
Abstract
The chestnuts and chinquapins are a group of about seven species of trees and shrubs in the genus Castanea. They are of considerable importance ecologically in all the areas of their natural occurrence, and the chestnuts especially are of great economic value for their lumber and for their nut crop wherever they are cultivated. Two catastrophic diseases, chestnut blight caused by the ascomycete fungus Cryphonectria parasitica (Murr.) Barr and Phytophthora root rot (ink disease) caused primarily by the soil-borne oomycetes Phytophthora cinnamomi Rands and P. cambivora (Petri) Buisman, have severely impacted chestnut in Europe and North America. Therefore, much of the breeding work continues to focus on breeding for resistance to these two diseases. The most serious insect pest of Castanea is the Asian chestnut gall wasp, Dryocosmus kuriphilus Yasumatsu. Variation in host tolerance to D. kuriphilus has led to development of new gall-resistant chestnut cultivars. Interspecific hybridization offers great opportunity to combine the most favorable traits found in the ample genetic diversity of the genus through introgression into locally-adapted populations. Chestnut breeders in eastern Asia have made great strides towards improvement of chestnut fruit quality and crop yields, and researchers in all chestnut growing regions have made gains in disease resistance by using molecular markers and other genomic tools to assist selection. Biotechnologies that include transmissible hypovirulence as a biocontrol for chestnut blight, tissue culture and other micropropagation techniques, and genetic engineering and transformation technologies are complementary to classical plant breeding programs.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Castanea
- Chestnut blight
- Cryphonectria parasitica
- Germplasm conservation
- Ink disease
- Phytophthora root rot
- Phytophthora cinnamomi
1 Introduction
The chestnuts and chinquapins are a group of tree and shrub species in the genus Castanea Miller of the Fagaceae. Depending on taxonomic treatment, 7–13 lower taxa are recognized within the genus (Mellano et al. 2012; Nixon 1997; Pereira-Lorenzo et al. 2016; Worthen et al. 2010). Chestnut researchers commonly recognize the following lower taxa: C. crenata Siebold & Zucc. (Japanese chestnut), C. mollissima Blume (Chinese chestnut), C. henryi Rehder & E.H. Wilson (willow leaf or pearl chestnut), and C. seguinii Dode from eastern Asia; C. sativa Mill. (European or sweet chestnut) from Europe and western Asia; C. dentata Borkh . (American chestnut), C. pumila (L.) Mill. var. pumila (Allegheny chinquapin), and C. pumila (L.) Mill. var. ozarkensis (Ashe) G.E. Tucker (Ozark chinquapin) from eastern North America (Conedera et al. 2004a; Johnson 1988; Pereira-Lorenzo et al. 2016; Pridnya et al. 1996; Worthen et al. 2010). In English-speaking countries, the common name chinquapin is often used in reference to Castanea species that typically produce one pistillate flower/nut per cupule, whereas the common name chestnut is applied to species that typically produce three pistillate flowers/nuts per cupule. The term chinquapin is also applied to other Fagaceae trees in Quercus, Chrysolepis and Castanopsis; chinquapin is derived from chechinquamins—John Smith’s seventeenth century transliteration of the word used by some indigenous peoples of Virginia to refer to C. pumila var. pumila (Barbour 1986). The native and introduced Castanea species found growing throughout the world today are listed in Table 5.1, and their areas of natural occurrence are mapped in Fig. 5.1.


Native and naturalized distributions of Castanea species worldwide. (a) Distributions of C. crenata , C. mollissima , C. henryi and C. seguinii in east Asia. Note: distributions of C. mollissima, Fig. 5.1 (continued) C. seguinii and C. henryi are largely sympatric in eastern China. (b) Distribution of C. sativa in Europe. (c) Distributions of C. dentata , C. pumila var. pumila and C. pumila var. ozarkensis in eastern North America. (Source: Maps created by Erin Taylor using the distribution shapefiles of Fei et al. (2012) for Asian and European species and Little (1977) for North American species. Distribution of C. crenata in the Korean peninsula digitized by Erin Taylor using occurrence data from Barstow (2018))
Castanea species are important nut crop and timber producers throughout most of their native and introduced ranges, and have been for many thousands of years (Avanzato and Bounous 2009). The major exception being in eastern North America, where introduced pests and pathogens severely limit cultivation of the native species. Nevertheless, the North American Castanea species were ecologically, economically and culturally important in eastern North America prior to the introduction of chestnut blight and Phytophthora root rot disease (ink disease) (Anagnostakis 2001; Crandall et al. 1945; Roane et al. 1986). As a result, C. dentata and C. pumila are the focus of extensive research and restoration breeding programs (Jacobs et al. 2013; Steiner et al. 2017; Thomas et al. 2015; Westbrook et al. 2019; Worthen et al. 2010).
Although there is still much debate about the history of artificial selection and domestication of Castanea species around the globe, evidence for selection of chestnut in East Asia and Western Europe dates back many thousands of years (Avanzato and Bounous 2009; Conedera et al. 2004b). In every case, our human ancestors were able to utilize the abundant annual production of fruits from wild chestnut trees, and to alter their environments (using fire, for example) in ways that benefitted both the trees and themselves (Avanzato and Bounous 2009; Krebs et al. 2004). LaBonte et al. (2018) provide genomic evidence for the ancient domestication of C. mollissima in China. From Pleistocene refugia in the Caucasus, parts of Italy and Switzerland, and probably also the Iberian Peninsula, C. sativa expanded rapidly to cover an area from the Caspian Sea to the Atlantic Ocean (Conedera et al. 2004; Krebs et al. 2004). Squatriti (2013) reviews evidence that improved cultivars selected from naturally occurring populations of C. sativa were being propagated by grafting in tenth century Italy. Pereira-Lorenzo et al. (2018) used microsatellite markers to investigate the role of graft propagation of local ecotypes in the domestication of C. sativa. They documented also the great age of some surviving grafts; the two oldest living trees they studied in Spain were grafted in the years 1427 and 1479. In Italy two ancient grafts of cv. Marrone Fiorentino were dated to 1650 and 1700. In contrast, while it is well established that the North American Castanea species were widely used by ancient peoples, convincing evidence has not been produced to support a history of artificial selection or domestication for Castanea in North America (but see Abrams and Nowacki 2008). Day (1953) describes early archeological evidence of C. dentata culture in New England, but no real documentation for selection or improvement prior to the nineteenth century.
Two major diseases—chestnut blight and Phytophthora root rot—have impacted chestnut growing globally, and most severely in Europe and North America. The chestnut blight pandemic began in Bronx, New York in 1904, caused by the ascomycete fungus Cryphonectria parasitica (Murr.) Barr, and within 50 years of its discovery had almost completely destroyed the American chestnut. Chestnut blight subsequently caused grave damage to the European chestnut throughout its native range (Anagnostakis 1987; Robin and Heiniger 2001). Phytophthora root rot—called ink disease in Europe—is caused primarily by the soil-borne oomycetes Phytophthora cinnamomi Rands and P. cambivora (Petri) Buisman, and has been a major limiting factor in chestnut culture in Europe and North America for at least 200 years (Crandall et al. 1945; Vettraino et al. 2005).
The most injurious insect pest of Castanea is the Asian chestnut gall wasp, Dryocosmus kuriphilus Yasumatsu. The gall former oviposits in buds during mid-summer, and the larvae deform shoots and leaves at budbreak the following spring, cause drastic crop losses and, in severe cases, death of the tree. The insect is native to China but was accidentally introduced to Korea and Japan, and then into North America and Europe. Variation in host tolerance to D. kuriphilus has led to development of new gall-resistant chestnut cultivars.
In this chapter, we review historical developments and recent advances in chestnut breeding for chestnuts as a nut crop, and for restoration of the American chestnut as a functioning member of Appalachian forest communities. We note progress and improvements in nut quality, crop yields and, most importantly, in achieving host tolerance to chestnut blight, Phytophthora root rot, and Asian chestnut gall wasp. We also identify important unanswered questions and areas of inquiry that are likely to benefit chestnut breeders in the future.
2 Cultivation and Traditional Breeding
Chestnut trees have been cultivated for thousands of years for their sweet, edible nuts, and were valued for their wood and other products by traditional cultures in Europe, Asia and eastern North America (Avanzato and Bounous 2009; Davis 2003). Migrating peoples have carried chestnuts well beyond their areas of natural occurrence, as far as Chile in South America (Grau 2003), southeastern Australia (Casey and Casey 2009), New Zealand (Klinac and Knowles 2009) and the West Coast of North America (Craddock 2009). Chestnuts dried and ground into flour were once a staple food in Mediterranean kitchens, and especially so in the mountainous regions of Turkey, Portugal, Spain, France and Italy, and on the island of Corsica where chestnut culture became a veritable chestnut civilization (Cherubini 1981; Pereira-Lorenzo et al. 2009a, b; Smith 1950; Soylu et al. 2009; Squatriti 2013). Almost every material need was provided for by the chestnut tree, not just food but also timber, poles, wicker, tannin and fuel from the chestnut coppice, bedding for animals and even medicinal astringents from the leaves (Bourgeois 1992; Bruneton-Governatori 1984; Pitte 1986). Along with the grapevine and the olive tree, the European chestnut is one of the great pillars upon which Mediterranean civilization was built (Bellini and Nin 2009; Bignami and Salsotto 1983). The Romans transported chestnut trees to all parts of their empire and chestnut culture later underwent a tremendous expansion during the late classical and early medieval period (Cherubini 1981; Conedera et al. 2004a, b; Jarman et al. 2019, Squatriti 2013). Chinese and Japanese chestnut trees played similar and no less important roles in the civilizations of China, Korea and Japan (Qin and Feng 2009; Saito 2009). A storehouse full of dried chestnuts represented food security – and wealth. Chestnuts were insurance against drought, flood, siege and any of the many other calamities that could befall an agriculture based on annual grain crops (Bruneton-Governatori 1984; Pitte 1986; Smith 1950; Squatriti 2013). Chestnut trees come very close to representing the archetypal tree crop (Smith 1950; Vieitez et al. 1996). They are very long lived; examples of Castanea sativa in Europe are known to be more than 1000 years old (Bounous 2002; Vieitez et al. 1996). And even at that great age, they continue to provide annual harvests of delicious, nutritious, easily gathered, easily stored food. Because a mature chestnut orchard resembles a temperate hardwood forest, in both structure and function, it provides significant ecological benefits with consequences far beyond merely producing a fruit (Vieitez et al. 1996). The tree offers habitat to myriad animal species and acts as the foundation of its community. It anchors soils to steep slopes that would otherwise be almost completely unsuitable for other forms of agriculture (Smith 1950). The chestnut grove moderates the local climate by shading the understory, and transpiring water into the atmosphere. It serves as host for symbiotic relationships with fungi, including the choice edible Cantharellus cibarius and Boletus edulis mushrooms whose harvest provides a highly valued forest product to the land’s stewards (Vieitez et al. 1996). These mutualistic symbionts join the ranks of other chestnut forest fungi involved in decomposition and recycling, mobilizing deep soil nutrients and minerals into the humus layers. The landscape value of the chestnut grove is almost unequalled by that of any other forest type (Bounous and DeGuarda 2002; Vieitez et al. 1996) possibly because the grove is evocative of a simpler, quieter past, when our relationship with nature was closer to the regular cycles of sun and moon and season (Fig. 5.2).

The mature chestnut orchard provides many of the same ecosystem services as the temperate hardwood forest. When the trees are widely spaced and pruned up, a rich grassland community can develop in the understory. (Photo by J.H. Craddock)
The result of centuries of cultivation and selection by growers is thousands of named varieties of Castanea crenata , C. mollissima , C. sativa , and their hybrids (Anagnostakis 2012; Miller et al. 1996; Pereira-Lorenzo et al. 2012; Rutter et al. 1991; Worthen et al. 2010). Dr. Sandra Anagnostakis, at the Connecticut Agricultural Experiment Station, maintains and updates the registry of chestnut cultivar names on her CAES web page: https://portal.ct.gov/CAES/ABOUT-CAES/Staff-Biographies/Sandra-L-Anagnostakis (Anagnostakis 2019).
The history of chestnut breeding was recently reviewed by Pereira-Lorenzo et al. (2016) in their very thorough paper on interspecific hybridization . They discuss germplasm and genetic resources, and genetic diversity and mating systems for the major Castanea species, as well as the development of molecular markers and genome-wide approaches to improve selection for desired traits in hybrid progenies. There is a greater focus on cultivar and rootstock development in the review by Pereira-Lorenzo et al. (2012). A history of chestnut breeding in the United States is well illustrated in the review by Anagnostakis (2012) of work begun by Walter Van Fleet in 1894 (Van Fleet 1914). Worthen et al. (2010) provide an excellent historical and theoretical context for breeding and selecting Castanea dentata for resistance to Cryphonectria parasitica using modern population genetic and genomic tools. Recently, Westbrook et al. (2019) provided genomic and quantitative genetic evidence that phenotypic selection can be used to move alleles conferring disease resistance from Asian Castanea species into an American chestnut genetic background (Figs. 5.3, 5.4 and 5.5).

Canker severity on American chestnut hybrids, following inoculation with Castanea parasitica strain EP155 (virus-free) and two isogenic hypovirulent strains (CHV1-EP713 and CHV1-Euro7). Host response varies from susceptible to resistant. The susceptible phenotypes are shown on the left. The resistant phenotypes are on the right. The top row are half-sibling B2F2s (2nd backcross F2 hybrids). The bottom row are full-sibling B3s (3rd backcross F1 hybrids). (Photos by J.H. Craddock)

Roguing susceptible progeny. In backcross breeding for Castanea parasitica resistance, American chestnut backcross hybrid progeny are inoculated in their 5th year, and selections are made from years 6 to 10. This selected third backcross tree had adequate blight tolerance, and the tree form and leaf and twig morphology of its C. dentata ancestors. (Photo by J.H. Craddock)

