
Abstract
The genus Castanea, chestnuts and chinkapins, belongs to the family Fagaceae, which includes other important timber producing genera such as Quercus and Fagus. The genus Castanea is divided into three geographically delimited sections with at least seven consistently recognized interfertile species: 4 species in Asia (C. mollissima, C. henryi, C. seguinii, and C. crenata), two or more species in North America (C. dentata, C. ozarkensis, and C. pumila) and one in Europe and Turkey (C. sativa). The two most important diseases of chestnut are ink disease (Phytophthora) and chestnut blight (Cryphonectria). Resistance to these is the major objective for rootstock breeding in Europe and scion breeding in North America. In both cases, the source of resistance was Asian species. European breeding programs developed resistant hybrid rootstocks, which are propagated by stooling, cuttings, or in vitro culture. A major pest of chestnut is the gall wasp Dryocosmus kuriphilus whose control is based on the spread of parasitoids but also on the selection of resistant cultivars. For nut production, the most important breeding objectives include the following: good horticultural traits, product quality, suitability to storage and processing, and ease of peeling. For timber, important characters include wood quality, rapid growth, and nonchecking of wood (ring-shake). Molecular maps have been developed, which has expanded the genetic knowledge of the chestnut. An efficient genetic transformation protocol for C. sativa through the coculture of somatic embryos with different strains of Agrobacterium tumefaciens has been described.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
In the world there are about 349,000 ha of orchards which produce 1,140,332 mt of chestnuts (mean value for 2000–2007). Chestnut production in Asia is almost 8 times that of Europe, with China being the dominant producer with an average production for 2000–2007 of 803,213 mt. China is reported to have about 130,000 ha in chestnut orchards, but Liu and Zhou (1999) estimated that the figure should be 670,000 ha, five times the FAO figure. Japan, Turkey, and Korea produce about 25,000, 50,000, and 78,000 mt respectively (FAO 2009, faostat.fao.org).
A survey among European countries estimates that there are 2.22 million ha of chestnut dominated forest (Conedera et al. 2004a). The main chestnut production is in Italy (24,000 ha) and Portugal (30,000 ha), with about 51,000 and 29,000 mt, respectively (FAO 2009, faostat.fao.org). Although the FAO accounts only 7,000 ha in Spain, Spanish statistics estimate 45,000 ha and 60,000 mt in 2006 (http://www.mma.es).
Chestnuts are multipurpose trees valued for nuts, timber, tannins, and landscape. They were historically distributed only throughout the northern hemisphere, but have more recently been introduced into Chile, Argentina, Australia, and New Zealand.
1.1 Taxonomy and Distribution
The genus Castanea (2n = 24), chestnuts and chinkapins, belongs to the family Fagaceae, which includes other important timber producing genera such as Quercus (oaks) and Fagus (beech) (Castroviejo et al. 1990). It is supported as a monophyletic clade most closely related to the genus Castanopsis (Manos et al. 2001).
The genus Castanea has been divided into three sections with at least seven consistently recognized species (Camus 1929; Johnson 1988). The section Eucastanon consists of five species characterized by three nuts per cupule; C. crenata Sieb. & Zucc. from Japan, C. mollissima Blume and C. seguinii Dode from China, C. sativa Miller from Europe, and C. dentata Borkhausen from North America. The section Balanocastanon is found exclusively in southeastern North America and characterized by a single nut per cupule. Although four to six imprecisely defined species are generally cited for this section (Graves 1961; Elias 1971; Jaynes 1972; Little 1979), more recently it has been proposed that it should be reduced to a single species, C. pumila Miller, with two varieties: var pumila and var. ozarkensis (Hardin and Johnson 1985; Johnson 1988). The section Hypocastanon consisting of C. henryi Rehder and Wilson from China is also characterized by a single nut per cupule.
UPGMA analysis of isozyme-based genetic distance estimates (Dane et al. 2003) and phylogenetic analysis based on cpDNA sequence data (Lang et al. 2006) suggest that Castanea species are geographically structured. This is inconsistent with the current phylogeny based on cupule characteristics. The section Eucastanon appears to be paraphyletic with the differentiation among species being best explained by their current geographical ranges. C. crenata appears to be the most basal taxa and sister to the remainder of the genus. The three Chinese species [C. mollissima and C. seguinii (Eucastanon) and C. henryi (Hypocastanon)] are supported as a single monophyletic clade and sister to a group containing the North American and European species. There appears to be weak but consistent support for a sister-group relationship between the North American and European species.
With cpDNA data, the chestnut appeared to expand westward from the extant Castanea species originating in eastern Asia, followed by intercontinental dispersion and divergence between the Chinese and European/North American species during the middle Eocene, followed by subsequent divergence between the European and North American species during the late Eocene (Lang et al. 2007). Morphological evolution of one nut per bur in the genus may have occurred independently on two continents.
1.2 Where Grown
There are three main areas where native chestnuts are found (Table 19.1, Fig. 19.1).

Main areas where chestnut (Castanea spp.) is grown
-
1.
In Asia, mainly in China, where C. mollissima Blume, C. henryi (Skan) Rehd. & E.H. Wils., and C. seguinii Dode are found in wild and cultivated stands. In the Korean Peninsula, Japan, and the temperate region of east Asia, the Japanese chestnut (Castanea crenata Sieb. & Zucc.) is naturally distributed and cultivated.
-
2.
In Europe and Turkey where C. sativa is predominant.
-
3.
In North America where C. dentata (Marsh.) Borkh. and C. pumila (L.) Mill. var. pumila were once widespread throughout the Appalachian Mountain Range, and where C. ozarkensis Ashe and C. alnifolia Nutt occupied small niches on the Ozark Plateau and in northern Florida.
The main species cultivated for fruit are C. mollissima, C. sativa, and C. crenata due to their large nut size (Table 19.2). Marrone types (C. sativa) are considered the most valuable for nut production. C. sativa and C. dentata are the most vigorous species, and they are also used for timber production. Interspecific hybrids which have emerged from disease resistance work are used for nut, timber and as rootstock.
1.3 Limits to Adaptation
In Europe, C. sativa is commonly found between 400 and 1,000 m above sea level depending on the latitude. The lowest elevations are recommended for the highest latitudes and vice versa (Bounous 2002). The early leafing of hybrids (mid March) restricts their use due to spring frosts to altitudes lower than 500 m.
The minimum rainfall for chestnut is 800 mm. Plants from this species are moderately thermophilic and well adapted to ecosystems with a year mean temperature ranging between 8 and 15°C and monthly mean temperatures during 6 months over 10°C.
Characterized as a mesophilic species, chestnut tree growth actually shows some limitations to high temperatures. European chestnut trees do not thrive in soil rich in active calcium, basic pH or with poor drainage. These are commonly grown in poor sandy to loamy soil on slopes but also in volcanic islands (Sicily, Canary, Madeira, and Azores). Deep soil and a deep root system are important to help trees maintain their water potential during the dry hot summer months (June to September) (Martins et al. 2005).
Chestnut is a dim-light species with better adaptation to shade and cold north-facing slopes, rather than south-facing ones (Gomes-Laranjo et al. 2007). The latter have higher mean temperatures, earlier leafing and flowering, and consequently a greater frost risk relative to north-facing ones.
2 Origin and Domestication of Scion Cultivars
2.1 Origin of the Cultivars
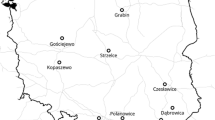
In Europe, the most probable natural range of the native chestnut species C. sativa, is delimited by six macroregions (Fig. 19.2): the Transcaucasian region, north-western Anatolia, the hinterland of the Tyrrhenian coast from Liguria to Lazio along the Apennine range, the region around Lago di Monticchio (Monte Vulture) in southern Italy, the Cantabrian coast on the Iberian Peninsula, and probably also the Greek Peninsula (Peloponnese and Thessaly), and north-eastern Italy (Colli Euganei, Monti Berici, Emilia-Romagna) (Krebs et al. 2004).

Distribution of Castanea sativa in Europe
The first evidence of active chestnut cultivation dates back to the third millennium before Christ in the eastern part of European range (Anatolian Peninsula, Northeastern Greece, and Southeastern Bulgaria). From there, Greeks first and then the Romans diffused the chestnut to the west (Pitte 1985, 1986; Conedera et al. 2004b) such that in the Middle Ages the cultivation of chestnut for timber production and as a staple food was a widespread component of the traditional farming system in most.
Mediterranean countries and southern parts of Central Europe (Conedera and Krebs 2008).
A large-scale molecular study based on simple sequence repeat (SSR) loci of the diversification process in chestnut cultivars from Portugal and Spain, from the northern Iberian Peninsula to the Canary Islands and the Azores, showed geographical and genetic structure in ten main cultivar groups (Pereira-Lorenzo et al. 2011). Cultivar origin and the diversification process was a combination of clonal propagation of selected seedlings, hybridization, and mutations. Mean value of clonality owing to grafting was 33%, mutations accounted for 6%, with hybridization being the main diversification process that can explain the great diversity found. Seedlings and graft sticks were transported in the colonization process, sometimes more than 3,000 km if we consider the Azores and the Canary Islands.
Although the phylogenetic map of the chestnut in Europe is not fully understood yet (Fineschi et al. 2000), the greater genetic similarity of chestnuts from the western Anatolia Peninsula to Italian and French populations than to the chestnut groves of eastern Anatolia facing the Black Sea (Villani et al. 1999) suggests that the flow of chestnut-related elements between Ancient Greece and the Italian world was not only cultural (Conedera et al. 2004b). It is probable that Greek colonists introduced chestnut cultivation in the Italian Peninsula from the main chestnut zones of Ancient Greece as they did for grapes (Dion 1977).
2.2 Brief History of Breeding
First hybridizations were made in 1884 in the USA (Van Fleet 1920), in 1926 in Spain (Gallastegui 1926) and in 1929 in Japan (Yamamoto et al. personal communication) (Table 19.3). Hybrid clones have been released in Japan, the USA, and Europe (France, Spain, and Portugal; Pereira-Lorenzo et al. 2010) and some of them are commercialized for nut, timber, and rootstocks.
2.3 Varietal Groups
The International Society for Horticultural Science maintains a registry of chestnut cultivars with names and their characteristics (Connecticut Agricultural Experiment Station, The USA, http://www.ct.gov/caes).
Most of the best cultivars recommended in France for new plantations are C. sativa cultivars, which produce marron nuts between 12 and 18 g (Table 19.4). French researchers have focused in the breeding of interspecific hybrids resistant to ink diseases suitable for fresh market use (Bergougnoux et al. 1978; Breisch 1995).
The most important cultivars in Spain are ‘Parede’ and ‘Longal’. They have been propagated profusely during the last 300 years (Pereira-Lorenzo et al. 2001a, 2006a). In Northern Spain, the most popular cultivars are ‘Amarelante’, ‘Negral’, ‘Famosa’, ‘Longal’, ‘Ventura’, ‘Garrida’, ‘Loura’, and ‘Luguesa’ (Table 19.5). In Extremadura, Central Spain, ‘Injerta’ and ‘Verata’ are cultivated, and in Southern Spain, ‘Planta Alajar’, ‘Temprana’, and ‘Pilonga’ are the best. In the Canary Islands, the most widespread cultivars are ‘Mulata’ in Tenerife and ‘Jabuda’ in La Palma (Pereira-Lorenzo et al. 2001b, c). In Spain, hybrids are considered an alternative in Atlantic areas where they show very good adaptability (Pereira-Lorenzo and Fernandez-Lopez 2001) with sufficiently large and monoembryonic nuts which are harvested before the 20th September.
In Portugal, ‘Longal’, one of the most ancestral varieties, is widely spread over all the chestnut regions (Trás-os-Montes, located in the northeast) and has been promoted as the best cultivar for industry. ‘Judía’ and ‘Martaínha’, due to their larger nut size, are usually preferred for the fresh market. ‘Judía frequently presents polyembryony (Table 19.6). Some cultivars such as ‘Longal’, ‘Amarelal’, and ‘Verdeal’ are found in North Spain and Portugal.
In California, the principal cultivar is ‘Colossal’ (C. sativa × C. crenata) grown with ‘Silverleaf’ (C. sativa), ‘Nevada’ (C. sativa × C. crenata), ‘Eurobella’ (C. sativa × C. crenata), or ‘Colossal’ seedlings as pollinizers (Vossen 2000).
In Italy, where the environmental conditions are favorable, the best cultivars are the marron type (‘Chiusa Pesio’, ‘Luserna’, ‘Val Susa’, ‘Castel del Rio’, ‘Marradi’, ‘Fiorentino’ from Italy) because of their large size which is preferred for the fresh and candy market (marrons glacés) (Table 19.7). Early maturing cultivars suitable for the premium market are ‘Tempurive’, ‘Castagne della Madonna’, and ‘Precoce di Roccamonfina’, while ‘Garrone Rosso’, ‘Garrone Nero’, ‘Gioviasca’, ‘Bionda di Mercogliano’, and ‘Montemarano’ produce large chestnuts for fresh market and candying. Many cultivars with small but very sweet and easy to peel nuts suitable for drying and flour production have been selected through the centuries in Italy: ‘Frattona’, ‘Gabbiana’, ‘Siria’, ‘Pastinese’, ‘Carpinese’ (Bounous 2002). In the 1980s, the University of Torino, Department of Arboriculture, released two Italian Euro-Japanese hybrids: ‘Primato’ and ‘Lusenta’.
