
Abstract:
Methanogenesis in hypersaline environments is determined by redox potential and permanency of anaerobic conditions, and by the concentration of other terminal electron acceptors, particularly sulfate, because sulfate-reducing bacteria have a greater affinity than methanogens for competitive substrates like hydrogen and acetate. Hypersalinity, however, is not an obstacle to methanogenesis; in many cases it provides higher concentrations of non-competitive substrates like methylamines, which derive from compatible solutes such as glycine-betaine that is synthesized by many microbes inhabiting hypersaline environments. Also, depletion of sulfate, as may occur in deeper sediments, allows increased methanogenesis. On the other hand, increasing salinity requires methanogens to synthesize or take up more compatible solutes at a significant energetic cost. Aceticlastic and hydrogenotrophic methanogens, with their lower energetic yields, are therefore more susceptible than methylotrophic methanogenesis, which further explains the predominance of methylotrophic methanogens like Methanohalophilus spp. in hypersaline environments. There are often deviations from the picture outlined above, which are sometimes difficult to explain. Identifying the role of uncultivated Euryarchaeota in hypersaline environments, elucidating the effects of different ions (which have differential stress effects and potential as electron acceptors) and understanding the effects of salinity on all microbes involved in methane cycling, will help us to understand and predict methane production in hypersaline environments.
Access provided by Autonomous University of Puebla. Download reference work entry PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
Hypersaline environments are simply defined as those with a greater concentration of salts than seawater. Such environments are many and varied, in terms of their overall salinity and predominant ions. Coastal environments, both man-made and natural are subject to desiccation, resulting in a wide variety of habitats from small, ephemeral salt pans within temperate salt marshes to large, permanently hypersaline sabkhas in sub-tropical regions (Hovorka, 1987). Similarly, inland salt lakes can be as large as the Great Salt Lake or a tiny spring. Salt deposits, often several hundred meters in thickness, lie beneath about a quarter of the Earth’s landmass, and contain brines from a cubic micrometer in volume to many cubic meters.
Hypersaline environments are widespread and were more prevalent in former geological times, for example much of northern Europe was covered by the salt-saturated Zechstein Sea during the Permo-Triassic period (Zharkov, 1981), and the Mediterranean Sea was desiccated more recently, with halite precipitation starting between 5.6 and 5.55 million years ago (Hsü et al., 1973). Deep-sea, anoxic, hypersaline brines, derived from dissolution of such ancient evaporites, form large lakes on the floor of the Gulf of Mexico, Mediterranean and Red Sea. Oil reservoirs are frequently associated with hypersaline environments, and many industrial waste streams are both anaerobic and hypersaline.
The inhabitants of hypersaline environments are generally termed halophiles, and the use and misuse of this term, together with all its qualifiers, as well as examples of the most ecologically important extreme halophiles, have been discussed by Oren (2008). The ability of microbes to tolerate hypersaline environments with different chemical compositions varies widely: Don Juan Pond, a CaCl2-saturated brine, appears to support no life (see Oren, 2002), whereas the African soda lakes are amongst the most productive environments in the world (Grant and Tindall, 1986). Salinity was found to be the main factor influencing microbial community composition in a recent synthesis of 111 studies (Lozupone and Knight, 2007), and contrary to popular perception, microbial diversity can be extremely high in environments where the salinity is about two to three times higher than seawater and where redox and light gradients exist. For example, 42 of the main bacterial phyla and 15 novel candidate phyla were reported in a microbial mat in Guerrero Negro with a salinity of 8% (Ley et al., 2006), representing an unprecedented level of microbial diversity (Lozupone and Knight, 2007). Also, where salinity gradients occur, microbial biodiversity, abundance and activity can also be greatly elevated owing to a cocktail of electron acceptors, donors, nutrients and carbon sources in the ionic and redox gradient (Daffonchio et al., 2006).
In most environments methanogens are in competition with sulfate-reducing bacteria for the products of fermentation, particularly hydrogen and acetate, and it is well documented that where sulfate concentrations are sufficiently high, e.g., saline and hypersaline environments, sulfate reduction will be the dominant terminal-electron-accepting process owing to the higher affinity for these competitive growth substrates (e.g., Lovley et al., 1982). Nevertheless, methanogenesis remains an important process in marine and hypersaline environments, for example in sulfate-depleted zones in deeper sediments (Wilms et al., 2007), in areas with elevated hydrogen production (Hoehler et al., 2001; Buckley et al., 2008), and where carbon sources are available that cannot be used by sulfate reducers (Oremland et al., 1982; Winfrey and Ward, 1983). Such non-competitive substrates include methanol, dimethylsulfide and methylated amines, of which the latter derive from salinity-induced compatible solutes. A scheme for the production of methylamines is indicated in Fig. 1 . Dimethylsulfide derives primarily from hydrolysis of its precursor molecule dimethylsulfoniopropionate, which, like glycine betaine, is a compatible solute, and so is common in saline and hypersaline environments (Kiene et al., 1986; Kiene and Visscher, 1987). Therefore methylotrophic methanogens are nearly always the dominant methanogens in hypersaline conditions, including salt-saturated environments (~5.2 M NaCl), although there are exceptions (Oremland and King, 1989).

The relative importance of methylotrophic methanogenesis is well illustrated by an analysis of the upper salinity (in parentheses) at which pure methanogenic cultures have been shown to grow with various substrates (Oren, 1999): methylamines (27%), hydrogen and carbon dioxide (12%) and acetate (4%). These salinities should not be considered as the upper limit of activity in situ, but indicative of the relative importance of these substrates at different salinities. Competition with sulfate reducers, and the consequent reduced pool of halophilic hydrogenotrophic and aceticlastic methanogens on which natural selection can act, partly explain the observed differences in salinity tolerance. However, this trend is also governed by the relative energy gain from different methanogenic reactions per mole of substrate (methylotrophic >> hydrogenotrophic ≥ aceticlastic), especially since halophiles must expend a lot of energy to maintain an osmotically balanced and functional cytoplasm via the biosynthesis and/or uptake of organic compatible solutes, and/or uptake of potassium ions (Oren, 1999).
