
Abstract
The true distribution of migrant species is rarely immediately apparent from the distribution of ring recoveries due to a heavy bias in regional recovery probabilities. For western Palearctic species, the recovery probability is especially low in Africa, but also varies within Europe. However, little work has been done to derive actual estimates of these recovery probabilities needed to infer the “true” underlying distribution. Here, we investigate the potential of using ringing data to estimate the seasonal distribution densities of migrant species. Using likelihoods based on a two point mixture distribution, the proportions of individuals wintering south of the Sahara are estimated using differences in recovery distributions among species in species groups where the location-specific probability of a ring recovery can be assumed to be essentially the same among species. We consider two such approaches. In the first, survival associated with a wintering area must be set constant across species. In the second, we assume the time series is long enough that a single binary response (recovered/not recovered) may be modeled independently of survival parameters. Under the first approach, we estimated the proportion of sub-Saharan migrants, together with 95% profile likelihood confidence intervals, for redstart as 0.84 [0.70,0.93], thrush nightingale 1.00 [0.49,1.00], garden warbler 0.95 [0.85;0.99], blackcap 0.60 [0.32;0.78], reed warbler 0.87 [0.72,0.95], and pied flycatcher 0.90 [0.76;0.97] using recovery data for birds ringed in Denmark and assuming that all robins winter north of Sahara. In the second approach, estimated proportions of sub-Saharan migrants were similar, but the confidence intervals were somewhat narrower. Although further work is required to examine the underlying assumptions, the models and analyses presented here provide a framework for making better use of existing ring recovery datasets to understand the “true” seasonal distribution patterns of European birds.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
1 Introduction
Birds are among the most mobile of all terrestrial organisms, and a few species travel up to more than 20 000 km annually between separate breeding and wintering areas. The task of defining the seasonal distribution of migratory species is considerably more complicated than for sedentary species. The recent demand for such data to assess risk of spread of diseases such as avian influenza (Delany et al. 2006) or effects of accelerating climate change on species of conservation concern (Walther et al. 2004) has created an urgent need to improve our ability to estimate the seasonal distribution and connectivity of migratory species.
From direct observations of birds in the field, we have acquired a general knowledge about seasonal species distributions (e.g. Cramp 1988). While different modeling approaches have been used to help explain variation in these observations (e.g. using climate variables, Walther et al. 2004), such approaches tell us little about the connectivity between populations and areas (Webster et al. 2002). While sophisticated approaches for tracking birds over long distances have been proposed using radio telemetry (Wikelski et al. 2007), these are not yet practical for most small passerines. As such, the most suitable available data bases for identifying connectivity are generally those based on ring recoveries. In Europe, very large ring recovery data bases exist. In Denmark alone, more than 4 million birds have been ringed and more than 180 000 of these have been recovered (Bønløkke et al. 2006). However, most analyses of these data to date have been qualitative or semi-quantitative (e.g. Zink 1973–1985, Zink and Bairlein 1995).
One obstacle to using such data for inference is the fact that spatial distributions of recoveries may provide biased estimates of the spatial distribution of birds due to differences in the regional recovery probabilities (Crissey 1955; Perdeck 1977). For western Palearctic bird migrants, the recovery probability is especially low in Africa, but it is also low in East Europe compared to West Europe. Because of difficulties in addressing spatial variation in recovery and survival probabilities, most analyses of ring recoveries from a larger spatial scale have not attempted any quantitative analysis and only presented the raw data for inferences on seasonal distributions (Bakken et al. 2003; Bønløkke et al. 2006; Fransson and Pettersson 2001; Wernham et al. 2002; Fig. 1; though see e.g. Kania 1981).

Typical presentation of ring recovery data in national recovery atlases. Here garden warblers ringed in Denmark (from Bønløkke et al. 2006)
A number of studies have investigated movements using multi-site (or multi-state) capture-recapture methods. However, this modeling approach has seen only a few applications for estimating species densities/distributions. A few single-species studies have estimated seasonal distributions. For example, Sibert et al. (1999) used an advection-diffusion model to investigate skipjack tuna movement. Similarly, Skalski et al. (2002) estimated route-specific passage of smolt, in a framework similar to the migration system considered here. However, they used radio telemetry and the robust design (Pollock 1982; Kendall et al. 1995) at certain points to estimate detectability.