In southern Switzerland, only two cultivars are ubiquitous and widespread throughout the chestnut area: the ‘Lüina’, a tree producing small-sized very sweet fruits for drying, and the ‘Verdesa’, a late ripening cultivar that keeps the fruit inside the bur allowing the chestnuts to be conserved over months for fresh consumption. It is not surprising, given the former staple food function of the chestnut in this mountain region in southern Switzerland, that the most suitable cultivars have excellent fresh and dry storage traits (Conedera et al. 1993).
Liu and Zhou (1999) identified the best six Chinese cultivars (C. mollissima) out of 28 examined. These include ‘Chu shu hong’, ‘Jiu jia zhong’, ‘Duan zha’, ‘Qin zha’, ‘Jiao zha’, and ‘Jian ding you li’. All of them produce nuts over 10 g (Jiao zha over 20 g), are easy to peel, and have excellent kernel quality for both cooking and roasting.
In Japan, most commercial cultivars belong to Japanese chestnut (Castanea crenata Sieb. & Zucc.), with only a few cultivars, such as ‘Riheiguri’, being hybrids between Japanese and Chinese chestnuts. In 2004, the most widely cultivated chestnut cultivars in Japan were ‘Tsukuba’ (23.3%), ‘Tanzawa’ (12.0%), ‘Ginyose’ (11.4%), ‘Ishizuchi’ (4.1%), ‘Riheiguri’ (4.0%), ‘Kunimi’ (3.7%), and ‘Ganne’ (2.9%) (Table 19.8). ‘Tsukuba’ (‘Ganne’ × ‘Hayadama’), the leading cultivar grown in Japan, was released as ‘Norin No.3’ in 1959 by NFIFTS (National Institute of Fruit Tree Science, formerly Fruit Tree Research Station). Although damage by gall wasps is serious, it is still a leading cultivar because of its high productivity. ‘Tanzawa’ (‘Otomune’ × ‘Taisyowase’) was named and released as ‘Norin No.1’ in 1959 by NFIFTS. ‘Ginyose’ is a rather old cultivar, which is thought to be derived from a chance seedling found at Toyono, Osaka around 1750, and is resistant to chestnut gall wasp. ‘Kunimi’ (‘Tanzawa’ × ‘Taisyowase’) was named and released as ‘Norin No.5’ in 1981 by NFIFTS. It is resistant to gall wasp, and suffers less damage from yellow peach moth than other cultivars. ‘Ishizuchi’ (‘Ganne’ × ‘Kasaharawase’) was named and released as ‘Norin No.4’ in 1968 by NFIFTS, and is resistant to chestnut gall wasp. ‘Riheiguri’ was selected and registered by K. Tsuchida in Gifu Prefecture in 1950. ‘Ganne’, a chance seedling, is resistant to chestnut gall wasp.
2.4 Rootstocks
Traditionally, growers have used as rootstocks, seedlings growing under grafted trees as well as seedlings from selected mother trees that gave seed with good emergence rate, growth and drought tolerance (Soylu and Serdar 2000) to establish the new orchards. These rootstocks have excellent graft compatibility and are suitable for poor soils; however, rootstocks from C. sativa are susceptible to ink disease.
At the beginning of the twentieth century, researchers from France, Spain, Italy and Portugal introduced seeds of C. crenata from Japan and C. mollissima from China into Europe. These species were resistant to ink disease, but their nuts were not appreciated by growers because of poor peeling, nor were they good for timber due to poor vigor. Later, they were tried as rootstocks, but incompatibility was common. Interspecific hybrids were made in Portugal, Spain and France and some clones were selected as rootstocks, combining resistance to ink disease, easy propagation, and good compatibility with good growth and production in poor soils where chestnut is normally cultivated.
Currently, only France and Spain commercialize their resistant hybrid rootstocks propagated by stooling, cuttings, or in vitro culture. Five French (Breisch 1995) and four Spanish hybrid clones (Pereira-Lorenzo and Fernández-López 1997; Pereira-Lorenzo et al. 1999) are recommended. Resistance to ink disease varies from low to very high for French rootstocks (Breisch 1995), while the Spanish hybrids vary from medium to very resistant (Fernández-López et al. 2002) (Table 19.9) (Breisch 1995; Pereira-Lorenzo and Fernández-López 1997). Their compatibility is excellent. Among the Spanish rootstocks, ‘CHR-151’ (‘HS’) which is easily propagated via in vitro culture, has been broadly used (Miranda-Fontaíña and Fernández-López 1992). No data are available about the importance of hybrid rootstocks in new plantations.
3 Genetic Resources
Genetic resources of chestnut have been collected by different institutions throughout the world (Bounous 2002). Cultivars, seedlings and interspecific hybrids are preserved in different institutions in Austria, China, France, Hungary, Korea, Italy, Portugal, Slovak Republic, Slovenia, Spain, Switzerland, Turkey, and the UK (Bounous 2002; Tables 19.4–19.8).
Species-level genetic diversity within the genus. Based on isoenzymes, C. dentata appears to be the least variable of the North American species H e ∼ 0.18 (Huang et al. 1994b; Huang et al. 1998). Levels of genetic diversity are higher for C. pumila H e ∼ 0.30 (Fu and Dane 2003) as well as for C. pumila var. ozarkensis H e ∼ 0.27 (Dane et al. 1999). Although estimates of gene diversity appear to be lower for C. dentata, this level of diversity is similar to that found in other woody plant species (Hamrick and Godt 1989). C. dentata also appears to harbor less variation (11%) among populations than does C. pumila (30.4%) or C. ozarkensis (14.7%). Huang et al. (1998) showed evidence for possible geographic structure in C. dentata, with southern populations showing higher levels of genetic diversity possibly related to their glacial refugium; they studied RAPD and SSR variation in ∼1,000 trees from 18 sample sites. Subsequent research using RAPDs and SSRs found that, although genetic differentiation among C. dentata populations has taken place, no disjunct regional pattern is apparent. C. dentata still exists as a highly variable species, even at the extremes of its natural range. Genetic variability in C. dentata follows a pattern consistent with the hypothesis of a single metapopulation where genetic drift will continue to play a major evolutionary role (Kubisiak and Roberds 2006). When compared to Asian and European Castanea (Huang et al. 1994a; Lang et al. 2007; Villani et al. 1991a, b), levels of genetic diversity based on isoenzymes in C. dentata appear to be similar to those reported for C. seguinii (H e ∼ 0.20), and levels reported for the two cultivars of C. pumila appear similar to those reported for all other Castanea: C. mollissima (H e ∼ 0.31), C. henryi (H e ∼ 0.26), and C. sativa (H e ∼ 0.24).
Levels of within and among natural diversity for specific species. In Europe, genetic variability in natural chestnut populations established that genes flow from East (Turkey) to the West (Italy) (Pigliucci et al. 1990a, b; Villani et al. 1991a, b, 1993; Aravanopoulos et al. 2002). Two main origins of variability in European cultivated chestnut were found in the Iberian Peninsula by SSRs, one in the North and a second in the Center (Pereira-Lorenzo et al. 2010).
The genetic diversity of wild chestnut (C. crenata) populations in northern Japan showed a high level of heterozygosity in wild populations (Tanaka et al. 2005). The H O and H E values in the chestnut (C. crenata) populations (H O: 0.727 and H E: 0.780) were similar to other Fagaceae such as Fagus sylvatica (0.727 and 0.753) (Pastorelli et al. 2003), Fagus orientalis (0.697 and 0.740) (Pastorelli et al. 2003), and Quercus rubra (0.679 and 0.737) (Aldrich et al. 2002).
Cultivar within species genetic diversity. Important efforts are being made in studying chestnut variability using morphological characteristics based on Breviglieri’s (1955) ‘Scheda Castanografica’, after the UPOV chestnut guideline (1988) and, more recently, applied to the Spanish cultivars (Pereira-Lorenzo et al. 1996a, 2006a) and different chestnut species (Oraguzie et al. 1998).
The first molecular markers based studies in chestnut used isoenzymes. Sawano et al. (1984) studied 16 clones (10 Japanese, 3 Chinese, and 2 hybrids). Wen and Norton (1992) studied isoenzymes and identified 22 Chinese cultivars. Other genetic analyses with isoenzymes were performed by Bonnefoi (1984), Malvotti and Fineschi (1987), Fineschi et al. (1990a, b), Huang et al. (1994a), and Pereira et al. (1999). RAPD markers were studied by Valdiviesso (1999), RAPD and ISSR by Goulao et al. (2001), and nuSSRs by Costa et al. (2008). The Portuguese cultivars showed great genetic variability, with multiple genotypes obtained per cultivar indicating their polyclonal origin (Costa et al. 2008). The different genotypes obtained for the main Portuguese cultivars have derived mainly from cross-pollination between them, but also from mutations. Four regions of Protected Denomination of Origin (DOP) were created to preserve the Portuguese cultivars: Castanha da Terra Fria, Castanha da Padrela, Castanha dos Soutos da Lapa and Castanha do Marvão. Molecular analysis showed the greatest variability (the largest number of genotypes) was located in the Northern regions (Castanha da Padrela) as compared to the southern region (Castanha do Marvão), which can be explained by the common practice of exchanging plant material for grafting in the Northern regions (Costa et al. 2008).
However, Fineschi et al. (1994) showed a relatively high degree of homogeneity both among individuals of the same cultivar and among cultivars of the same area in Italy, but a high genetic distance between geographic areas. Pereira-Lorenzo et al. (1996b, 2006a) studied the variability of the Spanish chestnut cultivars by isoenzymes and demonstrated that, in main cultivars. a main clone was predominant in orchards (over 60% of the samples), but intracultivar variability was important, surely due to the use of seedlings of those main cultivars by the growers. The H O and H E values obtained with isoenzymes in the Spanish chestnut cultivars were, on average, 0.398 and 0.333 respectively (Pereira-Lorenzo et al. 2006a). The excess of heterozygotes found in Galician chestnuts that were at least 300 years old was similar to that found in natural populations of C. dentata that were over 70 years old in Virginia, USA (Stilwell et al. 2003). The excess heterozygosity in these two populations may be due to the selection of heterozygous breeding material and by an absence of new recruits from other populations as suggested by Stilwell et al. (2003). Subsequent asexual propagation through grafting would maintain this situation. Subpopulations isolated in southern Spain had lower variation. Microsatellites confirmed the variability found with isoenzymes in Spanish cultivars (Pereira-Lorenzo et al. 2006a, 2010; Ramos-Cabrer et al. 2006) and heterozygosity was significantly higher as it occurs in other species.
In Italy, 33 microsatellite (SSR) loci were isolated in chestnut (Marinoni et al. 2003) and several oak loci (Steinkellner et al. 1997; Kampfer et al. 1998) were found to be polymorphic in Castanea sativa (Boccacci et al. 2004). Microsatellites are preferred for the DNA genotyping of cultivars aimed at identification, and were used in many studies of characterization of Castanea sativa Mill. cultivated germplasm, leading to the identification of over 70 cultivars.
Twenty cultivars from the North West Italian germplasm were characterized at 14 polymorphic loci (Marinoni et al. 2003). The total number of alleles was 90, and ranged from 4 to 10 per locus, with an average of 6.4. The mean expected heterozygosity was 0.72 (range: 0.65–0.83). The average observed heterozygosity (Ho) was 0.793 (range: 0.35–0.95). Further work was carried out within the EU project MANCHEST, and 121 North Italian accessions (Piemonte Region), including 39 Marrone individuals, were characterized (Botta et al. 2006) using a selected set of ten loci that included additional SSR markers isolated within the project by Buck et al. (2003). The loci (QrZAG96, QpZAG110, QpZAG119, CsCAT1, CsCAT3, CsCAT4, CsCAT6, CsCAT16, CsCAT17, and EMC15) were chosen on the basis of their position and distribution in the genome. Fifty-two genotypes were identified by the markers and were described by chemical and morphological traits.
Thirty Japanese chestnut accessions (Castanea crenata Sieb. & Zucc.) were evaluated by SSR markers, including 12 cultivars and 6 wild landraces originated in Japan, and 6 cultivars and 6 wild landraces originated in the Korean Peninsula (Yamamoto et al. 2003). The 14 polymorphic SSR loci produced 2–16 alleles per locus. The average values of heterozygosity and polymorphic information content among the 14 loci were 0.50 (0.10–0.93) and 0.54 (0.10–0.89), respectively. No differences on allele composition were observed between cultivated and wild landtraces as well as between Japanese and Korean origins. The results could indicate that the Japanese chestnuts originating from Japan and the Korean Peninsula showed similar genetic background, and that cultivated chestnuts might have been selected from wild chestnuts.
SSRs were also used to identify cultivars from Italy (Martín et al. 2010), Portugal and Spain (Pereira-Lorenzo et al. 2011). In the Iberian Peninsula, ten main groups of cultivars have been found related with the two main origins of variability, the Northern and the Central Iberian Peninsula. This study demonstrated that cultivar origin and the diversification process was a combination of clonal propagation of selected seedlings, hybridization, and mutations, which allowed high levels of diversity to be maintained with respect to selected clones for fruit production.