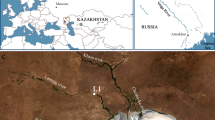
This review will focus more on those long-term, large-scale hypersaline anoxic environments, which have been studied in much greater depth, and their locations are shown in Fig. 2 . The emphasis will be on contemporary work which builds on studies outlined in some excellent reviews (e.g., Oremland and King, 1989; Ollivier et al., 1994; Oren, 1999; Marvin diPasquale et al., 1999; Oren, 2002). In order to understand the role of methanogenesis in carbon cycling in hypersaline environments evidence will be drawn from methane fluxes from field measurements, methane production rates (often from slurry experiments), cultivation, and investigation of uncultivated methanogenic communities. Methanogens are frequently studied without cultivation, owing to a generally good correspondence between phylogeny and phenotype that is less typical in other groups. Also, the mcrA gene, coding for methyl coenzyme-M reductase subunit-A, has proven valuable for investigating methanogens, and there is good correspondence between the phylogenies obtained with this functional gene and phylogenetic markers like 16S rRNA. This has resulted in widespread application of these markers to investigate methanogens in hypersaline (and many other) environments. Of course this is not absolute, and there is uncertainty over the phenotype of uncultivated organisms giving rise to 16S rRNA sequences that cluster within the Euryarchaeota but outside of known methanogens.

Locations of some of the main hypersaline environments where methanogensis has been studied.
2 Sedimentary Rocks
Waldron et al. (2007) exploited a natural salinity gradient from 8 mM to 3.5 M Cl− in the subsurface Antrim Shale, rich in methane derived from biodegraded hydrocarbons, to understand the salinity constraints on different types of methanogenesis. Methanogenesis was an important process in the shales, owing to a lack of competition with other terminal-electron-accepting processes, and it was evident that methanogens were capable of a high level of activity at their in-situ salinities, with the exception of the 3.5 M brine, in which there was no methane production. It is difficult to ascertain the percentage salinity of the two most saline brines (2.3 and 3.5 M Cl−) because the Na+ concentrations are 1.1 and 1.4 M respectively, implying the presence of other cations (perhaps K+, Mg2+, Ca2+) that were not measured. Based on most-probable-number enrichments, there is a clear change in methanogenic processes, with a mixture of hydrogenotrophic, aceticlastic and methylotrophic methanogenesis (primarily hydrogenotrophic) at low salinities and a predominance of methylotrophic methanogenesis at higher salinities (Waldron et al., 2007). This is supported by 16S rRNA clone libraries, in which all clone sequences resembled those from Methanocorpusculum spp. at 12.8 mM Cl−, while most resembled the methylotrophic Methanohalophilus spp. at 2.3 M Cl−. The second most abundant group from the high-salinity well was closely related to Methanoplanus petrolearius, previously shown to tolerate 0.86 M Cl−, and able to use a range of compounds including hydrogen and carbon dioxide (Ollivier et al., 1997), hinting that hydrogenotrophic methanogenesis may be possible at this very high salinity. Indeed Ollivier et al. (1998) isolated Methanocalculus haloterans from a hypersaline oil reservoir, a hydrogenotrophic methanogen capable of growth up to 12% NaCl (= 2 M Cl−). Perhaps, the long-term stability of the shale (>7000 years) and the absence of competing processes have enabled hydrogenotrophic methanogens to adapt to a higher salinity (Waldron et al., 2007).
3 Deep-Sea Hypersaline Anoxic Brine Lakes
There are numerous locations in the deep sea and the deep subsurface where dissolution of rock salt has resulted in hypersaline brine seeps and lakes, which are often associated with methane seeps. In the case of the deep-sea hypersaline brine lakes, the density gradient between the hypersaline brine and overlying seawater, coupled with weak currents at depth, restrict mixing, which results in the hypersaline brines becoming anoxic. Such pools and lakes have been discovered on the floor of the Gulf of Mexico, Red Sea and Mediterranean Sea. These hypersaline brines have commonly been shown to be a source of biogenic methane, often mixed with geogenic methane (Charlou et al., 2003; Joye et al., 2005). A small brine pool, called NR-1, in the Gulf of Mexico is even surrounded by dense beds of Bathymodiolus mussels which house chemoautotrophic bacteria presumably fed by sulfide or methane from the anoxic brine lake (MacDonald et al., 1990). In sediments fed by a sulfate-depleted hypersaline brine seep, Paull et al. (1985) found that the carbon in mussel tissue was isotopically light and hence presumably derived (via endosymbiotic methanotrophs) from brine-derived methane. Therefore, there is good evidence that food-webs are stimulated by reduced compounds derived from microbial activity in hypersaline brines analogous to hydrothermal vent communities (Martens et al., 1991).
In hypersaline brines on the floor of the Red Sea, such as the Kebrit and Shaban Basins, there are hints of the presence of methanogens from archaeal biomarkers (Michaelis et al., 1990) and euryarchaeal 16S rRNA gene amplicons (Eder et al., 2002), both of which could derive from non-methanogenic Archaea. More direct evidence has come from four of the eastern Mediterranean hypersaline basins where van der Wielen et al. (2005) detected methane production rates (μM CH4 d−1) of 85.8 (Urania), 16.9 (l’Atalante), 4.2 (Bannock) and 2.6 (Discovery). The most abundant uncultivated archaeal clones (termed MSBL-1) in most of these basins (van der Wielen et al., 2005) and in a hydrothermal mud vent beneath Urania hypersaline brine (Yakimov et al., 2007a), branched most closely to methanogens. The 16S rRNA signature from a very similar organism was also found in the anoxic hypolimnion of shallow hypersaline Solar Lake (Cytryn et al., 2000) and sediments of hypersaline Lake Chaka (Jiang et al., 2006). MSBL-1 Archaea probably represent a novel order, but despite their phylogenetic affiliation we currently have no firm evidence that they are methanogenic.