Open pollination and controlled pollination can be used to produce seed from selected hybrids. In this illustration, hand pollinations are being used to intercross selected 3rd backcrosses to produce 3rd backcross F2 (B3F2) seed for Tennessee seed orchards. (Photo by J.H. Craddock)
World chestnut production today is increasing. Global chestnut production (nuts and lumber products) declined precipitously in the late nineteenth and early twentieth centuries from a conspiracy of circumstances that included two world wars, two catastrophic diseases (chestnut blight, ink disease), the movement of invasive pests (such as the Asian chestnut gall wasp Dryocosmus kuriphilus) , changing patterns of settlement and employment (increased urbanization, mechanization of farm labor) and changes in the world’s economy and diet (Avanzato and Bounous 2009). But, the situation for chestnut is now improving after reaching a global nadir in the 1970s (Conedera et al. 2004b). Evidence of renewed interest since the 1990s is the number and frequency of international scientific meetings on chestnut worldwide (see the list of Acta on the ISHS web page). The availability of new, improved cultivars means that growers can now choose varieties resistant to the major pests and diseases that have plagued chestnut for much of the past century.
3 Germplasm Characterization and Conservation
The difficulty of conservation of Castanea genetic resources is compounded by chestnut blight, Phytophthora root rot, insect pests and habitat changes. Wild Castanea resources in situ are still plentiful but in every case there is special concern for genetic erosion. The Caucasus Biosphere Reserve in the (Republic of) Georgia harbors naturally occurring populations of C. sativa in an area thought to be close to the species’ center of diversity. Similarly, the Great Smoky Mountains National Park in Tennessee and North Carolina represents a reservoir of diversity close to the area of greatest abundance for C. dentata . However, C. parasitica and Phytophthora cinnamomi and P. cambivora , and Dryocosmus kuriphilus are perennial threats to that germplasm. In situ conservation depends therefore not only on the political integrity of the Reserve and the Park, but also on the phytopathological conditions present at those locations. There is no such system of parks or reserves in place to conserve germplasm of C. pumila var. pumila or C. pumila var. ozarkensis , so conservation is largely the responsibility of engaged citizens and private, non-governmental organizations like The American Chestnut Foundation. Ex situ conservation is also fraught with difficulties. The same diseases and pests conspire to destroy collections of susceptible germplasm, whether it is grown on its own roots or grafted onto Phytophthora-resistant rootstocks. The costs and challenges of maintaining collections of grafted plants is exacerbated by the phenomenon of delayed graft failure, which is particularly problematic for some C. mollissima genotypes and for interspecific graft combinations. A list of institutions that currently maintain and/or have access to Castanea germplasm resources is listed in Appendix I.
3.1 Genetic Variation
Germplasm characterization and conservation in the genus Castanea has been an active part of chestnut breeding in Europe, Asia and North America (Alexander et al. 2005; Bolvansky and Tarinova 2009). Government and university research programs in each of these regions are currently involved in germplasm conservation (Bounous 2002; Mellano et al. 2018). An important tool in germplasm conservation work has been the use of molecular methods to decipher genetic relationships among cultivars, and within and among species. And because the wild species are an important source of variation for breeding programs, many research efforts have focused on elucidating genetic variation in natural populations (reviewed by Pereira-Lorenzo et al. 2012, 2016; Worthen et al. 2010).
Values reported for within genus estimates of diversity allow us to rank the species: Castanea mollissima is apparently the most highly diverse, and this is maybe not surprising considering its enormous geographic range and the supposed great age of its populations, followed closely by C. pumila sensu lato and C. pumila var. ozarkensis . An intermediate level of diversity is exhibited by populations of C. crenata (Nishio et al. 2011b) and C. sativa (Mattioni et al. 2013). The structure of this diversity within C. sativa varies strongly with geography and is indicative of gene flow from east to west (Mattioni et al. 2013). Within species, diversity for C. sativa is generally much higher in the eastern (older) populations in the Caucasus, and generally much lower in the western (younger) populations, particularly in those most strongly influenced by clonal propagation of cultivars such as found in Italy. Castanea dentata is apparently lowest in species-level diversity (Huang et al. 1994; Kubisiak and Roberds 2006). However, the values reported by Huang et al. (1994) and Kubisiak and Roberds (2006) may not accurately represent the variation present in the southernmost regions of its range due to the low number of samples they collected. Chloroplast haplotype diversity in southerly populations of C. dentata are remarkably high, and chloroplast haplotype sharing has been documented between C. dentata and C. pumila (Shaw et al. 2012).
3.2 Japan
Castanea crenata is native to Japan (Fig. 5.1) and has been cultivated there for thousands of years. Descriptions of the oldest named cv. Chokoji appear in the literature in the late 1500s (Saito 2009). Cultivar development probably began with selection of wild chestnuts (Kotobuki 1994), but accelerated with urgency in the 1940s after the accidental introduction of gall wasp Dryocosmus kuriphilus from China. By the 1950s the gall-wasp resistant cvs. Tanzawa, Tsukuba and Ishizuchi were released. Ginyose, an older variety, was also recognized as resistant. Eventually the gall former adapted to the widely planted new varieties by overcoming their resistance. But, because of effective biocontrol of galls by the parasitoid Torymus sinensis, these cultivars are still grown today. Despite their adaptation to the soils and climate of Japan, and their high yields of very large chestnuts (the largest fruit of any chestnut species), these cultivars unfortunately suffer from a trait that was thought to characterize all forms of C. crenata – the nuts themselves are very difficult to peel and they have a thick, adherent, astringent and unpalatable pellicle. Hybridization efforts with the easy to peel C. mollissima from China did produce some cultivars – Riheiguri is one example – but for the most part the Chinese-Japanese hybrids were either not well adapted to the Japanese growing conditions or the nuts were of inferior quality. New screening techniques for rapid evaluation of pellicle peelability (Shoda et al. 2006) allowed for the discovery and release of the easy peeling cv. Porotan (Saito 2009) and a short list of other cultivars and selections (Takada et al. 2019). The genetics of the easy-peeling trait were worked out by pedigree analysis of the of the original crosses of two difficult-to-peel types, a half-diallel cross of Porotan and two other difficult-to peel cultivars, and a subsequent test cross of Porotan by its difficult-to-peel father and great grandfather Tanzawa (Takada et al. 2012). The authors conclude that the easy-peeling pellicle trait in C. crenata is controlled by a major recessive allele at a single locus. Takada et al. (2012) designated the easy-peeling locus P/p and offer evidence that the Tsukuba genotype is homozygous dominant (PP), the Tanzawa genotype is heterozygous (Pp), and the Porotan genotype is homozygous recessive (pp). They posit that the underlying mechanism of P/p in C. crenata may be different than the mechanisms underlying easy peeling in C. mollissima. This is based, in part, on the observation that all nuts produced by Porotan are easy to peel regardless of the pollen used to produce the nuts (Takada et al. 2010). This is in contrast with nuts produced by some easy to peel cultivars and hybrids of C. mollissima that are pollen-dependent, in other words, they produced difficult to peel nuts when pollinated by C. crenata but were easy to peel when pollinated by C. mollissima.
3.3 Korea
Castanea crenata also occurs naturally in Korea (Fig. 5.1). Although there is some speculation that Korean native chestnuts are hybrids between C. mollissima and C. crenata, and several taxon names have been proposed, most Korean cultivars are morphologically similar to their Japanese counterparts and many named cultivars are known (Kim 2006). Several Japanese cultivars of C. crenata were introduced to Korea subsequent to the appearance of the gall wasp Dryocosmus kuriphilus and are still widely grown there. Breeding efforts in Korea have focused on gall wasp resistance, chestnut blight resistance, and nut quality. Kim et al. (2005) announced the release of Daehan, a new blight-resistant, gall-wasp resistant cultivar with very high yields of large, good-quality nuts. The cross was made in 1980, preliminary selection was completed by 1990, advanced and regional trials were completed in 2003 and the cultivar was named Daehan in 2004 (Kim et al. 2005).
Variation in other agronomic and ornamental characteristics of Castanea crenata are also under selection by breeders (Kim et al. 2014, 2017; Saito 2009). Red leaves and red burs occur in some types and a new red bur cultivar was recently released in Korea (Kim et al. 2014).
3.4 China
Castanea mollissima is by far the most important of the Chinese species, as a nut crop, and from a breeding point of view (He et al. 2015). Vast amounts of morphological, physiological and genetic variation exists within the species (Huang 1998; Pereira-Lorenzo et al. 2016). Naturally occurring populations of C. mollissima are difficult to study because of the extent of alteration of the Chinese landscape by humans, and because the species exists almost entirely in cultivation (LaBonte et al. 2018). The other wild species in China include C. henryi and C. seguinii , and to a much lesser extent C. crenata (Fig. 5.1). Germplasm characterization of the Chinese species has recently included the complete chloroplast genome of C. mollissima cultivars (Zhu et al. 2019b), and complete chloroplast genome sequences for C. henryi (Gao et al. 2019) and C. seguinii (Zulfiqar et al. 2019). The response of C. mollissima to infestation by Dryocosmus kuriphilus was investigated by transcriptome analysis (Zhu et al. 2019a).
The ranges of Castanea henryi and C. seguinii largely overlap with that of C. mollissima, but they are found mostly in the wild state, or rarely cultivated locally (Fig. 5.1). Potentially useful genetic variation in tree form and habit, inflorescence type and phenology, and resistance to major diseases and pests exists in these two taxa (Gao et al. 2019; Zulfiqar et al. 2019). Castanea henryi is mostly important for its production of timber and other wood product, although the fruits, produced as a single nut per bur, are frequently found in markets throughout its range (Qin and Feng 2009). Some ecotypes of the naturally shrubby C. seguinii bloom more or less continuously throughout the growing season, with bisexual catkins at every node along the branch, and thus may have ripening fruits and fully-blooming flowers present on a single plant at the same time (Rutter et al. 1991). Castanea seguinii is regularly coppiced for firewood, but the new shoots are very precocious and will be bearing nuts, which are sold in local markets, within a year or so of cutting (Rutter et al. 1991).
Hundreds of named varieties of Castanea mollissima are known and grown in China, the world’s largest producer of chestnuts. Great variation exists regionally and many authors divide the cultivars into five separate regional groups: North, Northwest, Southwest, Southeast, and the Yangtze Valley Group. In the far northeast, adjacent to Korea, cultivars of C. crenata are grown. Many cultivars are propagated by grafting but seed propagation is also widespread. Cultivars in China are characterized by geographic origin (by regional group, by province or by growing area within province), fruit size and shape, quality, type and content of starches, density of kernels, and a long list of other agronomic characters and processing abilities. Yang et al. (2015) screened chestnut cultivars from ten different ecological regions. They tested a battery of compositional and nutritional factors and found cultivar differences and regional differences. In general, cultivars from the central regions (Hubei province) had higher total carbohydrate content. Cultivars from the deep south (Guandong and Zheijiang provinces) had the highest protein and polyphenol contents. Cultivars originating in the mid-south (Hunan) were highest in flavonoids and simple sugars, but lowest in fat content. A few of the best-known Chinese cultivars of C. mollissima are listed in Table 5.2.
3.5 Europe
Germplasm resources and characterization of European populations of Castanea sativa (Fig. 5.1) have been thoroughly studied and reviewed by Pereira-Lorenzo et al. (2006a, b, 2010, 2011, 2012). Mattioni et al. (2008) used genomic tools to investigate the active role of humans, over the past 9000 years, in determining not only the modern European distribution of C. sativa, but also the genetic structure of populations in naturalized forests, coppice stands (intensively managed for wood production), and chestnut orchards (managed for nut production). The species has ample genetic and adaptive variation across its wide geographic and climatic range, reflecting many distinct ecotypes (Casasoli et al. 2004, 2006; Martin et al. 2009; Villani et al. 2009). Boccacci et al. (2004) used SSRs developed in Quercus to type C. sativa cultivars. Costa et al. (2005) report the results of Portuguese chestnut germplasm characterization using SSRs. Variation exists in many traits of agronomic importance, including disease resistance (Vettraino et al. 2005). Native resistance to Dryocosmus kuriphilus in C. sativa has been reported recently in populations of grafted trees of the red salernitan ecotype (RSE) growing in the very important chestnut producing region of Campania, Italy (Nugnes et al. 2018). RSE is probably a group of closely related, morphologically similar cultivars including Mercoliana, Rossa di San Mango, Verdola and Palummina which differ, curiously, in their response to the gall former. The observation of resistance in RSE was based on lower oviposition rates, higher rates of larval mortality, fewer galls formed and smaller galls with higher rates of predation by Torymus sinensis. Prior to this report, there have been only two other indications of potentially resistant C. sativa cultivars; Pugnenga from Cuneo, and Savoye, from France (Sartor et al. 2015). The cultivar Palummina is marketed as, and protected by, and is the principal component of the Castagna di Montella IGP protected area, one of the oldest, most recognized and most prestigious denominations of geographic origin in Italy (Regione Campania 2015). Some of the C. sativa and hybrid cultivars most widely grown in Europe are listed in Table 5.2.
3.6 North America
There are currently recognized two species of Castanea naturally occurring in North America: C. dentata , the American chestnut, and C. pumila sensu lato, the perplexingly variable chinquapins (Craddock 1998; Miller et al. 1996; Perkins 2016; Rutter et al. 1991) (Fig. 5.1). Chinquapins vary widely in leaf shape and vestiture, tree stature, habit and form, across a wide geographic area. Along the Atlantic and Gulf coastal plains, most individuals are multi-stem shrubs maturing at less than 5 m tall. In the fire-prone sand hills of northern Florida, there are populations of C. pumila that mature at less than 2 m tall, forming 20-m wide or even larger thickets – each thicket composed apparently of a single, stoloniferous genet whose horizontal stems grow flat upon the ground and root themselves into the ground as they grow (Fig. 5.6). In the Appalachian Mountains, chinquapins are taller and more frequently fewer-stemmed. Some forms in northern Alabama reach heights of 10 m. The tallest chinquapins, in the pre-blight era, were recorded from the Ozark uplands of Arkansas and Missouri, and have been designated as a distinct taxon, C. pumila var. ozarkensis . The defining characteristic of C. pumila sensu lato is that it typically produces a single nut per involucre, and that the involucre opens along a single suture into two valves (Fig. 5.7).

Trailing chinquapins. At the extreme southernmost edge of its natural distribution, Castanea pumila assumes the habit of a stoloniferous shrub about 1 m tall, that forms large thickets up to 20 m (or greater) diameter, each apparently composed of a single genet. This photo was taken in Suwannee County, Florida and shows two adjacent genets that differ in height. Southern ecotypes of C. pumila have been assigned to several different taxa since their nineteenth century descriptions, but here we place them in C. pumila sensu lato pending further phylogenetic analysis at the population level. (Photo by J.H. Craddock)

The pistillate inflorescence of Castanea pumila will bear up to 9–12 (or more) cupules. Each cupule develops into a prickly involucre containing a single nut. (Photo by J.H. Craddock)
Only Castanea dentata was ever of any real economic importance, as a valuable source of excellent timber and for its delicious and nutritious nuts (Jaynes 1979; Smith 1950). Castanea dentata can grow to be a large tree, while C. pumila is a highly variable but usually shrubby, small tree (Paillet 1993). Both species had and continue to have important ecological functions in the forests of the eastern United States (Oak 2002, 2006; Southgate 2006). The chestnut blight pandemic, caused by Cryphonectria parasitica , completely removed C. dentata from its ecological niche in the forest canopy within 50 years of its first discovery in New York in 1904 (Anagnostakis 1987; Roane et al. 1986; Smith 2000). The rot-resistant remains of fallen giant American chestnut trees may still be found throughout eastern North America. But chestnuts are not extinct; the blight fungus only kills the shoots, not the roots. Although it continues to sprout new shoots from the bases of blight-killed stems, survival of the species varies greatly from site to site (Griffin 2000).
Populations of Castanea dentata persist as understory shrubs, suppressed by shade and eventually by blight (Griffin 1992; Parker et al. 1993). Many of the surviving specimens appear as clumps of multiple stems arising from a common root system (clones), although the connections are not always evident. The largest stems are often heavily cankered with blight and usually die before nuts are produced, however, individuals do rarely escape blight long enough to bloom (McWilliams et al. 2006; Stephenson et al. 1991). Surviving American chestnut trees, such as those frequently encountered in the mountains, are valuable for many reasons. Their potential as parents in a backcross breeding restoration program will confer local adaptation to the hybrids and ensure conservation of the native population’s genetic diversity (Hebard 2006). Even rarer are large surviving American chestnut trees (LSAs), characterized by swollen cankers with abundant callus. These very rare LSA individuals likely survive due to a combination of favorable circumstances including infection with attenuated (hypovirulent) strains of the blight fungus, ideal site conditions, and possibly a modicum of blight resistance (Fulbright 1999; Griffin 2000; Kolp et al. 2017). Overall, genetic diversity in C. dentata appears to be less than that of the other native species C. pumila sensu lato, and much lower than that of the Chinese chestnut, C. mollissima (Dane et al. 1998; Huang et al. 1994; Worthen et al. 2010).
The apparent distribution of genetic diversity in Castanea dentata is such that most of the variation can be found within populations, as is true for metapopulations of many wind-pollinated species that occur over broad ranges (Kubisiak and Roberds 2006), although recent work has shown much greater diversity in the southernmost populations, indicative of gene flow from southwest to northeast along the main axis of the Appalachian Mountains (Binkley 2008; Gailing and Nelson 2017; Li and Dane 2013; Shaw et al. 2012). The dominance of C. dentata in the forest canopy followed by its almost complete disappearance has caused major, long-term changes to the structure and health of the eastern North American forests (Oak 2002, 2006). Understanding the reproductive biology and natural ecology of C. dentata will be essential for its successful reintroduction (Dalgleish et al. 2016; Phelps et al. 2006; Wang et al. 2013). From a floristics perspective C. dentata is separated from C. pumila primarily by the number of pistils per cupule (Fig. 5.8). In the absence of flowering shoots the shrubby, understory post-blight form and habit of the two taxa can be confounding (Shaw et al. 2012).