4 Major Breeding Achievements
4.1 General Achievements
Diseases. Two main diseases, ink disease (Phytophthora spp.) and blight (Cryphonectria parasitica), threaten chestnut production. The European chestnut species C. sativa present less tolerance to main pest and diseases. Different genetic markers and different analytical approaches have shown a very significant amount of genetic variation for the whole range of species, pointing out the uniqueness of the Greek gene pool (Aravanopoulos et al. 2005).
Chestnut breeding in Europe began with the production of hybrids resistant to ink disease (Phytophthora spp.) to substitute the indigenous species. Initially, seedlings from Asian species C. crenata and C. mollissima were introduced between 1917 and 1940 (Elorrieta 1949) as a way to control ink disease, which was threatening the European chestnut orchards. Resistance on the Asian species was confirmed later, but these hybrids were in many traits inferior to the European species C. sativa; i.e. less vigor, lower quality of the nuts, bad affinity with the local cultivars, sensitivity to early spring frost and summer drought, and difficulty adapting to climatic characteristics of some areas in Europe (Elorrieta 1949; Pereira-Lorenzo and Fernandez-Lopez 2001). In 1989, a new program began to identify some hybrid clones that were interesting for timber, nut production or rootstocks (Pereira-Lorenzo and Fernández-López 1997, 2001).
In France, Asian species were introduced in 1925 and they showed high tolerance to ink disease but poor adaptation to soil and weather conditions. Schad et al. (1952) developed a breeding program to produce and select interspecific hybrids obtained by open or controlled crosses. Some of the French clones became very popular for nut production (Bergougnoux et al. 1978; Breisch 1995).
The first interspecific hybridizations in Portugal were initiated in 1947 by Bernardino Barros Gomes to introduce resistance to ink disease in C. sativa (Gomes Guerreiro 1948, 1957). More recently, interspecific crosses made between Castanea sativa ‘Aveleira’ (mother tree) with pollen of C. crenata (SC) or C. mollissima (SM) were done as a first step to identify molecular markers associated to ink and blight disease resistance in chestnut for developing marker assisted selection (MAS) and as a tool to identify genomic regions linked to resistance (QTLs) (Batista et al. 2008). Resistant C. sativa selections from COLUTAD in Portugal are being tested in a micropropagation program to rapidly make these available to the producers.
Chestnut blight is caused by Cryphonectria parasitica (Murr.) Barr (Syn. Endothia parasitica [Murr.] And.). It became the major disease of chestnut due to the sensitivity of C. dentata as well as, although to a lesser degree, of C. sativa to this fungus. Blight destroys the bark and the cambium causing the death of the branches or the tree above the wound when the disease girdles around them (Anagnostakis 1987; Heiniger and Rigling 1994). It was first observed in Europe in Genoa, Italy in 1938. The spread was quick through Italy and other European countries (Robin and Heiniger 2002), and less so in Southern UK, the Netherlands, Central and Southern Spain, and the Canary Islands. Blight almost eliminated the American chestnut (C. dentata) but European chestnut is recovering due to the natural occurrence of hypovirulence dsRNA hypovirus CHV1. Allemann et al. (1999) isolated five different CHV1 subtypes. Biological control is applied in Europe by hypovirulent strains of hypovirus growing cankers using Grente’s method (Grente and Berthelay-Sauret 1978).
Only two loci conferring resistance to Cryphonectria parasitica have been identified within germplasm of C. sativa, C. mollissima, and C. crenata (Sisco et al. 2005). The main origin of resistance to blight is coming from Asian species, mainly C. mollissima (Hebard and Stiles 1996). The American Chestnut Foundation has developed a backcross-breeding program to restore the American chestnut C. dentata. By the third backcross, the progenies reach on average 96% American background, which eventually exhibit entirely American characteristics in later generations (Diskin et al. 2006).
Pests. Oriental chestnut gall wasp (Dryocosmus kuriphilus Yasumatsu) causes very serious loss of nuts production in Japan, China and Korea. This insect is a tiny gall-forming wasp endemic to China, and was accidentally introduced into Japan (1941), Korea (1963), and the USA (1974). The larvae in the gall can cause extensive bud loss, decreased shoot and leaf growth and reduced nut production. Severely affected trees can die. Therefore, the Japanese chestnut breeding programs have focused their work over the last several decades on the development of resistant cultivars to the chestnut gall wasp. These programs have developed several resistant cultivars through the screening of germplasm and selective breeding. They have bred 4 resistant cultivars: ‘Tanzawa’, ‘Ibuki’, and ‘Tsukuba’ in 1959 and ‘Ishizuchi’ in 1968. The cultivars were planted extensively throughout Japan allowing a recovery of the chestnut industry. However, these resistances broke down, and two cultivars with better resistance to gall wasp were released in 1981 (‘Kunimi’) and 1992 (‘Shiho’).
Another strategy to overcome damage by chestnut gall wasp was biological control by the introduction of the natural enemy of the chestnut gall wasp, Torymus sinensis. It was introduced into Japan in 1979 in Fukuoka City in Kyushu, the western island of Japan, and then in 1980 at NIFTS (Tsukuba City) in Ibaraki in eastern Japan. As a result, it is difficult to find chestnut gall wasps in chestnut orchards today. Breeding of resistant cultivars combined with the use of biological control by natural enemies have contributed to overcome chestnut gall wasp.
In 2002, gall wasp was reported for the first time in Europe in northwest Italy. The cynipid there causes the development of galls on leaves, buds and inflorescences, resulting in a decrease in both growth and yield of the European chestnut. Although the biological control with the parasitoid Torymus sinensis Kamijo, recently introduced in northwest Italy from Japan, may be a promising method for reducing the pressure of the pest in chestnut forests, this will most likely not be enough to guarantee high yield and good nut quality in orchards. For this reason, the University of Torino (Sartor et al. 2007) is assessing the level of susceptibility to D. kuriphilus in Castanea sativa Miller and hybrid cultivars for developing a breeding program. So far C. sativa cultivars tested are susceptible to gall wasp, although at different levels. Yet, resistance sources were recently discovered in the C. sativa germplasm. In this case plants do not show gall development. Among the Euro-Japanese hybrids, ‘Bouche de Bétizac’ and ‘Marsol’ showed opposite reactions to the insect: no gall development was observed in ‘Bouche de Bétizac’, while the highest level of infestation was observed in ‘Marsol’. ‘Bouche de Bétizac’ has a hypersensitive response to infestation which results in larvae death at budburst. If the trait will remain stable, the selected materials will be used for orchard planting or for breeding resistant individuals. Preliminary work to understand the genetic mechanism of the susceptible response to the cynipid presence in chestnut tissues showed the expression of genes probably related to differentiation, nourishment, and the ability of the larvae to switch on parts of the seed development pathway.
Other two main insect pests are the moth larvae Cydia (=Laspeyresia) splendana Hb. and the weevil Curculio (=Balaninus) elephas Gyll. Cydia penetrate the nut through the bur as neonate larvae, and Curculio females oviposit through the bur. Debouzie et al. (1996) demonstrated that presence of chestnut moth larvae inhibited weevil egg-laying. It appears that Curculio ovoposits less in those cultivars with longest bur spines (Bergougnoux et al. 1978).
4.2 Scion
Nut quality. In a large study of Spanish cultivars (Pereira-Lorenzo et al. 2006a), only 7% of samples came from cultivars producing nuts over 15 g, and most cultivars produced smaller and less valued nuts (under 10 g). Of the four main nut shapes found, the most common are the elliptical-short (46%), the elliptical-triangular (30%), and elliptical-broad (18%). The most distinct and least common is the triangular shape (6%), which gives the name ‘Longal’ to the main cultivar in the Iberian Peninsula. Only 12% of the Spanish cultivars studied had more than 12% of multi embryo chestnuts. This indicates the strong selection made by growers to avoid peeling problems.
Asian species mature their nuts more quickly (early September) than does C. sativa (late October). This early harvest from the Asian species has been genetically transmitted to interspecific hybrids (Pereira-Lorenzo and Fernandez-Lopez 2001). However, important genetic variability is found in Spanish cultivars, a product of selection, with cultivars in South Spain production collected during the second half of September combined with cultivars harvested in October, and in the North with cultivars such as ‘Negral’ harvested at the beginning of October when the most frequent period is at the end of that month.
Chestnut pericarp split when the rainfall season is delayed till October, facilitating the development of fungus. This is not related to genetic variation (Ramos-Cabrer and Pereira-Lorenzo 2005) as embryo intrusions of the pericarp were found to be related to region and cultivar variation.
Another characterestic selected by growers is the shortness of the bur’s spines, only present in 14% of cultivars such as ‘Rapada’ or ‘Rapuga’, which are easy to harvest by hand (Pereira-Lorenzo et al. 2006a).
Larvae and weevils develop inside mature nuts, Cydia (=Laspeyresia) splendana Hb. and Curculio (=Balaninus) elephas Gyll. Cydia penetrate in the nut through the bur as neonate larvae and Curculio females make their ovoposition using with her long snout also through the bur. In Spanish cultivars, we confirmed the relationship between the length of the burr’s spines and lower attacks of Curculio as was previously reported by Bergougnoux et al. (1978). Burs with longest spines, longer than 1.5 cm, reduced the attacks up to 34%. Thus, the selection of long spur spines should be considered in breeding programs as an approach to develop Curculio resistant cultivars.
Chestnuts store starch in cotyledons, and the content is three- to fourfold higher than that found in other nuts (Ensminger et al. 1995). C. dentata and C. mollissima show a higher starch content (49%) than C. sativa (40%) (McCarthy and Meredith 1988). High starch content is important in cultivars for flour production or for animal feed. The highest average starch content in the Iberian Peninsula is produced by ‘Longal’ (Ferreria-Cardoso et al. 1993; Pereira-Lorenzo et al. 2006b) with 53–67% d.m. Another chemical characteristic that should be taken into account is high fiber content that reduces digestibility in some cultivars.
Concerning Spanish cultivars, Pereira-Lorenzo et al. (2006b) concluded that high variability in chemical composition between cultivars and regions corresponded to high genetic variability between cultivars. Correlations with environmental parameters were low, indicating that differences found between regions were probably due to the differences between cultivars. In Central and Southern Spain, some cultivars presented lowest moisture content due to the low summer rainfall in these regions. Lowest values of fiber content and ease of digestibility were found in cultivars from Galicia and Extremadura. No significant differences in Fe, Zn, and Cu were found, although Zn content is twice the value reported for European chestnuts.
Japanese chestnuts produce the largest nuts among Castanea spp., sometimes exceeding 30 g. Nut size is one of the most important morphological characteristics and many newly bred cultivars have nuts weighing 25–30 g. By contrast, the thick pellicle is often extensively invaginated into the nuts, which makes peeling difficult. The easy peeling characteristic in kernel is a major objective in Japanese chestnut breeding programs.
4.3 Rootstocks
Compatibility. The most popular clonal rootstocks are the Euro-Japanese hybrids selected in France. They are easy to propagate by layering or softwood cuttings, are tolerant to Phytophthora spp. and Cryphonectria parasitica and have genetic compatibility with most of the best cultivars. Popular rootstocks include: ‘CA 07’ (‘Marsol’) (moderately resistant to Phytophthora); ‘CA 74’ (‘Maraval’) (Phytophthora resistant, low vigor); ‘CA 118’ (‘Marlhac’) (moderately resistant to Phytophthora, but able to grow at temperature <−10°C); ‘CA 90’ (‘Ferosacre’) (Phytophthora resistant, but sensitive to temperatures <−10°C). European chestnut cultivars are usually grafted onto seedlings of C. sativa.
The ease of vegetative propagation and stock–scion compatibility are of primary importance in rootstock breeding. Chapa et al. (1990) and Bounous et al. (1992) found that C. crenata hybrids (C. crenata × C. sativa) are easier to propagate by cuttings or layering than C. sativa. Ease of propagation by layering or cuttings and Phytophthora-resistance of the French hybrids (‘Marsol’, ‘Marigoule’, ‘Maraval’, ‘Précoce Migoule’) make them useful as rootstocks or as direct producers. Unfortunately, graft incompatibility problems with many European cultivars have limited their wider application (Chapa et al. 1990; Ferrini et al. 1992; Breisch 1992; Pereira-Lorenzo and Fernández-López 1997).
Although environmental and stress factors may have a role, the success of a particular graft, stock–scion compatibility is most certainly under genetic control (Anagnostakis 1991). Three peroxidase isozyme genes known for Castanea (six types) may be involved with graft compatibility (Santamour et al. 1986). Graft incompatibility is also affected by ChMV (Desvignes 1996).
5 Current Goals of Breeding
The chestnut ideotype is a function of the final use (nuts or timber), and production and processing technology (harvesting systems, fresh or processed uses) (Table 19.10). For nut production the most important breeding objectives include the following: good horticultural traits, product quality, suitability to storage and processing, and easy peeling.
