
Abstract
Flax fibers (Linum usitatissimun L.) with good mechanical properties are required to reinforce polymers. This work provides an original method to determine the stiffness of dried or green fibers contained inside the plant. We studied three recent flax varieties (Marylin, Eden and TDL25) with distinct lodging resistances. After mechanical characterization on elementary fibers and morphological analysis on stems, we showed, by flexural tests on green and dried stems, that it was possible to correlate their stiffness with the Young modulus of the elementary fibers as well as their internal organization. This result confirmed the crucial role of fibers in supporting the plant and in particular in the lodging behaviour.
Access provided by Autonomous University of Puebla. Download conference paper PDF
Similar content being viewed by others

Keywords
Introduction
The use of flax fibers as composite reinforcements is justified by the renewable character of these fibers as well as by their good specific mechanical properties (Lefeuvre et al. 2014). Moreover, their mechanical performances are yearly reproducible, for a same variety (Lefeuvre et al. 2013) or for numerous batches (Baley and Bourmaud 2014). In order to ensure a reliable culture of flax, the lodging stability of the plant could be considered as a key point. Thus, an important varietal selection work is performed in order to develop new varieties with high fibers yield and good disease or lodging resistance, to ensure a sufficient income for the farmer.
The lodging originates because of buckling of the stems due to abundant rain, strong winds or numerous visiting animals (Menoux et al. 1982); in case of severe lodging, the plants might be lying down on the ground. The lodging of flax stems is highly influenced by the stem stiffness, the environmental solicitations and especially the additional water weight. Consequently, the distribution of the water drops on the plant increases its mass and, with wind, the risk of instability becomes even more significant. Moreover, plants could react to environmental stimuli by different ways; this phenomenon, called thigmomorphogenesis, generally induces a decrease into the plant height conjugated to an increase of their diameter and ramification (Telewski and Jaffe 1986). In addition, mechanically induced stress could cause an improvement of the cell wall stiffness (Biddington 1986).
In this article, we study the correlation between the stem stiffness and the elementary fibers modulus based on their distribution. Green and dried stems were both analysed. We selected three varieties of flax (Linum usitatissimum), Marylin, Eden and TDL25 having various lodging resistances. Firstly, we analysed the fiber yield for each variety as well as the stems morphology . Secondly, we studied the elementary fibers’ mechanical properties after dew retting and scutching according to the fibers’ location along the stem. The third part of the paper is dedicated to the measurement of the dried and green stem bending stiffness, giving an estimation of the modulus of elementary fibers in a green plant. The last part concerns an estimation of the risk of stem buckling.
Materials and Methods
Marylin, Eden and TDL 25 textile flax varieties were used for this study. Flax samples were obtained from the agricultural cooperative Terre De Lin (France). Eden is a variety registered since 2009 and is known for its lodging resistance, whereas TDL25 isn’t registered yet (recently selected) and presents a high lodging sensitivity. Flax stems were grown in France (Saint Pierre le Viger, France) in 2009, 2010, 2011 and 2012 (Marylin) or 2013 (Eden and TDL25) with almost same densities (around 1500 plants/m2) which corresponds to the conventional density for flax culture (Bert 2013). An original climatic scenario characterized each year. The year 2009 corresponded to a base/average year with accumulated sunshine hours and a pluviometry close to the mean values calculated over 30 years. The year 2010 exhibited a drought during the vegetative phase with accumulated sunshine hours slightly higher than the average value and with a water deficit in April and May. In 2011 there was a drastic drought during the vegetative phase with accumulated sunshine hours largely higher than the normal/than the average value; this drought was increased by a lack of water in March, April and May with water reserves already affected by the drought in the year 2010. The droughts during the vegetative phase had repercussions on the growth of the stem and generated short flax stems. Year 2012 exhibited an opposite climatic scenario compared to the previous years. The accumulated sunshine hours were close to the normal; however there was one important excess of water in April and June inducing lodging for some fields. Lastly, in 2013, weather conditions were favourable and close to normal ones; there was no lodging in the cultivated fields for this work.