Yakimov et al. (2007b) found a change in the main archaeal community across the ~2-meter halocline from oxic Mediterranean seawater (depth 3498.5 m) to almost NaCl-saturated, anoxic l’Atalante brine; with a group related to ANME-1 (putative anaerobic methane-oxidizing group) co-existing with the aforementioned MSBL-1 group near the top of the interface, i.e., where salinity is not extremely high but where oxygen is highly depleted. MSBL-1 was detected also in the brine, but Methanohalophilus-related 16S rRNA sequences were most abundant in the deeper parts of the halocline and in the hypersaline brine. Messenger RNA coding for methyl coenzyme-M reductase from Methanohalophilus spp. was detected in the l’Atalante hypersaline brine (Hallsworth et al., 2007), and we isolated strains from l’Atalante sediment with ~99% 16S rRNA sequence similarity to Methanohalophilus mahii using sulfate-free medium and non-competitive growth substrates (Sass, Timmis, McGenity, unpublished). This suggests that methanogenesis is occurring throughout the hypersaline brine of l’Atalante basin, and is primarily mediated by Methanohalophilus spp. using methylamines. In Bannock basin, which is chemically similar to l’Atalante but located on the opposite side of the Mediterranean trench, extremely low methane production rates were observed throughout the halocline from deep Mediterranean seawater to the hypersaline brine, but this increased to 3.5 μM CH4 d−1 in the near-salt-saturated brine (Daffonchio et al., 2006). 16S rRNA sequences from Methanohalophilus spp. and relatives were not found in Bannock interface or brine, whereas MSBL-1 was detected in both and ANME-1 in the interface (van der Wielen et al., 2005; Daffonchio et al., 2006), raising interesting questions about the factors influencing the distribution of methanogens.
Three of the eastern Mediterranean brine lakes are dominated by NaCl, but the Discovery brine is unusual in that it derives from the dissolution of bischofite and so is an almost pure, 5 molar MgCl2 brine, with a water activity (aw) of 0.37, the lowest recorded in a marine environment and far below the current known limit of life (0.605) (see Hallsworth et al., 2007). Moreover, MgCl2 destabilizes biological macromolecules at high concentrations, and above a concentration of 2.3 M (in the absence of compensating solutes) appears to be inhibitory to life (Hallsworth et al., 2007). In support of this notion, is the detection of mRNA, a highly labile indicator of active microbes, from sulfate reducers and methanogens, only in the upper half of the halocline from seawater to Discovery brine (<2.23 M MgCl2; aw of <0.801), whereas methanogen mRNA was detected in the NaCl-rich l’Atalante brine at a lower water activity of 0.741 (Hallsworth et al., 2007). Although mRNA was not detected in the lower half of the interface, 16S rRNA from both groups was found, and can be attributed to the exceptional preservation properties of MgCl2 (Hallsworth et al., 2007). Marvin diPasquale et al. (1999) found that rates of methanogenesis were very low in the Dead Sea sediments (salinity of ~30%, aw of ~0.669 (Krumgalz and Millero, 1982) and highly enriched in divalent cations, especially MgCl2), with only methanol additions in some samples permitting low levels of methane production. This is in contrast to the lower salinity (<18%) and NaCl-dominated Solar Lake sediments were rapid methanogenesis was observed with a wider range of non-competitive substrates. The relative influence of salinity/low water activity and divalent cation concentration on biogeochemical processes like methanogenesis warrants further investigation.
4 Guerrero Negro and Other Hypersaline Microbial Mats
As mentioned earlier the Guerrero Negro mats are hypersaline yet one of the most biodiverse environments on Earth. Hoehler et al. (2001) observed that, along a vertical transect of a subtidal microbial mat, high levels of methane production coincided with unusually high hydrogen concentrations (a consequence of cyanobacterial activity in the upper few millimeters of the mat), and concluded that competition with sulfate-reducing bacteria and hence hydrogen limitation for methanogenesis was therefore negated. In support, Skyring et al. (1989) had shown that inhibition of sulfate reduction left hydrogen consumption unaffected. However, no analysis of the methanogenic community was made to support this observation, and the fact that the surface of the mat also harbored the highest density of cyanobacteria which may be leaking the compatible solute, glycine betaine, giving rise to of methylamines (Fig. 1 ), could also explain high levels of methanogenesis, as observed by King (1988) in a different hypersaline, cyanobacteria-dominated mat, and explained in detail by Oren, (1990). Indeed, Smith et al. (2008) investigating the Guerrero Negro mats, found that methylated amines were the primary route to methane formation and that methylotrophic members of the Methanosarcinales were the only methanogens detected by mcrA amplification and sequencing. In an artificial system in which sulfate was removed over the course of a year, hydrogenotrophic methanogens were observed, but were still less abundant than methylotrophic methanogens. As anticipated, methanogenesis was much higher in the mat sample with sulfate concentrations below 200 μM compared with the in-situ level of 70 mM (Smith et al., 2008). Back-to-back studies on Guerrero Negro mats with salinities of 7–7.5% (Jahnke et al., 2008; Orphan et al., 2008) confirmed that methylotrophic Methanosarcinales were the dominant methanogens, although in most parts of the mat, methanogens were only a small fraction of the Archaeal community. Also, various perturbations and analyses revealed a stratification of Methanosarcinales, with the following trend: Methanolobus spp. in the photic zone, Methanohalophilus spp. in the middle, and Methanococcoides in the unconsolidated sediment below the mat, extending the salinity at which Methanococcoides spp. have been found by 3.5% (Orphan et al., 2008). Importantly, both studies revealed lipids that could serve as possible biomarkers for methanogenesis in microbial mats, both modern and ancient (Jahnke et al., 2008; Orphan et al., 2008). In a more hypersaline series of microbial mats, Mouné et al. (2003) did not detect methanogens at a salinity of 25–32%, but Methanosarcinales represented 1% of the prokaryote community in a mat with a lower salinity of 15–20%, thus further emphasizing the abundance of Methanosarcinales in methanogenesis in hypersaline microbial mats.
In a microbial mat in a saltern pond in Eilat with a salinity of 21.5%, Sørensen et al. (2004) estimated methane flux from the mat of 1.6 × 10–5 nmol cm−2 s−1, one order of magnitude lower than in a pond of lower salinity (13.8%) studied previously. Slurry experiments with salinities from 5 to 32% revealed that the methane-production rates increased between 15 and 25% salinity, coinciding with a decrease in the sulfate-reduction rate. When molybdate was added to prevent sulfate reduction, methanogenesis also occurred at 5–15% salinity, proving that sulfate reducers were outcompeting methanogens at the lower salinities. Despite this, at the in-situ salinity the rate of methanogenesis was only about 5% of sulfate reduction, and thus contributes marginally to carbon mineralization (Sørensen et al., 2004).
Giani et al. (1989) investigated methanogenesis as a function of depth and salinity in Kervalet saltern ponds (Bretagne) by placing gas domes on top of microbial mats. Methanogenesis was enriched by the addition of methylated amines and methanol but not by acetate, formate, H2/CO2 and glycerol. As found by Sørensen et al. (2004) rates of methane production were greater at the highest salinities tested (20–33%), both in the field and in laboratory simulations, and were up to 100 times higher than those reported for typical marine environments, despite elevated sulfate concentrations. Methane production decreased at intermediate salinities (7–12%), possibly due to increased anaerobic methane oxidation (Giani et al., 1989), but was again high at the lowest salinity (4%).