Ideally, we would like to model both the spatio-temporal distribution of recoveries as well as variation in survival probabilities for each population and age class. Apart from a general lack of extensive data sets necessary to estimate these many parameters, there is a general problem of parameter identifiability for such movement models. For example, Kendall et al. (2006) found that parameters are only identifiable if the number of “marking” areas with slightly different migration probabilities is greater or equal to the number of “recovery” areas.
Here, we present an approach for using ringing data to estimate the relative seasonal distribution (in winter) of migrant species based on a few simplifying assumptions. The rationale of this approach was already proposed by Busse and Kania (1977) and Kania and Busse (1987), who considered dividing the birds (of one species) into different groups. However, their model formulation only allowed for point estimation of parameters and thus statistical inference could not be based on their method.
The main focus will be estimating the species-specific proportions of songbirds wintering south of Sahara. In general, too little information is available in standard ringing data to separate survival, recovery and movement probabilities. However, standard ringing data generally includes many species, and the differences among species can provide the information necessary for estimating these parameters, if certain parameters such as survival and region-specific recovery probabilities can be treated as constant across species. In effect, this approach suffices to increase the number of “marking” areas (e.g., Kendall et al. 2006). We consider two methods for analyzing spatial ring recovery data. In the first, survival and ring recovery probabilities are assumed to be equal among species, but ring recovery probabilities are allowed to differ among regions. This approach utilizes traditional release-recovery arrays (e.g., Brownie et al. 1985). In the second method, the time series of recovery data are assumed to be long enough that survival is effectively zero (e.g., by censoring the last 15 years of ringing data), and only a binary response is modeled (recovered/not recovered). This second method requires that one throw out some of the data, but does not require the assumption that survival is equal among species. However, life-long ring reporting probabilities are still assumed to be constant across species within a region. Relaxation of the equal survival assumption in the second method will do little to alleviate bias if the reporting probabilities \(r\) differ between species. Because of the way in which \(r\) is defined, this can occur when sources of mortality within a region differ between species if each mortality source is associated with a different reporting rate. For both approaches, we restricted the species group considered to species where the variation among species in both survival and recovery probabilities are likely to be of smaller magnitude than the variation in recovery probabilities among regions.
2 Methods
We developed likelihoods based on a two point mixture distribution to estimate wintering distributions of songbirds. We assumed that individuals were ringed in the summer in Europe, and that the probability of capture for marking does not depend upon migration destination or individual attributes likely to affect survival or recovery probability. For simplicity, we consider the case where there are two wintering areas (A and B), and where interest focuses on estimating the proportion of birds that migrate to each area. We assume that an individual bird follows only one migration route; that is, an individual would not migrate to area A in one year and area B the next. As articulated in the introduction, we considered two possibilities for model construction.
