
Abstract
The theory of evolution has recently been in turmoil, with great interest in applying empirical information from EvoDevo, genomics, and ecology into the framework of quantitative genetic studies of evolution. Ciona is a small genus of sea squirts within the class Ascidiacea of the subphylum Tunicata, the sister group of vertebrates, a phylogenetic position that has contributed to fuel the interest in studying development and evolution in ascidians. Ciona species display several traits of evolutionary interest, e.g., conservative anatomy, high genetic polymorphism, cryptic speciation, metapopulation structure and invasive behavior. Some of these aspects may depend on the ecology of these marine animals, which display a great ecophysiological tolerance and unpredictable colonization capabilities. In addition, natural populations show the occurrence of spontaneous mutations with phylomimicking phenotypes. Here we review some key features of this talented marine organism that promise to provide insights in specific aspects of the expanded evolutionary biology.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 The Model System
The ascidian genus Ciona (Chordata, Tunicata) has attracted the interest of biologists for over a century because of the ecological importance and the key position in the evolutionary path leading to vertebrates. Ciona intestinalis (L. 1767) is a marine invertebrate that lives on shallow hard bottoms, where it can represent a major component of coastal benthic ecosystems. The life cycle is characterized by a short pelagic stage with a chordate-like tadpole larva that, upon settlement and metamorphosis, loses the chordate body plan and generates an invertebrate form. To increase the rate of successful reproduction, Ciona is hermaphrodite like all tunicates, and like many ascidians, it spawns large numbers of gametes on a daily basis. Moreover, due to the high tolerance of C. intestinalis to ecophysiological parameters, such as euthrophic conditions following anthropization, this opportunistic species is considered a sentinel of environmental conditions in coastal biotopes. These biological traits may contribute to the cosmopolitan, and sometimes invasive, distribution of intestinalis in temperate to sub-boreal waters of both hemispheres. Phylogenetic and comparative genomic studies have demonstrated that the Linnean taxon is a complex of at least four (C. intestinalis spA-D) morphologically cryptic but genetically distinct sibling species of disjoint geographical distribution (see Sect. 6.2; Suzuki et al. 2005; Caputi et al. 2007; Nydam and Harrison 2007; Zhan et al. 2010) (Fig. 6.1). C. intestinalis spA is the cryptic species for which the genome sequence is available (Dehal et al. 2002).

Sympatric individuals of Ciona intestinalis spA and spB from North European coasts. Photo by L. Caputi
Evolutionary developmental biology is providing strong evidence that embryonic processes are conserved between basal chordates and vertebrates. Therefore, it is not surprising that this animal occupies a prominent role in several branches of biology (Satoh 1994). A unique blend of advantageous features for experimental analysis has made it a powerful model organism for genetics and genomics of chordates due to ease of manipulability. Its genome carries fewer genes and less genetic redundancy than in vertebrates (Dehal et al. 2002; Small et al. 2007a). In addition, the application of bioinformatics and experimental approaches to the Ciona system, such as means for manipulating gene expression, provides reverse and forward genetics tools for studying the genetic basis of cellular and developmental processes common to all chordates (Hendrickson et al. 2004; Sasakura et al. 2007; Sordino et al. 2008; Veeman et al. 2008).
Besides advancements in culturing and formal genetics, yet research with C. intestinalis is largely based on sampling in nature. Indeed, growing Ciona strains is labor intensive, and sporadic loss of genotypes occurs due to yet suboptimal culturing conditions (Kano 2007; Cirino et al. 2002; Joly et al. 2007). Simple, efficient, and rapid method for long-term storage of Ciona sperm in liquid nitrogen can greatly facilitate strain management and experimental design. Obligation to sample natural populations for laboratory needs has prompted better understanding of genetic diversity in the wild as well as of Ciona population biology sensu lato, also in the light of preserving natural resources.
However, knowledge of classical genetics is still fragmentary. Progress in genetic studies is revealing a high degree of genetic polymorphisms at individual and population scales (Dehal et al. 2002; Boffelli et al. 2004). Therefore, to understand the complex genetic structure observed between natural populations at different geographical scales (Schmidtke and Engel 1980; Kano et al. 2001; Small et al. 2007b; Sordino et al. 2008; Zhan et al. 2010Caputi et al. Personal Communication), it is important to address genetic polymorphism and gene flow within and among distant and close populations by means of unlinked markers of nuclear, mitochondrial, and ribosomal origins (see Sect. 6.3). This allows to resolve historic phylogeographical patterns of C. intestinalis populations and to determine current patterns of allelic flow between genotypes. Resolution of the genetic structure of populations is also a prerequisite for undertaking analysis of environmental selection across gradients of physical conditions within specific areas.