5 Buried Salt Deposits
Many microbes, most notably Halobacteriaceae, have been isolated and their 16S rRNA signatures detected in salt crystals from buried evaporite deposits (see McGenity et al., 2000, 2008), but to date methanogens have evaded detection. Nevertheless, in salt mines naked flames are not permitted and there are records of methane-induced explosions; also gas-induced popping salt is a common threat to mine workers. The redox potentials of fluid inclusions in halite are generally negative, usually from −10 to −130 mV (summarized by Roedder, 1984), and methane, amongst other gases, is commonly detected (Roedder, 1984). Perhaps methanogens lack the capacity for long-term survival as seen in Halobacteriaceae (their close cousins who have adapted to aerobic conditions), in which case, the presence of methane can be explained by trapping of the gas during initial halite precipitation or more typically during recrystallization of buried halite (Pironon et al., 1995a,b). Alternatively, methane may be produced in young halite crystals by co-entombed methanogens which subsequently die, and over geological time their necromass serves as a source of carbon and energy for heterotrophic extremely halophilic microbes.
6 Other Environments
In addition to the MSBL-1 group discussed previously, there are several reports of methanogenesis in hypersaline environments in which the associated archaeal clone libraries contain Euryarchaeota that do not fall within clusters of known methanogens, e.g., Solar Lake hypolimnion (salinity >15%; Cytryn et al., 2000) and Quinghai Lake, China (salinity, 12.5%; Dong et al., 2006). Studying a soil-salinity gradient, Walsh et al. (2005) detected Methanosarcinales, including Methanohalophilus, as a minor component of the archaeal clones from the least saline (7%) soil, whereas at this salinity and higher there were numerous Euryarchaeota of unknown function.
Methanogenesis has been observed in many soda lakes with high pH. In slurries from Searles Lake (30% salinity) and Mono Lake (9% salinity) incubated at 25°C at pH 9.8, methane-production rates decreased from ~20–100 mmol cm−3 d−1 to ~0.5–1 mmol cm−3 d−1 with an increase in salinity from 2.5 to 35% (Kulp et al., 2007), implying that many of the methanogens in these lakes survive periods of elevated salinity but are better adapted to lower salinity, and are primed to become more active during wetter periods. It is notable therefore, that in relatively recent history (thousands of years), both lakes were much fresher than currently. In the Tuva and Southeastern Transbaikal soda lakes, methane is formed by methylotrophic and hydrogenotrophic pathways, respectively, with little contribution from aceticlastic methanogens (Zavarzin et al., 1996; Namsaraev et al., 1999).
Low temperatures are also not a barrier to methanogens. For example, in the Arctic Gypsum Hill spring (7.5% salinity; 5–7°C) 4% of the archaeal community was represented by close relatives of methylotrophic, psychrotolerant Methanococcoides spp., however, they were not detected in Color Peak spring (salinity 15.5%; Perreault et al., 2007). Instead, in the higher salinity spring there was an increase in putatively methanogenic Euryarchaeota to 16% of the archaeal community compared with 8% in the lower salinity spring (Perreault et al., 2007), again raising the important issue of understanding the true ecological roles of the Euryarchaeota giving rise to these 16S rRNA sequences.
7 Cycling of Methane
In many hypersaline environments methanogenesis is quantitatively far less important than sulfate reduction in the cycling of carbon. Nevertheless, if aerobic methane-oxidizing bacteria are inhibited to a much greater extent than methanogens, then overall methane flux to the atmosphere could be significant. Using a suite of incubation and inhibition techniques and measuring with gas chromatography, Conrad et al. (1995) found no evidence of methane oxidation in Solar Lake or in mats from the Eilat salterns, with salinities of 9 and 13%, respectively. By contrast, using 14C-labelled methane, methanotrophy was observed in Crimean microbial mats with salinities up to 33% (Sokolov and Trotsenko, 1995). There are also reports of pure methanotrophic cultures that can grow up to 15% NaCl (Heyer et al., 2005), and in-situ methanotrophy has been observed in the alkaline-saline Mono Lake (salinity ~9%) where both type-I and type-II methanotrophs were shown to be responsible, and rates of aerobic methane oxidation exceed those found in many marine environments (Carini et al., 2005; Lin et al., 2005).
There is also geochemical evidence of anaerobic methane oxidation in Mono Lake (Joye et al., 1999) and Big Soda Lake (Iversen et al., 1987). More recently, Lloyd et al. (2006) examined sediments overlying a brine-pool methane seep in the Gulf of Mexico, and showed that the archaeal community consisted predominantly of ANME-1b methane oxidizers (i.e., a phylogenetically distinct group of methanogens previously shown to oxidize methane by reverse methanogenesis in cooperation with sulfate-reducing bacteria), particularly concentrated around the methane-sulfate interface, where the salinity reaches ~13%. In lower salinity environments where anaerobic methane oxidation is occurring, other ANME groups are usually present. But in addition to the Gulf of Mexico (Lloyd et al., 2006), ANME-1-related 16S rRNA sequences were common in Mediterranean deep-sea hypersaline anoxic basins (Yakimov et al., 2007a, b), thus suggesting that the ANME-1 group is better adapted to hypersaline conditions than other anaerobic methane oxidizers.
As mentioned previously, methylamines are probably the main carbon and energy source for halophilic methanogens, and these largely derive from compatible solutes such as glycine betaine, which enter the environment upon cell death. Zhilina and Zavarzin isolated the first halophilic homoacetogen, Acetohalobium arabaticum, which converted glycine betaine to acetate and trimethylamine. When grown in co-culture with a Methanohalophilus strain, only acetate and methane accumulated in the medium, demonstrating the trophic link between an abundant compatible solute and methanogenesis in hypersaline environments (Zhilina and Zavarzin, 1990). Subsequently several other members of the Halanaerobiales able to produce methylamines from betaine have been described (see Oren, 2002).