B. Multinomial cell probabilities | |||||
|---|---|---|---|---|---|
Year of release | Group | Strata recovered | Year of recovery | ||
1 | 2 | 3 | |||
1 | 1 | A | \(\uppi_{11}^A f_1^A\) | \(\uppi_{11}^A S_1^A f_2^A\) | \(\uppi_{11}^A S_1^A S_2^A f_3^A\) |
1 | B | \((1-\uppi_{11}^A)f_1^B\) | \((1-\uppi_{11}^A)S_1^B f_2^B\) | \((1-\uppi_{11}^A)S_1^B S_2^B f_3^B\) | |
2 | 1 | A | \(\uppi_{12}^A f_2^A\) | \(\uppi_{12}^A S_{2}^A f_3^A\) | |
1 | B | \((1-\uppi_{12}^A)f_2^B\) | \((1-\uppi_{12}^A)S_2^B f_3^B\) | ||
3 | 1 | A | \(\uppi_{13}^A f_3^A\) | ||
1 | B | \((1-\uppi_{13}^A)f_3^B\) | |||
1 | 2 | A | \(\uppi_{21}^A f_1^A\) | \(\uppi_{21}^A S_1^A f_2^A\) | \(\uppi_{21}^A S_1^A S_2^A f_3^A\) |
2 | B | \((1-\uppi_{21}^A)f_1^B\) | \((1-\uppi_{21}^A)S_1^B f_2^B\) | \((1-\uppi_{21}^A)S_1^B S_2^B f_3^B\) | |
2 | 2 | A | \(\uppi_{22}^A f_2^A\) | \(\uppi_{22}^A S_{2}^A f_3^A\) | |
2 | B | \((1-\uppi_{22}^A)f_2^B\) | \((1-\uppi_{22}^A)S_2^B f_3^B\) | ||
3 | 2 | A | \(\uppi_{23}^A f_3^A\) | ||
2 | B | \((1-\uppi_{23}^A)f_3^B\) | |||
2.1 Likelihood 1: Survival and Recovery Probabilities Constant Across Species
If we assume that survival is constant across species that use similar migration routes, we can specify a multinomial model for the spatial and temporal distribution of ring recoveries (Table 1). The parameters of the model include \(\uppi_{gi}^A\), the probability that an individual of group \(g\), initially ringed in year \(i\), is a migrant to wintering area A; \(S_i^A\) and \(S_i^B\), the probability of annual survival from the time of ringing in year \(i\) for those birds migrating to wintering area A and B, respectively; and \(f_i^A\) and \(f_i^B\), the probabilities that individual migrants to areas A and B are recovered in \((i,\ i+1)\), given that they are alive at time \(i\). Groups in this case can include members of different species or different banding locations if only one species is under consideration. Whatever groups are considered, it is assumed that survival and recovery probabilities are the same for different groups.
Analytical and analytic-numeric (Burnham et al. 1987) methods were used to explore parameter identifiability with this model. As was found by Kendall et al. (2006) in the case of stochastic transitions to wintering areas, an argument based on the number of equations and number of unknowns suggested that parameters would only be identifiable if the number of groups (banding locations, number of species) were greater or equal to the number of wintering locations. Further, the proportion of individuals migrating to different wintering areas needs to differ among the groups under consideration; as a result, partitioning data from one species and banding location into two subsets will not remedy the parameter identifiability problem.
2.2 Likelihood 2: Long Term Survival Assumed to Be Zero
Eventually, all birds ringed will die. If ringing records are censored for a long enough duration at the end of the study (but recovery records are still compiled), one may eliminate one of the assumptions required by Model 1 by setting \(S_\ast^A=S_\ast^B=0\), where \(S_\ast\) is survival to the end of the study. Under this approach, each ringing event is accompanied by a binary response variable which equals 1 if the individual’s ring is recovered later and 0 if not. Under this approach, we ignore temporal variation in ring recovery probabilities, and write a likelihood for ringing and recovery data as
where \(r^A\) and \(r^B\) give time constant ring reporting probabilities for wintering areas A and B in the sense of Seber (1982), \(R_{gi}\) gives the number of individuals ringed and released in group \(g\) at time \(i,\ m_{gi}^A\) and \(m_{gi}^B\) give the number of these individuals later recovered in wintering areas A and B, respectively, and \(T\) gives the number of years that ringing data are modeled. Requirements for parameter identification are similar in this approach as they were with Likelihood 1.
2.3 Example Analyses
We coded the preceding likelihoods into program SURVIV (White 1983) to estimate species-specific differences in the proportions of birds wintering in Europe/North Africa and in sub-Saharan Africa, respectively, for 7 species of songbirds. We analyzed data on redstart Phoenicurus phoenicurus, thrush nightingale Luscinia luscinia, European robin Erithacus rubecula, reed warbler Acrocephalus scirpaceus, garden warbler Sylvia borin, blackcap Sylvia atricapilla and pied flycatcher Ficedula hypoleuca ringed in Denmark 1899–2002. These are all small passerines passing Denmark on migration in reasonable numbers. For these species, annual numbers ringed are known as well as detailed data on recoveries. For this analysis, only birds recovered dead were included. For the species included, live recaptures are rare and their associated probabilities are probably even more heterogeneous and have more complicated spatial and temporal variation than that for dead recoveries, as they depend on the distribution of ringers. For the species considered, most of the birds ringed are young birds but due to the low number of recoveries for several of the species, we pooled all age classes together.