What ecological and evolutionary forces generate and maintain variation? Which and how many loci are involved? Which is their effect? Answering these questions may help to elucidate the particular evolutionary and demographic history of natural populations, identifying new genomic regions and candidate genes of evolutionary significance (Stinchcombe and Hoekstra 2007). The increasing knowledge concerning C. intestinalis population biology makes this species an ideal tool for studying microevolutionary processes. The advent of next-generation genomic technologies in C. intestinalis research provides a useful instrument to study environmental impact and adaptation of this invasive species by population genomics studies, as well as to understand the involvement of epigenetic mechanisms in the control of ascidian development. Genomic technologies can be used to analyze the dynamic distribution of epigenetic marks such as cytosine methylation, posttranslational modification of histone tails, and nucleosome composition, at distinct stages of development and in different environmental settings. The resulting spatiotemporal information will provide detailed insights into how the ascidian transcriptome is controlled by epigenetic processes, and what is the effect of environmental cues on the chromatin landscape. We suggest that combining ecology, population genetics and genomics, and the study of spontaneous mutations in Ciona may aid in understanding which types of microevolutionary mechanisms and factors do generate variation (see Sect. 6.5).
2 Distribution and Ecology
Ciona intestinalis has always been considered as a species with cosmopolitan distribution, and broad environmental tolerance to unstable or fluctuating environments (Hoshino and Nishikawa 1985; Therriault and Herborg 2008a, b). Nevertheless, the recent finding of cryptic species within the so-called C. intestinalis, has questioned this general assumption. Two C. intestinalis cryptic species are more widely distributed and have a mostly disjoint distribution, which overlaps only in few areas. C. intestinalis spA, in fact, lives in temperate seas around the globe (Mediterranean Sea, south European and South American Atlantic coasts, and Pacific Ocean), while C. intestinalis spB is found in North Atlantic waters (Caputi et al. 2007; Zhan et al. 2010). They coexist in the English Channel, where they can theoretically hybridize (Caputi et al. 2007; Nydam and Harrison 2011a, b). A third and a fourth cryptic species, defined as spC and spD, have been described only for one location in the Mediterranean Sea and the Black Sea, respectively, within the distribution range of spA (Nydam and Harrison 2007, 2010; Zhan et al. 2010) (Fig. 6.2).

Worldwide distribution of Ciona intestinalis cryptic species
Specific studies on life cycle and life history of C. intestinalis spA are lacking. As a matter of fact, most published information deals with the biology and ecology of C. intestinalis spB (Suzuki et al. 2005; Caputi et al. 2007), referring to Scandinavian-Danish (Svane and Havenhand 1993; Petersen and Svane 1995; Petersen et al. 1995; Petersen 2007) and eastern Canadian populations (Ramsay et al. 2008, 2009). It has been shown that recruitant waves of spB larvae settle close to or on adults individuals, forming multigenerational clusters (Havenhand and Svane 1991), and that larval settlement is enhanced by the formation of a fouling canopy on a bare substratum that is subsequently monopolized by ascidian clusters (Schmidt 1983). Thus, the environment into which C. intestinalis settles is strongly affected by the occurrence and density of conspecific individuals (Marshall and Keough 2003). Recruitment is also driven by the seasonal fluctuations of density that C. intestinalis experiences in the different sites the taxon colonizes, as a function of the year and particularly season (Lambert and Lambert 1998). Moreover, predation does not seem to be a selective factor for recruitment as C. intestinalis lives in environments where the number of predators is reduced (Petersen and Svane 1995; de Oliveira Marins et al. 2009).
According to known distribution, spB is more adapted to cold water, as also reflected in its response to the unfavorable seasons. C. intestinalis spB populations, in fact, experience a reduction in density of individuals in winter (Petersen and Svane 1995), while spA populations completely disappear in late summer and winter, reoccurring when the environmental conditions are again favorable in spring and autumn as shown for the southern California harbors (Pérès 1952; Sabbadin 1958; Lambert and Lambert 1998). Due to the lack of specific information, many questions related to the ecology and distribution of spA are still open. For example, how T° - S ‰ seasonal variations influence the reoccurrence of the species in the benthic community, being the taxon offspring sensible to the quality of the environment (Marshall and Keough 2003) even if the species tolerates moderate excess of nitrogen, reduced water clarity, low oxygen levels, and periodic algal blooms (Carman et al. 2007). C. intestinalis inhabits harbors and confined natural environments where thanks to its capacity to tolerate a wide range of temperature and salinity, produces viable gametes in different environmental conditions, which are an important extrinsic ecological forcing influencing its life history, in particular the occurrence timing (Dybern 1965; Lambert and Lambert 1998; Carver et al. 2003). Tolerance to salinity and temperature also varies ontogenetically, embryos and larvae being less tolerant to high temperatures, and young post-metamorphic ascidians more resistant than adults to low temperatures. The favorable temperature range is between 15°C and 20°C, with a salinity value of 35‰ (Marin et al. 1987). C. intestinalis has a remarkable filter suspension feeder capacity with a high efficiency in terms of particles range in relation to temperature which affects the clearance rates (Petersen 2007). Examining stomach and gut contents of C. intestinalis, it is possible to distinguish an amorphous matrix made of particulate matter rich of pelagic and benthic diatoms (Fig. 6.3). The species represents also a useful sentinel organism being able to tolerate oil pollution, industrial and radioactive wastes into marine environments concentrated in its tunic or filtered from the water column. Recently, the invasive behavior of Ciona species has been reported throughout the globe (e.g., Canada, Argentina, South Africa, New Zealand; reviewed in Zhan et al. 2010). In most newly introduced areas, C. intestinalis is a very competitive species that rapidly covers nearly 100% of the available substratum excluding native species in a short period of time (Ramsay et al. 2008). The ecological impact caused by the introduction of nonindigenous ascidian species in natural coastal environments has raised a growing concern in recent years (Whitlatch et al. 1995; Lambert 2001). Nonindigenous species cause different types of influences, as the reduction of biodiversity, the alteration of ecosystems, and the decline of native species (Everett 2000; Pimentel et al. 2000). The ecological impact also reflects on the coastal human activities, with severe economic consequences. For this reason, understanding the evolutionary and ecological causes responsible for the rapid spread of invasive species represents one of the main challenges for conservation biologists and environmental managers.