From the above discussion it is clear that the relationship between methanogenesis and salinity is far from simple, and is dependent on the geological, historical and current physico-geochemical situation of the environment. Methanogenesis may decrease with increasing salinity owing to a concomitant increase in sulfate concentration, which in turn provides an abundant supply of terminal electron acceptor for sulfate-reducing bacteria, which out-compete methanogens for common resources such as acetate or hydrogen. Alternatively, methanogenesis may increase as salinity rises, because many microbes use glycine betaine as a compatible solute to cope with high salinity; and upon release from the cell, fermentative organisms can convert glycine betaine to methylamines (Fig. 1 ) which provide an energy-rich compound that can be used by many halophilic and halotolerant methanogens, but not by sulfate-reducing bacteria. Additionally, competition may be diminished by sulfate being used up, e.g., at depth or by precipitation of sulfate minerals. Nevertheless, growth at very high salinities is energetically demanding, and so often serves to inhibit methanogenesis.
8 Cultivated Halophilic Methanogens
Cultivated, taxonomically described halophilic methanogens are shown in Table 1 , and in addition there are several interesting halophilic strains whose names have not been validly published, especially in the genera Methanohalophilus and Methanohalobium (see Zhilina, 2001). Strain OCM 283, from an oil-reservoir brine, has a specific requirement for calcium and magnesium ions, reflecting the ionic composition of the oil-field brine from which it was isolated (Obraztsova et al., 1988). It was originally named “Methanococcoides euhalobius” (Obraztsova et al., 1988), but based on its wide NaCl tolerance (1–14%, optimum 6%), growth with methanol and methylamines and 16S rRNA sequence comparison, it was proposed to transfer it to the genus Methanohalophilus (Davidova et al., 1997), however this name has never been validated. A strain whose name was validly published as Halomethanococcus doii (Yu and Kawamura, 1987), but which probably belongs to the genus Methanohalophilus, has been lost (Boone, 2001). Nakatsugawa (1991) described strain NY-218 which grew optimally at 14–18% NaCl and grew well on methylamines and methanol, and moderately on acetate. Methanocalculus halotolerans is the most halotolerant strain known that uses hydrogen and carbon dioxide, as well as formate, as growth substrates (Ollivier et al., 1998). However, Pérez-Fillol et al. (1985) reported an extremely halophilic, methanogen that was both hydrogenotrophic and methylotrophic. The genus Methanolobus has several slightly halophilic strains, such as Methanolobus oregonensis DSMZ 5435, which grows up to 9% NaCl, which, in addition to using methanol and methylamines, can use dimethylsulfide and methanethiol (Liu et al., 1990). The combined properties of halophily, alkaliphily and the ability to convert methanethiol to methane by Methanolobus oregonensis and related species are actively being tested for desulfurization of petroleum and gas (van Leerdam et al., 2008).
Methanogens cope with elevated salinity by the accumulation of organic compatible solutes, often together with potassium ions which serve to counter the charge of anionic solutes. Additionally, enhanced expression of the gene coding for ClpB, a chaperone of the AAA+ superfamily, was identified in response to hyper- and hypo-osmotic stress (Shih and Lai, 2007). However, addition of glycine betaine reduced expression of clpB upon hyperosmotic shock, indicating that it has an important role in protein protection at high salinity. The common compatible solute, glycine betaine can be synthesized and accumulated by many methanogens, but they also use a variety of uncommon solutes, for example Methanohalophilus spp. use Nε-acetyl-β-lysine, β-glutamine, Lα-glutamate and α-glucosylglycerate (Lai et al., 1991; Roberts, 2005). Thus, on the one hand, not only does glycine betaine serve as a source of carbon and energy after fermentation to methylamines by other organisms, it is also an important osmoprotectant. It would be interesting to learn the extent to which halophilic methanogens ‘monitor’ the concentration of dissolved glycine betaine in the environment, and adjust its uptake and metabolism accordingly, on the one hand taking advantage of a pre-made compatible solute, while on the other retaining sufficient for future growth.
9 Research Needs
Few halophilic methanogens have been isolated recently, and publication dates in Table 1 indicate that none has been taxonomically described in the last decade. Also, the previous discussion highlights the presence of numerous Euryarchaeota that may be halophilic methanogens, but which remain to be cultivated. Bringing such organisms into culture should be a research priority, as was done recently for peat-bog methanogens, now represented by ‘Methanoregula boonei’ (Bräuer et al., 2006), and with a fully sequenced genome. Molecular biological tools are under-developed for methanogens, which, coupled with the paucity of genome sequences of halophilic methanogens, means that we are only just beginning to understand the generic and methanogen-specific features of adaptation to high salinity. Certainly, the potential interaction between compatible-solute and chaperone networks deserves further investigation (Shih and Lai, 2007). Cross-referencing clone libraries of mcrA and 16S rRNA sequences is valuable to detect trends such as determining which uncultivated methanogens are common to hypersaline environments. However, doing this manually is inefficient and, especially with the plethora of pyrosequencing data emerging, it is important to develop and apply software to study methanogen biogeography. For methanogens that evade cultivation, metagenomics, single-celled genomics, and stable-isotope probing, following 13C (e.g., from methylamines and other growth substrates) into lipids and nucleic acids, can provide a link between function and phylogeny.
There are numerous other areas that need further research if we are to truly understand carbon cycling past, present and future: it is important to understand how methanogens cope with desiccation (from rice paddies to salt marshes) because not only must methanogens tolerate low water activity, but also increased exposure to oxygen. What are the constraints of growth at high salinity – energetic (e.g., cost of making compatible solutes), competition, nutrient uptake etc., and how do these affect methane producers and consumers over space and time? Research into these areas may help us to learn whether and how aerobic Halobacteriaceae evolved from strictly anaerobic methanogens, and direct the search for putative missing links between the two archaeal groups.
References
Boone DR, Mathrani IM, Liu Y, Menaia JAGF, Mah RA, Boone JE (1993) Isolation and characterization of Methanohalophilus portucalensis sp. nov. and DNA reassociation study of the genus Methanohalophilus. Int J Syst Bacteriol 43: 430–437.
Boone DR (2001) Genus IV Methanohalophilus. In Bergey’s Manual of Systematic Bacteriology 2nd edn. vol. 1: The Archaea and the Deeply Branching and Phototrophic Bacteria. Garrity GM (ed.). New York: Springer, 281–283.
Bräuer SL, Cadillo-Quiroz H, Yashiro E, Yavitt JB, Zinder SH (2006) Isolation of a novel acidophilic methanogen from a peat bog. Nature 442: 192–194.