For timber, important characters include wood quality, rapid growth, and nonchecking of wood (ring-shake). Ease of propagation and resistance to major diseases and pests are common for nut and timber types. A few chestnut cultivars such as ‘Garrida’, ‘Loura’, and ‘Parede’ combine both timber and chestnut production under the appropriate climatic conditions.
Japanese chestnut breeding trials started in 1929 at the Agricultural Experiment Station in Niigata Prefecture. Originally, the Japanese chestnut was crossed with a Chinese chestnut in an attempt to introgress the easy peeling kernel trait from Chinese chestnut. However, these efforts were unsuccessful. In 1947, national chestnut breeding programs started at the Horticultural Research Station (now National Institute of Fruit Tree Science: NIFTS). The breeding target at that time was to produce a cultivar with the Japanese chestnut characters of high yield and big nut size combined with the character of easy peeling kernel of Chinese chestnut. Before this was achieved, the objective was changed to chestnut gall wasp resistance in 1952 because the chestnut gall wasp which appeared in Okayama Prefecture in western Japan around 1941 quickly spread to the whole country. Since the Chinese chestnut breeding material had little resistance to gall wasp, two gall wasp resistant native cultivars ‘Ginyose’ and ‘Ganne’ were used to breed gall wasp resistant cultivars.
Once the damage caused by the chestnut gall wasp decreased, the goal of Japanese chestnut breeding apparently shifted from insect resistance to the nut quality. The decrease of insect damage enabled the use of the Chinese chestnut for breeding. At the same time, a new rapid screening system for removing the chestnut pellicle after deep frying in cooking oil (HOP method, High-temperature Oil Peeling method) was developed and applied to chestnut breeding. By using the HOP method, easy peeling Japanese chestnut cultivars were identified and used in breeding. The newest promising cultivar with an easy peeling trait, ‘Porotan’, bred and registered in 2007. But it was developed from Japanese chestnuts without the easy peeling trait. The genetic control of the easy peeling trait of ‘Porotan’ is unknown.
5.1 Plant Characteristics
Semicompact, medium, or low vigor are the most suitable features for medium or high density plantations. Other valuable cultivar characteristics include the following: early maturity, precocious bearing, regular and high yields, strong branches, good pollinizer ability, and intercompatibility with the best cultivars. Harvesting is one of the most costly aspects of chestnut production. Harvest-related traits include upright habit for mechanical shaking and low detaching force to shake off burs from the tree. Mechanical harvesting of the nuts from the ground may be easier with nuts that fall closed in the burs (to prevent nuts from infection), than with nuts which fall free from dehiscent burs. For timber production, trees have to demonstrate high vigor, high wood production, straight trunk, self-pruning ability, and wood not subject to ring-shake or radial checking.
5.2 Nut Characteristics
Large nut size is desirable from the standpoint of harvesting, handling, fresh marketing and making candy (marrons glacés), while a small or medium size nut may be used for dried chestnuts or as a vegetable. The marketing of peeled or processed chestnuts puts less emphasis on size. Evenness of shape, shiny color, dark brown stripes, flavor, and firm texture are valuable traits for fresh marketing. Other desirable traits are easy pellicle removal, no pellicle intrusion, no hollow kernel, no multiembryo nuts, and resistance to pests (Cydia, Curculio and others) and to storage diseases (Cyboria and others).
Japanese chestnut breeding programs have focused on breeding excellent cultivars with high nut quality. The major selection criteria for the NIFTS programs are large nut size, low pericarp splitting, low polyembryony, white color of the steamed kernel, high sweetness, flavor, and kernel quality. In addition, an easy peeling pellicle is highly desired but a difficult characteristic to obtain for more than 50 years of breeding. The newly released cultivar ‘Porotan’ is the most prominent cultivar with good nut quality and an easy peeling trait for East Asian Markets.
6 Breeding Methods and Techniques
6.1 Ploidy, Karyotype, and Genome Size in Castanea
Castanea spp. are generally recognized as having 2n = 24 chromosomes (Jaynes 1962), the number characteristic of most of the Fagaceae studied to date (Mehra et al. 1972; Ohri and Ahuja 1991; D’Emerico et al. 1995). With only minor exceptions such as occasional triploidy and aneuploidy (Jaynes 1962), the normal somatic number of chromosomes in species and hybrids in the genus is 24. In general, a rather high degree of fertility has been observed among interspecific hybrids (Jaynes 1964, 1972). Although pairing appears to be normal in many interspecific hybrids, the presence of segregation distortion in some mapping populations (Kubisiak et al. 1997; Kubisiak, unpublished data) and abnormal pairing in F1 hybrid pollen mother cells (Faridi et al. 2008), suggest that significant chromosomal differences such as translocations and/or inversions are likely to exist among Castanea species. A better understanding of chromosome-level genomic differences between species will be important for effective breeding using interspecific hybrids. Genome size appears to be fairly conserved among species (2C = 1.57–1.67 pg), only five times that reported for Arabidopsis. A tractable genome size and abundant genetic and genomic resources make Castanea a good candidate for future targeted- or whole-genome sequencing (Kremer et al. 2007).
6.2 Mating System
Chestnut is a monoecious species presenting male flowers in catkins and female flowers that develop at the base of bisexual catkins. Some chestnut cultivars present morphological male sterility. Soylu (1990) proposed a genetic model based on two genes and five morphotypes: astaminate (xxzz), brachystaminate (xxZz), mesostaminate (Xxzz), and longistaminate long/short (XXZZ/XxZz). Astaminate flowers do not produce pollen, brachystaminate can produce very limited quantities of pollen, while longistaminate catkins are those that produce more pollen. Astaminate catkins are supposed to be more frequent in some of the best ‘marron’ type cultivars. Male sterility may be related to lower energy consumption during flowering. Male sterility has been found in up to 21% of Spanish cultivars. Up to 8% of the cultivars presented astaminate catkins, and 13% had brachystaminate catkins. Those cultivars require pollinizers, mainly with longistaminate catkins (Pereira-Lorenzo et al. 2006a).
Pistillate flowers have six to eight styles whose tips are hollow at full bloom. The ovary presents seven (rarely six or eight) carpels. Each flower has 10–16 anatropous ovules. The bearing monoembryonic seeds (marron type) have been related to a high occurrence of anomalies, such as delayed embryo sac differentiation and the presence of supernumerary nuclei in the embryo sac (Botta et al. 1995).
Very little is still known about the genetic system controlling mating and the self-incompatibility system in chestnut, although it is considered to be of gametophytic type (Breviglieri 1951; Brewbaker 1957; Jaynes 1975). Cross-pollination is compulsory.
6.3 Hybridization
Interspecific hybridizations between most of the chestnut species are possible. Main problems are related with the different flowering time between species. Asian species are more precocious than European chestnut. Hybridization was the main breeding method used to incorporate resistance to blight into American chestnut, ink disease into European chestnut and wall gasp into Japanese chestnut (Table 19.3). It was also used in Japan to incorporate the easy peeling trait into Japanese cultivars, which were normally difficult to peel.
Meiosis in pollen mother cells occurs 10 to 15 days before anthesis, in the first week of June in Italian cultivars (Botta et al. 1995). Pollen viability varied from 81.3 ± 6.1% based on fluorochromatic reaction, to 58.2 ± 7.0% on hanging drops and 50.1 ± 4.5% germination on agar media.
Pollen is easily collected from the longistaminate pollinizers and desiccated to be stored in a refrigerator for short-term storage or in a freezer for long-term storage. Emasculation and female flower isolation is needed to avoid unknown pollination. Male flowers from the bisexual catkins must be also removed. Bags are attached to the base of the catkins. Hand pollination with fresh catkins brushing over pistils has been detailed by Nienstaedt (1956). The bags are removed after setting. Nuts are collected when burs begin to crack and nuts begin to turn brown.
European breeding programs to incorporate resistance to ink disease (Phytophthora spp.) produced interspecific hybrids of first generation. In the USA, a backcrossing program using blight resistant Chinese species as the donor and American chestnuts (C. dentata) as the recurrent parent has produced blight resistant American chestnut (Diskin et al. 2006). Diskin et al. (2006) pointed out that, in the BC3-F2 generation, 93% of seedlings showed morphological characteristics of American chestnut incorporating 100% Chinese resistance to blight. The program to obtain such material required more than 20 years.
6.4 Propagation
Seeds. Seedlings are used in breeding programs based on hybridization (Table 19.3), as local rootstocks for main producing areas (Pereira-Lorenzo et al. 2007), but also to propagate selected forest progenies in Europe. Wild progenies from Europe have shown adaptive response to water stress such as a lower growth rate (Pliura and Eriksson 2002; Lauteri and Villani 2004; Eriksson et al. 2005) and different bud break (Blanco Silva et al. 2005).
Chilling releases embryos from endodormancy. To avoid dehydration which reduces germination, chestnuts are stratified in sand or in humid peat moss at 1–2°C for 4 or 5 months.
Pregerminated nuts are sown in raised beds. The apex can be cut off to permit the formation of a well expanded root system. The depth of seeding must not exceed 3–5 cm, spacing in the row is usually 30–40 cm, with rows 80–100 cm apart.
At the end of the first growing season (August–September), the seedlings are 100–150 cm tall with a diameter of 8–12 mm and are ready to be chip budded in fall or grafted or budded the following spring.
Grafting. Grafting is used not only for propagation of selected cultivars but also in breeding programs to establish core collections and germplasm banks, to propagate mature origin such as plus trees, and to rejuvenilize plant material for later propagation by cuttings, layering, and micropropagation. Chestnut cultivars have been traditionally propagated using bark graft in spring or flute graft, and continue to be used in new orchards when in situ graft is made. Nurseries use various budding and grafting techniques. Summer budding (patch and T-bud) are easier and more effective since the higher temperatures cause rapid healing (Pereira-Lorenzo and Fernández-López 1997). However, summer budded trees are not ready for the market until the following winter. By contrast, spring grafting (chip, cleft, and whip) can produce finished plants for the same winter, although they need a second year growth to get to comparable growth with those summer-budded plants.
Cuttings. Vegetative propagation of chestnut is limited by the age of the mother plant from which the cuttings are taken. Cuttings from material of mature origin are difficult, if not impossible, to root (Vieitez 1974). The most efficient system is through the use of greenwood or softwood cuttings collected from 3- to 4-year-old mother plants, which can be grown in the open or as potted plants in the greenhouse. Cuttings, bearing 3–4 buds (10–15 cm) and leaving half of the uppermost leaf, are collected from May to July and their bases treated with indole-3-butyric acid (IBA) or naphthalene-3-acetic acid (NAA) at concentrations ranging from 1,000 to 4,000 ppm (Rodríguez et al. 2005). The cuttings are subsequently placed in trays, containing suitable substrates, in rooting tunnels equipped with a fog system. Rodríguez et al. (2005) showed that the production efficiency of the system is related to the genotype.
Layering. Chestnut layering is another vegetative propagation method mainly used in European countries for the clonal propagation of rootstocks, resistant Euro-Asian hybrids or direct producer trees. Sprouts are developed annually from stumps and, regardless of the age of the mother plant, exhibit a juvenile physiological condition, allowing them to root after proper auxin treatments. When the sprouts reach a length of 30–35 cm (May), their basal parts (8–10 cm) are stripped of leaves and a paste containing IBA and NAA (or both) at a rate of 2 g/kg Vaseline® is applied to this part of the sprout, which is finally covered with soil. When the shoots reach the rest period (November), the rooted shoots are excised from the mother stump and planted in the nursery to strengthen their root system. The number of shoots produced yearly per stump (10–20) and the rooting rates (60–90%) are clearly genotype dependent.
In vitro. Efforts are being made to establish reliable in vitro regeneration systems that allow chestnut clonal propagation. The two principal micropropagation systems are based on somatic embryogenesis and on micropropagation through axillary shoot development. Although somatic embryogenesis is theoretically more efficient for clonal mass production than propagation via axillary shoot proliferation, several difficulties need to be overcome to render it commercially viable, particularly when cultures originate from adult tissues (Corredoira et al. 2006). By contrast, chestnut can currently be micropropagated from both juvenile and mature material using the axillary shoot multiplication method, although it is common for the protocol to require optimization for a specific cultivar, making the large-scale propagation in many cases challenging (Vieitez et al. 2007).
Juvenile plant material may be collected from seedlings conventionally obtained in greenhouse or climate chamber, as well as from seedlings obtained by in vitro culture of embryonic axes (Sánchez et al. 1997a). In the case of mature material, the use of stump sprouts (juvenile parts of mature trees) or basal shoots grown on the lower part of the trees as source of explants allows the micropropagation of chestnut. Cuttings taken from these materials are collected in winter, stored at 4°C and forced to flush in a climate chamber, after which the primary explants are taken from the flushed shoots. By contrast, the reactivity of crown-derived primary explants is poor, and reinvigoration methods must be applied using pretreatments such as etiolation (Ballester et al. 1989) or grafting onto seedling rootstocks (Sánchez et al. 1997b).
The primary explants from which chestnut shoot cultures are initiated are generally shoot tips and nodes bearing 1 or 2 axillary buds. After excision from the plant source (juvenile or mature trees), they must be sterilized and established in vitro on a number of different initiation media supplemented with cytokinins. After 6–8 weeks of culture in initiation medium, new shoots develop which can be subcultured at 4- to 5-week intervals for the shoot multiplication culture stage. From the different systems employed for rooting of micropropagated chestnut shoots, the culture for 24 h in a rooting medium containing 25–50 mg/l IBA followed by transference to either an auxin-free root expression medium or to a substrate mixture appears as the most appropriate for obtaining acceptable rooting frequencies.