Stomach content of Ciona intestinalis spA. Note particulate matter embedding fragments and recognizable pelagic diatoms, such as one belonging to the genus Thalassiosira sp. (light gray arrow), and an unidentified species. Scale bar 5 μm
3 Population Genetics and Phylogeography
Studies of phylogeography structure, genetic diversity, and evolutionary changes can help in understanding the potential for colonization and establishment of alien species, the geographic patterns of invasion and range expansion, and the potential for evolutionary responses to novel environments, giving important insights in the definition of management practices (Everett 2000; Holland 2000; Pimentel et al. 2000). Understanding population dynamics and spread potential of invasive species means also determining the degree of population connectivity and investigating factors driving genetic exchange at various geographical scales. Population connectivity in aquatic invasive species can be influenced by water currents, natural and human-mediated pathways of propagule dispersal, species’ life histories, and by variation in environmental and community composition across geographical scales (Lee 2002; Darling and Folino-Rorem 2009; Jesse et al. 2009; Goldstien et al. 2010; Sorte et al. 2010).
In C. intestinalis, allelic DNA polymorphism across the entire genome is very high (1.2%, based on Single Nucleotide Polymorphisms – SNPs, and insertion/deletions; Dehal et al. 2002), including non-synonymous SNP frequency in important developmental genes such as Hox5 and Hox10 (0.27% and 1.89% in spA and spB, respectively) (Caputi et al. 2008). The average genome-wide SNP heterozygosity in C. savignyi is even higher, reaching 4.5% (Small et al. 2007b). Because of that, it has been argued that C. savignyi exhibits the highest levels of SNP variation and structural polymorphism among multicellular organisms (Small et al. 2007b).
In Ciona populations, high levels of genetic diversity may derive from several factors, among which the mutation–selection balance between reappearing phenotypes and their elimination by natural selection, the reproductive strategies, the geographical connectivity, and the association with the large effective population size. From here on we focus our attention on population genetic structure and population connectivity at both large and regional geographical scales of C. intestinalis spA.
3.1 Genetic Diversity
A high level of polymorphism has been demonstrated at various enzyme (Schmidtke and Engel 1980) and microsatellite loci (Procaccini et al. 2000; Andreakis et al. 2007; Zhan et al. 2010). In allozymes, average heterozygosity was 0.319 and the average number of alleles per locus was 2.6 (Schmidtke and Engel 1980). For microsatellites, allelic richness is higher, ranging from 5.1 to 5.9 (Zhan et al. 2010). The high degree of polymorphism observed in C. intestinalis populations determines extremely high values of expected heterozygosity (HE) in all analyzed populations, with values ranging from 0.65–0.69 on eight populations distributed worldwide (Zhan et al. 2010), to 0.73 on three populations in the Gulf of Naples (Tyrrhenian Sea) (Sordino et al. 2008) (Table 6.1). These values are higher than those recorded in other solitary tunicates such as Styela clava (HE = 0.48–0.63, Dupont et al. 2009, 2010; Goldstien et al. 2010). Small et al. (2007b) suggested that extreme polymorphism in Ciona is probably associated with large effective population size rather than an elevated mutation rate.
Despite the high expected heterozygosis, populations show a relevant heterozygote deficit, which determines a significant deviation from Hardy–Weinberg equilibrium (HWE) (P < 0.001; Sordino et al. 2008; Zhan et al. 2010). Similar or higher departure ratio from HWE was previously reported in other ascidians including the solitary S. clava (Dupont et al. 2010) and the colonial Botryllus schlosseri (Ben-Shlomo et al. 2006). The departure resulting from heterozygote deficit might be explained in three different ways. First, recurrent inbreeding. Different studies suggested that C. intestinalis larvae generally settle very close to the adult individuals (Petersen and Svane 1995; Howes et al. 2007), increasing the possibility of breeding with related individuals. Additionally, C. intestinalis self-sterility is not complete; about 15–20% of individuals can self-fertilize (Rosati and Santis 1978; Kawamura et al. 1987). Second, subpopulation structure. The possibility that a temporal and⁄or spatial Wahlund effect can be another cause for the massive heterozygote deficit, is suggested by the enhanced subpopulation structure, the high connectivity among population, which are highly dynamic in their fluctuations, and the metapopulation structure shown at small geographic scale. Third, the presence of null alleles. Null alleles are very likely to be present in C. intestinalis, due to the highly polymorphic genome, and have been recognized to be one important cause for heterozygote deficiency in many marine species (e.g., Hedgecock et al. 2004; Zhan et al. 2007).