The pistillate inflorescences of Castanea are contained in prickly involucres (the cupules) that typically bear three pistils, and are found at the base of the bisexual catkins. Illustrated here are the pistillate flowers of C. dentata at full bloom (peak of pollen receptivity). (Photo by J.H. Craddock)
Many authors have suggested that interspecific hybridization has played a role in forming the highly diverse North American chinquapins (Binkley 2008; Camus 1929; Dode 1908; Elias 1971; Kubisiak and Roberds 2006; Li and Dane 2013; Little 1977; Nixon 1997; Shaw et al. 2012). Chloroplast haplotype sharing between species does occur in areas of sympatry for Castanea dentata and C. pumila in the southern Appalachian region (Binkley 2008; Li and Dane 2013; Perkins 2016; Shaw et al. 2012), however, the relative importance of interspecific hybridization versus retention of polymorphisms predating speciation has not been investigated.
The commercial chestnut industry in North America is based entirely on introduced germplasm (Craddock 2002, 2009; Craddock and Pellegrino 1992). Nut production is divided into two very separate geographic regions with very different climates, soils and cultures: The East and the West (Nave 1998). The native and introduced Castanea species found growing in North America today are listed in Table 5.1. The native species C. dentata, and C. pumila, which occur naturally only in the East, are not important as nut crops. Commercial cultivars available through the nursery industry are listed in Table 5.2. Distribution of cultivars varies by region: in the West, the most widely-grown cultivars of C. sativa and Euro-Japanese hybrids are of recent introduction. Craddock and Pellegrino (1992) list the cultivars they successfully sent through the required 2-year post-entry quarantine. Of those on the list, Marigoule, Bouche de Bétizac and several Italian Marrone cvs. are now grown commercially in Washington, Oregon, California and Michigan. In the East, varieties and hybrids of C. mollissima dominate (Craddock et al. 2005). Some varieties, however, are grown only to a very limited extent (one or two growers). California and the Pacific Northwest states of Oregon and Washington produce chestnuts in an area of ideal climate, free of chestnut blight, and free of most of the major insect pests that plague chestnut growers in the East (Table 5.3). The single most important cultivar in the western region is the Euro-Japanese variety Colossal, which is vigorous and very productive. Chestnut flour made from dried Colossal is available commercially. Several western growers are topworking Colossal orchards to the recently introduced Italian Marrone varieties because of the higher nut quality of the Marrone types. Michigan represents a chestnut growing area typical of the American Midwest: very cold winters and hot, humid summers. Although Colossal is grown to a limited extent in the Midwest, it does not have suitable blight resistance for sustained harvests. Most Midwestern production is based on C. mollissima . Traditionally, most orchards were planted as seedling trees, which vary greatly in all agronomic characters. Newer orchards include grafted trees of improved cultivars. Problems of limited availability of nursery stock and delayed graft failure of the grafted trees have slowed the development of the Midwestern commercial chestnut industry. The University of Missouri Center for Agroforestry has had extensive chestnut cultivar trials underway since 1996. Recent work on cultivar evaluation has uncovered several promising cultivars of C. mollissima (Hunt et al. 2004). Shing has large nuts and is exceedingly vigorous and productive. Qing, Payne, Eaton, Homestead and Sleeping Giant all have great commercial potential (Metaxas 2013). In the Appalachian region, where there is still considerable nostalgia for the American chestnut, several cultivars of C. mollissima have been developed locally, including Amy, Gideon, Kohr, Mossbarger and Peach. Chinese chestnut is also employed in agroforestry and silvopasture applications in the small farms of Appalachia (Lovell et al. 2018) (Fig. 5.9).

Sheep are particularly well suited to the chestnut orchard, and in a managed silvopasture system, can contribute significantly to a diversified farm economy. (Photo by J.H. Craddock)
The introduction of Asian chestnut gall wasp into Georgia in the 1970s effectively destroyed the chestnut orchard industry there, but populations of the parasite are apparently now under biocontrol due to the parasitoid wasp Torymus sinensis. However, the Southeast has produced several outstanding cultivars of merit, most notably Nanking. Metaxas (2013) presents results of a formal chestnut cultivar evaluation and breeding program at the University of Tennessee at Chattanooga (Fig. 5.10). Qing, Shing and Payne appear outstanding among the Castanea mollissima cultivars for precocity and yield. Qing and Payne (Fig. 5.11a) are compact plants and Shing is extremely vigorous. Gideon may be the best all-around cultivar (Fig. 5.11b) for the eastern United States. Colossal showed great promise for precocity and nut size among the hybrid cultivars but soon proved too susceptible to chestnut blight. Eaton and Sleeping Giant have produced very attractive and good-flavored nuts although Eaton may not have adequate blight tolerance and has suffered from delayed graft failure in Tennessee (Metaxas 2013).

The Euro-Japanese cv. Marigoule was used in crosses with Castanea mollissima cv. Gideon in an effort to combine the good tree form and high yields of cv. Marigoule with the better disease resistance and excellent fruit quality of Gideon. (Photo by J.H. Craddock)

Castanea mollissima cv. Payne produces an excellent quality nut on a semi-dwarf tree that may be suitable for high density orchard plantings (a). Castanea mollissima cv. Gideon produces regular crops of an excellent quality, medium sized nut. The cultivar is widely adapted for cultivation across eastern North America (b). (Photos by J.H. Craddock)
4 Cytogenetics
Early investigations of genetic differences between Castanea species involved cytogenetic methods. Jaynes (1962) performed chromosome counts in accessions representing each Castanea species and nine interspecific hybrids. He found that all Castanea species and most of the interspecific hybrids had a somatic chromosome number of 2n = 2× = 24. However, triploidy and aneuploidy were observed in two interspecific hybrids resulting from crosses between Castanea species from North America and Eastern Asia (Jaynes 1962). More recently, work by Islam-Faridi et al. (2009, 2011, 2016) has revealed evidence of structural chromosomal differences between Castanea species. Islam-Faridi et al. (2011) found that the satellite region distal of the major 18S–28S ribosomal DNA (rDNA) locus in C. mollissima is larger than its counterpart in C. dentata . By using florescence in situ hybridization (FISH) to investigate a hypothesized reciprocal translocation between chestnut species, Islam-Faridi et al. (2016) found evidence that the C. mollissima cv. Vanuxem is heterozygous for a translocation involving linkage groups (LG) H and L.
In addition to the cytogenetic work indicating structural chromosomal differences between Castanea species, Sisco et al. (2014) recently shed light on cytoplasmic genome variation among chestnut species. Male sterility in chestnut was observed in the progeny of many intra- and interspecific crosses in the genus (Jaynes 1963, 1964; McKay 1942; Omura and Akihama 1980; Soylu 1992). In North America, male sterility has presented an obstacle to the introgression of disease resistance from C. mollissima to C. dentata because the male sterile phenotype has been observed in F1, BC1 and BC2 progeny descended from interspecific crosses between C. dentata and C. mollissima (Shi and Hebard 1997; Sisco et al. 2014) (Fig. 5.12). As a result, chestnut breeders involved in breeding C. dentata for blight resistance were limited in crossing designs that would produce male-fertile progeny. By analyzing noncoding chloroplast DNA sequences from the parents and progeny of 17 F1 and BC1 crosses involving C. dentata, C. mollissima and C. crenata in various combinations, Sisco et al. (2014) found that male sterility in interspecific hybrids was correlated perfectly with the D chloroplast haplotype of C. dentata. Interestingly, C. dentata trees containing non-D chloroplast haplotypes (e.g. M and P chloroplast haplotypes) did not produce male-sterile progeny when crossed with C. mollissima. Trees with these non-D haplotypes were primarily found in the southern half of the range of C. dentata. Based on the association of the male-sterile and male-fertile phenotypes (Fig. 5.12) with distinct chloroplast haplotypes of C. dentata, the authors posited that the sterility observed in hybrids between C. dentata and east Asian Castanea spp. is cytoplasmic male sterility (CMS), resulting from an interaction between mitochondrial genes inherited from C. dentata and nuclear genes inherited from C. mollissima . The findings of Sisco et al. (2014) were of practical value to chestnut breeders because they demonstrated that (1) substantial cytoplasmic diversity of phenotypic significance exists across the range of C. dentata and (2) because male sterility in crosses between C. dentata and Asian Castanea spp. is of the CMS type, male-fertile hybrids can be produced by using D cytotype C. dentata as the male parent in crosses with East Asian Castanea spp. or by using M or P cytotype C. dentata in any combination with East Asian Castanea spp. (Sisco et al. 2014).

The longistaminate catkins in Castanea dentata usually reach anthesis before the bisexual catkins do, and before the stigmas of the pistillate flowers are pollen receptive (a and b). Cytoplasmic male sterility in interspecific hybrids is frequently encountered with certain cytotype/genotype combinations. Many American x Chinese hybrids exhibit completely male-sterile, astaminate catkins, as shown here in a B2F2 at full bloom (c and d). (Photos by J.H. Craddock)
5 Molecular Breeding
5.1 Genetic and Genomic Methods in Chestnut Breeding
The recent availability of molecular genetic and genomic tools has allowed investigators to address several recurring questions in the field of chestnut breeding. What is the genetic architecture of resistance to the various pathogens of chestnut? Are genes that confer resistance to one pathogen tightly linked to genes controlling resistance to other pathogens or other desired phenotypes? Can selection strategies informed by molecular genetic and genomic methods produce desired phenotypes more quickly than breeding programs based entirely upon phenotypic selection? Over the past few decades researchers have used quantitative trait locus (QTL) mapping to identify genomic regions controlling several traits of importance for chestnut breeders and growers—namely, resistance to chestnut blight, resistance to Phytophthora root rot, nut traits and adaptive traits (Kubisiak et al. 1997, 2013; Nishio et al. 2018b; Santos et al. 2017b; Zhebentyayeva et al. 2014). Comparative genomic and transcriptomic studies have proposed candidate genes for disease resistance (Barakat et al. 2009, 2012; Kubisiak et al. 2013; Serrazina et al. 2015). And, most recently, the availability of genomic resources is beginning to make possible the use of genomic selection in chestnut breeding (Nishio et al. 2018a; Steiner et al. 2017; Westbrook et al. 2019). Nishio et al. (2018b) report genetic maps and QTLs for eight traits based on analysis of single nucleotide polymorphisms (SNPs) and simple sequence repeats (SSRs) in two breeding populations of Castanea crenata cultivars and selections. They showed very strong effects for QTLs associated with harvest date and pericarp splitting, and predict that marker-assisted selection can greatly improve breeding efficiency (Nishio et al. 2018b). Pereira et al. (2012) have reviewed the integration of new biotechnologies into chestnut breeding including molecular markers for selection at the genome level. Historically, isoenzymes, random amplified polymorphic DNAs (RAPDs) and amplification fragment length polymorphisms (AFLPs) were used. More recently SSRs and SNPs have come to the fore. SSR and SNP depend on four primary sources of genomic information: expressed sequence tags (ESTs); genomic DNA libraries enriched for repeating sequences; bacterial artificial chromosomes (BACs) and, increasingly, whole genome sequences (Fang et al. 2013).
5.2 Molecular Approaches for Blight Resistance Breeding
In North America, molecular breeding approaches have been used solely within the context of Castanea dentata restoration efforts (Burnham et al. 1986; Diskin et al. 2006; Westbrook 2018a, b; Westbrook et al. 2019; Wheeler and Sederoff 2009; Worthen et al. 2010). In an early paper describing his plan to use the backcross method to introgress blight resistance from C. mollissima into C. dentata , Burnham (1988) proposed that restriction fragment length polymorphisms (RFLPs) could be used to accelerate the breeding program by selecting backcross progeny that most closely resemble the genotype of the C. dentata recurrent parent, then subjecting the marker-selected progeny to phenotypic selection for chestnut blight resistance. Bernatzky and Mulcahy (1992) expanded upon the proposal to use marker-assisted selection (MAS) in C. dentata breeding by highlighting the need for genetic linkage maps that can be used to identify DNA markers linked to the genes controlling blight resistance.
Kubisiak et al. (1997) were the first in North America to use genetic mapping to assist chestnut breeding efforts. They used isozyme, RFLP and random amplified polymorphic DNA (RAPD) markers to create a genetic linkage map for an F2 family derived from two F1 crosses between Castanea dentata and C. mollissima and used phenotypic data from inoculation of the F2 progeny with chestnut blight fungus to identify seven genomic regions that appeared to condition a resistance response in the host. Only three of the seven regions were found to have an intermediate to large effect on resistance. The study of Kubisiak et al. (1997) was encouraging to chestnut breeders because it provided evidence that high levels of blight resistance could be conferred by a limited number of genomic regions, thus indicating that introgression of blight resistance from C. mollissima into C. dentata may be a realistic objective.
Taking advantage of new genomic resources, Kubisiak et al. (2013) proposed a list of candidate genes for blight resistance that are located within the three QTL intervals identified in the earlier work of Kubisiak et al. (1997). By comparing a genetic map of Castanea mollissima with the peach (Prunus persica (Rosaceae)) reference genome assembly, Kubisiak et al. (2013) found multiple segmentally homologous regions in the peach genome that span the three blight resistance QTLs in chestnut. The authors found that two of these regions in the annotated peach genome sequence contain genes for resistance to powdery mildew disease. Kubisiak et al. (2013) posited that the orthologous relationships supported by syntenic positions and sequence similarities between peach and chestnut suggest that these genomic regions may contain a set of conserved (prior to the divergence of Fagaceae and Rosaceae) genetic elements whose products respond to fungal invasion.
The development of Castanea mollissima genomic resources (Cannon et al. 2017; Sisco et al. 2005) by plant breeders has positioned the Chinese chestnut as a model organism for the study of evolution of woody plants and their relationships to other more closely related herbaceous species. Staton et al. (2015) used the reference genome to focus on disease resistance QTLs of potential importance to chestnut breeders and compare them to sequence data from other species. The unexpected levels of synteny preservation found among unrelated tree species imply that life history may play a more important role than has previously been recognized (Staton et al. 2015).
Recently, genomic selection has been proposed to accelerate recovery of blight resistance in The American Chestnut Foundation’s (TACF) backcross breeding program (Steiner et al. 2017; Westbrook 2017, 2018a, b). In the past, TACF has used progeny testing to identify advanced backcross (BC3F2) trees possessing high levels of blight resistance to supply seed for forest plantings. At current rates of progeny testing, it is expected to take 50 years to complete selection on the 5000 mother trees in TACF’s seed orchards (Westbrook 2017). Genomic selection, however, can greatly accelerate the process when used with accurate prediction models (Westbrook et al. 2019) (Figs. 5.13 and 5.14). Westbrook et al. (2019) used a genotyping by sequencing (GBS) approach to develop and apply a genomic prediction model to populations of segregating B3F2 American chestnut hybrids. They used a training population of more than 7000 B3F3 progeny of more than 500 open-pollinated, selected B3F2s. The B3F2s were descended from two Castanea mollissima sources of resistance (cvs. Clapper and Graves) and were planted in a seed orchard experimental design which had been screened by inoculation with two strains of C. parasitica (SG2-3, Ep155) and culled (Hebard 1994, 2006). The genomic predictions of progeny canker severity were as accurate as phenotypic estimates for Clapper B3F2s, but heritability was too low for the descendants of cv. Graves (Westbrook et al. 2019). Westbrook et al. (2019) conclude that the forthcoming chromosome-scale reference genome for C. dentata , combined with the genotypes for thousands of backcross individuals currently in the TACF state chapter programs, will greatly facilitate the elucidation of the C. mollissima contributions to blight resistance.