All birds were considered to have been ringed in summer of year 0, as data sparseness precluded investigation of models with temporal variation in model parameters. The birds were primarily ringed in the migration seasons (spring and autumn), and very few were ringed in the winter (European robin only). The wintering period was defined for all species as December-February,, and no recoveries outside this season were included. For simplicity, we only modeled recoveries for a period of five years after ringing.
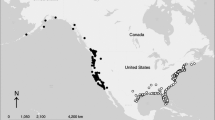
Each recovery was either in Europe/North Africa or sub-Saharan Africa (Fig. 2). Due to data sparseness, we set mixture, survival, and recovery probabilities constant over time, and constrained survival to be independent of wintering area. There were only four recoveries in sub-Saharan Africa, which rendered parameters non-estimable if mixture parameters were included for all species. Thus, we also set the probability of migration to sub-Saharan Africa to zero for the European robin (which does not occur in sub-Saharan Africa; Keith et al. 1992), allowing estimation of mixture parameters for other species. For each likelihood formulation, two models were fit to the data. In the first, recovery probabilities were allowed to differ between Europe/North Africa and sub-Saharan Africa. In the second, recovery probability was set equal between the two wintering areas.

Spatial distribution of ring recoveries of 7 species ringed in Denmark and recovered in winter (December–February)
3 Results
Under likelihood 1, the model with differences in recovery probability between Europe/North Africa and sub-Saharan Africa was strongly favored by \({\rm AIC}_{\rm c}\) (\(\Delta{\rm AIC}_{\rm c} = 150.8\) relative to the model with a constant recovery probability; see Burnham and Anderson 2002). Under this model, the estimated ring recovery probability for Europe/North Africa was 4.6E-04, with a 95% profile likelihood interval of [4.1E-04; 5.2E-04] and that in sub-Saharan Africa 1.1E-05 [4.4E-06; 2.6E-05], thus around 40 times lower in sub-Saharan Africa. The common estimate of annual survival probability was 0.34 [0.30; 0.38]. Estimated probabilities of migrating to sub-Saharan Africa ranged from 0.60 in the blackcap to 1.0 in thrush nightingale (Table 2). These estimates can be compared with each species’ actual recoveries from either region (Table 2 and Fig. 2).
Model selection criterion was similarly dismissive of the model with constant recovery probability when likelihood 2 was employed \((\Delta{\rm AIC}_{\rm c} = 178.6)\). In this case, the estimated cumulative probability of a ring being reported over the lifetime of an individual was 1.4E-03 (95% profile CI: 1.3E-03, 1.6E-03) for birds wintering in Europe and North Africa, and 2.8E-05 (95% profile CI: 0.9E-05, 6.4E-05) for birds wintering in sub-Saharan Africa. Estimated probabilities of wintering in sub-Saharan Africa were similar to those from likelihood 1 (Table 2).
4 Discussion
The estimates of recovery probabilities and movement probabilities were obtained using standard ringing data with only limited trapping information. Only numbers of birds ringed each year are known. This is a common limitation for many of the ringing data bases in Europe, in that full information has only been computerized for birds recovered later in standard ringing data, but annual ringing totals for each species can generally be obtained with reasonable effort. The two approaches produced similar estimated movement probabilities that are in general similar to what one would expect based on biology of the species, and, despite the wide confidence limits, useful as quantitative estimates. For all species included, we estimated that the majority of individuals wintered in sub-Saharan Africa. These species are known to winter in large numbers in sub-Saharan Africa and they are comparatively rare in Europe in winter (Cramp 1988, 1992).