3.2 Genetic Structure
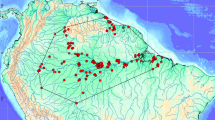
Despite high genetic polymorphism of populations, geographic isolation and theoretical short-distance dispersal of larvae, an impressive level of genetic homogeneity is revealed by microsatellites and mtDNA analyses. In the Mediterranean basin, C. intestinalis spA shows a single mitochondrial COI haplotype and low degree of microsatellite-based differentiation (Caputi et al. 2007; Caputi et al. personal communication), suggesting a recent bottleneck event followed by rapid recolonization by a dominant mitochondrial haplotype with high dispersal capacity (Fig. 6.4). In comparison, spB mitochondrial COI shows higher geographical structure, suggestive of fixed populations (Caputi et al. 2007; Zhan et al. 2010). From this point of view, the Mediterranean Sea seems to act as a metapopulation formed by genetically homogeneous clusters of spA individuals occupying fragmented habitats in completely different environmental conditions (Caputi et al. 2007). Another theoretical factor linked to the observed lack of sharp genetic structure is the absence of barriers to gene flow among populations that may dilute the effects of genetic bottlenecks and decrease the genetic differentiation among populations (Nei et al. 1975). Nevertheless, we hypothesize that the short larval dispersal and ecological preference for enclosed habitats of C. intestinalis act as barriers to natural patterns of gene flow.

Nested Clade Analysis of C. intestinalis spA populations performed on the COI statistical parsimony cladogram. Modified from Caputi et al. personal communication
Global Fst values, ranging from 0.0353 to 0.0543, are dramatically lower than the average values (0.2354) recorded for populations of the same areas, suggesting that gene flow among populations at a large geographical distance is more active than gene flow among close-by localities (Zhan et al. 2010). These results are in agreement with preliminary observations showing high levels of migration between continents (Caputi et al. personal communication).
How can we explain the lower gene flow at regional scale, compared to global scale, coupled with the absence of genetic structure and low theoretical dispersal of larvae? In our opinion there are two possible reasons. The most obvious explanation is that the geographic structure of C. intestinalis spA does not reflect only natural dispersal patterns but results also from propagation vectors of anthropogenic origin. The second is that local populations can be adapted to specific environmental conditions and isolated from local gene flow, due to their persistence in enclosed environments such as lagoons and marinas.
Larvae can be entrapped in water currents, which may offer an effective method of long-distance dispersal in “open” environments lacking of physical barriers (Palumbi 1992). By the way, this dispersal method appears to have a limited range and is not expected to be sufficient to homogenize genetic variation at an intercontinental scale (Dupont et al. 2010). Generally, human-mediated vectors such as ballast water exchange, vessel hull fouling, and aquaculture trading have been considered responsible for such a genetic pattern and the recent range expansion of C. intestinalis (Carver et al. 2003; Lambert 2007). Looking at the isolation of populations living in enclosed environments, no significant differentiation was detected between open-shore and lagoon sites in the Gulf of Naples (Tyrrhenian Sea, Italy). The only exception is represented by the innermost population of the Fusaro lagoon (Sordino et al. 2008), which shows genotypic proportions deviating significantly from HWE expectations at several loci. This population occupies specific microhabitats and it oscillates between periodical extinction events from the upper layers and subsequent recolonization with the likely contribution of deeper refugees (Riisgard et al. 1998). These environmental constraints coupled with nonrandom mating, or eventually selfing, offer the reproductive assurance of selected mating combinations and an increased probability that locally adapted genotypes are able to persist (Fields and Johnston 2005). In general, we can assume that local adaptation and physical barriers experienced by populations living in confined environments can add an important variable in the interpretation of neutral patterns of gene flow in C. intestinalis (Fig. 6.5).

Principal coordinate analysis showing significant differentiation between isolated (FuI) and open-sea (VC/CdS) populations. (Modified from Sordino et al. 2008)
4 Naturally Occurring Mutations
One of the oldest and unsolved problems in evolutionary biology is which mutations generate evolutionarily relevant phenotypic variation. To understand the genetic basis of evolutionary change in nature, we need to address questions about the relative importance of coding vs. regulatory DNA in morphological variation, the origin of phenotypic traits by either natural selection or genetic drift, and how the ecology mediates such processes. Because the genetic variation responsible for phenotypic variation is but a subset of the genetic variation in general, it is essential to understand the evolutionary processes underlying this variation if we are to identify genes that cause morphological transitions. Furthermore, it is clear that allelic variation affects responses to key environmental stressors.
Several traits make ascidians suitable for studying the role of developmental genes in natural variation. Transparent and externally fertilized embryos develop by bilaterally symmetrical cleavage, according to a determined pattern, with a defined and well-characterized cell lineage and segregation of developmental fate of cells until the end of gastrulation. Larvae are valuable for understanding the elements of structure, developmental processes, and possibly also the evolution of the vertebrate body plan, including organogenesis of the chordate nervous system.