In addition to clonal propagation, in vitro tissue culture is a useful technique for germplasm conservation. The cold storage of cultures represents a procedure for medium-term conservation, and Janeiro et al. (1995) reported the possibility of keeping chestnut cultures at 2–4°C for up to 1 year without subculture. In addition, a successful cryopreservation system of chestnut shoot tips has been reported (Vidal et al. 2005) allowing long-term storage of chestnut genotypes in liquid nitrogen. A detailed protocol for micropropagation of European chestnut, including storage and molecular marker analysis to determine the genetic stability of in vitro regenerated plants, has recently been published (Vieitez et al. 2007).
7 Integration of New Biotechnologies in Breeding Programs
7.1 Genomic Resources for Castanea
A number of molecular marker systems have been used in Castanea for applications such as cultivar identification, population genetics, linkage analysis, and marker-assisted selection. These marker systems consisted initially of isoenzymes, followed by random amplified polymorphic DNAs (RAPDs), inter-simple sequence repeats (ISSRs), and amplification fragment length polymorphisms (AFLPs), and more recently by simple sequence repeats (SSRs) and single nucleotide polymorphisms (SNPs). Given that SSRs and SNPs are rapidly becoming the markers of choice due to their informativeness, high transferability across related taxa, prevalence in the genome, and amenability to automated high-throughput analysis, we have chosen to focus primarily on the development and use of these newer marker systems.
There are currently four main sources from which SSR or SNP markers are being developed: genomic DNA libraries enriched for repeat-containing sequences; expressed sequence tags (ESTs); bacterial artificial chromosome (BAC) sequencing, or from whole genome sequencing efforts. Owing to a previous lack of DNA sequence data available for the Fagaceae, and in particular for Castanea, most SSRs currently available have been developed from repeat-enriched genomic DNA libraries. A small number of SSR markers developed from Quercus sequences are proving useful in Castanea (Aldrich et al. 2003; Barreneche et al. 2004). Of those developed from Fagus (Tanaka et al. 1999; Pastorelli et al. 2003), comparatively fewer are proving to be useful within the Castanea (Kremer et al. 2007; Kubisiak, unpublished data). The transferability of SSRs from other genera within the Fagaceae to Castanea will largely depend upon their evolutionary distance, with higher levels of transferability expected between more closely related genera such as Castanopsis (Manos et al. 2001).
Sequence data for at least 83 SSR primer pairs developed specifically from Castanea are currently publicly available. Forty-six SSR primer pairs developed from C. sativa have been characterized (Buck et al. 2003; Marinoni et al. 2003). Yamamoto et al. (2003) characterized 15 SSR primer pairs developed from C. crenata. More recently, 22 SSR primer pairs were developed from C. mollissima and characterized in both C. mollissima and C. crenata (Inoue et al. 2009). In general, these markers appear to be highly transferable across the Castanea. SSRs are already proving useful for cultivar identification and typing (Boccacci et al. 2004; Beccaro et al. 2004; Pereira-Lorenzo et al. 2010), linkage analysis (Sisco et al. 2005), and QTL analyses (Kubisiak, unpublished data; Casasoli et al. 2004, 2006).
Previously, the availability of DNA sequence data for Castanea was extremely limited (Connors et al. 2001). More recently, a genomic tool development project for various members of the Fagaceae was initiated (Sederoff et al. 2008), with C. mollissima being a key model species. A large component of this project is focused on the transcriptomes of C. mollissima and C. dentata (Carlson et al. 2007, 2008). Large EST databases are being created with significant numbers of sequence contigs showing similarity to predicted proteins in Populus trichocarpa. Additional SSRs and a significant number of SNPs are being identified in EST sequence data that should prove to be an invaluable resource for the community interested in all aspects of the genetics, breeding, and biotechnology of Castanea. An integrated Web-based resource for the Castanea genetics/genomics community [Fagaceae Genomic Database (FGD): http://www.fagaceae.org] has been developed (Ficklin et al. 2007), and relevent sequence information, homology results, genetic/physical map information, SSRs, SNPs, and other genomic data are being posted as it become available.
Another large component of the genomic tool development project is the production of genetic and physical mapping resources for C. mollissima. A BAC library has been constructed which consists of ∼20× coverage (http://www.fagaceae.org/progress/NE1015/Tomkins_FingerprintingAndcDNAUpdate.ppt). The entire library is currently being fingerprinted by high information content fingerprinting. A subset of clones consisting of the minimum tiling path will be selected for BAC-end sequencing. Genetic markers developed from BAC-end sequence and hybridization of genetic marker probes to high-density BAC colony filters will be used to combine the genetic and physical maps. An integrated genetic/physical map will become the platform for future targeted genome sequencing of regions harboring resistance genes and will be an invaluable resource for gene cloning studies in Castanea.
Three partial gene encoding proteins described as pathogenesis-related were isolated and cloned from infected resistant chestnut plants: a cystatin, a beta 1,3 glucanase isoform, and a thaumatin-like protein gene using the RT-PCR technology (Serrazina 2004). The nucleotide sequences and amino acid deduced sequences have high homology with resistance genes’ sequences of other plant species in the GenBank database. A partial gene encoding AOC (allene oxide cyclase) was also cloned, similar to the Lycopersicum esculentum gene. The gene expression analysis by Northern Blotting of aoc, cist, gluc, and pttaum points out to a straight relation of these genes with C. sativa resistance to P. cinnamomi (Serrazina 2004). Tobacco explants were transformed by particle bombardment, to study the overexpression effect of the isolated genes on plant resistance to P. cinnamomi. The observation of inoculated transformed and nontransformed plantlets suggests that the constitutive expression of aoc, cist, gluc, and pttaum genes (separately) attenuated the pathogenic effects of P. cinnamomi in transformed tobacco plants (Serrazina 2004).
To gain insight into genetic reorganization, which leads to the formation of ectomycorrhiza, a cDNA microarray was constructed and used to study the interaction of Castanea sativa roots and Pisolithus tinctorius during the first hours of contact. (Sebastiana 2006). Statistical analysis of microarray results identified a set of 32 C. sativa genes and 8 Pisolithus tinctorius genes with altered expression in response to the interaction between the two organisms. Differentially expressed genes identified in C. sativa roots displayed significant sequence similarities to proteins involved in cellular processes such as defense response, protein maturation/degradation, cell wall modification, primary metabolism, signal transduction, and cytoskeletal organization. Fungal genes regulated by the interaction with C. sativa roots displayed significant sequence similarities to proteins involved in cell wall structure, protein maturation/degradation, and cellular organization (Sebastiana 2006).
Linkage relationships between isoenzymes and morphological traits in interspecific crosses were found (Huang et al. 1996). Molecular maps have been developed (Kubisiak et al. 1997; Casasoli et al. 2001) opening a new way to the genetic knowledge of chestnut.
7.2 Transgenics
A conventional chestnut breeding program requires around 15–20 years to incorporate a new disease resistance allele and to reach the BC3F2 generation. The great advantage of genetic transformation, as a complementary breeding system, is that a new set of genes can be transferred into chestnut somatic cell lines in a matter of 2–3 years. This is possible today because, after decades of research on in vitro tissue culture of both European and American chestnut (Vieitez and Merkle 2004), a repetitive and reproducible system useful for genetic transformation through somatic embryogenesis has been described for both species (Robichaud et al. 2004; Corredoira et al. 2006). Although in most cases the induction of somatic embryos was carried out from immature material (embryonic axes), Corredoira et al. (2003) have shown that embryogenic cultures can be initiated from leaf explants of seedlings, opening up the possibility of propagating and transforming mature material in the future.
The first attempts to transform European chestnut used hypocotyl segments from in vitro-germinated seedlings and stem segments of in vitro-grown shoots, which were cocultured with Agrobacterium tumefaciens (Seabra and Pais 1998, 1999). Unfortunately, no transgenic plants were obtained as there were a large number of escapes (97%) and gene integration was transient. On the other hand, some transformation experiments were carried out by coculturing cotyledonary node explants with A. tumefaciens harboring reporting genes, with 2.3% of explants developing kanamycin-resistant shoots, although no transgenic plants were recovered (Corredoira et al. 2005).
An efficient genetic transformation protocol for C. sativa has been described for the first time (Corredoira et al. 2004) through the coculture of somatic embryos with different strains of Agrobacterium tumefaciens carrying marker genes. The plasmids contain the nptII genes driven by the nos promoter for kanamycin selection and the β-glucuronidase reporter uidA gene (gus) driven by either ubiquitin (Ubi-1) or the CaMV 35S promoter. Following 4 days of coculture and after 12 weeks of culture in selection medium, cotyledonary stage-regenerated embryos were isolated from GUS-positive lines and subcultured on selection medium to establish and proliferate embryogenic transgenic lines. The presence of the nptII and the uidA genes in GUS-positive embryogenic lines was assessed by PCR and Southern blot analyses. Transformation efficiencies as high as 25%, were recorded. This transformation protocol was improved by studying the effect of both the genotype and the type of initial explant (Corredoira et al. 2007). In order to increase the tolerance of European chestnut to fungi diseases, attempts to transform embryogenic lines with a thaumatin-like protein (CsTL1), which has antifungal activity in vitro, are currently in progress (Maynard et al. 2008).
The first reports on transgenic American chestnut tissues were carried out by Carraway et al. (1994) who used microprojectile bombardment (biolistics) to transform proembryogenic masses derived from immature zygotic embryos, although no stable transformation events were recovered. Subsequently, a coculture transformation experiment was carried out with Agrobacterium tumefaciens containing the plasmid construct pDVspB-OxO, which included a germin-like oxalate oxidase gene (OxO) to enhance blight resistance, a phosphinothricin acetyltransferase gene (bar) as a selectable marker, and a green fluorescent protein gene (mgfp5-ER) as a visual marker (Polin et al. 2006; Maynard et al. 2008). To increase the transformation rate, the plate flooding system approach was assayed (Rothrock et al. 2007).
One problem affecting both European and American chestnut is the relatively low conversion rate of somatic embryos into plants (Corredoira et al. 2003, 2004). However, germinating embryos with only shoot development are also produced in germination medium, and these shoots can be multiplied successfully by axillary shoot proliferation, giving rise to an unlimited number of transgenic shoots to be rooted. This constitutes a valuable alternative for plant regeneration from transformed germinating embryos in which plantlet conversion is not achieved. Both European and American transgenic chestnut plants were acclimatized in phytotron and grown in the greenhouse and in the open fields (Maynard et al. 2008).
In addition to this consolidated transformation procedure, Fernando et al. (2006) described a promising preliminary approach consisting in the transformation of America chestnut pollen. The method, in which in vitro tissue culture technology is not required, makes use of transformation via particle bombardment. Only transient green fluorescent protein (GFP) expression was recorded and the highest values were achieved using ungerminated pollen.
References
Aldrich, P.R., Michler, C.H., Sun, W., Romero-Severson, J. (2002) Microsatellite markers for northern red oak (Fagaceae: Quercus rubra). Mol. Ecol. Notes., 2:472–474.
Aldrich, P.R., Parker, G.R., Michler, C.H., Romero-Severson, J. (2003) Whole-tree silvic identifications and the microsatellite genetic structure of a red oak species complex in an Indiana old-growth forest. Can. J. Forest Res., 33:2228–2237.
Allemann, C., Hoegger, P., Heiniger, U., Rigling D. (1999) Genetic variation of Cryphonectria hypoviruses (CHV1) in Europe, assessed using restriction fragment length polymorphism (RFLP) markers. Mol. Ecol., 8:843–854.
Anagnostakis, S.L. (1987) Chestnut blight: The classical problem of an introduced pathogen. Mycologia, 29:23–37.
Anagnostakis S.L. (1991) Peroxidase allozyme phenotypes in Castanea and their segregation among progeny. HortSci., 26:1424.
Antonaroli, R., Bagnaresi, U., Bassi, D. (1984). Indagini sulla variazione di alcuni caratteri morfologici in popolazioni di castagno da frutto nella provincia di Bologna. Monti e Boschi, 2, 47–50.
Aravanopoulos, F.A., Drouzas, A.D., Alizoti, P.G. (2002) Electrophoretic and quantitative variation in chestnut (Castanea sativa Mill.) in Hellenic populations in old-growth natural and coppice stands. For. Snow Landsc. Res., 76(3):429–434.
Aravanopoulos, F.A., Bucci, G., Akkak, A., Blanco Silva, R., Botta, R., Buck, E.,Cherubini, M., Drouzas, A.D., Fernandez-Lopez, J., Mattioni, C., Marinoni, D., Papadima, A., Russel, K., Zas, R., Villani, F. (2005) Molecular population genetics and dynamics of chestnut (Castanea sativa Mill.) in Europe: inferences for gene conservation and tree improvement. Acta Hort., 693:403–411.
Ballester,A., Sánchez, M.C., Vieitez, A.M. (1989) Etiolation as a pretreatment for in vitro establishment and multiplication of mature chestnut. Physiol. Plant., 73:77(3):395–400.