Buckley DH, Baumgartner LK, Visscher PT (2008) Vertical distribution of methane metabolism in microbial mats of the Great Sippewissett salt marsh. Environ Microbiol 10: 967–977.
Carini S, Bano N, LeCleir G, Joye SB (2005) Aerobic methane oxidation and methanotroph community composition during seasonal stratification in Mono Lake, California (USA). Environ Microbiol 7: 1127–1138.
Charlou JL, Donval JP, Zitter T, Roy N, Jean-Baptiste P, Foucher JP, Woodside J (2003) Evidence of methane venting and geochemistry of brines on mud volcanoes of the eastern Mediterranean Sea. Deep-Sea Res Pt I 50: 941–958.
Conrad R, Frenzel P, Cohen Y (1995) Methane emission from hypersaline microbial mats – lack of aerobic methane oxidation activity. FEMS Microbiol Ecol 16: 297–305.
Cytryn E, Minz D, Oremland RS, Cohen Y (2000) Distribution and diversity of archaea corresponding to the limnological cycle of a hypersaline stratified lake (Solar Lake, Sinai, Egypt). Appl Environ Microbiol 66: 3269–3276.
Daffonchio D, Borin S, Brusa T, Brusetti L, van der Wielen PWJJ, Bolhuis H, D’Auria G, Yakimov M, Giuliano L, Tamburini C, Marty D, McGenity TJ, Hallsworth JE, Sass AM, Timmis KN, Tselepides A, de Lange GJ, Huebner A, Thomson J, Varnavas SP, Gasparoni F, Gerber HW, Malinverno E, Corselli C, (2006) Stratified prokaryote network in the oxic-anoxic transition of a deep-sea halocline. Nature 440: 203–207.
Davidova IA, Harmsen HJM, Stams AJM, Belyaev SS, Zehnder AJB (1997) Taxonomic description of Methanococcoides euhalobius and its transfer to the Methanohalophilus genus. Antonie van Leeuwenhoek 71: 313–318.
Dong HL, Zhang GX, Jiang HC, Yu BS, Chapman LR, Lucas CR, Fields MW (2006) Microbial diversity in sediments of saline Qinghai Lake, China: Linking geochemical controls to microbial ecology. Microbial Ecol 51: 65–82.
Eder W, Schmidt M, Koch M, Garbe-Schonberg D, Huber R (2002) Prokaryotic phylogenetic diversity and corresponding geochemical data of the brine-seawater interface of the Shaban Deep, Red Sea. Environ Microbiol 4: 758–763.
Giani D, Jannsen D, Schostak V, Krumbein WE (1989) Methanogens in a saltern in the Bretagne (France). FEMS Microbiol Ecol 62: 143–150.
Grant WD, Tindall BJ (1986) The alkaline saline environment. In Microbes in Extreme Environments. RA Herbert and GA. Codd (eds.). London: Academic Press, 25–54.
Hallsworth JE, Yakimov MM, Golyshin PN, Gillion JLM, D'Auria G, de Lima Alves F, La Cono V, Genovese M, McKew BA, Hayes SL, Harris G, Giuliano L, Timmis KN, McGenity TJ (2007) Limits of life in MgCl2-containing environments: chaotropicity defines the window. Environ Microbiol 9: 801–813.
Heyer J, Berger U, Hardt M, Dunfield PF (2005) Methylohalobius crimeensis gen. nov., sp. nov., a moderately halophilic, methanotrophic bacterium isolated from hypersaline lakes of Crimea. Int J Syst Evol Microbiol 55: 1817–1826.
Hoehler TM, Bebout BM, Des Marais DJ (2001) The role of microbial mats in the production of reduced gases on the early Earth. Nature 412: 324–327.
Hovorka S (1987) Depositional environments of marine-dominated bedded halite, Permian San Andres Formation, Texas. Sedimentology 34: 1029–1054.
Hsü KJ, Ryan WBF, Cita MB (1973) Late Miocene desiccation of the Mediterranaean. Nature 242: 240–244.
Iversen N, Oremland RS, Klug MJ (1987) Big Soda Lake (Nevada). 3. Pelagic methanogenesis and anaerobic methane oxidation. Limnol Oceanogr 32: 804–814.
Jahnke LL, Orphan VJ, Embaye T, Turk KA, Kubo M, Summons RE, Des Marais DJ (2008) Lipid biomarker and phylogenetic analyses to reveal archaeal biodiversity and distribution in a hypersaline microbial mat and underlying sediment. Geobiology 6: 394–410.
Jiang HC, Dong HL, Zhang GX, Yu BS, Chapman LR, Fields MW (2006) Microbial diversity in water and sediment of Lake Chaka, an athalassohaline lake in northwestern China. Appl Environ Microbiol 72: 3832–3845.
Joye SB, Connell TL, Miller LG, Oremland RS, Jellison RS (1999) Oxidation of ammonia and methane in an alkaline, saline lake. Limnol Oceanogr 44: 178–188.
Joye SB, MacDonald IR, Montoya JP, Peccini M (2005) Geophysical and geochemical signatures of Gulf of Mexico seafloor brines. Biogeosciences 2: 295–309.
Kiene RP, Visscher PT (1987) Production and fate of methylated sulfur compounds from methionine and dimethylsulfoniopropionate in anoxic salt marsh sediments. Appl Environ Microbiol 53: 2426–2434.
Kiene RP, Oremland RS, Catena A, Miller LG, Capone DG (1986) Metabolism of reduced methylated sulfur compounds in anaerobic sediments and by a pure culture of an estuarine methanogen. Appl Environ Microbiol 52: 1037–1045.
King GM (1988) Methanogenesis from methylated amines in a hypersaline algal mat. Appl Environ Microbiol 54: 130–136.
Krumgalz BS, Millero FJ (1982) Physico-chemical study of the Dead Sea waters. I. Activity coefficients of major ions in Dead Sea water. Mar Chem 11: 209–222.
Kulp TR, Han S, Saltikov CW, Lanoil BD, Zargar K, Oremland RS (2007) Effects of imposed salinity gradients on dissimilatory arsenate reduction, sulfate reduction, and other microbial processes in sediments from two California soda lakes. Appl Environ Microbiol 73: 5130–5137.
Lai M-C, Sowers KR, Robertson DE, Roberts MF, Gunsalus RP (1991) Distribution of compatible solutes in the halophilic methanogenic archaebacteria. J Bacteriol 173: 5352–5358.