Natural variation and neutral population genetics can be used for the characterization of phylogenetically relevant mutations in C. intestinalis, under the assumption that the accumulation and distribution of mutant phenotypic classes is mostly influenced by historical and environmental factors (i.e., effective population size, genetic variability, reproductive strategies, geographical barriers, environmental harshness). It has been previously shown that up to 20% individuals from natural populations are heterozygote carriers of developmental mutations that segregate in a way typical of recessive alleles (Hendrickson et al. 2004; Sordino et al. 2008). The percentage of heterozygote carriers varies from 13.4% to 19.5% among populations, as well as the relative contribution of lethal and nonlethal mutant phenotypes that may be sharply different (20–60% and 40–80%, respectively). It has been argued that these phenotypes contribute to local adaptation but do not represent potentially independent evolutionary lineages, as suggested by the low morphological complexity of the species, which is adapted to an ecological scenario featuring low intraspecific competition and stressful environmental conditions (Sordino et al. 2008).
Changes in terms of presence, position, size, and structure of the larval brain sensory organs have occurred repeatedly in ascidian phylogeny (Jeffery 2004). In C. intestinalis, mutations that affect sensory organ phenotype are of special interest to uncover the changes in regulatory gene networks that underlie morphological diversity. Phylomimicking mutations, as they are called, have been also termed “hopeful monsters” by Richard Goldschmidt, who proposed that they were generated through alterations of development (Goldschmidt 1940). Which are the molecular networks controlling organ specification and how morphogenetic structures evolved in the chordate lineage? Which seemingly different mechanisms drove the remodelling of the larval body plan in ascidians, as seen in the independent loss of tail and brain sensory organs in Molgulid and Clavelinid species? Mutations that affect development of the Ciona embryo in a similar way might have played analogous roles during evolution, but the degree to which these mutations are identical to those upon which ascidian evolution depends is unknown. The molecular identity of Ciona mutation can be disclosed by positional cloning via bulked segregant analysis (BSA) with AFLP reactions and the support of genetic and physical linkage maps and the C. intestinalis genomic sequence (Dehal et al. 2002; Kano et al. 2006; Kano 2007; Veeman et al. 2008). Then, the orthologue of the genes mutated in Ciona can be isolated from specific ascidian lineages, and compared at the levels of sequence, expression, function, and regulation in order to address their implication in phenotypic radiation. At the population level, the molecular identity of specific mutations that underlie evolutionary shift can be correlated with the performance in natural environments, e.g., fitness of mutated sensory organs compared with the wild type, and with the rules of population genetics that allow their repeated fixation under conditions of natural selection.
The combination of marker-assisted analysis of population structure and screening of phenotypic classes is central not only in shaping models in developmental biology but also in evolutionary genetics of populations. The comparison between frequencies of natural variation and population genetic patterning represents a unique opportunity to quantify the importance of biogeographic and anthropogenic factors in affecting mutation–selection dynamics (Sordino et al. 2008).
5 Ciona Intestinalis as a Model for the Ecology and Genetics of Adaptation
Assessing the dynamics and timescales of phenotypic and genetic polymorphism in different environments requires the study of the dynamics of natural populations through extensive field work. C. intestinalis represents an ideal species for studies in the wild (see Sect. 6.2). Here we suggest that Ciona species can become a model organism for deciphering the evolutionary potential of invasive species in the newly colonized habitats and, more in general, for understanding the microevolution of natural populations in response to changing environmental conditions. This is particularly important if we consider the continuous and increasing impact of human activities on natural environments. One of the main challenges in evolutionary biology is the understanding of evolutionary processes in natural populations and their relationship with environmental conditions and environmental quality. In a recent review, Charmantier and Garant (2005) discuss three important points, which should be taken into consideration for evaluating the evolutionary potential of a species in the natural environment. First, unfavorable environmental conditions determine lower rate of evolution although the rapid increasing of human disturbance on natural environments makes it difficult to derive general trends for all the species. Second, different genotypes can have different levels of interaction with the environment, which reflects on a different evolutionary potential in stressful conditions. Third, authors stress that environments can be variable in space and time, making that the evolutionary potential of a single population can vary when environmental conditions change. In this case, the potential for microevolution is constrained by either a lack of heritable variation in unfavorable environments, or by a reduced strength of selection in favorable environments (Wilson et al. 2006).
In species which are displaced from their natural habitat and move to colonize new habitats, the change of environmental conditions to which they must adapt can be orders of magnitude faster than what would occur in the same natural habitat. Successful invasive species show to possess high evolutionary potential, which is theoretically proportional to the amount of additive genetic variation present (Fisher 1930). Nevertheless, very little is known about the role of genetic diversity in process of adaptation to new environments.
C. intestinalis seems to possess very high genetic polymorphism (see Sect. 6.3), which could be at the basis of its invasive success, in particular in some areas (Ramsay et al. 2008). Also, the existence of a significant number of natural mutants seems to confirm the high evolutionary potential of the species. Nevertheless, the life cycle of the species, the ecological requirements which seem to favor its life in isolated and enclosed environments, with low water quality, make the scenario more complicated. The deficit in heterozygosis encountered in most of the natural populations analyzed so far (Sordino et al. 2008; Zhan et al. 2010) seems to counteract the positive effect toward rapid evolution provided by the high genetic polymorphism.
