parasites: pathobiology and protection. CABI Publishing, Wallingford, pp 30–54
Wright HA, Wootton RJ, Barber I (2006) The effect of Schistocephalus solidus infection on meal size of three-spined stickleback. J Fish Biol 68:801–809
Yokoyama H, Grabner D, Shirakashi S (2012) Transmission biology of the Myxozoa. In: Carvalho E (ed) Health and environment in aquaculture. INTECH Open Access Publisher, Rijeka, Croatia
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2015 Springer International Publishing Switzerland
About this chapter
Cite this chapter
Kuhn, T., Klapper, R., Münster, J., Dörge, D.D., Kochmann, J., Klimpel, S. (2015). Remote Control: Parasite Induced Phenotypic Changes in Fish. In: Mehlhorn, H. (eds) Host Manipulations by Parasites and Viruses. Parasitology Research Monographs, vol 7. Springer, Cham. https://doi.org/10.1007/978-3-319-22936-2_9
Download citation
DOI: https://doi.org/10.1007/978-3-319-22936-2_9
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-22935-5
Online ISBN: 978-3-319-22936-2
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)