Mechanical properties (Young’s modulus, ultimate strength and failure strain) of single flax fibers were obtained from tensile tests. Fibers are hand-extracted and bonded onto a paper frame and clamped on a universal MTS type tensile testing machine equipped with a 2 N capacity load cell. Tensile tests were carried out according to the AFNOR XP T-501-2 standard (AFNOR 2009) taking into account the compliance of the cell-load. The gauge length was 10 mm and fibers were tested at a crosshead displacement speed of 1 mm min−1 and at controlled temperature (23 °C) and humidity (50 %). The mean diameter used for calculations is the average of 6 values measured along the fiber (Lefeuvre et al. 2013). For each sampling area (bottom and middle of the stem), at least 50 elementary fibers were tested.
External dimensions (height and external diameter) of the stems were assessed using a tape measure and a calliper. The bottom of the stem was defined as the beginning of the aerial part of the plant. Internal morphology of the stems and characterization of the fibers were performed by using histological cuts. The flax stem was embedded in elder marrow and cut with a Ranvier’s microtome and a razor blade. Cuts on green stems were performed about 30 min after being removed from the freezer to avoid drying. Cuts on dried stems were made after immersion for 20 min in a 50 % ethanol-50 % water solution.
Sections were observed by using an optical microscope and pictures were taken. To enable the detection of fibers, these were first manually encircled using the Gimp® software. Images were then analysed with image J®, an image analyser program including a cell-counter. Thus, the following information was obtained: fiber number, areas, and filling rate, as well as the area of transverse section and stem diameter. The analysis was first performed on green stems and then on dried stems in order to measure the variation of these previous parameters according to the stem water content.
For 3 points bending tests, flax stems were cut into 15 cm long segments. Each section was weighed and submitted to a three-point bending test. Tests were performed by using a MTS universal testing machine with a 50 N load cell with a span length of 120 mm. Given the diameter of the stem (around 2 mm) and the selected span length, the shear effect can be ignored here. The crosshead speed is set at 20 mm min−1. The force displacement curve was registered and used to determine the bending stiffness EI of the stem. Considering the stem as a uniform beam with circular cross section, Eq. 1 gives displacement in the middle of the section:
where F is the applied force, L the span length and EI the bending stiffness.
Assuming that fibers are the strengthening tissues responsible for the lodging resistance, the flax stem is considered as a hollow circular beam. Cross-sections obtained by using a microtome were observed and the axial second moment of the area was determined using image analysis, as given by Eq. 2:
where D and d are the external and internal fiber crown, respectively.
Thus, Young’s modulus E of the stem is given by the formula:
where dF/dY is the initial slope of the force displacement curve.
The tests were first performed on green stems 30 min after being removed from the freezer. The tests were stopped before the rupture of the stem to stay in the elastic zone. The stems were then stored at a controlled temperature and humidity for drying. Once drying was completed, similar tests were performed on these same stems to measure the influence of water content on the bending rigidity of the stem.
Results
In first time, we selected Marylin flax stem cultivated in the same geographical area over 4 successive years. Various parameters influencing the stem stiffness were analysed: the height and the diameter of the stems, their internal organization including the proportion of fiber bundles, the diameter and mechanical properties of elementary fibers.
Stem Height and Fibre Yield
Figure 1 presents the relationship between the technical length of long fibres and the production of scutched fibres by hectare. It clearly evidences that the more the stem height increased the more important the percentage of collected fibres. Today in France, according to the professional data (Bert 2013), the average mass of fibres (mass of the long fibres and tows after scutching) collected by hectare is around 2500 kg. The average yield for the 11 batches was 2059 ± 941 kg/ha. The fibre yields were significantly lower than the average for the samples submitted in drastic drought periods in 2011 and significantly higher for one sample, which received the highest amount of rainfall in 2012. Thus, fibre production was strongly correlated with the climatic conditions, decreasing with the drought intensity and increasing with rainfall. Thus, in normal rain-fall and sunshine conditions, the shorter plants could exhibit a significant scutched fibre yield. This phenomenon could be due to a better fibre thickening or to differences in the fibre number.