Barreneche, T., Casasoli, M., Russell, K., Akkak, A., Meddour, H., Plomion, C., Villani, F., and Kremer, A. (2004) Comparative mapping between Quercus and Castanea using simple-sequence repeats (SSRs) Theor. Appl. Genet., 108:558–566.
Bassi, D., Sbaragli, E. (1984) Indagine pomologica su alcuni cloni di castagno da frutto (C.sativa Mill) Rivista di Frutticoltura Nº 6. Italia.
Batista, D., Valdiviesso T., Santos, L. Paulo, O., Laranjo, J, Costa, R. (2008) Genotyping Castanea sativa × C. crenata and C. sativa x C. mollissima F1 Hybrids using Nuclear SSRs. Acta Hort., 784:107–112.
Beccaro, G. L., Botta, R., Torello Marinoni, D., Akkak, A., Bounous, G., (2004) Application and evaluation of morphological, phoenological and molecular techniques for the characterization of Castanea sativa Mill. cultivars. Acta Hort., 693:453–457.
Bergougnoux, F., Verlhac, A., Breisch, H., Chapa, J. (1978) Le châtaignier. INVUFLEC, Paris, 192 p.
Blanco Silva, R., Silva, R., Zas, R., Fernández López, J. (2005) Geographic differentiation in adaptive traits of wild chestnut Spanish populations (Castanea sativa Miller). Investigación agraria. Sistemas y recursos forestales, 14:13–26.
Boccacci, P., Akkak, A., Marinoni, D.T., Bounous, G., Botta, R. (2004) Typing European chestnut (Castanea sativa Mill.) cultivars using oak simple sequence repeat markers. HortSci., 39:1212–1216.
Bonnefoi, C. (1984) Etude du polymorphism enzymatique des populations forestières de chataignier, Castanea sativa Miller. Thèse de Doctorat, Université des Sciences et Techniques du Languedoc, Académie de Montpellier, 142 pp.
Botta, R., Vergano, G., Me, G., Vallania, R. (1995) Floral biology and embryo development in chestnut (Castanea sativa Mill.). HortSci., 30:1283–1286.
Botta, R., Guaraldo, P., Mellano, M. G., Bounous, G. (2006) DNA typing and quality evaluation of chestnut (Castanea sativa Mill.) cultivars. International Symposium “Optimization, Productivity and Sustainability of Chestnut Ecosystems in Mediterranean Europe” Catania (Italy), 23–26 February 2005. Advances Hort. Sci., 1:96–100.
Bounous, G. (2002) Propagazione. In: Il castagno: Coltura, ambiente ed utilizzazioni in Italia e nel mondo. Bounous, G (ed.) Edagricole, Bologna, Italy, pp 47–63.
Bounous, G., Barone, E., Gioffré, D., Inglese, P, Zappia, R., Peano, C. (1989) Primi risultati dell’indagine sulle cultivar di castagno da fruttodiffuse in Calabria. L’Informatore Agrario, 45:53–57.
Bounous G., Bouchet M., Gourdon L. (1992) Reconstruction of traditional chestnut orchard. Experiences in Piedmont and Southern France. L’Inf. Agr., 9:155–160.
Bounous, G., Torello Marinoni, D. (2005) Chestnut: Botany, Horticulture, and Utilization. Hort. Reviews, Edit. J. Janick; Wiley & Sons Inc., New Jersey, vol. 31:291–347.
Breisch H. (1992) Compatibility tests between the main French varieties of chestnut trees and ink-resistant hybrid rootstocks. Proc. World Chestnut Industry Conference, Morgantown, West Virginia: 41–53.
Breisch, H. (1995) Châtaignes et marrons. CTIFL, 239 p.
Breviglieri N. (1951) Research on the flower and fruit biology in Castanea sativa and Castanea crenata in Vallombrosa territory. Publication n.1, Centro di Studio sul Castagno (C.N.R.) La Ricerca Scientifica (Suppl.) 21:15–49.
Breviglieri, N. (1955) Indagini ed osservazioni sulle migliori varietà italiane di Castagno. Centro di Studio Sul Castagno, Pubblicazione N.2, Supplemento a la Ricerca Scientifica, pp. 27–164.
Brewbaker, J.L. (1957) Pollen cytology and incompatibility systems in plants. J. Hered., 48: 217–277.
Buck E. J, Hadonou M., James C.J., Blakesley D., Russell K. (2003) Isolation and Characterisation of polymorphic microsatellites in European chestnut (Castanea sativa Mill.) Mol. Ecol. Notes, 10:1046–1048.
Burnham, C. R. (1988) The restoration of the American chestnut. American Scientist 76:478–487.
Camus, A. (1929) Les châtaigniers. Encyclopédie économique de sylviculture. P. Lechevalier, 3, 600 p.
Carlson, J., Powell, W., Tomkins, J., Ficklin, S., and Smith, C. (2007) GS20 454 sequencing of the chestnut transcriptome: results of a pilot study. Plant & Animal Genomes XV Conference, January 13–17, 2007, San Diego, CA. W127.
Carlson, J., Barakat, A., DiLoreto, D.S., Smith, C. (2008) The chestnut transcriptome. Plant & Animal Genomes XVI Conference, January 12–16, 2008, San Diego, CA. P25.
Carraway, D.T., Wilde, H.D., Merkle, S.A. (1994) Somatic embryogenesis and gene transfer in American chestnut. J. American Chestnut Foundation, 8:29–33.
Casasoli, M., Mattioni, C., Cherubini, M., Villani, F. (2001) A genetic linkage map of European chestnut (Castanea sativa Mill.) based on RAPD, ISSR and isozyme markers. Theor. Appl.Genet., 102:1190–1199.
Casasoli, M., Pot, D., Plomion, C., Monteverdi, M.C., Barreneche, T., Lauteri, M., Villani, F. (2004) Identification of QTLs affecting adaptive traits in Castanea sativa Mill. Plant Cell Environ., 27:1088–1101.
Casasoli, M., Derory, J., Morera-Dutrey, C., Brendel, O., Porth, I., Guehl, J-M., Villani, F., Kremer, A. (2006) Comparison of QTLs for adaptive traits between oak and chestnut based on an EST consensus map. Genetics, 172:533–546.
Castroviejo, S., Lainz, M., López, G., Montserrat, P., Muñoz, F., Paiva, J., Villar, L. (1990) Flora Ibérica. Plantas Vasculares de la Península Ibérica e Islas Baleares. Real Jardín Botánico, C.S.I.C., 2, 10–15 pp.
Chapa, J., Chazerans, P., Coulie, J. (1990) Multiplication végétative du châtaignier, amelioration par greffage de printemps et bouturage semi-légneux. L’Arboriculture Fruitierè, 431:41–48.
Conedera, M., Krebs, P. (2008) History, Present Situation and Perspective of Chestnut Cultivation in Europe. In: Abreu, C.G.; Peixoto, F.P.; Gomes-Laranjo, J. (eds): Proceedings of the second Iberian Chestnut Congress, Vila Real (P), June 20-22th 2007. Acta Hort., 784:23–27.
Conedera, M., Marcozzi, M., Jud, B. (1993) Banque de données sur les incendies de forêt au Sud des Alpes suisses. Proceedings of the Symposium “Contribution of European Engineers to Reduction of Natural Disasters”, Lausanne, 29–30 Sept. 1993. 165–171.
Conedera, M., Manetti, M-C., Giudici, F., Amorini, E. (2004a) Distribution and economic potential of the Sweet chestnut (Castanea sativa Mill.) in Europe. Ecologia mediterranea, 30: 2 p. 179–183.
Conedera, M., Krebs, P., Tinner, W., Pradella, M., Torriani, D. (2004b) The cultivation of Castanea sativa (Mill.) in Europe, from its origin to its diffusion on a continental scale. Veget Hist Archaeobot, 13:161–179.
Connors, B.J., Maynard, C.A., Powell, W.A. (2001) Expressed sequence tags from stem tissue of the American chestnut, Castanea dentata. Biotech. Letters, 23:1407–1411.
Corredoira, E., Ballester, A., Vieitez, A.M. (2003) Proliferation, maturation and germination of Castanea sativa Mill somatic embryos originated from leaf explants. Ann. Bot., 92:129–136.
Corredoira, E., Montenegro, D., San-José, M.C., Vieitez, A.M., Ballester, A. (2004) Agrobacterium-mediated transformation of European chestnut embryogenic cultures. Plant Cell Rep., 23: 311–318.
Corredoira, E., San-José, M.C., Ballester, A., Vieitez, A.M. (2005) Genetic transformation of Castanea sativa Mill. by Agrobacterium tumefaciens. Acta Hort., 693:387–393.
Corredoira, E., Ballester, A., Vieitez, F.J., Vieitez, A.M. (2006) Somatic embryogenesis in chestnut. In: Plant Cell Monographs, Vol. 2, Somatic Embryogenesis. Mujib, A. and Samaj, J. (eds.). Springer, Berlin, Heidelberg, 177–199.
Corredoira, E., San-José, M.C., Vieitez, A.M., Ballester, A. (2007) Improving genetic transformation of European chestnut and cryopreservation of transgenic lines. Plant Cell Tissue Organ Cult., 91:281–288.
Costa, R., Ribeiro, C., Valdiviesso, T., Afonso, S., Borges, O., Soeiro,,J., Costa, H., Fonseca, L., Augusta, C., Cruz, M.H., Salazar, M., Matos Soares, F., Sequeira, J., Assunção, A., Correia, P., Lima, M.J. (2008) Variedades de Castanha das Regiões Centro e Norte de Portugal. 80 pp. Ed. INRB.I.P.
Dane, F., Hawkins, L.K., Huang, H. (1999) Genetic variation and population structure of Castanea pumila var. ozarkensis. J. Amer. Soc. Hort. Sci., 124:666–670.
Dane, F., Lang, P., Huang, H., Fu, Y. (2003) Intercontinental genetic divergence of Castanea species in eastern Asia and eastern North America. Heredity, 91:314–321.
Debouzie, D., Heizmann, A., Desouhant, E., Manu, F. (1996) Interference of several temporal and spatial scales between two chestnut insects. Oecologia, 108:151–158.
D’Emerico, S., Bianco, P., Medagli, P., and Schirone, B. (1995) Karyotype analysis in Quercus spp. (Fagaceae). Silvae Genet., 44:66–70.
Desvignes J.C. (1996) L’incompatibilitè du châtaignier induite par le chestnut mosaic virus ChMV. Infos-Ctifl, n° 121.
Diller, J. D. and R. B. Clapper. (1969) Asiatic and hybrid chestnut trees in the eastern United States. J. For., 67:328–331.
Dion, R. (1977) Histoire de la vigne et du vin en France dês origines au XIX siècle. Flammarion, Paris.
Diskin, M., Steiner, K. C., Hebard, F. V. (2006) Recovery of American chestnut characteristics following hybridization and backcross breeding to restore blight-ravaged Castanea dentata. For. Ecol. Manage., 223:439–447.
Elias, T.S. (1971) The genera of Fagaceae in the southeastern United States. J. Arnold Arboretum 52:159–195.
Elorrieta, J. (1949) El castaño en España. Ministerio de Agricultura Pesca y Alimentación No. 48. Madrid, 303 pp.
Ensminger, A. H., Ensminger, M. E., Konlande, J. E., Robson, J. R. K. (1995) The Concise Encyclopedia of Foods and Nutrition. 2nd ed., CRC Press, Boca Raton, FL.
Eriksson, G., Jonson, A., Laureti, M., Pliura, A. (2005) Genetic Variation in Drought Response of Castanea sativa Mill. Seedlings. Acta Hort., 693:247–254.
FAO (2009) http://faostat.fao.org/.
Faridi, N., Nelson, C.D., Banda, H., Abdul Majid, M., Kubisiak, T.L., Hebard, F.V., Sisco, P.H., Phillips, R. (2008) Cytogenetic analysis of a reciprocal translocation in F1 hybrid between American and Chinese chestnuts. Plant & Animal Genomes XVI Conference, January 12–16, 2008, San Diego, CA. W346.
Fernández-López, J., Vázquez-Ruíz-de-Ocenda, R.A., Díaz-Vázquez, R.D., Pereira-Lorenzo, S. (2002) Evaluation of resistance of Castanea sp. clones to Phytophthora sp. using excised chestnut shoots. For. Snow. Landsc. Res., 76:451–454.
Fernando, D.D., Richards, J.L., Kikkert, J.R. (2006) In vitro germination and transient GFP expression of American chestnut (Castanea dentata) pollen. Plant Cell Rep., 25:450–456.
Ferreria-Cardoso, J. V., Fontainhas-Fernandes A.A., Torres-Pereira, M.G. (1993) Nutritive value and technological characteristics of Castanea sativa Mill. fruits - comparative study of some Northeastern Portugal cultivars. In: International Congress on Chestnut, Spoleto. Proceedings. Spoleto, Italy.
Ferrini F., Mattii G.B., Nicese F.P., Pisani P.L. (1992) Investigation for realisation of clonal rootstock for chestnut trees. Proceedings of a scientific meeting SOI 1992, Ravello, Rome, Italy: 412–413.