Until now, no studies have specifically focused on the relationship between genetic polymorphism and adaptive potential in C. intestinalis, taking into consideration the life cycle of the species and its ecological requirements. Recently, we started a multidisciplinary research program which aims to study population ecology of the adult and larval stages, population genetics of natural populations, and evolutionary significance of natural mutants, in relation to the biotic and abiotic component of a coastal lagoon. The available molecular tools existing for this species (see Sect. 6.1) makes it an excellent model for this type of integrative studies. Actually, being C. savignyi and the two cryptic species C. intestinalis spA and spB separated by comparable genetic distance (13–15%), they altogether offer a unique experimental model for approaching levels of evolutionary divergence in developmental programs among sister taxa. Our research aims at gaining insights in population structure and diversity, to relate with temporal and spatial population dynamics; to correlate population genetic structure and the occurrence of abnormal phenotypes as an important focus for understanding selective forces that shape natural finite populations, and to gain insights into the embryological and evolutionary mechanisms that generate animal diversity. We also hope to give insights in the understanding of the evolutionary potential of invasive species in general. Whether a new mutation can contribute to morphological and, ultimately, taxonomic radiation, entails the possibility that the frequency of this genetic change increases in the population, leading progressively to reproductive isolation and fixation of the change. As future perspective, we are considering the possibility to apply a genome scan approach using gene-linked polymorphic markers in order to identify genes under selection to be related to both the presence of mutants and the population genetic patterns observed.
References
Andreakis N, Caputi L, Sordino P (2007) Characterization of highly polymorphic nuclear microsatellite loci from the ascidian Ciona intestinalis. Mol Ecol Notes 7:610–612
Ben-Shlomo R, Paz G, Rinkevich B (2006) Postglacialperiod and recent invasions shape the population genetics of botryllid ascidians along european atlantic coasts. Ecosystems 9:1118–1127
Boffelli D, Weer CV, Weng L, Lewis KD, Shoukry MI, Pachter L, Keys DN, Rubin EM (2004) Intraspecies sequence comparisons for annotating genomes. Genome Res 14:2406–2411
Caputi L, Andreakis N, Mastrototaro F, Cirino P, Vassillo SP, Sordino P (2007) Cryptic speciation in a model invertebrate chordate. Proc Natl Acad Sci USA 104:9364–9369
Caputi L, Andreakis N, Affinito O, Vassillo M, Procaccini G, Sordino P (submitted) Recent expansion and global divergence of Ciona intestinalis sp. A, a strong marine competitor
Caputi L, Borra M, Andreakis N, Biffali E, Sordino P (2008) SNPs and Hox gene mapping in Ciona intestinalis. BMC Genomics 9:39–50
Carman MR, Bullard SG, Donnelly JP (2007) Water quality, nitrogen pollution, and ascidian diversity in coastal waters of southern Massachusetts, USA. J Exp Mar Biol Ecol 342:175–178
Carver CE, Chisholm A, Mallet AL (2003) Strategies to mitigate the impact of Ciona intestinalis (L.) biofouling on shellfish production. J Shellfish Res 22:621–631
Charmantier A, Garant D (2005) Environmental quality and evolutionary potential: lessons from wild populations. Proc R Soc Lond B 272:1415–1425
Cirino P, Toscano A, Caramiello D et al (2002) Laboratory culture of the ascidian Ciona intestinalis (L.): a model system for molecular developmental biology research. Mar Mod Elec Rec. http://www.mbl.edu/html/BB/MMER/CIR/CirCon.html
Darling JA, Folino-Rorem NC (2009) Genetic analysis across different spatial scales reveals multiple dispersal mechanisms for the invasive hydrozoan Cordylophora in the Great Lakes. Mol Ecol 18:4827–4840
Dehal P, Satou Y, Campbell RK et al (2002) The draft genome of Ciona intestinalis: insights into chordate and vertebrate origins. Science 298:2157–2167
Dupont L, Viard F, Dowell MJ, Wood C, Bishop JDD (2009) Fine- and regional-scale genetic structure of the exotic ascidian Styela clava (Tunicata) in southwest England, 50 years after its introduction. Mol Ecol 18:442–453
Dupont L, Viard F, Davis MH, Nishikawa T, Bishop JDD (2010) Pathways of spread of the introduced ascidian Styela clava (Tunicata) in Northern Europe, as revealed by microsatellite markers. Biol Invasions 12:2707–2721
Dybern BI (1965) The life cycle of Ciona intestinalis (L.) f. typica in relation to the environmental temperature. Oikos 16:109–131