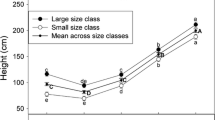
American chestnut hybrid seed orchard. A minimum of 20 genetic families (non-first cousin lines) of B3F2 hybrid seeds are planted at very high density (0.6 m × 1.2 m) in 150-tree plots, in anticipation of high selection pressure. Genomic prediction of Castanea parasitica tolerance, which is currently under development, will allow us to choose the very best 2–5 trees per 150-tree plot. The fully selected seed orchard should contain 40–100 blight tolerant trees with C. dentata morphology . (Photo by J.H. Craddock)

Three stages of selection in an American chestnut seed orchard. The B3F2 trees in the foreground are at the end of their first growing season after planting at very high density (0.3 m × 2 m). In the middle ground are 3-year-olds that have been inoculated with virus-free isolates of Castanea parasitica. The taller trees in the background are 6-year-olds that were selected for C. parasitica tolerance, tree form, and C. dentata leaf and twig morphological characters. (Photo by J.H. Craddock)
5.3 Molecular Approaches for Phytophthora Root Rot Resistance Breeding
Amid the growing realization that Phytophthora root rot (PRR) is an important obstacle to growth of chestnut in the southern United States, multiple groups began investigation of the genetic architecture of PRR resistance in the first decade of the twenty-first century. Kubisiak (2010) presented a major effect QTL for PRR resistance on chestnut linkage group (LG) E in BC1 progeny derived from crossing a PRR-susceptible Castanea dentata individual with a C. dentata-mollissima F1 descended from the C. mollissima cv. Nanking source of PRR resistance. In a study of multiple BC1 and BC4 mapping populations descended from PRR resistance sources C. mollissima cv. Nanking and C. mollissima cv. Mahogany, Zhebentyayeva et al. (2014) identified a major effect QTL for PRR resistance on LG_E, providing further support of the hypothesis of a limited number of genomic regions controlling PRR resistance.
Because PRR is also a major impediment to cultivation of Castanea sativa in Europe, genetic and genomic tools have been used to understand and employ PRR-resistance in breeding of this species (Robin et al. 2006). Santos et al. (2015b, 2017b) developed and used SSR and SNP markers to map QTLs for PRR resistance in two F1 families derived from interspecific crosses between the PRR-susceptible species C. sativa and the PRR-resistant species C. crenata . Santos et al. (2017b) identified QTLs for PRR resistance on LG_E and LG_K, which was consistent with preliminary results obtained by Zhebentyayeva (pers comm to Santos (2017b)) from descendants of crosses between C. dentata and C. mollissima . Santos et al. (2017b) hypothesized that the finding of common resistance QTLs within C. crenata and C. mollissima may indicate shared resistance mechanisms to fungal or fungal-like pathogens across the Castanea genus.
In Europe, comparison of the root transcriptomes of PRR-susceptible Castanea sativa and PRR-resistant C. crenata after inoculation with Phytophthora cinnamomi resulted in the identification of 283 genes that are differentially expressed in response to the pathogen. Santos et al. (2017a) filtered this list of 283 differentially expressed genes to identify 8 candidate genes for resistance to P. cinnamomi. Using digital polymerase chain reaction (dPCR), Santos et al. (2017a) identified one gene, Cast_Gnk2-like, that best discriminates between susceptible and resistant genotypes. Santos et al. (2017a) also found that pre-formed defenses are crucial for the resistance of C. crenata to P. cinnamomi infection.
Perkins et al. (2019) report on the discovery of new sources of host resistance to Phytophthora cinnamomi in the TACF backcross breeding program (Fig. 5.15). They screened ten interspecific hybrid families in a nursery setting, and although the primary scope of their effort was introgression of disease resistance into Castanea dentata for ecological restoration purposes, the first backcross progeny used in the trial all descend from C. mollissima cultivars of merit: Amy, Byron, Gideon, Lindstrom-99, Payne and Petersburg and so on may be of interest in the development of new nut cultivars with the flavor profile of C. dentata (Perkins et al. 2019).

Resistance to Phytophthora cinnamomi can be determined by screening trees in containers during their first year of growth in the nursery. Resistant progeny are then transplanted into PRR symptomatic orchards for screening against Cryphonectria parasitica. Phenotypic selection at the F1 and BC1 generations was sufficient to improve the average PRR resistance for these lines (Perkins et al. 2019). These first backcross American hybrids descend from a 2004 cross of a wild type Tennessee C. dentata x C. mollissima cv. Gideon. The selected F1s were backcrossed to other wild type Tennessee C. dentata in 2014. (Photo by J.H. Craddock)
5.4 Genetic Resources for Understanding Insect Resistance
Alma (2002) lists more than 40 kinds of insects and mites known to cause damage to Castanea species. They attack the roots, bark, twigs, leaves and fruits. Some of the most injurious pests are listed in Table 5.3. On a global scale, the chestnut gall wasp, Dryocosmus kuriphilus is surely the one insect that causes the most damage to chestnut production; when introduced from China to Japan in the 1940s, it caused drastic reductions in the harvest of C. crenata (Saito 2009), and soon after its discovery in the state of Georgia, USA, commercial production based on C. mollissima was almost completely wiped out (Payne et al. 1983). Its accidental introduction to Italy was quickly followed by a similar, catastrophic drop in yield – some areas in Cuneo Province lost the entire crop within a few years of infestation (Brussino et al. 2002; Sartor et al. 2015). A biocontrol, based on the parasitoid wasp Torymus sinensis, native to China, is currently deployed in Japan, Korea, Italy and USA, but the gall wasp remains problematic; episodic infestations continue to impact harvests worldwide, and the pest continues to spread in Europe (Ferracini et al. 2019; Gehring et al. 2018; Quacchia et al. 2008; Sartor et al. 2015). The best long term control may be genetic host resistance to infestation. Variation in gall wasp tolerance was first explored in the 1940s in Japan (Oho and Shimura 1970), and several gall-wasp resistant selections of C. crenata were released as cultivars in the 1950s (Saito 2009; Shimura 1972). Despite the continued ravages of the insect and its apparent ability to overcome resistance, Tsukuba, Tanzawa, Ishizuchi and Ginyose are still the most widely grown cultivars in Japan today (Pereira-Lorenzo et al. 2012; Saito 2009). The better, more resistant cvs. Kunimi and Shiho, were released in the 1980s but they are not so widely grown because about the time of their first availability the gall wasp parasitoid T. sinensis was released and began effecting biocontrol of D. kuriphilus. Since the 1990s, the focus of the Japanese chestnut breeders has returned to improvements in nut quality, particularly ease of pellicle removal in cultivars of C. crenata (Nishio et al. 2013; Saito 2009).
Dini et al. (2012) report on the discovery of a possibly novel mechanism for gall wasp resistance in the Euro-Japanese hybrid Bouche de Bétizac. They describe what they interpret to be a hypersensitive response in swelling buds to infestation by Dryocosmus kuriphilus . They used 3,3′-diaminobenzidine (DAB) in an assay for in vivo production of H2O2, itself an indicator of the stress-response glycoproteins germin and germin-like proteins (GLPs). GLPs are markers for stress response gene products and have oxalate oxidase (OxO) activity related to programmed cell death and the hypersensitive response (HR). They observed a positive DAB response in all buds of the resistant cv. Bouche de Bétizac but in none of the buds of the susceptible cv. Madonna, regardless of infestation state. They detected production of putative GLP using Real Time PCR. The observed hypersensitive response in Bouche de Bétizac has allowed the cultivar to remain free of galls after more than 10 years of exposure (Dini et al. 2012).
The molecular basis of Castanea mollissima responses to Dryocosmus kuriphilus infestations was very thoroughly investigated by Zhu et al. (2019a) through transcriptomic analysis of differentially-expressed genes during the different gall-formation stages in the life cycle of the wasp. In response to attack by the gall former, plant responses included plant hormone signaling, changes in transcription products, stimulation of Ca2+-mediated signal transduction pathways, and activation of secondary metabolic and other stress induced changes. The authors conclude that the genes involved represent C. mollissima gall defense genes (Zhu et al. 2019a).
5.5 Deciphering the Genetics of Agronomic and Nut Traits
The genetic architecture of important agronomic and nut traits in Castanea crenata and C. mollissima has been investigated by researchers in Japan, China and the USA. Researchers at the NARO Institute of Fruit Tree and Tea Science, Japan, used genetic linkage mapping and pedigree analysis to understand genetic control of nut traits (Nishio et al. 2011a, 2014b). The easy-peeling pellicle trait in the C. crenata cv. Porotan, the only commercial cultivar of C. crenata that possesses this trait, was investigated by Nishio et al. (2013) and Takada et al. (2019). Nishio et al. (2013) found that the easy-peeling pellicle was conferred in their C. crenata breeding populations by a recessive allele at a single gene. They also identified multiple SSR markers tightly linked to this locus. The difficulty of pellicle removal in C. crenata is a major disadvantage for commercial use of the species; thus, the SSR markers developed by Nishio et al. (2013) and the germplasm discovered by Takada et al. (2019) are currently being used to breed new C. crenata cultivars with easy-peel nuts (Nishio et al. 2018a, b; Takada et al. 2019).
With MAS for easy-peeling pellicles currently in use at the Institute of Fruit Tree and Tea Science, an additional objective of the breeding program is to release cultivars with easy-peeling pellicles and different nut harvesting dates from Castanea crenata cv. Porotan (Nishio et al. 2018b). For this purpose, Nishio et al. (2018b) used QTL mapping to study the genetic control of several important agronomic and nut traits in C. crenata: nut harvest date, nut weight, pericarp splitting, peach moth (Conogethes punctiferalis) infestation, number of burs per tree, number of nuts per tree, trunk diameter and yield per tree. One significant QTL each was identified for nut weight, peach moth infestation, number of burs per tree, number of nuts per tree, trunk diameter and yield per tree, while multiple significant QTLs were identified for both harvest date and pericarp splitting. The percentage of total phenotypic variance explained by the QTLs for harvest date and pericarp splitting was high (47.5–60.8% and 33.4–41.7%, respectively) and the SSR and SNP markers used in the study are expected to have immediate use for MAS in C. crenata breeding (Nishio et al. 2018b). Next, Nishio et al. (2018a) revisited study of many of these traits using a genome-wide association study on 99 phenotypically diverse C. crenata cultivars. Of the 12 total QTLs identified, 4 were associated with harvest date, 4 with nut weight, 3 with peach moth infestation and 1 with pericarp splitting. Accuracies of genomic selection were high for nut harvest (0.841) and moderate for peach moth infestation (0.604), indicating that genomic selection may reduce the cost of phenotypic evaluation of these traits by allowing selection at the seedling stage.
Microsatellite-based paternity analyses, using SSRs, allowed Nishio et al. (2014a, 2019) to estimate the effective pollen dispersal distance for cross-pollination in orchards of Castanea crenata cultivars. Because many Japanese chestnut cultivars are difficult to peel, and because it is difficult to separate the easy-peel nuts of cv. Porotan from the difficult-to-peel types after they have been harvested, the recommendation is to plant cv. Porotan alone in solid rows or blocks so that the easy-peel nuts can be harvested separately. The authors note that the effective pollen distance is not very far and thus suggest that the pollinizer varieties should be in adjacent rows or staggered in the row of the main cultivar (Nishio et al. 2014a, 2019).
QTL mapping has been used to facilitate marker-assisted selection in Castanea mollissima breeding programs in China where the desired traits mostly concern nut quality and harvest date. Ji et al. (2018) used the genotyping-by-sequencing method to construct a high density linkage map and identify QTLs for five nut traits. Three of the QTLs were associated with single nut weight, two were associated with nut thickness, five with nut width, one for nut height, and six QTLs associated with ripening period (Ji et al. 2018).
5.6 Future Prospects in Molecular Breeding of Chestnut
Despite the recent advances listed above, several important questions remain unanswered. Do the same genes control blight resistance in different chestnut species? Do the same genes control blight resistance in different cultivars within the same species? Are there different alleles encoding blight resistance within the same loci? Are there different loci for Phytophthora root rot resistance across different species and cultivars? What about viruses? Although the focus of many molecular studies in chestnut have been on breeding for disease resistance, several other important traits that vary in interesting ways may be amenable to molecular breeding approaches. These include nut crop yield, bud-bust phenology, tree architecture, and drought tolerance, nut flavors, nut higher lipid contents, nut morphology (Fig. 5.16). Moreover, untapped genetic potential exists in North American species. The excellent taste of American chestnuts and Ozark chinquapins, related to their lipid contents (Senter et al. 1994), and adaptation to fire disturbance in coastal chinquapins are traits of potential agronomic/culinary utility. Genetic study of the stoloniferous shrub (<1 m) plant architecture of the Castanea pumila coastal ecotype might facilitate production of a high-quality nut crop. Hybridization of trailing chinquapin with disease-resistant East Asian species that produce large chestnuts could produce dwarf cultivars suitable for intensive, high-density cultivation.