Ficklin, S., Smith, C., Sisco, P., Wheeler, N., Carlson, J., Sederoff, R., Tomkins, J. (2007) Fagaceae genomic database (FGD): an integrated web-based resource for tree genomics. Plant & Animal Genomes XV Conference, January 13–17, 2007, San Diego, CA. P848.
Fineschi, S., Gillet, E., Malvolti, M.E. (1990a) Genetics of sweet chestnut (Castanea sativa Mill.). Silvae Genet., 39:188–193.
Fineschi, S., Malvolti, M.E., Morgante, M., Vendramin, G.G., Paciucci, M. (1990b) Genetic studies on cultivated chestnut. Abstract, Congreso ISHS, Florencia, Agosto 1990.
Fineschi, S., Malvolti, M.E., Morgante, M., Vendramin, G.G. (1994) Can. J. For. Res., 24:1160–1165.
Fineschi, S., Taurchini, D., Villani, F., Vendramin, G.G. (2000) Chloroplast DNA polymorphism reveals little geographical structure in Castanea sativa Mill.(Fagaceae) throughout southern European countries. Molecular Ecology, 9:1495–1503.
Fu, Y., Dane F. (2003) Allozyme variation in endangered Castanea pumila var. pumila. Annals Bot., 92:223–230.
Gallastegui, C. (1926) Técnica de la hibridación artificial del castaño. Boletín de la Real Sociedad Española de Historia Natural. Tomo XXVI, pp. 88–94.
Gomes-Laranjo, J., Coutinho, J.P., Peixoto, F., Araujo-Alves, J. (2007) Ecologia do castanheiro. In J Gomes-Laranjo, J Ferreira-Cardoso, E Portela, CG Abreu, eds, Castanheiros. UTAD, Vila Real, pp 109–149.
Gomes Guerreiro, M. (1948) Acerca do uso da análise discriminatória: comparação entre duas castas de castanhas. – Sep. Das Publicações da Direcção Geral dos Serviços Florestais e Aquícolas, Vol.XV, Tomo I e II, p.137–151.
Gomes Guerreiro, M. (1957) Castanheiros: alguns estudos sobre a sua ecologia e o seu melhoramento genético. Instituto Superior de Agronomía, Lisboa.
Goulao, L., Valdiviesso, T., Santana, C., Oliveira, C.M. (2001) Comparison between phenetic characterisation using RAPD and ISSR markers and phenotypic data of cultivated chestnut (Castanea sativa Mill.). Gen. Res. Crop Evol., 48:329–338.
Graves, A.H. (1961) Keys to chestnut species. Annu. Rep. North. Nut Grow. Assoc., 61:78–90.
Grente, J., Berthelay-Sauret, S. (1978) Biological control of chestnut blight in France. En: Proc. Am. Chestnut Symp. Morgantown: Virginia University Books pp 30–34.
Hamrick, J.L., Godt, M.J.W. (1989) Allozyme diversity in plants. In: Brown, A.H.D., Clegg, M.T., Kahler, A.L., and Weir, B.S., eds. Plant population genetics, breeding and genetic resources. Sunderland, MA: Sinauer Associates Inc.
Hardin, J.W., Johnson, G.P. (1985) Atlas of foliar surface features in woody plants, VIII. Fagus and Castanea (Fagaceae) of eastern North America. Bull. of the Torrey Bot. Club, 112:11–20.
Hebard, F.V., Stiles, S. (1996) Backcross breeding simplified. Journal of the American Chestnut Foundation, 10:35–39.
Heiniger,U., and Rigling, D. (1994) Biological Control of Chestnut Blight in Europe. Ann. Rev. Phytopathol., 32:581–589.
Huang, H., Dane, F., Norton, J.D. (1994a) Genetic analysis of 11 polymorphic isozyme loci in chestnut species and characterization of chestnut cultivars by multi-locus allozyme genotypes. J. Amer. Soc. Hort. Sci., 119:840–849.
Huang, H., Dane, F., Norton, J.D. (1994b) Allozyme diversity in Chinese, Seguin and American chestnut (Castanea spp.). Theor. Appl. Genet., 88:981–985.
Huang, H., Dane, F., Norton, J.D. (1996) Linkage relationships of isozymes and morphological traits in interspecific chestnut crosses. HortSci., 31:419–420.
Huang, H., Dane, F., Kubisiak, T.L. (1998) Allozyme and RAPD analysis of the genetic diversity and geographic variation in wild populations of the American chestnut (Fagaceae). Amer. J. Bot., 85:1013–1021.
Inoue, E., Ning, L., Hara, H., Ruan, S., Anzai, H. (2009) Development of Simple Sequence Repeat Markers in Chinese Chestnut and Their Characterization in Diverse Chestnut Cultivars. J. Am. Soc. Hort. Sci., 134:610–618.
Janeiro, L.V., Vieitez, A.M., Ballester, A. (1995) Cold storage of in vitro cultures of wild cherry, chestnut and oak. Ann. Sci. For., 52:287–293.
Jaynes, R.A. (1962) Chestnut chromosomes. For. Sci., 8:372–377.
Jaynes, R.A. (1964) Interspecific crosses in the genus Castanea. Silvae Genet., 13:146–154.
Jaynes, R.A. (1972) Genetics of chestnut. USDA Forest Serv. Res. Pap. WO-17, 13p.
Jaynes, R.A. (1975) Chestnuts. Pages 490–503 in: Janick, J and Moore, J.N. (eds.) Advances in Fruit Breeding. Purdue Univ. Press, West Lafayette, IN.
Johnson, G.P. (1988) Revision of Castanea sect. Balanocastanon (Fagaceae). J. Arnold Arboretum, 69:25–49.
Kampfer, S., Lexer, C., Glössi, J., Steinkellner, H. (1998) Characterization of (GA)n microsatellite loci from Quercus robur. Hereditas, 129:183–186.
Krebs, P., Conedera, M., Pradella, M., Torriani, D., Felber, M., Tinner, W. (2004) Quaternary refugia of the sweet chestnut (Castanea sativa Mill.): an extended palynological approach. Veget Hist Archaeobot, 13:145–160.
Kremer, A., Casasoli, M., Barreneche, T. [and others]. 2007. Chapter 5 Fagaceae Trees. In: Genome Mapping and Molecular Breeding in Plants Vol. 7; Forest Trees. (Kole, C. Ed.), Springer Publ, Leipzig, Germany.
Kubisiak, T.L., Roberds, J.H. (2006) Genetic structure of American chestnut populations based on neutral DNA markers. In: Steiner, K.C., and Carlson, J.E., eds. Restoration of American Chestnut to Forest Lands – Proceedings of a Conference and Workshop. May 4–6, 2004, The North Carolina Arboretum, Natural Resources Report NPS/NCR/CUE/NRR – 2006/001, National Park Service, Washington, DC.
Kubisiak, T.L., Hebard, F.V., Nelson, C.D., Zhang, J., Bernatzky, R., Huang, H., Anagnostakis, S.L., Doudrick, R.L. (1997) Molecular mapping of resistance to blight in an interspecific cross in the genus Castanea. Phytopathol., 87:751–759.
Lafitte G (1946) Le Châtaignier Japonais en Pays Basque. Mendionde 69.
Lang, P., Dane, F., Kubisiak, T.L. (2006) Phylogeny of Castanea (Fagaceae) based on chloroplast trnT-L-F sequence data. Tree Gen. and Genom., 2:132–139.
Lang P, Dane F, Kubisiak TL, Huang H. (2007) Molecular evidence for an asian origin and a unique westward migration of species in the genus Castanea via Europe and North America. Mol. Phylogen. Evol., 43:49–59.
Lauteri, M., Villani, F. (2004) Identification of QTLs affecting adaptive traits in Castanea sativa Mill. Plant Cell Environ., 27:1088–1101.
Little, E.L. Jr. (1979) Checklist of United States trees (native and naturalized). US Dept. of Agric. Handbook No. 541, Washington, DC. 375 p.
Liu, L.; Zhou, J.Y. (1999) Some considerations on chestnut development in the 21st century in China. Acta Hort., 494:85–88.
Malvotti, M.E., Fineschi, S. (1987) Analysis of enzyme systems in chestnut (Castanea sativa Mill.). Genet. Agr., 41:243–256.
Manos, P.S., Zhe-Kun Zhou, Z-K., Cannon, C.H. (2001) Systematics of Fagaceae: phylogenetic tests of reproductive trait evolution. Int. J. Plant Sci., 162:1361–1379.
Marinoni, D., Akkak, A., Bounous, G., Edwards, K.J., Botta, R. (2003) Development and characterization of microsatellite markers in Castanea sativa (Mill). Mol. Breed., 11:127–136.
Martín, M.A.., Mattioni, C., Cherubini, M., Taurchini, D., Villani, F. (2010) Genetic characterisation of traditional chestnut varieties in Italy using microsatellites (simple sequence repeats) markers. Annals Applied Biol., 157:37–44.
Martins, A., Linhares, I., Raimundo, F., Coutinho, J.P., Gomes-Laranjo, J., Borges, O., Sousa, V. (2005) The importance of deep soil layers to supply water to agro-forestry systems: A case study of a mature chestnut orchard in Northern Portugal. Acta Hort., 693:663–670.
Maynard, C.A., Powell, W.A., Polin-McGuigan, L.D., Viéitez, A.M., Ballester, A., Corredoira, E., Merkle, S.A. and Andrade, G.M. (2008) Chestnut. In: Kole, C. and Hall, T.C. (eds.) A Compendium of Transgenic Crop Plants: Transgenic Forest Trees Species. Volume 9. Blackwell Publishing, Oxford, UK, pp. 169–192.
McCarthy, M. A., Meredith, F.I. (1988) Nutrient data on chestnuts consumed in the United States. Econ. Bot., 42:29–36.
Mehra, P.N., Hans, A.S., Sareen, T.S. (1972) Cytomorphology of Himalayan Fagaceae. Silvae Genet., 21:102–109.
Miranda-Fontaíña, M.E., Fernández-López, J. (1992) Micropropagation as nursery technique in chestnut compared with stooling. Proceedings of the World Chestnut Industry Conference, July 8–10, 1992. Morgantown, West Virginia, USA, 54–60.
Nienstaedt, H. (1948) Notes on the Chestnut: Breeding, Culture, and Botanical Characters of Species and Hybrids. Master’s Thesis, Yale School of Forestry, 104 p + XIII.
Nienstaedt, H. (1956) Receptivity of the pistillate flowers and pollen germination test in genus Castanea. Zeitschrift Forstgenetik Forstplanzenz, 5:40–45.
Ohri, D., Ahuja, M.R. (1991) Giesma C-banding in Fagus sylvatica L., Betula pendula Roth and Populus tremula L. Silvae Genet., 40:72–75.
Oraguzie, N.C., McNeil, D.L., Paterson, A.M., Chapman, H.M. (1998) Comparison of RAPD and MorphoNut markers for revealing genetic relationships between chestnut species (Castanea spp) and New Zealand chestnut selections. N.Z. J. Crop & Hort. Sci., 26:109–15.
Pastorelli, R., Smulders, M.J., Van’t Westende, W.P.C, Vosman, B, Gianinini, R, Vettori, C, Vendramin G.G. (2003) Characterization of microsatellites markers in Fagus sylvatica L. and Fagus orientalis Lipsky Mol. Ecol. Notes, 3:76–78.
Pereira, M.J.P., Castro, L.F.T., Torres-Pereira, J.M.G., Pereira-Lorenzo, S. (1999) Isozyme polymorphisms in portuguese chestnut cultivars. Acta Hort., 494:283–286.
Pereira-Lorenzo, S., Fernández-López, J. (1997) Propagation of chestnut cultivars by grafting: methods, rootstocks and plant quality. J. Hort. Sci., 72:731–739.
Pereira-Lorenzo, S., Fernandez-Lopez, J. (2001) El castaño. In: La Horticultura Española (F. Nuez y G. Llácer, eds.), Madrid, 280–284.
Pereira-Lorenzo, S., Ramos-Cabrer, A.M. (2004) Chestnut, and ancient crop with future. Production Practices and Quality Assessment of Food Crops, Volume 1, Preharvest Practice (Ramdane Dris and S. Mohan Jain ed.), Kluwer Academic Publishers, 1:105–161. ISBN: 1-4020-1698-0.
Pereira-Lorenzo, S., Fernández-López, J., Moreno-González, J. (1996a) Variability and grouping of Northwestern Spanish Chestnut Cultivars (Castanea sativa). I. Morphological traits. J. Amer. Soc. Hort. Sci., 121:183–189.
Pereira-Lorenzo, S., Fernández-López, J., Moreno-González, J. (1996b) Variability and grouping of Northwestern Spanish Chestnut Cultivars (Castanea sativa). II. Isoenzyme traits. J. Amer.Soc. Hort. Sci., 121:190–197.
Pereira-Lorenzo, S.; Ascasibar, J., Ramos, A. (1999) Spanish scheme in chestnut improvement for nut and timber production. FAO-CIHEAM-Nucis- Newsletter, 8:32–34.
Pereira-Lorenzo, S., Díaz-Hernández, B., Ciordia-Ara, M., Ascasibar-Errasti, J., Ramos-Cabrer, A.M., Sau, F. (2001a) Spanish chestnut cultivars. HortSci., 36:344–347.