Relationship between the length of technical long fibres and the fibre yield. The dashed line is the usual yield from the professions’ data
Regarding the average fibre yield after retting and scutching, the average was estimated to be 36.5 ± 3.5 %; this value is well correlated with the usual yield published by the profession (37 %) (Bert 2013). Globally the fibre rate increased when the technical length increased. This effect was the most remarkable within the 2011 year due to the drastic drought occurring this year. For each year, 2009–2011, there were shorter stems producing larger fibre rates, suggesting that the relationship was complex. This might happen, especially because the fibre rate depended not only on the fibre biosynthesis but also on the retting extent as well as the degree of humidity in the scutched plant. On a general way, the data pointed out the strong impact of an extreme drought climate on the fibre yield and rate of fibres in the biomass. On the other hand, there was no major effect of an excess rainfall compared with normal growth conditions.
Elementary Fibres Mechanical Properties
Mechanical properties of the fibers were investigated. The studied fibres, whatever their batch, exhibited a Young modulus of 57.5 ± 0.3 GPa, a strength at rupture of 1034 ± 6 MPa and an elongation of 2.0 ± 0.1 %. Baley and Bourmaud (2014) analyzed the tensile mechanical properties of 50 batches of flax fibres (Linum usitatissimum) cultivated in France between 1993 and 2011 submitted to dew retting and scutching. They were characterized under the same conditions as those of the present study in accordance with the XPT 25-501-2 (AFNOR 2009) standard (constant T° and hygrometry). The average Young modulus of these 50 batches was 52.6 ± 8.6 GPa, the strength at rupture 945 ± 200 MPa and the elongation at rupture 2.07 ± 0.45 %. Interestingly, most of our data exhibited higher values for E and σ than the mean estimated for the 50 reference samples (Fig. 2).

Relationship between the strength at break and the Young modulus of flax fibres (Baley and Bourmaud 2014)
Due to their low density (1.53 vs. 2.54 g/cm3 for the glass fibres), the Marylin fibres average specific stiffness and stress at rupture were estimated to be 36.9 GPa cm3/g and 646 MPa cm3/g, respectively. These values are close to, and sometimes, for the Young’s modulus, better than those of glass fibres (28.3 GPa cm3/g and 695 MPa cm3/g) (Coroller et al. 2013). Moreover, although the flax fibre tensile properties are widely scattered, this phenomenon also occurred with the breaking properties of the glass fibres in opposition to their Young’s modulus, which is more stable.
Organization and Morphology of the Stems
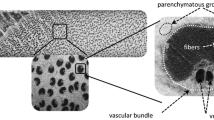
Figure 3 presents a section of a stem of flax (Fig. 3a) as well as a zoom on the peripheral area (Fig. 3b) where the fibres are arranged into bundles. The fibres were considered as support materials to reinforce the stem structure. For each batch, three stems were studied. Table 1 shows the results of the morphological analysis of the stems at middle height.

Optical image of a stem section a (X 10) and focus on a fibre-bundle area b (X 40)
We can observe that the number of fibres per section and their diameter increased with the diameter of the stems. Thus, the drought had a direct impact on the number of thickened fibres per section. Either, consequently to the plant adaptation to drastic environmental conditions, there was a reduction of the division of fibre cells at the level of the meristem or a lack of cellulose deposition (hence of thickening). Such a relationship also existed in the genetic variability (Brutch et al. 2011) (R2 = 0.74).
In parallel, the diameter of fibres increased significantly with the height. When Brutch et al. (2011) compared 13 lines, no correlation was found between the diameter of fibres and the stem height (genotypical variation). Thus, ecological regulation (this study) and genotypic relation showed opposite trends within a line (Marylin, this study). Both stem diameter and cell diameter increased with height suggesting some kind of synchronous regulation of most of the morphological parameters.
Consequently, the conjugated increase of the fibre number and of their diameter induced an increase of the diameter of the stems. Thus, the number of thickened fibres within a bundle is not a constant for a given variety: there is a strong impact due to the climate, via the regulation of the stem diameter and height (R2 = 0.94). The use of image analysis showed that for a short stem of 25 cm there was an average of 2.6 fibres in the thickness of a bundle and 4.3 in the case of an 80 cm length stem. It is average information because the number of fibres in a bundle and the section of the latter are not perfectly constant. This is explained, amongst other things, by the existence of leafs fixed on the stems which could disturb the bundles organization.