The prickles on the bur of the American chestnut may be the longest, proportionately, of any species (a). The fruits of Castanea dentata are typically covered in silky hairs (b). (Photo by J.H. Craddock)
6 Biotechnology
Biotechnologies , including biological controls, tissue culture techniques, genetic engineering and genomic marker systems, have great potential to advance restoration of Castanea dentata in eastern North America and to improve chestnut culture worldwide. The contributions, prospects and limitations of novel biotechnologies to forest tree breeding have been reviewed by Häggman et al. (2016) and by Tuskan et al. (2018).
6.1 Induced Mutation Breeding
Chestnut breeding using induced mutations was attempted in the USA after the chestnut blight pandemic decimated Castanea dentata populations in the twentieth century. Starting in the 1950s, multiple breeding programs irradiated thousands of C. dentata nuts and scions with the goal of generating blight-resistant mutants (Diller and Clapper 1965; Samman and Thor 1975; Singleton and Dietz 1974). Large plantings containing irradiated seedlings and their progeny were established in five U.S. states. However, no blight-resistant mutants were identified (Burnham 1988). Dietz (1978) attempted induced mutation breeding using ionizing radiation to achieve resistance to C. parasitica. Some of his irradiated trees, and some of their intercrossed progeny still exist in orchard plantings in Maryland, and are maintained by The American Chestnut Foundation (Burnworth 2002). Recent examination of surviving trees by Craddock (pers obs) indicated canker severity well within the range of C. dentata wild type trees infected with hypovirulent strains of C. parasitica, with no real evidence of increased resistance in irradiated trees or their progeny. Interest in American chestnut irradiation programs began to dwindle in the 1980s (Curry 2014).
6.2 Hypovirulence
Biological control of chestnut blight is based on transmissible hypovirulence, a phenomenon marked by the reduced virulence of the pathogenic fungus, rendering the fungus less damaging to the host plant (Grente 1965). Effective biocontrol was first observed on Castanea sativa in Italy (Biraghi 1950a, b) although the nature of the causal agent was not determined for more than 20 years. A dsRNA virus transmits the hypovirulent phenotype and the viral RNA can transform normal lethal cankers into slower-growing superficial bark cankers that do not kill the tree (Ćurković-Perica et al. 2017; Grente and Berthelet-Sauret 1978; Peever et al. 1997; Roane et al. 1986). In the 1970s hypoviruses were found in Michigan associated with recovering stands of C. dentata (Roane et al. 1986), although some of the Michigan hypovirulent strains were associated with mutations in the C. parasitica mitochondrial DNA (Fulbright 1999). By slowing the growth of the fungus, the virus allows the tree to live and bear fruit (Griffin 2000).
The biology of hypovirulence has been studied and reviewed extensively (Anagnostakis et al. 1998; Baidyaroy et al. 2000; Chen and Nuss 1999; Dierauf et al. 1997; Fulbright 1999; Griffin et al. 2006; MacDonald and Double 2006; Monteiro-Vitorello et al. 2000). Hebard et al. (1984) showed that host resistance in Castanea is a key factor allowing expression of hypovirulence (superficial cankers with reduced canker expansion). Barriers to virus transmission, one of the chief limitations to effective biocontrol, can be overcome using biotechnologies (Choi et al. 2012; Double et al. 2017; Zhang and Nuss 2016).
6.3 Genetic Engineering
The marriage of in vitro culture and molecular biology of trees, including chestnut, was reviewed by Chang et al. (2018). Beginning around the same time as the American Chestnut Foundation’s backcross breeding program, scientists at the State University of New York’s College of Environmental Science and Forestry (SUNY-ESF) and collaborators have pursued a genetic engineering approach to increase blight resistance in Castanea dentata (Chang et al. 2018; Steiner et al. 2017). An important breakthrough in this area occurred with development of a method to establish C. dentata somatic embryo cultures (Caraway et al. 1994; Merkle et al. 1991). Newhouse et al. (2014) used in vitro methods to study heterologous expression of oxalate oxidase in transformed C. dentata that show high levels of tolerance to C. parasitica infection. In vitro homologous expression of a CsCh3 chitinase in transformed somatic embryos of C. sativa with potential resistance to C. parasitica was reported by Corredoira et al. (2012, 2016). The use of somatic embryos facilitated the design of an effective transformation system for C. dentata (Andrade et al. 2009, Polin et al. 2006; Xing et al. 1999). The main blight tolerance gene chosen by researchers in this area is a gene encoding oxalate oxidase, which catalyzes the degradation of oxalate into H2O2 and CO2 and is produced in a variety of unrelated plants, such as strawberry, beet, peanut, apricot and most cereal grains (Chang et al. 2018; Steiner et al. 2017). Oxalate production by some plant pathogenic fungi, including the chestnut blight fungus, induces a programmed cell death response in plant tissue that is required for disease development (Hebard and Shain 1988; Kim et al. 2008). Based on observations that virulent strains of C. parasitica produce greater quantities of oxalate than hypovirulent strains (Chen et al. 2010; Havir and Anagnostakis 1983), the oxalate oxidase gene was chosen as a lead candidate gene for transformation (Chang et al. 2018). Tests of transgenic C. dentata plants expressing the oxalate oxidase transgene have shown that these plants exhibit enhanced resistance to C. parasitica and transmit blight resistance to a portion of their progeny (Newhouse et al. 2014; Zhang et al. 2013). The first field trials of transgenic C. dentata were planted in 2006 under a permit from the United States Department of Agriculture, Animal Plant Health Inspection Service (USDA-APHIS) (Steiner et al. 2017). There is some concern in the scientific community about public resistance to a transgenic approach (Barnhill-Dilling and Delborne 2019). Researchers at SUNY-ESF are currently in the process of seeking deregulation of the best-performing transgenic lines through USDA-APHIS (Chang et al. 2018).
Researchers in the United States have proposed that genetic engineering can be merged with backcross breeding to create Castanea dentata populations that combine most, if not all, of the following attributes: (1) blight resistance from the OxO transgene, (2) Phytophthora root rot resistance introgressed from the East Asian Castanea species, (3) blight resistance introgressed from the East Asian Castanea species, (4) adaptation to local environmental conditions from a diverse set of C. dentata recurrent parents and (5) an effective population size sufficiently large as to be at minimal long-term risk of extinction due to inbreeding and genetic drift (Steiner et al. 2017; Westbrook 2018a, b). The long-term breeding goal for C. dentata in North America is unique in that resistance to two diseases must be introgressed into wild populations to facilitate restoration of the species (Westbrook et al. 2019; Worthen et al. 2010).
Biotechnology research on chestnut in Europe and Asia is fundamentally different from that in North America because the main emphasis is on cultivar development and preservation, rather than species restoration. As a result, more effort has been spent on micropropagation and preservation of superior genotypes than on transformation. One very effective use of micropropagated material in plant breeding is the ability to clone hybrid progeny before screening. Santos et al. (2015a, 2017a) used plantlets generated by in vitro propagation to study the genetics of resistance to ink disease. The ability to clone progenies, before screening, is a powerful tool for selection for disease resistance (Santos et al. 2015a). Researchers in Europe are also in the process of using genetic engineering to increase resistance to both blight and ink disease in C. sativa . Corredoira et al. (2012) engineered C. sativa to overexpress a native thaumatin-like protein gene (CsTL1), which encodes a protein that was earlier shown to possess in vitro antifungal activity against Trichoderma viride and Fusarium oxysporum. A gene from C. sativa was used to transform hybrid poplars by Moreno-Cortez et al. (2012). They used the C. sativa RAV1 gene which is homologous to an Arabidopsis TEM gene. The transgenic poplars overexpressed CsRAV1 and showed early formation of current-growing-season branching, a possibly adaptive trait the authors consider important to biomass production (Moreno-Cortez et al. 2012).
7 Conclusions and Prospects
Chestnut researchers and growers now have at their disposal new tools, new techniques, new information and better access to genetic resources that should, in the near future, permit great gains in the movement towards several important breeding objectives: increased resistance to the most limiting chestnut diseases, tolerance to the most troublesome pests and improvements in fruit quality and yields. Cutting-edge gene editing technologies such as those based on the CRISPR/Cas9 systems may have tremendous applications for chestnut (Häggman et al. 2016; Tuskan et al. 2018). The American Chestnut Foundation is currently funding range-wide, and genome-wide sampling in an effort to document adaptive variation in Castanea dentata that may be useful to the species as it confronts rapid climate change (Westbrook 2018a, b; Westbrook et al. 2019), and conversations have begun within the scientific community about possible assisted migration as a conservation strategy (Fig. 5.17). Habitat modelling to predict future geographic ranges can be based on current and historic occurrence data for the trees and for their pathogens and pests (Phillips et al. 2006; Wang et al. 2010, 2013). Application of the knowledge presented here offers great opportunities to improve forest stands of chestnut for the sustainable production of high-quality lumber and other forest products in East Asia, North America and in Europe.