Everett RA (2000) Patterns and pathways of biological invasions. Trends Ecol Evol 15:177–178
Fields S, Johnston M (2005) Whither model organism research? Science 307:1885–1886
Fisher RA (1930) The genetical theory of natural selection. Oxford University Press, Oxford
Goldschmidt R (1940) The material basis of evolution. Yale University Press, New Haven
Goldstien SJ, Schiel DR, Gemmell NJ (2010) Regional connectivity and coastal expansion: differentiating pre-border and post-border vectors for the invasive tunicate Styela clava. Mol Ecol 19:874–885
Havenhand JN, Svane I (1991) Roles of hydrodynamics and larval behaviour in determining spatial aggregation in the tunicate Ciona intestinalis. Mar Ecol Prog Ser 68:271–276
Hedgecock D, Li G, Hubert S, Bucklin K, Ribes V (2004) Widespread null alleles and poor cross-species amplification of microsatellite DNA loci cloned from the Pacific oyster, Crassostrea gigas. J Shellfish Res 23:379–385
Hendrickson C, Christiaen L, Deschet K et al (2004) Culture of adult ascidians and ascidian genetics. Methods Cell Biol 74:143–170
Holland BS (2000) Genetics of marine bioinvasions. Hydrobiologia 420:63–71
Hoshino Z, Nishikawa T (1985) Taxonomic studies of Ciona intestinalis (L.) and its allies. Publ Seto Mar Biol Lab 30:61–79
Howes S, Herbinger CM, Darnell P, Vercaemer B (2007) Spatial and temporal patterns of recruitment of the tunicate Ciona intestinalis on a mussel farm in Nova Scotia, Canada. J Exp Mar Biol Ecol 342:85–92
Jeffery WR (2004) Evolution and development of brain sensory organs in molgulid ascidians. Evol Dev 6:170–179
Jesse R, Pfenninger M, Fratini S et al (2009) Disjunct distribution of the Mediterranean freshwater crab Potamon fluviatile - Natural expansion or human introduction? Biol Invasions 11:2209–2221
Joly JS, Kano S, Matsuoka T, Auger H, Hirayama K et al (2007) Culture of Ciona intestinalis in closed systems. Dev Dyn 236:1832–1840
Kano S (2007) Initial stage of genetic mapping in Ciona intestinalis. Dev Dyn 236:1768–1781
Kano S, Chiba S, Satoh N (2001) Genetic relatedness and variability in inbred and wild populations of the solitary ascidian Ciona intestinalis revealed by arbitrarily primed polymerase chain reaction. Mar Biotechnol 3:58–67
Kano S, Satoh N, Sordino P (2006) Primary genetic linkage maps of the ascidian, Ciona intestinalis. Zoolog Sci 23:31–39
Kawamura K, Fujita H, Nakauchi M (1987) Cytological characterization of self incompatibility in gametes of the ascidian, Ciona intestinalis. Dev Growth Diff 29:627–642
Lambert G (2001) A global overview of ascidian introductions and their possible impact on the endemic fauna. In: Sawada H, Yokosawa H, Lambert CC (eds) The biology of ascidians. Springer, Tokyo
Lambert G (2007) Invasive sea squirts: a growing global problem. J Exp Mar Biol Ecol 342:3–4
Lambert CC, Lambert G (1998) Non indigenous ascidians in southern California harbors and marinas. Mar Biol 130:675–688
Lee CE (2002) Evolutionary genetics of invasive species. Trends Ecol Evol 17:386–391
Marin MG, Bressan M, Beghi L, Brunetti R (1987) Thermo-aline tolerance of Ciona intestinalis (L., 1767) at different developmental stages. Cah Biol Mar 28:47–57
Marshall DJ, Keough MJ (2003) Effects of settler and density on early post-settlement survival of Ciona intestinalis in the field. Mar Ecol Prog Ser 259:139–144
Nei M, Maruyama T, Chakraborty R (1975) Bottleneck effect and genetic-variability in populations. Evolution 29:1–10
Nydam ML, Harrison R (2007) Genealogical relationships within and among shallow-water Ciona species (Ascidiacea). Mar Biol 151:1839–1847
Nydam ML, Harrison RG (2010) Polymorphism and divergence within the ascidian genus Ciona. Mol Phylogenet Evol 56:718–726
Nydam ML, Harrison RG (2011a) Introgression despite substantial divergence in a broadcast spawning marine invertebrate. Evolution 65:429–442
Nydam ML, Harrison RG (2011b) Reproductive protein evolution in two cryptic species of marine chordate. BMC Evol Biol 11:18
Oliveira Marins de F, Silva Oliveira da C, Viera Maciel NM, Skinner LF (2009) Reinclusion of Ciona intestinalis (Ascidiacea: Cionidae) in Brazil –a methodological view. JMBA2 – Biodivers Rec. 1–5
Palumbi SR (1992) Marine speciation on a small planet. Trends Ecol Evol 7:114–118
Pérès JM (1952) Recherches sur le cycle sexuel de «Ciona intestinalis (L.)». Arch Anat Microsc Morphol Exp 41:153–183
Petersen JK (2007) Ascidian suspension feeding. J Exp Mar Biol Ecol 342:127–137
Petersen JK, Svane I (1995) Larval dispersal in the ascidian Ciona intestinalis (L.). Evidence for a closed population. J Exp Mar Biol Ecol 186:89–102
Petersen JK, Schou O, Thor P (1995) Growth and energetics in the ascidian Ciona intestinalis. Mar Ecol Prog Ser 120:175–184