In complement we can notice that the number of fibres in a section changed according to the position along the stem height, being most numerous in the middle of the stem. This was previously described for another commercial variety, Natasha (Morvan et al. 2003). Whatever the stem height, a little variation in the number of fibres is observed between the bottom and middle position whereas a significant fibre number decrease is observed for the top position. Moreover, the bibliography shows that the mechanical properties are the best in middle part of the plant (Charlet et al. 2007, 2009). The mechanical and morphological differences observed at various levels in the height of the plants might be explained by the cellulose filling of the secondary wall which occurred after the cell elongation and differentiation steps.
Correlation Between the Stem Stiffness or Morphology and the Fibre Mechanical Properties
3-point bending tests were carried out on fractions of stems taken from the central part of the plant in order to determine their flexure stiffness. The total area of the fibre bundles over a section was estimated using image analysis. Knowing the flexure stiffness of the stems and the area and thickness of the fibre crown, it was possible to estimate the Young modulus of fibres in the stems thanks to the method described in the experimental part. The hypothesis was that only the fibres contributed to the stiffness of the stems.
Figure 4 shows the calculated fibre Young’s modulus for the different tested stems as a function of their length. Whatever the plant height, the calculated fibre stiffness is stable. This point confirmed the results previously obtained by elementary fibre tensile characterization (Fig. 2). The average Young modulus (deflection tests on stems) of fibres in the stems was estimated to be 63.9 ± 8.6 GPa; this value was in the same range as the stiffness measured by tensile tests (56.5 ± 4.4 GPa). The slight difference was considered to be acceptable taking account the uncertainty of the crown diameter measurements, as well as the possible modification of the fibre diameters induced by the ethanol-water immersion before cutting. Moreover, we hypothesized that only fibres contribute to the plant stiffness; the role of the woody core was neglected in our approach. On the other hand, the agro industrial processes (retting and scutching), as well as the dew retting, might have reduced the stiffness potential of the fibres.

Elementary fibres Young’s modulus estimated from the stem stiffness and its internal organization
It was shown that the fiber yield and number of fibers increased with the height of the plants, the number of fibers being maximal at medium height. In addition, the fibers’ diameter decreases with the height of the stem because of the growth mechanisms. Lastly, flexural tests on stems made it possible to correlate their stiffness with the Young modulus of the elementary fibers as well as their internal organization.
In a second time, we studied two recent flax varieties (Eden and TDL25) with a distinct lodging resistance. The results of the mechanical characterization indicate that the Eden variety has a superiority concerning fiber stiffness . The analysis of the localized bending stiffness of a dried stem and of its fiber distribution in the corresponding cross section allowed us to estimate the fibers average modulus. As detailed previously for Marylin flax, results obtained by this method were very close to the tensile tests values. The same procedure was used on green stems to approach the living state of the plant and to determine the Young’s modulus of green fibers. Figure 5 shows the comparison between dried or green fibers mechanical properties.

Evolution of fibers’ rigidity between the green and dried state
The results highlighted a variation of fiber stiffness between the green state and the dried state (around +25 %). The results enabled the use of a simplified buckling model, which confirmed the superiority of the Eden variety in lodging resistance. Thus, the analysis of the mechanical properties of flax stems and their structure could be a selection criterion.
Conclusions
During this work, we highlighted important differences in the structure of the stems or flax fibres according to the years of growth; they could be linked to the climatic conditions suffered by the plants. Firstly, the study was conducted on Marylin flax. The fibre yield was strongly correlated to the fibre height and morphological differences occurred in length and diameter of the stems (both characters being highly correlated indicating a synchronous bi-dimensional regulation of the fibre dimensions). Moreover, the number of thickened fibres and their diameters were shown to vary according to the location of the plant and also according to the height of the stems. Thus, the stem diameter increased with the height of the plants as well as the thicknesses of the fibre bundles whereas the average diameter of fibres decreased from the bottom to the top of the stem. According to the environment, the plant will thus develop in a different way and adapt its morphology to be able to resist as much as possible to the external aggressive conditions and in particular, to lodging.