These third backcross American chestnut hybrids were bred from some of the southernmost populations of Castanea dentata . They exhibit good tree form, adequate blight resistance, and represent a potentially very important germplasm resource for adaptive variation as the species copes with climate change. (Photo by J.H. Craddock)
References
Abrams MD, Nowacki GJ (2008) Native Americans as active and passive promoters of mast and fruit trees in the eastern USA. The Holocene 18(7):1123–1137
Alexander MT, Worthen LM, Craddock JH (2005) Conservation of Castanea dentata germplasm of the southeastern United States. Acta Hortic 693:485–490
Alma A (2002) Insetti e acari. In: Bounous G (ed) Il castagno: coltura, ambiente ed utilizzazioni in Italia e nel mondo. Edagricole, Bologna, pp 115–123
Anagnostakis SL (1987) Chestnut blight: the classical problem of an introduced pathogen. Mycologia 79(1):23–37
Anagnostakis SL (2001) The effect of multiple importations of pests and pathogens on a native tree. Biol Invasions 3(3):245–254
Anagnostakis SL (2012) Chestnut breeding in the United States for disease and insect resistance. Plant Dis 96:1392–1403
Anagnostakis SL (2019) The Connecticut agricultural experiment station. https://portal.ct.gov/CAES/ABOUT–CAES/Staff–Biographies/Sandra–L–Anagnostakis. Accessed 28 Mar 2019
Anagnostakis SL, Chen B, Geletka LM, Nuss DL (1998) Hypovirus transmission to ascospore progeny by field-released transgenic hypovirulent strains of Cryphonectria parasitica. Phytopathology 88(7):598–604. https://doi.org/10.1094/PHYTO.1998.88.7.598
Andrade GM, Nairn CJ, Le HT, Merckle SA (2009) Sexually mature transgenic American chestnut trees via embryogenic suspension-based transformation. Plant Cell Rep 28:1385–1397. https://doi.org/10.1007/s00299-009-0738-7
Avanzato D, Bounous G (eds) (2009) Following chestnut footprints (Castanea spp.). Scrip Hortic 9:1–175
Baidyaroy D, Huber DH, Fulbright DW, Bertrand H (2000) Transmissible mitochondrial hypovirulence in a natural population of Cryphonectria parasitica. Mol Plant-Microbe Interact 13:88–95
Barakat A, Loreto DS, Zhang Y et al (2009) Comparison of the transcriptomes of American chestnut (Castanea dentata) and Chinese chestnut (Castanea mollissima) in response to the chestnut blight infection. BMC Plant Biol 9:51. https://doi.org/10.1186/1471–2229–9–51
Barakat A, Staton M, Cheng CH et al (2012) Chestnut resistance to the blight disease: insights from transcriptome analysis. BMC Plant Biol 12:38. https://doi.org/10.1186/1471-2229-12-38
Barbour PL (ed) (1986) The complete works of captain John Smith (1580–1631) vol 1. A map of Virginia. With a description of the countrey [sic], the commodities, people, government and religion. University of North Carolina Press, Chapel Hill
Barnhill-Dilling S, Delborne JA (2019) The genetically engineered American chestnut tree as opportunity for reciprocal restoration in the Haudenosaunee communities. Biol Conserv 232:1–7
Barstow M (2018) Castanea crenata. In: The IUCN red list of threatened species 2018. https://www.iucnredlist.org/species/62004433/62004435. Accessed 1 July 2018
Bellini E, Nin S (2009) Italy. In: Avanzato D, Bounous G (eds) Following chestnut footprints (Castanea spp.). Scrip Hortic 9:85–95
Bernatzky R, Mulcahy DL (1992) Marker-aided selection in a backcross breeding program for resistance to chestnut blight in the American chestnut. Can J For Res 22(7):1031–1035. https://doi.org/10.1139/x92-137
Bignami GR, Salsotto A (1983) La civiltà del castagno. L’Arciere, Cuneo
Binkley MA (2008) The phylogeography of North American chestnuts and chinquapins (Castanea Mill., Fagaceae). MS thesis, University of Tennessee at Chattanooga. https://scholar.utc.edu/theses/198
Biraghi A (1950a) Caratteri di resistenza in Castanea sativa nei confronti di Endothia parasitica. Boll Staz Patol Veg Roma 7:161–171
Biraghi A (1950b) La distribuzione del cancro del castagno in Italia. Ital J For Mt Environ 5:18–21
Boccacci P, Akkak A, Torello Marinoni D et al (2004) Typing European chestnut (Castanea sativa Mill.) cultivars using oak simple sequence repeat markers. HortSci 39:1212–1216
Bolvansky M, Tarinova D (2009) Slovakia. In: Avanzato D, Bounous G (eds) Following chestnut footprints (Castanea spp.). Scrip Hortic 9:122–127
Bounous G (ed) (2002) Il castagno: coltura, ambiente ed utilizzazioni in Italia e nel mondo. Edagricole, Bologna
Bounous G, De Guarda A (2002) In: Bounous G (ed) Il castagno: coltura, ambiente ed utilizzazioni in Italia e nel mondo. Edagricole, Bologna, pp 173–176
Bourgeois C (1992) Le châtaignier: un arbre, un bois. Institut pour le Developpement Forestier, Paris
Bruneton-Governatori A (1984) Le pain de bois: ethnohistoire de la châtaigne et du châtaignier. Eché, Toulouse
Brussino G, Bosio G, Baudino M et al (2002) Pericoloso insetto esotico per il castagno europeo. Inf Agrar 37:59–62
Burnham CR (1988) The restoration of the American chestnut. Am Sci 76:478–487
Burnham CR, Rutter PA, French DW (1986) Breeding blight-resistant chestnuts. Plant Breed Rev 4:347–397
Burnworth E (2002) A brief history of the efforts of Stronghold, Inc to restore the American chestnut: 1969 to the present. https://www.acf.org/wp-content/uploads/2016/09/StrongholdHist.pdf. Accessed 14 Jan 2019
Camus A (1929) Les chataigniers. Monographie des genres Castanea et Castanopsis. In: Encyclopedie economique de sylviculture. Paul Lechevalier, Paris
Cannon N, Staton ME, Addo-Quaye C et al (2017) The physical and genetic structure of the Chinese chestnut (Castanea mollissima) genome. Plant and animal genome XXV conference, San Diego, CA, Jan 14–18 2017
Carraway DT, Wilde HD, Merkle SA (1994) Somatic embryogenesis and gene transfer in American chestnut. J Am Chestnut Found VIII(1):29–33
Casasoli M, Pot D, Plomion C et al (2004) Identification of QTLs affecting adaptive traits in Castanea sativa Mill. Plant Cell Environ 27:1088–1101
Casasoli M, Derory J, Morera-Dutrey C et al (2006) Comparison of quantitative trait loci for adaptive traits between oak and chestnut based on an expressed sequence tag consensus map. Genetics 172(1):533–546. https://doi.org/10.1534/genetics.105.048439
Casey J, Casey B (2009) Australia. In: Avanzato D, Bounous G (eds) Following chestnut footprints (Castanea spp.). Scrip Hortic 9:14–19
Chang S, Mahon EL, MacKay HA et al (2018) Genetic engineering of trees: progress and new horizons. In Vitro Cell Dev Biol Plant 54:341–376. https://doi.org/10.1007/s11627-018-9914-1
Chen B, Nuss DL (1999) Infectious cDNA clone of hypovirus CHV1-Euro7: a comparative virology approach to investigate virus–mediated hypovirulence of the chestnut blight fungus Cryphonectria parasitica. J Virol 73:985–992
Chen C, Sun Q, Narayanan B et al (2010) Structure of oxalacetate acetylhydrolase, a virulence factor of the chestnut blight fungus. J Biol Chem 285(34):26685–26696
Cherubini G (1981) La civiltà del castagno in Italia alla fine del medioevo. Archeologia medievale 8:247–280
Choi GH, Dawe AL, Churbano A et al (2012) Molecular characterization of vegetative incompatibility genes that restrict hypovirus transmission in the chestnut blight fungus Cryphonectria parasitica. Genetics 190:113–127
Conedera M, Krebs P, Tinner W et al (2004a) The cultivation of Castanea sativa (Mill.) in Europe, from its origin to its diffusion on a continental scale. Veget Hist Archaeobot 13:161–179. https://doi.org/10.1007/s00334–004–0038–7
Conedera M, Manetti MC, Giudici F, Amorini E (2004b) Distribution and economic potential of the sweet chestnut (Castanea sativa Mill.) in Europe. Ecol Mediterr 30:179–193
Corredoira E, Valladares S, Allona I et al (2012) Genetic transformation of European chestnut somatic embryos with a native thaumatin-like protein (CsTL1) gene isolated from Castanea sativa seeds. Tree Physiol 32(11):1389–1402. https://doi.org/10.1093/treephys/tps098
Corredoira E, San José MC, Vieitez AM et al (2016) Agrobacterium-mediated transformation of European chestnut somatic embryos with a Castanea sativa (Mill.) endochitinase gene. New For 47:669. https://doi.org/10.1007/s11056-016-9537-5
Costa R, Valdiviesso T, Marum L et al (2005) Characterization of traditional Portuguese chestnut cultivars by nuclear SSRs. Acta Hortic 693:437–440
Craddock JH (1998) Chestnut resources in North America. Ann Rep North Nut Growers Assoc 89:19–30
Craddock JH (2002) Stati Uniti. In: Bounous G (ed) Il castagno: coltura, ambiente ed utilizzazioni in Italia e nel mondo. Edagricole, Bologna, pp 278–283
Craddock JH (2009) USA. In: Avanzato D, Bounous G (eds) Following chestnut footprints (Castanea spp.). Scrip Hortic 9:168–174
Craddock JH, Pellegrino S (1992) Recent introduction of European chestnut varieties into the USA. In: Double ML, MacDonald WL (eds) Proceedings of the international chestnut conference. West Virginia University Press, Morgantown, pp 197–200
Craddock JH, Alexander MT, Alexander SH et al (2005) Chestnut cultivar evaluations in Tennessee: orchard establishment and early germplasm characterization. Acta Hortic 693:465–467
Crandall BS, Gravatt GF, Ryan MM (1945) Root disease of Castanea species and some coniferous and broadleaf nursery stocks, caused by Phytophthora cinnamomi. Phytopathology 35:162–180
Ćurković-Perica M, Ježić M, Karin V et al (2017) Biocontrol of chestnut blight: (in) stable infection of the chestnut blight fungus by Cryphonectria hypovirus 1. 7th Congress of European microbiologists, Federation of European Microbiological Societies, 9–13 July 2017, Valencia, Spain, abstract 713
Curry HA (2014) Radiation and restoration; or, how best to make a blight-resistant chestnut tree. Environ Hist 19:217–238
Dalgleish HJ, Nelson CD, Scrivani JA, Jacobs DF (2016) Consequences of shifts in abundance and distribution of American chestnut for restoration of a foundation forest tree. Forests 7(1):4. https://doi.org/10.3390/f7010004
Dane F, Harrison S, Huang H (1998) Evaluation of the genetic diversity in Castanea pumila variety ozarkensis through isozyme analysis and DNA amplification fingerprinting. Ann Rep North Nut Grow Assoc 89:31–35
Davis DE (2003) Where there are mountains: an environmental history of the southern Appalachians. University of Georgia Press, Athens
Day G (1953) The Indian as an ecological factor in the northeastern forest. Ecology 34:329–346
Dierauf T, Artman J, Elkins JR et al (1997) High level of chestnut blight control on grafted American chestnut trees inoculated with hypovirulent strains. J Arbor 23:87–88
Dietz A (1978) The use of ionizing radiation to develop a blight resistant American chestnut, Castanea dentata, through induced mutations. In: MacDonald WL, Cech FC, Luchok J, Smith C (eds) Proceedings of the American chestnut symposium. West Virginia University Press, Morgantown, pp 17–20
Diller JD, Clapper RB (1965) A progress report on attempts to bring back the chestnut tree in the eastern United States, 1954–1964. J For 63:186–188
Dini F, Sartor C, Botta R (2012) Detection of a hypersensitive reaction in the chestnut hybrid ‘Bouche de Bétizac’ infested by Dryocosmus kuriphilus Yasumatsu. Plant Physiol Biochem 60:67–73
Diskin M, Steiner KC, Hebard FV (2006) Recovery of American chestnut characteristics following hybridization and backcross breeding to restore blight-ravaged Castanea dentata. For Ecol Manag 223:439–447
Dode ML-A (1908) Notes dendrologiques sur les châtaigniers. Bull Soc Dendrol France 8:140–159
Double ML, Nuss DL, Rittenour WR et al (2017) Long-term field study of transgenic hypovirulent strains of Cryphonectria parasitica in a forest setting. For Pathol 47(6). https://doi.org/10.1111/efp.12367
Elias TS (1971) The genera of Fagaceae in south-eastern United States. J Arnold Arbor 52:159–195
Fang G-C, Blackmon BP, Staton ME et al (2013) A physical map of the Chinese chestnut (Castanea mollissima) genome and its integration with the genetic map. Tree Genet Genomes 9:525–537. https://doi.org/10.1007/s11295–012–0576–6
Fei S, Liang L, Paillet FL et al (2012) Modelling chestnut biogeography for American chestnut restoration. Divers Distrib 18:754–768
Ferracini C, Ferrari E, Pontini M et al (2019) Effectiveness of Torymus sinensis: a successful long-term control of the Asian chestnut gall wasp in Italy. J Pest Sci 92:353–359. https://doi.org/10.1007/s10340–018–0989–6
Fulbright DW (1999) Chestnut blight and hypovirulence. In: Stacey G, Keen NT (eds) Plant-microbe interactions, APS Press, vol 4. St. Paul, Minnesota, pp 57–79
Gailing O, Nelson CD (2017) Genetic variation patterns of American chestnut populations at EST-SSRs. Botany 95:799–807
Gao XX, Yan F, Liu M et al (2019) The complete chloroplast genome sequence of an endemic species Pearl chestnut (Castanea henryi). Mitochon DNA B Resour 4(1):551–552. https://doi.org/10.1080/23802359.2018.1553522
Gehring E, Bellosi B, Quacchia A, Conedera M (2018) Assessing the impact of Dryocosmus kuriphilus on the chestnut tree: branch architecture matters. J Pest Sci 91:189–202. https://doi.org/10.1007/s10340–017–0857–9
Grau PB (2003) Introduction of European chestnut cultivars (Castanea sativa Mill.), Euro-Japanese hybrids (Castanea crenata x Castanea sativa) and Japanese cultivars (Castanea crenata Sieb et Zucc.) to Chile. First Results. Agricult Técn 63(3):329–336
Grente J (1965) Les formes hypovirulentes d’Endothia parasitica et les espoirs de lutte contre le chancre du châtaignier. Academie d’Agriculture de France, Extrait du Proces-verbal de la Seance 51:1033–1037
Grente J, Berthelay-Sauret S (1978) Biological control of chestnut blight in France. In: MacDonald WL, Cech FC, Luchok J, Smith C (eds) Proceedings of the American chestnut symposium. West Virginia University Press, Morgantown, pp 30–34
Griffin GJ (1992) American chestnut survival in understory mesic sites following the chestnut blight pandemic. Can J Bot 70:1950–1956
Griffin GJ (2000) Blight control and restoration of the American chestnut. J For 98(2):22–27
Griffin GJ, Elkins JR, McCurdy D, Griffin SL (2006) Integrated use of resistance, hypovirulence, and forest management to control blight on American chestnut. In: Steiner KC, Carlson JE (eds) Restoration of American chestnut to forest lands: proceedings of a conference and workshop, May 4–6 2004, North Carolina Arboretum, Asheville, NC. Natural resources report NPS/NCR/CUE/NRR – 2006/001, National Park Service, Washington, DC, pp 97–107
Häggman H, Sutela S, Fladung M (2016) Genetic engineering contribution to forest tree breeding efforts. In: Vettori C et al (eds) Biosafety of forest transgenic trees. Springer, Dordrecht. https://doi.org/10.1007/978-94-017-7531-1_2
Havir EA, Anagnostakis SL (1983) Oxalate production by virulent but not by hypovirulent strains of Endothia parasitica. Physiol Plant Pathol 23(3):369–376. https://doi.org/10.1016/0048–4059(83)90021–8
He SA, Yi TS, Pei SJ, Huang HW (2015) Crop plants and their wild relatives. In: Hong DY, Blackmore S (eds) Plants of China: a companion to the flora of China. Cambridge University Press, Cambridge, UK, pp 283–308
Hebard FV (1994) Inheritance of juvenile leaf and stem morphological traits in crosses of Chinese and American chestnut. J Hered 85:440–446
Hebard FV (2006) The backcross breeding program of the American Chestnut Foundation. J Am Chestnut Found XIX:55–78
Hebard FV, Shain L (1988) Effects of virulent and hypovirulent Endothia parasitica and their metabolites on ethylene production by bark of American and Chinese chestnut and scarlet oak. Phytopathology 78(6):841–845
Hebard FV, Griffin GJ, Elkins JR (1984) Developmental histopathology of cankers incited by hypovirulent and virulent isolates of Endothia parasitica on susceptible and resistant chestnut trees. Phytopathology 74(2):140–149
Huang HW (1998) Review of current research of the world Castanea species and importance of germplasm conservation of China native Castanea species. J Wuhan Bot Res 16:171–176
Huang H, Dane F, Norton JD (1994) Allozyme diversity in Chinese, Seguin and American chestnut (Castanea spp.). Theor Appl Genet 88:840–849
Hunt KL, Gold MA, Warmund MR (2004) Chinese chestnut cultivar performance in Missouri. Acta Hortic 693:145–148
Islam-Faridi N, Nelson CD, Sisco PH et al (2009) Cytogenetic analysis of American chestnut (Castanea dentata) using fluorescent in situ hybridization. Acta Hortic 844:207–210
Islam-Faridi N, Majid MA, Nelson CD (2011) Physical location of 18s–28s and 5s ribosomal RNA genes in Chinese chestnut (Castanea mollissima). In: Nelson CD, Rousseau RJ, Yuceer C (eds) Proceedings of the 31st southern forest tree improvement conference, Biloxi, Mississippi, 14–16 June 2011, pp 28–31. http://www.sftic.org
Islam-Faridi N, Majid MA, Zhebentyayeva T et al (2016) FISH confirmation of a reciprocal translocation in chestnut. Cytogenet Genome Res 148(2–3):144
Jacobs DF, Dalgleish HJ, Nelson CD (2013) A conceptual framework for restoration of threatened plants: the effective model of American chestnut (Castanea dentata) reintroduction. New Phytol 197:378–393