Pimentel D, Lach L, Zuniga R, Morrison D (2000) Environmental and economic costs of non-indigenous species in the United States. Bioscience 50:53–65
Procaccini G, Pischetola M, Di Lauro R (2000) Isolation and characterization of microsatellite loci in the ascidian Ciona intestinalis (L.). Mol Ecol 9:1924–1926
Ramsay A, Davidson J, Landry T, Arsenault G (2008) Process of invasiveness among exotic tunicates in Prince Edward Island, Canada. Biol Invasions 10:1311–1316
Ramsay A, Davidson J, Bourque D, Stryhn H (2009) Recruitment patterns and population development of the invasive ascidian Ciona intestinalis in Prince Edward Island, Canada. Aquat Invasions 4:169–176
Riisgard HU, Jensen AS, Jürgensen C (1998) Hydrography, near-bottom currents and grazing impact of the filter-feeding ascidian Ciona intestinalis in a Danish fjord. Ophelia 49:1–16
Rosati F, Santis RD (1978) Studies on fertilization in the ascidians. I. Self-sterility and specific recognition between gametes of Ciona intestinalis. Exp Cell Res 112:111–119
Sabbadin A (1958) Il ciclo biologico di Ciona intestinalis (L.), Molgula manhattensis (De Kay) e Styela plicata (Lesuer) nella laguna veneta. Arch Oceanogr Limnol 11:1–28
Sasakura Y, Oogai Y, Matsuoka T, Satoh N, Awazu S (2007) Transposon mediated transgenesis in a marine invertebrate chordate: Ciona intestinalis. Genome Biol 8(Suppl 1):S3
Satoh N (1994) Developmental biology of ascidians. Cambridge University Press, New York
Schmidt GH (1983) The hydroid Tubularia larynx causing “bloom” of the ascidians Ciona intestinalis and Ascidiella aspersa. Mar Ecol Prog Ser 12:103–105
Schmidtke J, Engel W (1980) Gene diversity in tunicate populations. Biochem Genet 18:503–508
Small KS, Brudno M, Hill MM, Sidow A (2007a) A haplome alignment and reference sequence of the highly polymorphic Ciona savignyi genome. Genome Biol 8:R41
Small KS, Brudno M, Hill MM, Sidow A (2007b) Extreme genomic variation in a natural population. Proc Natl Acad Sci USA 104:5698–5703
Sordino P, Andreakis N, Brown ER et al (2008) Natural variation of model mutant phenotypes in Ciona intestinalis. PLoS One 3:e2344
Sorte CJ, Williams SL, Carlton JT (2010) Marine range shifts and species introductions: comparative spread rates and community impacts. Global Ecol Biogeogr 19:303–316
Stinchcombe JR, Hoekstra HE (2007) Combining population genomics and quantitative genetics: finding the genes underlying ecologically important traits. Heredity 100:158–170
Suzuki M, Nishikawa T, Bird A (2005) Genomic approaches reveal unexpected genetic divergence within Ciona intestinalis. J Mol Evol 61:627–635
Svane I, Havenhand JN (1993) Spawning and dispersal in Ciona intestinalis (L.). P.S.Z.N.I. Mar Ecol 14:53–66
Therriault TW, Herborg L (2008a) A qualitative biological risk assessment for vase tunicate Ciona intestinalis in Canadian waters: using expert knowledge. ICES J Mar Sci 65:781–787
Therriault TW, Herborg L (2008b) Predicting the potential distribution of the vase tunicate Ciona intestinalis in Canadian waters: informing a risk assessment. ICES J Mar Sci 65:788–794
Veeman MT, Nakatani Y, Hendrickson C, Ericson V, Lin C, Smith WC (2008) chongmague reveals an essential role for laminin-mediated boundary formation in chordate convergence and extension movements. Development 135:33–41
Whitlatch RB, Osman RW, Frese A (1995) The ecology of two introduced marine ascidians and their effects of epifaunal organisms in Long Island Sound. In: Balcom N (ed) Proceedings of the Northeast Conference on Non-Indigenous Aquatic Nuisance Species: Reg Conf, pp 29–48
Wilson AJ, Pemberton JM, Pilkington JG et al (2006) Environmental coupling of selection and heritability limits evolution. PLoS Biol 4:e216
Zhan A, Bao Z, Hui M et al (2007) Inheritance pattern of EST-SSRs in self-fertilized larvae of the bay scallop Argopecten irradians. Ann Zoolog Fennici 44:259–268
Zhan A, Macisaac HJ, Cristescu ME (2010) Invasion genetics of the Ciona intestinalis species complex: from regional endemism to global homogeneity. Mol Ecol 19:4678–4694
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2011 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Procaccini, G., Affinito, O., Toscano, F., Sordino, P. (2011). A New Animal Model for Merging Ecology and Evolution. In: Pontarotti, P. (eds) Evolutionary Biology – Concepts, Biodiversity, Macroevolution and Genome Evolution. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-20763-1_6
Download citation
DOI: https://doi.org/10.1007/978-3-642-20763-1_6
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-20762-4
Online ISBN: 978-3-642-20763-1
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)