Pereira-Lorenzo, S., Ramos-Cabrer, A.M., Rios-Mesa, D., Perdomo, A., González-Pérez, J. (2001b) Update of the Spanish Chestnut Inventory of Cultivars. FAO-CIHEAM-Nucis-Newsletter, 10:34–37.
Pereira-Lorenzo, S., Rios, D., Cubas, F., Calzadilla, C., Ramos-Cabrer, A.M., Sánchez-Sánchez, A. (2001c) Chestnut cultivars on the Canary Islands. Forest Snow Lands. Res., 76:445–450.
Pereira-Lorenzo, S., Díaz-Hernández, M.B., Ramos-Cabrer, A.M. (2006a) Use of highly discriminating morphological characters and isoenzymes in the study of Spanish chestnut cultivars. J. Amer. Soc. Hort. Sci., 131:770–779.
Pereira-Lorenzo, S., Ramos-Cabrer, A.M., Díaz-Hernández, M.B., Ciordia-Ara, M., Rios-Mesa, D. (2006b) Chemical composition of chestnut cultivars from Spain. Sci. Hort., 107:306–314.
Pereira-Lorenzo, S., Ríos-Mesa, D., González-Díaz, A.J., Ramos-Cabrer, A.M. (2007) Los castañeros de Canarias. CCBAT - Cabildo de Tenerife; CAP - Cabildo de La Palma, 1–136.
Pereira-Lorenzo, S., Costa, R., Ramos-Cabrer, A.M., Ribeiro, C., Serra da Silva, C., Manzano, G., Barreneche, T. (2010). Variation in grafted European chestnut and hybrids by microsatellites reveals two main origins in the Iberian Peninsula. Tree Gen. Genom., 6:701–715.
Pereira-Lorenzo, S., Lourenço Costa, R.M., Ramos-Cabrer, A.M., Ciordia-Ara, M., Marqués Ribeiro, C.A., Borges, O., Barreneche, T. (2011). Chestnut cultivar diversification process in the Iberian Peninsula, Canary Islands and Azores. Genome, 54:301–315. Doi:10.1139/G10-122.
Pigliucci, M., Benedettelli, S., Villani, F. (1990a) Spatial patterns of genetic variability in Italian chestnut (Castanea sativa). J. Can. Bot., 68:1962–1967.
Pigliucci, M., Villani, F., Benedettelli, S. (1990b) Geographic and climatic factors associated with the spatial structure of gene frequencies in Castanea sativa Mill. from Turkey. J. Genet., 69: 141–149.
Pitte, J.R. (1985) Le châtaignier en Gaule et dans les provinces voisines. En: Le Bois dans la Gaule romaine et les provinces voisines, ERRANCE, 21:185–190.
Pitte, J.R. (1986) Terres de Castanide. Fayard, 479 pp.
Pliura, A., Eriksson, G. (2002) Genetic Variation in Juvenile Height and Biomasa of Open-pollinated Familias of six Castanea sativa Mill. Populations in a 2 x 2 Factorial Temperature x Watering Experiment. Silvae Gent., 51:152–160.
Polin, L.D., Liang, H., Rothrock, R.E., Nishii, M., Diehl, D.L., Newhouse, A.E., Nairn, C.J., Powell, W.A., Maynard, C.A. (2006) Agrobacterium-mediated transformation of American chestnut (Castanea dentata (Marsh.) Borkh.) somatic embryos. Plant Cell Tissue Organ Cult., 84:69–79.
Ramos-Cabrer, A.M., Pereira-Lorenzo, S. (2005) Genetic Relationship between Castanea sativa Mill. Trees from North-western to South Spain Based on Morphological Traits and Isoenzymes. Gen. Res. Crop Evol., 52:879–892.
Ramos-Cabrer, A.M., Díaz-Hernández, M.B., Ciordia-Ara, M., Rios-Mesa, D., Gonzalez-Díaz, J., Pereira-Lorenzo, S. (2006) Study of Spanish chestnut cultivars using SSR markers. Adv. Hort. Sci., 20:113–116.
Robichaud, R.L., Lessard, V.C., Merkle, S.A. (2004) Treatments affecting maturation and germination of American chestnut somatic embryos. J. Plant Physiol., 161:957–969.
Robin, C., Heiniger, U. (2002) Chestnut blight in Europe: diversity of Chryphonectria parasitica, hypovirulence and biocontrol. For. Snow Landsc. Res., 76:361–367.
Rodríguez, L., Cuenca, B., Pato, B., Cámara, M.J., Ocaña, L. (2005) Cost and efficiency of propagating Castanea sativa hybrids in vitro and by cutting in a commercial nursery. Acta Hort., 693:305–311.
Rothrock, R.E., Polin-Mcguigan, L.D., Newhouse, A.E., Powell, W.A., Maynard, C.A. (2007) Plate flooding as an alternative Agrobacterium-mediated transformation method for American chestnut somatic embryos. Plant Cell, Tissue Organ Cult., 88:93–99.
Sánchez, M.C., San-José, M.C., Ferro, E., Ballester, A., Vieitez, A.M. (1997a) Improving micropropagation conditions for adult-phase shoots of chestnut. J. Hortic. Sci., 72:433–443.
Sánchez, M.C., Ballester, A. and Vieitez, A.M. (1997b) Reinvigoration treatments for the micropropagation of mature chestnut trees. Ann. Sci. For., 54:359–370.
Santamour F.S. Jr., McArdle A.J., Jaynes R.A. (1986) Cambial isoperoxidase patterns in Castanea. J. Environ. Hort., 4:14–16.
Sartor, C., Mellano, M.G., Quacchia, A., Alma, A., Botta, R. (2007) Cinipide galligeno del castagno: prospettive di impiego di strategie da affiancare alla lotta biologica. Riassunto dei lavori, VIII giornate scientifiche SOI. Italus Hortus, 14:130–133.
Sawano, M., Ichii, T., Nakanishi, T., Kotera, Z. (1984) Studies on identification of chestnut species and varieties by isozyme analysis. Science Reports of Faculty of Agriculture, Kobe University, 16:67–71.
Schad, C., Solignat, G., Grente, J., Venot, P. (1952) Recherches sur le châtaignier à la Station de Brive. Ann. Amel. Plantes, 3:376–458.
Seabra, R.C., Pais, M.S. (1998) Genetic transformation of European chestnut. Plant Cell Rep., 17:177–182.
Seabra, R.C., Pais, M.S. (1999) Genetic transformation of European chestnut (Castanea sativa Mill.) with genes of interest. Acta Hort., 494:407–413.
Sebastiana, M. (2006) Identificação de genes envolvidos no reconhecimento hóspede/hospedeiro em ectomicorrizas. Ph.D. Dissertation.127 pp.
Sederoff, R., Nielson, D., Smith, C., Tomkins, J., Atkins, M., Blackmon, B., Staton, M., Ficklin, S., Hebard, F., Sisco, P., Carlson, J., Diloreto, S., Barakat, A., Powell, W., Baier, K., Anagnostakis, S., Kubisiak, T., Wheeler, N. (2008) Genomic tool development for the Fagaceae. Plant & Animal Genomes XVI Conference, January 12–16, 2008, San Diego, CA. p494.
Serrazina, S. (2004) Isolamento e Caracterização de Genes de Resistência à Doença da Tinta em Castanea sativa Mill. Ph.D. Dissertation. 301 pp.
Sisco, P.H., Kubisiak, T.L., Casasoli, M., Barreneche, T., Kremer, A., Clark, C., Sederoff, R.R., Hebard, F.V., Villani, F. (2005) An improved genetic map for Castanea mollissima/Castanea dentata and its relationship to the genetic map of Castanea sativa. Acta. Hort., 693:491–495.
Soylu A. (1990) Heredity of male sterility in some chestnut cultivars (Castanea sativa Mill.) XXIII Int. Congr. Florence.
Soylu, A., Serdar, U. (2000) Rootstock selection on chestnut (Castanea sativa Mill.) in the middle of black sea region in Turkey. Acta Hort., 538:483–487.
Steinkellner H., Fluch S., Turetschek E., Lexer C., Streiff R., Kremer A., Burg K., Glössl J. (1997) Identification and characterization of (GA/CT)n -microsatellite loci from Quercus petraea. Plant Mol. Biol., 33:1093–1096.
Stilwell, Kevin L., Wilbur, Henry M., Werth, Charles R., Taylor, Douglas R. (2003) Heterozygote advantage in the American chestnut, Castanea dentata (Fagaceae) Am. J. Bot., 90:207–213.
Tanaka, K., Tsumura, Y., Nakamura, T. (1999) Development and polymorphism of microsatellite markers for Fagus crenata and the closely related species, F. japonica. Theor. Appl. Genet., 99:11–15.
Tanaka, T, Yamamoto, T, Suzuki, M. (2005) Genetic Diversity of Castanea crenata in Northern Japan Assessed by SSR Markers. Breed. Sci., 55:271–277.
Taylor, W. A. and H. P. Gould. (1914) Promising new fruits. pp 122–124 IN: Yearbook of Agriculture for 1913. USDA, Washington, D.C.
UPOV (1988) Draft guidelines for the conduct of tests for distinctness, homogeneity and stability (CHESNUT). TG/124/1(proj.), 23 p.
Urquijo, P. (1944) Aspectos de la obtención de híbridos resistentes a la enfermedad del castaño. Bol. Veg. Ent. Agr. XIII: 447–462.
Urquijo, P. (1957) La regeneración del castaño. Bol. de Pat. Veg. y Entomología Agrícola, XXII:217–232.
Valdiviesso, M.T. (1999) Estudo sobre a reproduçao sexuada e caracterizaçao de cultivares de Castanea sativa Mill. Thesis Doctoral, Alcobaça.
Van Fleet, W. (1920) Chestnut work at Bell Experiment Plot. 11th annual report of the Northern Nut Growers Association, pp 16–21.
Vidal, N., Sánchez, C., Jorquera, L., Ballester, A., Vieitez, A.M. (2005) Cryopreservation of chestnut by vitrification of in vitro-grown shoot tips. In Vitro Cell. Dev. Biol.-Plant, 41:63–68.
Vieira Natividade, J. (1947) Quatro anos na defesa da campanha e Reconstituição dos Soutos. Edição da Junta Nacional das frutas. Lisboa.
Vieitez, E. (1974) Vegetative propagation of chestnut. N.Z. For. Sci., 4:242–252.
Vieitez, F.J., Merkle, S.A. (2004) Fagaceae. In: Litz RE (ed) Biotechnology of fruit and nut crops. CAB International, Wallinford, pp 263–296.
Vieitez, A.M., Sánchez, M.C., García-Nimo, M.L., Ballester, A. (2007) Protocol for micropropagation of Castanea sativa. In: Protocols for Micropropagation of Woody Trees and Fruits. AM Jain and H Häggman, eds. Springer, pp 299–312.
Villani, F.S., Pigliucci, M., Benedettelli, S., Cherubini, M. (1991a) Genetic differentiation among Turkish chestnut (Castanea sativa Mill.) populations. Heredity, 66:131–136.
Villani, F.S., Benedettelli, S., Paciucci, M., Cherubini, M., and Pigliucci, M. (1991b) Genetic variation and differentiation between natural populations of chestnut (Castanea sativa Mill.) from Italy. In: Fineschi, S., Malvolti, M.E., Cannata, F., Hattemer, H.H., eds. Biochemical markers in the population genetics of forest trees. The Netherlands: SPB Academic Publishing BV Press.
Villani, F.; Pigliucci, M.; Cherubini, M.; Sun, O.; Parducci, L. (1993) Genetic diversity of Castanea sativa Mill. in Europe: Theorical aspects and applied perspectives. Abstracts, International Congress on Chestnut, Spoleto, Italia.
Villani, F., Sansotta, A., Cherubini, M., Cesaroni, D., Sbordoni, V. (1999) Genetic structure of Castanea sativa in Turkey: evidence of a hybrid zone. J. Evol. Biol., 12:233–244.
Vossen, P. (2000) Chestnut culture in California. University of California, Division of Agriculture and Natural Resources, Publication 8010, 17 p.
Wen, H., Norton, J. (1992) Enzyme Variation in Chinese Chestnut Cultivars. Abstract, International Chestnut Conference, Morgantown, USA.
Yamamoto, T., Tanaka, T., Kotobuki, K., Matsuta, N., Suzuki, M., Hayashi, T. (2003) Characterization of simple sequence repeats in Japanese chestnut. J. Hort. Sci. Biotech., 78: 197–203.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2012 Springer Science+Business Media, LLC
About this chapter
Cite this chapter
Pereira-Lorenzo, S. et al. (2012). Chestnut. In: Badenes, M., Byrne, D. (eds) Fruit Breeding. Handbook of Plant Breeding, vol 8. Springer, Boston, MA. https://doi.org/10.1007/978-1-4419-0763-9_19
Download citation
DOI: https://doi.org/10.1007/978-1-4419-0763-9_19
Published:
Publisher Name: Springer, Boston, MA
Print ISBN: 978-1-4419-0762-2
Online ISBN: 978-1-4419-0763-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)