Jarman R, Hazell Z, Campbell G et al (2019) Sweet chestnut (Castanea sativa Mill.) in Britain: re-assessment of its status as a Roman archaeophyte. Britannia 10:1–26. https://doi.org/10.1017/S0068113X19000011
Jaynes RA (1962) Chestnut chromosomes. For Sci 8:372–377. https://doi.org/10.1093/forestscience/8.4.372
Jaynes RA (1963). Male sterility in Castanea. Proceedings of the XI international congress of genetics. Hague, Netherlands
Jaynes RA (1964) Interspecific crosses in the genus Castanea. Silvae Genet 13:146–154
Jaynes RA (1979) Chestnuts. In: Jaynes RA (ed) Nut tree culture in North America. Northern Nut Growers Association, Hamden, pp 111–127
Ji F, Wei W, Liu Y et al (2018) Construction of a SNP-based high-density genetic map using genotyping by sequencing (GBS) and QTL analysis of nut traits in Chinese chestnut (Castanea mollissima Blume). Front Plant Sci 9:816. https://doi.org/10.3389/fpls.2018.00816
Johnson GP (1988) Revision of Castanea sect. Balanocastanon (Fagaceae). J Arnold Arbor 69:25–49
Kim MJ (2006) Chestnut cultivars in Korea. Korea Forest Research Institute, Suwon, pp 37–176
Kim MJ, Kim S-C, Hwang M-S et al (2005) A new high-yielding, large nut chestnut cultivar, “Daehan” suitable for processing. Korean J Breed 37(1):71–72
Kim KS, Min J-Y, Dickman MB (2008) Oxalic acid is an elicitor of plant programmed cell death during Sclerotinia sclerotiorum disease development. Mol Plant-Microbe Interact 21(5):605–612
Kim MJ, Lee U, Park Y (2014) ‘Jahong’: A new indigenous Korean chestnut cultivar with reddish burrs. HortSci 49(12):1588–1589
Kim SH, Lee A, Kwon HY et al (2017) Analysis of flowering and nectar characteristics of major four chestnut cultivars (Castanea spp.). Kor J Apic 32(3):237–246
Klinac D, Knowles R (2009) New Zealand. In: Avanzato D, Bounous G (eds) Following chestnut footprints (Castanea spp.). Scrip Hortic 9:103–105
Kolp M, Double M, Fulbright DW et al (2017) Fungal community structure in chestnut blight cankers on American chestnut in Michigan and Wisconsin. Ecological Society of America annual meeting, 6–11 August 2017, Portland
Kotobuki K (1994) Chestnut. In: Matsuo T (ed) Collected data of plant genetic resources. Kodansya, Tokyo, pp 1174–1184
Krebs P, Conedera M, Pradella M et al (2004) Quaternary refugia of the sweet chestnut (Castanea sativa Mill.): an extended palynological approach. Veg Hist Archaeobotany 13:145–160. https://doi.org/10.1007/s00334–004–0041–z
Kubisiak TL (2010) QTL analysis of Phytophthora resistance in a BC1 population. NE-1033 Annual Meeting Minutes, 2010, Maggie Valley
Kubisiak TL, Roberds JH (2006) Genetic structure of American chestnut populations based on neutral DNA markers. In: Steiner KC, Carlson JE (eds) Restoration of American chestnut to forest lands – proceedings of a conference and workshop, May 4–6 2004, North Carolina Arboretum, Asheville, NC. Natural resources report NPS/NCR/CUE/NRR – 2006/001, National Park Service, Washington, DC, pp 109–122
Kubisiak TL, Hebard FV, Nelson CD et al (1997) Molecular mapping of resistance to blight in an interspecific cross in the genus Castanea. Phytopathology 87(7):751–759
Kubisiak TL, Nelson CD, Staton ME et al (2013) A transcriptome-based genetic map of Chinese chestnut (Castanea mollissima) and identification of regions of segmental homology with peach (Prunus persica). Tree Genet Genomes 9:557–571. https://doi.org/10.1007/s11295–012–0579–3
LaBonte NR, Zhao P, Woeste K (2018) Signatures of selection in the genomes of Chinese chestnut (Castanea mollissima Blume): the roots of nut tree domestication. Front Plant Sci 9:810. https://doi.org/10.3389/fpls.2018.00810
Li X, Dane F (2013) Comparative chloroplast and nuclear DNA analysis of Castanea species in the southern region of the USA. Tree Genet Genomes 9:107–116. https://doi.org/10.1007/s11295–012–0538–z
Little EL (1977) Atlas of United States trees, vol 4. US Government printing office, Washington, DC, p 17
Lovell ST, Dupraz C, Gold MA et al (2018) Temperate agroforestry research – considering multifunctional woody polycultures and the design of long-term field trials. Agrofor Syst 92(5):1397–1415. https://doi.org/10.1007/s10457–017–0087–4
MacDonald WL, Double ML (2006) Hypovirulence: use and limitations as a chestnut blight biological control. In: Steiner KC, Carlson JE (eds) Restoration of American chestnut to forest lands – proceedings of a conference and workshop, May 4–6 2004, North Carolina Arboretum, Asheville, NC. Natural resources report NPS/NCR/CUE/NRR – 2006/001, National Park Service, Washington, DC, pp 87–95
Martin MA, Alvarez JB, Mattioni C et al (2009) Identification and characterization of traditional chestnut varieties of southern Spain using morphological and simple sequence repeat (SSRs) markers. Ann Appl Biol 154:389–398
Mattioni C, Cherubini M, Micheli E et al (2008) Role of domestication in shaping Castanea sativa genetic variation in Europe. Tree Genet Genomes 4:563–574. https://doi.org/10.1007/s11295–008–0132–6
Mattioni C, Martin MA, Villani F (2013) Microsatellite markers reveal a strong geographical structure in European populations of Castanea sativa (Fagaceae): evidence for multiple glacial refugia. Am J Bot 100(5):951–961
McKay JW (1942) Self-sterility in the Chinese chestnut (Castanea mollissima). Proc Am Soc Hortic Sci 4:156–160
McWilliams W, Lister T, LaPoint E et al (2006) Current status of chestnut in eastern US forests. In: Steiner KC, Carlson JE (eds) Restoration of American chestnut to forest lands – proceedings of a conference and workshop, May 4–6 2004, North Carolina Arboretum, Ashville, NC. Natural resources report NPS/NCR/CUE/NRR – 2006/001, National Park Service, Washington, DC, pp 31–39
Mellano MG, Beccaro GL, Donno D et al (2012) Castanea spp. biodiversity conservation: collection and characterization of the genetic diversity of an endangered species. Genet Resour Crop Evol 59:1727–1741
Mellano MG, Torello-Marinoni D, Boccacci P et al (2018) Ex situ conservation and characterization of the genetic diversity of Castanea spp. Acta Hort 1220:1–6. https://doi.org/10.17660/ActaHortic.2018.1220.1
Merkle SA, Wiecko AT, Watson-Pauley BA (1991) Somatic embryogenesis in American chestnut. Can J For Res 21:1698–1701
Metaxas AM (2013) Chestnut (Castanea spp.) cultivar evaluation for commercial chestnut production in Hamilton County, Tennessee. MS thesis, University of Tennessee at Chattanooga. https://scholar.utc.edu/theses/339
Miller G, Miller DD, Jaynes RA (1996) Chestnuts. In: Janick J, Moore JN (eds) Fruit breeding, vol III. Nuts. John Wiley, New York, pp 99–123
Monteiro-Vitorello CB, Baidyaroy D, Bell J et al (2000) A circular mitochondrial plasmid incites hypovirulence in some strains of Cryphonectria parasitica. Curr Genet 37:242–256
Moreno-Cortes A, Hernandez-Verdeja T, Sanchez-Jimenez P et al (2012) CsRAV1 induces sylleptic branching in hybrid poplar. New Phytol 194:83–90. https://doi.org/10.1111/j.1469-8137.2011.04023.x
Nave JM (1998) Large-fruited chestnuts grown in North America. Ann Rep North Nut Grow Assoc 89:42–82
Newhouse AE, Polin-McGuigan LD, Baier KA et al (2014) Transgenic American chestnuts show enhanced blight resistance and transmit the trait to T1 progeny. Plant Sci 228:88–97
Nishio S, Yamamoto T, Terakami S et al (2011a) Novel genomic and EST-derived SSR markers in Japanese chestnuts. Sci Hortic 130:838–846
Nishio S, Yamamoto T, Terakami S et al (2011b) Genetic diversity of Japanese chestnut cultivars assessed by SSR markers. Breed Sci 61:109–120. https://doi.org/10.1270/jsbbs.61.109
Nishio S, Takada N, Yamamoto T et al (2013) Mapping and pedigree analysis of the gene that controls the easy peel pellicle trait in Japanese chestnut (Castanea crenata Sieb. et Zucc.). Tree Genet Genom 9:723–730
Nishio S, Iketani H, Fujii H et al (2014a) Use of population structure and parentage analyses to elucidate the spread of native cultivars of Japanese chestnut. Tree Genet Genom 10:1171–1180. https://doi.org/10.1007/s11295–014–0751–z
Nishio S, Yamada M, Takada N et al (2014b) Environmental variance and broad-sense heritability of nut traits in Japanese chestnut breeding. HortSci 49:696–700
Nishio S, Hayashi T, Yamamoto T et al (2018a) Bayesian genome-wide association study of nut traits in Japanese chestnut. Mol Breed 38:99. https://doi.org/10.1007/s11032–018–0857–3
Nishio S, Terakami S, Matsumoto T et al (2018b) Identification of QTLs for agronomic traits in the Japanese chestnut (Castanea crenata Sieb. et Zucc.) breeding. Hortic J 87(1):43–54. https://doi.org/10.2503/hortj.OKD–093
Nishio S, Takada N, Terakami S et al (2019) Estimation of effective pollen dispersal distance for cross-pollination in chestnut orchards by microsatellite-based paternity analyses. Sci Hortic 250:89–93
Nixon KC (1997) Fagaceae Dumortier, beech family. In: Flora of North America Editorial Committee (ed) Flora of North America north of Mexico, vol 3. Oxford University Press, New York, pp 436–437
Nugnes F, Gualtieri L, Bonsignore CP et al (2018) Resistance of a local ecotype of Castanea sativa to Dryocosmus kuriphilus (Hymenoptera: Cynipidae) in southern Italy. Forests 9(2):94. https://doi.org/10.3390/f9020094
Oak S (2002) From the Bronx to Birmingham: impact of chestnut blight and management practices on forest health risks in the southern Appalachian Mountains. J Am Chestnut Found 16(1):32–41
Oak S (2006) Forest health impacts of the loss of American chestnut. In: Steiner KC, Carlson JE (eds) Restoration of American chestnut to forest lands – proceedings of a conference and workshop, May 4–6 2004, North Carolina Arboretum, Asheville, NC. Natural resources report NPS/NCR/CUE/NRR – 2006/001, National Park Service, Washington, DC, pp 21–29
Oho N, Shimura I (1970) Process of study on Dryocosmus kuriphilus Yasumatsu (Hymenoptera: Cynipidae) and several problems about recent infestation. Shokubutsu Boeki (Plant Prot) 24:421–427
Omura M, Akihama T (1980) Male sterility in chestnuts: a tentative plan for the seed propagation of fruit trees. Gamma Field Symp 19:77–89
Paillet FL (1993) Growth form and life histories of American chestnut and Allegheny and Ozark chinkapin at various North American sites. Bull Torrey Bot Club 120(3):257–268
Parker GG, Hill SM, Kuehnel LA (1993) Decline of understory American chestnut [Castanea dentata (Marsh.) Borkh.] in a southern Appalachian forest. Can J For Res 23(2):259–265
Payne JA, Jaynes RA, Kays SJ (1983) Chinese chestnut production in the United States: practice, problems and possible solutions. Econ Bot 37:187–200
Peever TL, Liu YC, Milgroom MG (1997) Diversity of hypoviruses and other double-stranded RNAs in Cryphonectria parasitica in North America. Phytopathology 87:1026–1033
Pereira-Lorenzo S, Diaz-Hernandez MB, Ramos-Cabrer AM (2006a) Use of highly discriminating morphological characters and isozymes in the study of Spanish chestnut cultivars. J Am Soc Hortic Sci 131(6):770–779
Pereira-Lorenzo S, Ramos-Cabrer AM, Diaz-Hernandez MB et al (2006b) Chemical composition of chestnut cultivars from Spain. Sci Hortic 107:306–314
Pereira-Lorenzo S, Diaz-Hernandez MB, Ramos-Cabrer AM (2009a) Spain. In: Avanzato D, Bounous G (eds) Following chestnut footprints (Castanea spp.). Scrip Hortic 9:134–142
Pereira-Lorenzo S, Rios-Mesa D, Gonzalez-Diaz AJ, Ramos-Cabrer AM (2009b) Spain – Canary Islands. In: Avanzato D, Bounous G (eds) Following chestnut footprints (Castanea spp.). Scrip Hortic 9:143–148
Pereira-Lorenzo S, Costa R, Ramos-Cabrer AM et al (2010) Variation in grafted European chestnut and hybrids by microsatellites reveals two main origins in the Iberian Peninsula. Tree Genet Genomes 6:701–715. https://doi.org/10.1007/s11295–010–0285–y
Pereira-Lorenzo S, Costa R, Ramos-Cabrer AM et al (2011) Chestnut cultivar diversification process in the Iberian Peninsula, Canary Islands, and Azores. Genome 54:301–315. https://doi.org/10.1139/G10–122
Pereira-Lorenzo S, Ballester A, Corredoira E et al (2012) Chestnut. In: Badenes ML, Byrne D (eds) Fruit breeding/Springer series. Handbook of plant breeding. Springer, New York, pp 729–769
Pereira-Lorenzo S, Costa R, Anagnostakis S et al (2016) Interspecific hybridization of chestnut. In: Mason AS (ed) Polyploidy and hybridization for crop improvement. CRC Press, Boca Raton, pp 377–407
Pereira-Lorenzo S, Ramos-Cabrer AM, Barreneche T et al (2018) Instant domestication process of European chestnut cultivars. Ann Appl Biol 174:74–85. https://doi.org/10.1111/aab.12474
Perkins, MT (2016) Chloroplast DNA phylogenetics of the North American chestnuts and chinquapins (Castanea Mill., Fagaceae). MS thesis, University of Tennessee at Chattanooga. https://scholar.utc.edu/theses/485
Perkins MT, Robinson AC, Cipollini ML, Craddock JH (2019) Identifying host resistance to Phytophthora cinnamomi in hybrid progeny of Castanea dentata and Castanea mollissima. HortSci 54(2):221–225. https://doi.org/10.21273/HORTSCI13657–18
Phelps T, Steiner KC, Chen CC, Zaczek J (2006) Planting trials of American chestnut in central Appalachian forests. In: Steiner KC, Carlson JE (eds) Restoration of American chestnut to forest lands- proceedings of a conference and workshop, May 4–6 2004, North Carolina Arboretum, Asheville, NC. Natural resources report NPS/NCR/CUE/NRR – 2006/001, National Park Service, Washington, DC, pp 161–166
Phillips SJ, Anderson RP, Schapire RE (2006) Maximum entropy modeling of species geographic distributions. Ecol Model 190:231–259
Pitte JR (1986) Hommes et paysages du châtaignier de l’antiquité à nos jours. Fayard, Paris
Polin LD, Liang H, Rothrock RE et al (2006) Agrobacterium-mediated transformation of American chestnut [Castanea dentata (Marsh.) Borkh.] somatic embryos. Plant Cell Tissue Organ Cult 84:69–79
Pridnya MV, Cherpakov VV, Paillet FL (1996) Ecology and pathology of European chestnut (Castanea sativa) in the deciduous forests of the Caucasus Mountains in southern Russia. Bull Torrey Bot Club 123:213–222
Qin L, Feng Y (2009) China. In: Avanzato D, Bounous G (eds) Following chestnut footprints (Castanea spp.). Scrip Hortic 9:36–39
Quacchia A, Moriya S, Bosio G et al (2008) Rearing, release and settlement prospect in Italy of Torymus sinensis, the biological control agent of the chestnut gall wasp Dryocosmus kuriphilus. BioControl 53:829–839. https://doi.org/10.1007/s10526–007–9139–4
Regione Campania (2015) Castagna di Montella IGP. http://wwwagricolturaregionecampaniait. Accessed 28 Mar 2019
Roane MK, Griffin GJ, Elkins JR (1986) Chestnut blight, other Endothia diseases, and the genus Endothia. American Phytopathological Society, St. Paul
Robin C, Heiniger U (2001) Chestnut blight in Europe: diversity of Cryphonectria parasitica, hypovirulence and biocontrol. For Snow Landsc Res 76(3):361–367
Robin C, Morel O, Vettraino AM et al (2006) Genetic variation in susceptibility to Phytophthora cambivora in European chestnut (Castanea sativa). For Ecol Manag 226:199–207. https://doi.org/10.1016/j.foreco.2006.01.035
Rutter PA, Miller G, Payne JA (1991) Chestnuts (Castanea). Acta Hortic 290:761–788
Saito T (2009) Japan. In: Avanzato D, Bounous G (eds) Following chestnut footprints (Castanea spp.). Scrip Hortic 9:96–99
Samman SN, Thor E (1975) Breeding for resistance to chestnut blight at the University of Tennessee. Proceedings of the 23rd northeastern forest tree improvement conference, 4–7 Aug 1975, New Brunswick
Santos C, Machado H, Correia I et al (2015a) Phenotyping Castanea hybrids for Phytophthora cinnamomi resistance. Plant Pathol 64:901–910. https://doi.org/10.1111/ppa.12313
Santos C, Zhebentyayeva T, Serrazina S et al (2015b) Development and characterization of EST-SSR markers for mapping reaction to Phytophthora cinnamomi in Castanea spp. Sci Hortic 194:181–187
Santos C, Duarte S, Tedesco S et al (2017a) Expression profiling of Castanea genes during resistant and susceptible interactions with the oomycete pathogen Phytophthora cinnamomi reveal possible mechanisms of immunity. Front Plant Sci 8:515. https://doi.org/10.3389/fpls.2017.00515
Santos C, Nelson CD, Zhebentyayeva T et al (2017b) First interspecific genetic linkage map for Castanea sativa x Castanea crenata revealed QTLs for resistance to Phytophthora cinnamomi. PLoS One 12(9). https://doi.org/10.1371/journal.pone.0184381
Sartor C, Dini F, Torello Marinoni D et al (2015) Impact of the Asian wasp Dryocosmus kuriphilus (Yasumatsu) on cultivated chestnut: yield loss and cultivar susceptibility. Sci Hortic 197:454–460
Senter SD, Payne JA, Miller G, Anagnostakis SL (1994) Comparison of total lipids, fatty acids, sugars, and non-volatile organic acids in nuts from four Castanea species. J Sci Food Agric 65:223–227
Serrazina S, Santos C, Machado H et al (2015) Castanea root transcriptome in response to Phytophthora cinnamomi challenge. Tree Genet Genomes 11:6. https://doi.org/10.1007/s11295–014–0829–7
Shaw J, Craddock JH, Binkley MA (2012) Phylogeny and phylogeography of North American Castanea Mill. (Fagaceae) using cpDNA suggests gene sharing in the southern Appalachians (Castanea Mill., Fagaceae). Castanea 77:186–211