Ley RE, Harris JK, Wilcox J, Spear JR, Miller SR, Bebout BM, Maresca JA, Bryant DA, Sogin ML, Pace NR (2006) Unexpected diversity and complexity of the Guerrero Negro hypersaline microbial mat. Appl Environ Microbiol 72: 3685–3695.
Lin JL, Joye SB, Scholten JCM, Schafer H, McDonald IR, Murrell JC (2005) Analysis of methane monooxygenase genes in Mono Lake suggests that increased methane oxidation activity may correlate with a change in methanotroph community structure. Appl Environ Microbiol 71: 6458–6462.
Liu Y, Boone DR, Chay C (1990) Methanohalophilus oregonense sp. nov., a methylotrophic methanogen from an alkaline, saline aquifer. Int J Syst Bacteriol 40: 111–116.
Lloyd KG, Lapham L, Teske A (2006) An anerobic methane-oxidizing community of ANME-1b Archaea in hypersaline Gulf of Mexico sediments. Appl Environ Microbiol 72: 7218–7230.
Lovley DR, Dwyer DF, Klug MJ (1982) Kinetic analysis of competition between sulfate reducers and methanogens for hydrogen in sediments. Appl Environ Microbiol 45: 187–192.
Lozupone CA, Knight R (2007) Global patterns in bacterial diversity. Proc Natl Acad Sci USA 104: 11436–11440.
MacDonald IR, Reilly JF, Guinasso NL, Brooks JM, Carney RS, Bryant WA, Bright TJ (1990) Chemosynthetic mussels at a brine-filled pockmark in the northern Gulf of Mexico. Science 248: 1096–1099.
McGenity TJ, Gemmell RT, Grant WD, Stan-Lotter H (2000) Origins of halophilic microorganisms in ancient salt deposits. Environ Microbiol 2: 243–250.
McGenity TJ, Hallsworth JE, Timmis KN (2008) Connectivity between ‘ancient’ and ‘modern’ hypersaline environments, and the salinity limits of life. In: CIESM Workshop Monographs n°33: The Messinian Salinity Crisis from mega-deposits to microbiology – A consensus report. Almeria (Spain), 7–10 November 2007, 115–120.
Martens CS, Chanton JP, Paull CK (1991) Biogenic methane from abyssal brine seeps at the base of the Florida escarpment. Geology 19: 851–854.
Marvin diPasquale M, Oren A, Cohen Y, Oremland RS (1999) Radiotracer studies of bacterial methanogenesis in sediments from the Dead Sea and Solar Lake (Sinai). In Microbiology and Biogeochemistry of Hypersaline Environments. A Oren (ed.). Boca Raton FL: CRC Press, 149–160.
Mathrani IM, Boone DR, Mah RA, Fox GE, Lau PP (1988) Methanohalophilus zhilinae sp. nov., an alkaliphilic, halophilic, methylotrophic methanogen. Int J Syst Bacteriol 38: 139–142.
Michaelis WA, Jenisch A, Richnow HH (1990) Hydrothermal petroleum generation in Red Sea sediments from the Kebrit and Shaban Deeps. Appl Geochem 5: 103–114.
Mouné S, Caumette P, Matheron R, Willison JC (2003) Molecular sequence analysis of prokaryotic diversity in the anoxic sediments underlying cyanobacterial mats of two hypersaline ponds in Mediterranean salterns. FEMS Microbiol Ecol 44: 117–130.
Nakatsugawa N (1991) Novel methanogenic archaebacteria which grow in extreme environments. In Superbugs: Microorganisms in Extreme Environments. K Horikoshi and WD Grant (eds.). Berlin: Japan Scientific Societies Press, Tokyo, 212–220.
Namsaraev BB, Zhilina TN, Kulyrova AV, Gorlenko VM (1999) Bacterial methanogenesis in soda lakes of the southeastern Transbaikal region. Microbiology (Russia) 68: 586–591.
Obraztsova AJ, Shipin OV, Bezrukova LV, Beliaev SS (1988) Properties of the coccoid methylotrophic methanogen, Methanococcoides euhalobius sp. nov. Microbiology 56: 523–527.
Ollivier B, Caumette P, Garcia, JL, Mah RA (1994) Anaerobic bacteria from hypersaline environments. Microbiol Rev 58: 27–38.
Ollivier B, Cayol JL, Patel BKC, Magot M, Fardeau ML, Garcia JL (1997) Methanoplanus petrolearius sp. nov., a novel methanogenic bacterium from an oil-producing well. FEMS Microbiol Lett 147: 51–56.
Ollivier B, Fardeau ML, Cayol JL, Magot M, Patel BKC, Prensier G, Garcia JL (1998) Methanocalculus halotolerans gen. nov., sp. nov., isolated from an oil-producing well. Int J Syst Bacteriol 48: 821–828.
Oremland RS, Marsh LM, Polcin S (1982) Methane production and simultaneous sulphate reduction in anoxic, salt marsh sediments. Nature 296: 143–145.
Oremland RS, King GM (1989) Methanogenesis in hypersaline environments. In Microbial Mats: Physiological Ecology of Benthic Microbial Communities. Y Cohen and E Rosenberg (eds.). Washington, DC: American Society for Microbiology, 180–190.
Oren A (1990) Formation and breakdown of glycine betaine and trimethylamine in hypersaline environments. Antonie van Leeuwenhoek 58: 291–298.
Oren A (1999) Bioenergetic aspects of halophilism. Microbiol Mol Biol Rev 63: 334–348.
Oren A (2002) Halophilic Organisms and their Environments. Dordrecht: Kluwer Academic Publishers.
Oren A (2008) Microbial life at high salt concentrations: phylogenetic and metabolic diversity. Sal Syst 4: 2.
Orphan VJ, Jahnke LL, Embaye T, Turk KA, Pernthaler A, Summons RE, Des Marais DJ (2008) Characterization and spatial distribution of methanogens and methanogenic biosignatures in hypersaline microbial mats of Baja California. Geobiology 6: 376–393.
Paterek JR, Smith PH (1988) Methanohalophilus mahii gen. nov., sp. nov., a methylotrophic halophilic methanogen. Int J Syst Bacteriol 38: 122–123.
Paull CK, Jull AJT, Toolin LJ, Linick T (1985) Stable isotope evidence for chemosynthesis in an abyssal seep community. Nature 317: 709–711.