The rather strong assumptions made here are of course likely to bias the parameter estimates. Assuming equal survival probabilities, finding and reporting, and recovery probabilities among species and within regions are clearly not realistic. For example, mortality is not likely to be equal among short- and long-distance migrants. However, the variation in these parameters is likely to be on a smaller scale than the variation in the estimated parameters. The small differences between the parameter estimates under the two approaches provides some indication that our assumption about equal survival probabilities across species is not seriously violated. However, a full scale sensitivity analysis would likely be needed to confirm this assertion. Larger differences in parameter estimates between the two approaches are likely to arise if using less similar species.
Several possibilities exist to improve the estimates. First of all, the data set could be extended to include data from more countries, as these should be readily available, e.g. through the EURING data bank (http://www.euring.org/edb/). Another reasonable extension would be to include more species (e.g. wildfowl to assess spread of avian influenza). In that case, it would be necessary to model differences among species in survival and regional recovery probabilities. As long as these could be considered constant within groups this should still be possible. Alternatively, they could be modeled as relative differences, i.e. the recovery probability at site B could be constrained to be a constant proportion of the probability at site A. Estimates of the regional recovery probabilities could be further improved by using the seasonal changes in distribution for each species, since constant recovery probability within regions through seasons is probably a reasonable assumption. This does, however, cause some trouble due to the fact that many ringing schemes have not computerized detailed ringing information. Sensitivity to this can be modeled, but hopefully our approach may give some additional incentive for ringing organizations to digitize historical ringing records.
Extending the data set to include more species and more fine-scaled regions (and even time-specific probabilities from 100 years of ringing) raises the question of how to deal with high dimensional parameter sets. A custom-made framework is probably necessary for this type of analysis since complicated models are not easily accommodated in SURVIV.
The parameter estimates presented here were conditional on highly constrained models and few recoveries. As such, they are certainly not definitive. However, the analysis should serve as a template for how this framework could be extended to analyze larger, richer datasets. Even basic science may benefit from quantitative distribution maps, where previous studies on e.g. the migratory orientation program have used qualitative patterns only (Mouritsen 1998; Thorup and Rahbek 2004).
References
Bønløkke J, Madsen JJ, Thorup K, Pedersen KT, Bjerrum M, Rahbek C (2006) The Danish Bird Migration Atlas. Rhodos, Humlebæk.
Bakken V, Runde O, Tjorve E (2003) Norwegian Bird Ringing Atlas. Stavanger, Stavanger Museum.
Brownie C, Anderson DR, Burnham KP, Robson DS (1985) Statistical Inference from Band-Recovery Data: A Handbook, 2nd edition. U. S. Fish and Wildlife Research Service, Resource Publication 156. U. S. Department of Interior, Washington D.C.
Burnham KP, Anderson DR (2002) Model Selection and Multimodel Inference: a Practical Information-Theoretic Approach, 2nd edn. Springer Verlag, New York.
Burnham KP, Anderson DR, White GC, Brownie C, Pollock KH (1987) Design and analysis for fish survival experiments based on release-recapture. American Fisheries Society Monograph 5. Bethesda, Maryland, USA.
Busse P, Kania W (1977) A quantitative estimation of distribution of ringed birds on the basis of recovery dispersal. Notatki Ornitologiczne 18:79–93. [Polish with English summary]
Cramp S (ed.) (1988) The Birds of the Western Palearctic. Vol. V. Oxford University Press, Oxford.
Cramp S (ed.) (1992) The Birds of the Western Palearctic. Vol. VI. Oxford University Press, Oxford.
Crissey WF (1955) The use of banding data in determining waterfowl migration and distribution. Journal of Wildlife Management 19:75–83.
Delany S, Veen J, Clarke J (Eds.) (2006) Urgent preliminary assessment of ornithological data relevant to the spread of Avian Influenza in Europe. Report to the European Commision.
Fransson T, Pettersson J (2001) Swedish Bird Ringing Atlas. Stockholm.
Kania W (1981) The autumn migration of the chaffinch Fringilla coelebs over the Baltic coast in Poland. Acta Ornithologica 18:371–414.