Shi Y, Hebard FV (1997) Male sterility in the progeny derived from hybridization between Castanea dentata and C. mollissima. J Am Chestnut Found 11(1):38–47
Shimura I (1972) Breeding of chestnut varieties resistant to chestnut gall wasp, Dryocosmus kuriphilus Yamatsu. Japan Agric Res Q 6(4):224–230
Shoda M, Takada N, Saito T et al (2006) A method for quickly removing pellicles from chestnuts by deep frying cooking oil. Bull Natl Inst Fruit Tree Sci 5:21–27
Singleton WR, Dietz A (1974) Use of mutation breeding to revive the American chestnut. Va J Sci 24:133
Sisco PH, Kubisiak TL, Casasoli M et al (2005) An improved genetic map for Castanea mollissima/Castanea dentata and its relationship to the genetic map of Castanea sativa. Acta Hortic 693:491–496
Sisco PH, Neel TC, Hebard FV et al (2014) Cytoplasmic male sterility in interspecific hybrids between American and Asian Castanea species is correlated with the American D chloroplast haplotype. Acta Hortic 1019:215–222
Smith JR (1950) Tree crops: a permanent agriculture. Devin-Adair Company, New York
Smith DM (2000) American chestnut: ill-fated monarch of the eastern hardwood forest. J For 98(2):12–15. https://doi.org/10.1093/jof/98.2.12
Southgate EWB (2006) Historical ecology of American chestnut (Castanea dentata). In: Steiner KC, Carlson JE (eds) Restoration of American chestnut to forest lands – proceedings of a conference and workshop, May 4–6 2004, North Carolina Arboretum, Asheville, NC. Natural resources report NPS/NCR/CUE/NRR – 2006/001, National Park Service, Washington, DC, pp 13–19
Soylu A (1992) Heredity of male sterility in some chestnut cultivars (Castanea sativa Mill.). Acta Hortic 317:181–186. https://doi.org/10.17660/ActaHortic.1992.317.21
Soylu A, Serdar U, Ertan E, Mert C (2009) Turkey. In: Avanzato D, Bounous G (eds) Following chestnut footprints (Castanea spp.). Scrip Hortic 9:155–159
Squatriti P (2013) Landscape and change in early medieval Italy: chestnuts, economy, and culture. Cambridge University Press, Cambridge, UK
Staton M, Zhebentyayeva T, Olukolu B et al (2015) Substantial genome synteny preservation among woody angiosperm species: comparative genomics of Chinese chestnut (Castanea mollissima) and plant reference genomes. BMC Genomics 16:744. https://doi.org/10.1186/s12864–015–1942–1
Steiner KC, Westbrook JW, Hebard FV et al (2017) Rescue of American chestnut with extraspecific genes following its destruction by a naturalized pathogen. New For 48:317–336. https://doi.org/10.1007/s11056–016–9561–5
Stephenson SL, Adams HS, Lipford ML (1991) The present distribution of chestnut in the upland forest communities of Virginia. Bull Torrey Bot Club 118(1):24–32
Takada N, Sawamura Y, Nishio S, Saito T (2010) Influence of pollen parents on removability of pellicle and fruit weight of chestnut cultivar ‘Porotan’. Acta Hortic 866:239–242
Takada T, Nishio S, Yamada M et al (2012) Inheritance of the easy-peeling pellicle trait of Japanese chestnut cultivar ‘Porotan’. HortSci 47(7):845–847
Takada N, Yamada M, Nishio S et al (2019) Existence of genetic differences in pellicle peelability in Japanese chestnut (Castanea crenata Sieb. et Zucc.) cultivars and selections with difficult-peeling pellicles. Hortic J 86(4):456–462. https://doi.org/10.2503/hortj.OKD–030
Thomas AL, Avery JD, Byers PL et al (2015) Ozark chinkapin demonstrates compatibility as a scion grafted to Chinese chestnut rootstocks: implications for ex situ conservation. Native Plant J 16(2):117–124
Tuskan GA, Groover AT, Schmutz J et al (2018) Hardwood tree genomics: unlocking woody plant biology. Front Plant Sci 9:1799. https://doi.org/10.3389/fpls.2018.01799
Van Fleet W (1914) Chestnut breeding experience: how the American chestnut, threatened by extinction, may be saved to horticulture, even if not to forestry – promising crosses between chinquapin and Asiatic species – trees bear early, produce good nuts and show resistance to blight. J Hered 5(1):19–25. https://doi.org/10.1093/jhered/5.1.19
Vettraino AM, Morel O, Perlerou C et al (2005) Occurrence and distribution of Phytophthora species in European chestnut stands, and their association with ink disease and crown decline. Eur J Plant Pathol 111:169–180
Viéitez Cortizo E, Viéitez Madriñán ML, Viéitez Madriñán FJ (1996) El castaño. Edilesa, Caixa Ourense
Villani F, Mattioni C, Cherubini M et al (2009) An integrated approach to assess the genetic and adaptive variation in Castanea sativa Mill. Acta Hortic 866:91–95. https://doi.org/10.17660/ActaHortic.2010.866.8
Wang XY, Huang XL, Jiang LY, Qiao GX (2010) Predicting potential distribution of chestnut phylloxerid (Hemiptera: Phylloxeridae) based on GARP and Maxent ecological niche models. J Appl Entomol 134:45–54. https://doi.org/10.1111/j.1439-0418.2009.01447.x
Wang GG, Knapp BO, Clark SL, Mudder BT (2013) The silvics of Castanea dentata (Marsh.) Borkh., American chestnut, Fagaceae (beech family). General technical report SRS-173. USDA Forest Service, Southern Research Station, Asheville
Westbrook JW (2017) Genomic selection for disease resistance in American chestnut backcross populations. Chestnut J Am Chestnut Found 31(3):38–42
Westbrook JW (2018a) A plan to diversify a transgenic blight-tolerant American chestnut population. Chestnut J Am Chestnut Found 32(3):31–36
Westbrook JW (2018b) Merging backcross breeding and transgenic blight resistance to accelerate restoration of the American chestnut: the American Chestnut Foundation’s breeding and selection plan 2015–2025. https://www.acf.org/wp-content/uploads/2018/03/TACF_2015-2025_BreedingSciencePlan_updated2_9_18.pdf. Accessed 16 Feb 2019
Westbrook JW, Zhang Q, Mandal MK et al (2019) Genomic selection for blight–tolerance in American chestnut (Castanea dentata) backcross populations. Evol Appl. In press
Wheeler N, Sederoff R (2009) Role of genomics in the potential restoration of the American chestnut. Tree Genet Genomes 5:181–187. https://doi.org/10.1007/s11295–008–0180–y
Worthen LM, Woeste KE, Michler CH (2010) Breeding American chestnuts for blight resistance. In: Janick J (ed) Plant breeding reviews, vol 33. Wiley-Blackwell, Hoboken, pp 305–339
Xing Z, Powell WA, Maynard CA (1999) Development and germination of American chestnut somatic embryos. Plant Cell Tissue Organ Cult 57(1):47–55
Yang F, Liu Q, Pan S et al (2015) Chemical composition and quality traits of Chinese chestnuts (Castanea mollissima) produced in different ecological regions. Food Biosci 11:33–42
Zhang DX, Nuss DL (2016) Engineering super mycovirus donor strains of chestnut blight fungus by systematic disruption of multilocus vic genes. Proc Natl Acad Sci USA 113:2062–2067
Zhang B, Oakes AD, Newhouse AE et al (2013) A threshold level of oxalate oxidase transgene expression reduces Cryphonectria parasitica-induced necrosis in a transgenic American chestnut (Castanea dentata) leaf bioassay. Transgenic Res 22:973–982. https://doi.org/10.1007/s11248–013–9708–5
Zhebentyayeva T, Chandra A, Abbott AG et al (2014) Genetic and genomic resources for mapping resistance to Phytophthora cinnamomi in chestnut. Acta Hortic 1019:263–270
Zhu C, Shi F, Chen Y et al (2019a) Transcriptome analysis of Chinesec (Castanea mollissima Blume) in response to Dryocosmus kuriphilus Yasumatsu infestation. Int J Mol Sci 20(4):855. https://doi.org/10.3390/ijms20040855
Zhu C, Shi F, Wang M et al (2019b) The complete chloroplast genome of a variety of Castanea mollissima ‘Hongli’ (Fagaceae). Mitochondrial DNA B Resour 4(1):993–994. https://doi.org/10.1080/23802359.2019.1580160
Zulfiqar S, Gao X-X, Sun Y-W et al (2019) The complete chloroplast genome sequence of Seguin chestnut (Castanea seguinii). Mitochond DNA B Resour 4(1):342–343. https://doi.org/10.1080/23802359.2018.1544047
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Appendices
Appendices
1.1 Appendix I: Research Institutes Relevant to Chestnut
Institute | Specialization and research activities | Contact information, including website |
|---|---|---|
The American Chestnut Foundation, Asheville, NC, USA | American chestnut restoration | Dr. Jared Westbrook email: chestnut@acf.org website: https://www.acf.org/ |
American Chestnut Research and Restoration Project, SUNY College of Environmental Science and Forestry, Forest Biotechnology Working Group Syracuse, NY, USA | Basic and applied research that will lead to the development of a blight-resistant American chestnut tree. Perform cutting edge research that will enhance our understanding of forest tree biology and lead to improved productivity and biodiversity of our forested ecosystems | Dr. William A. Powell email: wapowell@esf.edu website: https://www.esf.edu/chestnut/ |
Beijing Advanced Innovation Center for Tree Breeding by Molecular Design Beijing University of Agriculture Beijing, China | Genetics and molecular breeding of tree crops | Dr. Ling Qin email: qinlingbac@126.com |
Centre for Research and Technology of Agro-Environment and Biological Sciences University of Trás-os-Montes e Alto Douro Vila Real, Portugal | Chestnut physiology, abiotic and biotic stresses, and breeding | Dr. José Gomes-Laranjo email: jlaranjo@utad.pt |
Chattanooga Chestnut Project, University of Tennessee at Chattanooga, TN, USA | Cultivar development, American chestnut restoration, genetic diversity and evolution of North American Castanea species | Dr. J. Hill Craddock email: hill-craddock@utc.edu website: https://www.utc.edu/biology-geology-environmental-science/profiles/faculty/wmn758.php |
Chestnut Research and Development Centre Chiusa Pesio, Italy | Research on several topics related to chestnut cultivation, including chestnut germplasm, advanced propagation techniques, optimization of cultural practices, pests and diseases management, nursery and extension services | Prof. Gabriele Beccaro email: centro.castanicoltura@unito.it web: |
Dept. of Biological Sciences Mississippi State University Starkville, MS, USA | Molecular interactions of plant pathogenic fungi and their hosts, particularly between chestnut blight fungus and the American chestnut | Dr. Angus Dawe email: dawe@biology.msstate.edu website: https://www.biology.msstate.edu/people/staff.php?id=ald662 |
Dept. of Biological Sciences University of Notre Dame Notre Dame, IN, USA | Population dynamics, spatial genetic structure, gene flow, adaptive genetic variation and spontaneous interspecific hybridization | Dr. Jeanne Romero-Severson email: jromeros@nd.edu website: https://biology.nd.edu/people/jeanne-romero-severson/ |
Department of Crop Production and Engineering Projects High Polytechnic School Lugo Campus University of Santiago de CompostelaLugo Spain | Research in chestnut with morphological and molecular markers, genetic diversity evaluation, identification and catalogue of local cultivars, | Dr. Santiago Pereira-Lorenzo email: santiago.pereira.lorenzo@usc.es website: http://www.usc.es/en/departamentos/prodvexg/profesores/pereiralorenzosantiagopublicacion |
Dept. of Plant and Environmental Sciences Clemson University Clemson, SC, USA | Diseases caused by Phytophthora spp. and rust diseases, with current work on root rot of chestnut | Dr. Steven Jeffers email: sjffrs@clemson.edu website: https://www.clemson.edu/cafls/faculty_staff/profiles/sjffrs |
Dept. of Plant Biology Michigan State University East Lansing, MI, USA | Interactions between American chestnut, its blight pathogen, and intracellular hyperparasites that alter pathogen virulence | Dr. Andrew M. Jarosz email: amjarosz@msu.edu website: https://plantbiology.natsci.msu.edu/directory/andrew-m-jarosz/ |
Division of Plant and Soil Sciences West Virginia University Morgantown, WV, USA | Biological control of chestnut blight using transmissible hypovirulence | Dr. William L. MacDonald email: macd@wvu.edu website: https://www.davis.wvu.edu/faculty-staff/directory/william-macdonald |
Faculty of Agriculture Dept. of Horticulture Ondokuz Mayis University Samsun, Turkey | Evaluation of chestnut hybrids for cultivar and rootstock characteristics, postharvest treatment of chestnuts | Prof. Dr. Ümit Serdar email: userdar@omu.edu.tr |
Forest Health Research and Education Center University of Kentucky Lexington, KY, USA | Conservation and restoration of forested ecosystems using genetics-based biological research, social science research, and educational outreach on factors affecting tree and forest health | Dr. C. Dana Nelson email: dananelson@fs.fed.us |
Institute of Botany, Jiangsu Province and Chinese Academy of Sciences Nanjing, China | Genetics and breeding of C. mollissima | Dr. Guomin Geng, email: 13951970164@163.com |
ISHS Chestnut Working Group | Coordinates chestnut research worldwide | Chairman: Dr. José Gomes Laranjo email: jlaranjo@utad.pt |
NARO Institute of Fruit Tree and Tea Science Tsukuba, Japan | Cultivar development, production systems, integrated pest management, elucidation of fruit functionality, regulatory mechanisms governing fruit quality | Dr. Toshihiro Saito email: www.naro.affrc.go.jp |
Research Institute on Terrestrial Ecosystems Italian National Research Council Porano, Italy | Population genetics and genomics of C. sativa | Dr. Fiorella Villani email: fiorella.villani@ibaf.cnr.it website: https://www.cnr.it/en/institute/125/research-institute-on-terrestrial-ecosystems-iret |
The Schatz Center for Tree Molecular Genetics Pennsylvania State University University Park, PA, USA | Research and training in forest genetics related projects in the areas of genomics, conservation genetics, and biotechnology | Dr. John Carlson email: jec16@psu.edu |
Scientific-Research Centre of Agriculture Tbilisi, Georgia | In situ and ex situ management of C. sativa genetic resources in Georgia | Dr. Zviad Bobokashvili email: bobokashvili@hotmail.com website: http://srca.gov.ge/en |
Shandong Provincial Key Laboratory of Fruit Tree Biotechnology Breeding, Shandong Institute of Pomology Tai’an, China | Genetic diversity and breeding of C. mollissima , home of the national chestnut germplasm repository of China | Dr. Qingzhong Liu email: qzliu@sdip.cn |
Sistemas Agrários e Florestais e Sanidade Vegetal Instituto Nacional de Investigação Agrária e Veterinária Lisbon, Portugal | Forest genetics, molecular biology and genomics of woody species including C. sativa, and understanding resistance of Castanea spp. to Phytophthora | Dr. Rita Lourenço Costa email: rita.lcosta@iniav.pt |
South China Botanical Garden Chinese Academy of Sciences Guangzhou, China | Plant ex situ conservation and inventory, plant conservation genetics | Dr. Hongwen Huang email: huanghw@mail.scbg.ac.cn website: http://english.scbg.cas.cn/ |
Southern Institute of Forest Genetics, US Forest Service Saucier, MS, USA | Genetics/genomics of disease resistance, genetic diversity of American chestnut | Dr. C. Dana Nelson email: dananelson@fs.fed.us |
Swiss Federal Institute for Forest, Snow and Landscape Research WSL Cadenazzo, Switzerland | WSL explores the dynamics of the terrestrial environment, and the use and protection of natural habitats and cultural landscapes. | Dr. Marco Conederaemail: marco.conedera@wsl.ch |
Tree Genetics and Breeding Lab, Aristotle University of Thessaloniki Thessaloniki, Greece | Genetics and epigenetics of C. sativa, breeding and micropropagation of C. sativa, | Prof. F.A. Aravanopoulos, email: aravanop@for.auth.gr |
Unité Mixte de Recherche Biodiversité, des Gènes aux Communautés Institut National de la Recherche Agronomique Bordeaux, France | Genetic determinism of diseases of chestnut; interactions between gall wasp, C. parasitica and endophytic fungi; evolution of C. parasitica and its hypovirus | Dr. Cécile Robin email: robin@bordeaux.inra.fr https://www6.bordeaux-aquitaine.inra.fr/biogeco_eng/Staff/Staff-directory/Q-Z/Robin-Cecile |
USDA Forest Service Hardwood Tree Improvement and Regeneration Center, Department of Forestry and Natural Resources Purdue University West Lafayette, IN, USA | Genetics of forest trees including development and use of DNA markers and sequence data to study genetic variability, evolution and spatial distribution of variability, breeding trees for plantations and species restoration | Dr. Keith Woeste, email: woeste@purdue.edu website: https://htirc.org/ |
1.2 Appendix II: Chestnut Genetic Resources
Information is presented in Table 5.2.
Rights and permissions
Copyright information
© 2019 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Craddock, J.H., Perkins, M.T. (2019). Chestnut (Castanea spp. Miller) Breeding. In: Al-Khayri, J., Jain, S., Johnson, D. (eds) Advances in Plant Breeding Strategies: Nut and Beverage Crops. Springer, Cham. https://doi.org/10.1007/978-3-030-23112-5_5
Download citation
DOI: https://doi.org/10.1007/978-3-030-23112-5_5
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-23111-8
Online ISBN: 978-3-030-23112-5
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)