Interestingly, the mechanical properties of the fibres (taken from the middle) remain constant overall whatever the morphology or the structure of the stems and the diameter of the fibres. We showed that the morphology of the cell walls is directly related to the stem stiffness and that it is possible, through bending tests on portions of stems, to estimate the stiffness of elementary fibres by knowing their arrangement within the stem. These results were confirmed on two other varieties (Eden and TDL25) and the same approach was used on green stems to be close to that of a living plant. The estimate of the average stiffness of the green fibers was around 25 % lower than the dried fibers. Thus, with a simplified model, the risk of buckling (lodging) of green plants was then approached. The model takes into account the plant mass, the stem height and the flexural stem stiffness (bottom part); showing that the bending stiffness of the stem is a very important parameter for the resistance to lodging which is a buckling instability. The stem stiffness appeared to be mainly influenced by the fibers’ rigidity in the present study, explaining the superiority of Eden for lodging.
In future works, it would be interesting to study the length of the elementary fibers in the plant height and the evolution (microstructure, composition) of cell walls during drying. Also, the study of the resistance to lodging in real conditions (living flax) could be very instructive.
References
Lefeuvre A, Bourmaud A, Morvan C, Baley C. Tensile properties of elementary fibres of flax and glass: Analysis of reproducibility and scattering. Mater Lett 2014;130:289–91.
Lefeuvre A, Bourmaud A, Lebrun L, Morvan C, Baley C. A study of the yearly reproducibility of flax fiber tensile properties. Ind Crops Prod 2013;50:400–7.
Baley C, Bourmaud A. Average tensile properties of French elementary flax fibers. Mater Lett 2014;122:159–61.
Menoux Y, Katz E, Eyssautier A, De Parcevaux S. Résistance à la verse du lin textile: influence du milieu et critères de verse proposés. Agronomie 1982;2:173–80.
Telewski FW, Jaffe MJ. Thigmomorphogenesis: Field and laboratory studies of Abies fraseri in response to wind or mechanical perturbation. Physiol Plant 1986;66:211–8.
Biddington N. The effects of mechanically-induced stress in plants — a review. Plant Growth Regul 1986;4:103–23.
Bert F. Lin Fibre: culture et transformation. 2013.
AFNOR. Determination of tensile properties of elementary flax fibres XPT 25-501-2. 2009.
Coroller G, Lefeuvre A, Le Duigou A, Bourmaud A, Ausias G, Gaudry T, et al. Effect of flax fibres individualisation on tensile failure of flax/epoxy unidirectional composite. Compos Part A Appl Sci Manuf 2013;51:62–70.
Brutch NB, Sharov IY, Pavlov A V, Porokhovinova EA. Diversity of flax characters associated with fibre formation and environmental influence on their formation. Russ J Genet Appl Res 2011;1:361–70.
Morvan C, Andème-Onzighi C, Girault R, Himmelsbach DS, Driouich A, Akin DE. Building flax fibres: more than one brick in the walls. Plant Physiol Biochem 2003;41:935–44.
Charlet K, Jernot JP, Gomina M, Bréard J, Morvan C, Baley C. Influence of an Agatha flax fibre location in a stem on its mechanical, chemical and morphological properties. Compos Sci Technol 2009;69:1399–403.
Charlet K, Baley C, Morvan C, Jernot JP, Gomina M, Bréard J. Characteristics of Hermès flax fibres as a function of their location in the stem and properties of the derived unidirectional composites. Compos Part A Appl Sci Manuf 2007;38:1912–21.
Acknowledgments
The authors would like to thank Terre de Lin and Jean Paul Trouvé for supplying flax fibers and stems, and ADEME for the financial support.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 RILEM
About this paper
Cite this paper
Bourmaud, A., Gibaud, M., Lefeuvre, A., Morvan, C., Baley, C. (2016). Influence of Stem Morphology and Fibers Stiffness on the Loading Stability of Flax. In: Fangueiro, R., Rana, S. (eds) Natural Fibres: Advances in Science and Technology Towards Industrial Applications. RILEM Bookseries, vol 12. Springer, Dordrecht. https://doi.org/10.1007/978-94-017-7515-1_4
Download citation
DOI: https://doi.org/10.1007/978-94-017-7515-1_4
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-017-7513-7
Online ISBN: 978-94-017-7515-1
eBook Packages: EngineeringEngineering (R0)