Kania W, Busse P (1987) An analysis of the recovery distribution based on finding probabilities. Acta Ornithologica 23:121–128.
Keith S, Urban EK, Fry CH (1992) The Birds of Africa. Vol. IV. Academic Press, London.
Kendall WL, Conn PB, Hines JE (2006) Combining multistate capture-recapture data with tag recoveries to estimate demographic parameters. Ecology 87:169–177.
Kendall WL, Pollock KH, Brownie C (1995) A likelihood-based approach to capture-recapture estimation of demographic parameters under the Robust Design. Biometrics 51:293–308.
Mouritsen H (1998) Modelling migration: the clock-and-compass model can explain the distribution of ringing recoveries. Animal Behaviour 56:899–907.
Perdeck AC (1977) The analysis of ringing data: Pitfalls and prospects. Vogelwarte 29 Sonderheft 33–44.
Pollock KH (1982) A capture-recapture design robust to unequal probability of capture. Journal of Wildlife Management 46:757–760.
Seber GAF (1982) The Estimation of Animal Abundance and Related Parameters, 2nd Edition. Griffin, London.
Sibert JR, Hampton J, Fournier DA, Bills PJ (1999) An advection-diffusion-reaction model for the estimation of fish movement parameters from tagging data, with application to skipjack tuna (Katsuwonus pelamis). Canadian Journal of Fisheries Aquatic Sciences 56:925–938.
Skalski JR, Townsend R, Lady J, Giorgi AE, Stevenson JR, McDonald RD (2002) Estimating route-specific passage and survival probabilities at a hydroelectric project from smolt radiotelemetry studies. Canadian Journal of Fisheries and Aquatic Sciences 59:1385–1393.
Thorup K, Rahbek C (2004) How do geometric constraints influence migration patterns. Animal biodiversity and conservation 27(1):319–329.
Walther BA, Wisz MS, Rahbek C (2004) Known and predicted African winter distributions and habitat use of the endangered Basra reed warbler (Acrocephalus griseldis) and the near-threatened cinereous bunting (Emberiza cineracea). Journal of Ornithology 145:287–299.
Webster MS, Marra PP, Haig SM, Bensch S, Holmes RT (2002) Links between worlds: unraveling migratory connectivity. Trends in Ecology & Evolution 17(2):76–83.
Wernham CV, Toms MP, Marchant JH, Clark JA, Siriwardena GM, Baillie SR (eds.) (2002) The Migration Atlas: movements of the birds of Britain and Ireland. Poyser, London.
White GC (1983) Numerical estimation of survival rates from band recovery and biotelemetry data. Journal of Wildlife Management 47:716–728.
Wikelski M, Kays R, Kasdin J, Thorup K, Smith JA, Cochran WW, Swenson GW Jr. (2007) Going wild: what a global small-animal tracking system could do for experimental biologists. Journal of Experimental Biology 210:181–186.
Zink G (1973–1985) Der Zug Europäischer Singvögel. Ein Atlas der Wiederfunde beringter Vögel. Teil 1–4. Radolfzell.
Zink G, Bairlein F (1995) Der Zug Europäischer Singvögel. Ein Atlas der Wiederfunde beringter Vögel. Teil 5. Radolfzell.
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2009 Springer Science+Business Media, LLC
About this chapter
Cite this chapter
Thorup, K., Conn, P.B. (2009). Estimating the Seasonal Distribution of Migrant Bird Species: Can Standard Ringing Data Be Used?. In: Thomson, D.L., Cooch, E.G., Conroy, M.J. (eds) Modeling Demographic Processes In Marked Populations. Environmental and Ecological Statistics, vol 3. Springer, Boston, MA. https://doi.org/10.1007/978-0-387-78151-8_52
Download citation
DOI: https://doi.org/10.1007/978-0-387-78151-8_52
Publisher Name: Springer, Boston, MA
Print ISBN: 978-0-387-78150-1
Online ISBN: 978-0-387-78151-8
eBook Packages: Mathematics and StatisticsMathematics and Statistics (R0)