Pérez-Fillol M, Rodriquez-Valera F, Ferry JG (1985) Isolation of methanogenic bacteria able to grow in high salt concentration. Microbiología SEM 1: 29–33.
Perreault NN, Andersen DT, Pollard WH, Greer CW, Whyte LG (2007) Characterization of the prokaryotic diversity in cold saline perennial springs of the Canadian High Arctic. Appl Environ Microbiol 73: 1532–1543.
Pironon J, Pagel M, Lévêque MH, Mogé M (1995a) Organic inclusions in salt. Part 1. Solid and liquid organic matter, carbon dioxide and nitrogen species in fluid inclusions from the Bresse Basin (France). Org Geochem 23: 391–402.
Pironon J, Pagel M, Walgenwitz F, Barrès O (1995b) Organic inclusions in salt. Part 2. Oil, gas and ammonium in inclusions from the Gabon margin. Org Geochem 23: 739–750.
Roberts MF (2005) Organic compatible solutes of halotolerant and halophilic microorganisms. Sal Syst 1: 5
Roedder E (1984) The fluids in salt. Am Mineral 69: 413–439.
Skyring GW, Lynch RM, Smith GD (1989) Quantitative relationships between carbon, hydrogen, and sulphur metabolism in cyanobacterial mats. In Microbial Mats: Physiological Ecology of Benthic Microbial Communities. Y Cohen and E Rosenberg (eds.). Washington, DC: American Society for Microbiology, 170–179.
Shih C-J, Lai M-C (2007) Analysis of the AAA+ chaperone clpB gene and stress-response expression in the halophilic methanogenic archaeon Methanohalophilus portucalensis. Microbiology 153: 2572–2583.
Smith JM, Green SJ, Kelley CA, Prufert-Bebout L, Bebout BM (2008) Shifts in methanogen community structure and function associated with long-term manipulation of sulfate and salinity in a hypersaline microbial mat. Environ Microbiol 10: 386–394.
Sokolov AP, Trotsenko YA (1995) Methane consumption in (hyper)saline habitats of Crimea (Ukraine). FEMS Microbiol Ecol 18: 299–304.
Sørensen KB, Canfield DE, Oren A (2004) Salinity responses of benthic microbial communities in a solar saltern (Eilat, Israel). Appl Environ Microbiol 70: 1608–1616.
van der Wielen PWJJ, Bolhuis H, Borin S, Daffonchio D, Corselli C, Giuliano L, de Lange GJ, Huebner A, Varnavas SP, Thomson J, Tamburini C, Marty D, McGenity TJ, Timmis KN, (2005) The enigma of prokaryotic life in deep hypersaline anoxic basins. Science 307: 121–123.
van Leerdam RC, Bonilla-Salinas M, de Bok FAM, Bruning H, Lens PNL, Stams AJM, Janssen AJH (2008) Anaerobic methanethiol degradation and methanogenic community analysis in an alkaline (pH 10) biological process for Liquefied Petroleum Gas (LPG) desulfurization. Biotechnol Bioeng 101: 691–701.
Waldron PJ, Petsch ST, Martini AM, Nuslein K (2007) Salinity constraints on subsurface archaeal diversity and methanogenesis in sedimentary rock rich in organic matter. Appl Environ Microbiol 73: 4171–4179.
Walsh DA, Papke RT, Doolittle WF (2005) Archaeal diversity along a soil salinity gradient prone to disturbance. Environ Microbiol 7: 1655–1666.
Welsh DT (2000) Ecological significance of compatible solute accumulation by micro-organisms: from single cells to global climate. FEMS Microbiol Rev 24: 263–290.
Wilms R, Sass H, Köpke B, Cypionka H, Engelen B (2007) Methane and sulfate profiles within the subsurface of a tidal flat are reflected by the distribution of sulfate-reducing bacteria and methanogenic archaea. FEMS Microbiol Ecol 59: 611–621.
Winfrey MR and Ward DM (1983) Substrates for sulphate reduction and methane production in hypersaline sediments. Appl Environ Microbiol 45: 193–199.
Yakimov MM, Giuliano L, Cappello S, Denaro R, Golyshin PN (2007a) Microbial community of a hydrothermal mud vent underneath the deep-sea anoxic brine lake Urania (Eastern Mediterranean). Origins Life Evol B 37: 177–188.
Yakimov MM, La Cono V, Denaro R, D’Auria G, Decembrini F, Timmis KN, Golyshin PN, Giuliano L (2007b) Primary producing prokaryotic communities of brine, interface and seawater above the halocline of deep anoxic lake L’Atalante, Eastern Mediterranean Sea. ISME J 1: 743–755.
Yu IK, Kawamura F (1987) Halomethanococcus doii gen. nov., sp. nov.: an obligate halophilic methanogenic bacterium from solar salt ponds. J Gen Appl Microbiol 33: 303–310.
Zavarzin GA, Zhilina TN, Pikuta EV (1996) Secondary anaerobes in haloalkaliphilic communities in lakes of Tuva. Microbiology (Russia) 65: 480–486.
Zharkov MA (1981) History of Paleozoic Salt. Berlin: Springer.
Zhilina TN (1983) New obligate halophilic methane-producing bacterium. Microbiology 52: 290–297.
Zhilina TN (2001) Genus III Methanohalobium. In Bergey’s Manual of Systematic Bacteriology 2nd edn. vol. 1: The Archaea and the Deeply Branching and Phototrophic Bacteria. GM Garrity (ed.). New York: Springer, 279–281.
Zhilina TN, Zavarzin GA (1987) Methanohalobium evestigatum gen. nov., sp. nov., extremely halophilic methane-producing archaebacteria. Dokl Akad Nauk SSSR 293: 464–468.
Zhilina TN, Zavarzin GA (1990) Extremely halophilic, methylotrophic, anaerobic bacteria. FEMS Microbiol Rev 87: 315–322.
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2010 Springer-Verlag Berlin Heidelberg
About this entry
Cite this entry
McGenity, T.J. (2010). Methanogens and Methanogenesis in Hypersaline Environments. In: Timmis, K.N. (eds) Handbook of Hydrocarbon and Lipid Microbiology. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-540-77587-4_53
Download citation
DOI: https://doi.org/10.1007/978-3-540-77587-4_53
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-540-77584-3
Online ISBN: 978-3-540-77587-4
eBook Packages: Biomedical and Life SciencesReference Module Biomedical and Life Sciences