
Abstract
A longer growing season with climate change is expected to increase net primary productivity of many rangeland types, especially those dominated by grasses, although responses will depend on local climate and soil conditions. Elevated atmospheric carbon dioxide may increase water use efficiency and productivity of some species. In many cases, increasing wildfire frequency and extent will be damaging for big sagebrush and other shrub species that are readily killed by fire. The widespread occurrence of cheatgrass and other nonnatives facilitates frequent fire through annual fuel accumulation. Shrub species that sprout following fire may be quite resilient to increased disturbance, but may be outcompeted by more drought tolerant species over time.
Adaptation strategies for rangeland vegetation focus on increasing resilience of rangeland ecosystems, primarily through non-native species control and prevention. Ecologically based non-native plant management focuses on strategies to repair damaged ecological processes that facilitate invasion, and seeding of desired natives can be done where seed availability and dispersal of natives are low. Proactive management to prevent establishment of non-native species is also critical (early detection-rapid response), including tactics such as weed-free policies, education of employees and the public, and collaboration among multiple agencies to control weeds. Livestock grazing can also be managed through the development of site-specific indicators that inform livestock movement guides and allow for maintenance and enhancement of plant health.
Access provided by CONRICYT-eBooks. Download chapter PDF
Similar content being viewed by others


Keywords
- Rangelands
- Vulnerability
- Climate change
- Nonnative plants
- Adaptation
- Sagebrush
- Woodlands
- Grasslands
- Shrublands
6.1 Introduction
Rangelands , including grassland , shrubland , desert , alpine, and some woodland ecosystems, are dominated by grass, forb, or shrub species (Lund 2007). Rangelands occupy more than 26 million hectares in the Northern Rockies (Reeves and Mitchell 2011), producing forage for domestic and wild ungulates , providing critical habitat for numerous species such as greater sage-grouse (Centrocercus urophasianus), and providing many recreational opportunities.
Climate change, combined with residential development, energy development , and invasive (nonnative) species (e.g., cheatgrass , wild horses and burros), create a significant challenge for resource managers charged with ensuring sustainability of ecosystem services . The effects of climate change on rangelands have been studied less than effects on forests, but the effects of (past and future) human land-use activities on rangelands will probably exceed those of climate change, at least in the short term. This assessment focuses on regeneration success, response to disturbance (especially wildfire), and life history traits in rangelands , rather than on explicit estimates of future land-use change. The focus on life history traits combined with the concepts of resilience and resistance can help with understanding the effects of climate change. Resilience is the capacity of ecosystems to regain structure, processes, and function in response to disturbance (Holling 1973; Allen et al. 2005), whereas resistance is the capacity to retain these attributes in response to disturbance (Folke et al. 2004). These concepts are especially helpful for understanding establishment of nonnative plants and interactions between climate change stressors (Chambers et al. 2014), as demonstrated in Fig. 6.1, which shows that management for ecosystem services derived from rangelands will be most effective in mesic rangelands.

Resilience to disturbance (a) and resistance to cheatgrass (b) over a typical temperature/precipitation gradient in cold desert (modified from Chambers et al. 2014). Dominant ecological sites range from Wyoming big sagebrush on warm, dry sites (left); to mountain big sagebrush on cool, moist sites (middle); to mountain big sagebrush and root-sprouting shrubs on cold, moist sites (right). Resilience increases along the temperature/precipitation gradient, influenced by site characteristics (e.g., aspect). Resistance also increases along the gradient, influenced by disturbances and management treatments that alter vegetation structure and composition. ARTRw Wyoming big sagebrush (Artemisia tridentata ssp. wyomingensis ); ARTRv mountain big sagebrush (A. tridentata ssp. vaseyana ), SYOR mountain snowberry (Symphoricarpos oreophilus)
6.2 Rangeland Vegetation
This assessment focuses on vegetation types and individual species for which sufficient information and data exist to make inferences about the effects of climate change. First, we reviewed the extent of rangelands in the Northern Rockies and generally confined analysis to U.S. Forest Service (USFS) rangelands (Reeves and Mitchell 2011) in the Northern Rockies. We determined that the complexity of rangeland vegetation, combined with a paucity of climate change effects studies, suggests that a grouping of individual vegetation types into classes is appropriate. Therefore, we assessed the following vegetation classes:
-
Northern Great Plains —This vegetation class is broadly distributed, including a mixture of cool-season (C3) and warm-season (C4) grass species.
-
Montane shrubs—Includes a broad diversity of shrub species, many of which are important for browsing by native ungulates .
-
Montane grasslands —This relatively scarce vegetation class is dominated by cool-season species, often intermixed with forest vegetation.
-
Sagebrush systems—Dominated by species in the genus Artemisia , this is a ubiquitous and iconic vegetation class in much of the western United States, providing critical wildlife habitat for many species, including greater sage-grouse .
Sagebrush systems dominated by big sagebrushes (Wyoming big sagebrush [Artemisia tridentata ssp. wyomingensis ], mountain big sagebrush [A. t. ssp. vaseyana ], and basin big sagebrush [A. t. ssp. tridentata ]) have been widely studied, at least partially as a result of recent research on sage-grouse habitat. Therefore, inferences about the vulnerability of these sagebrush species to climate change is supported by relatively more information than for other species. Four sagebrush types were delineated for this assessment: big sagebrushes (Wyoming big sagebrush , basin big sagebush), low sagebrushes (low sagebrush [A. arbuscula ], black sagebrush [A. nova ]), sprouting sagebrushes (silver sagebrush [A. cana ], three-tip sagebrush [A. tripartita ]), and mountain big sagebrush .
Wyoming and basin big sagebrush types were aggregated because they have similar life histories, stature, and areal coverage in the Northern Rockies, and because they represent critical habitats for many species of animals. Basin big sagebrush occupies sites with relatively deeper soils that retain sufficient moisture for perennial bunchgrasses , suggesting these sites may be more resilient and resistant to a drier climate (Chambers et al. 2007). Silver sagebrush and three-tip sagebrush can resprout after fire, making them unique among the sagebrush species. Communities dominated by Wyoming big sagebrush are by far the most common and occupy the most area (Table 6.1), whereas the low sagebrush type occupies the least. Although basin and Wyoming sagebrush are common throughout the Northern Rockies, mountain big sagebrush communities occupy the greatest extent on lands managed by the USFS .
6.3 Management Issues
Fire regimes , improper grazing , and nonnative species are concerns for rangeland management in the Northern Rockies. Uncharacteristic fire regimes threaten most rangeland habitats, especially sagebrush steppe, across much of the western United States. On one hand, “too much fire” may affect the landscape relative to historical fire regimes, because many sagebrush habitats now have shortened fire return intervals , resulting in increasing dominance of nonnative invasive annual grasses that create fuel conditions that facilitate more frequent combustion (Chambers et al. 2007). On the other hand, fire exclusion has led to longer fire return intervals that may be responsible for conifer encroachment in montane grasslands (Arno and Gruell 1986) and higher elevation sagebrush habitats, especially those dominated by mountain big sagebrush (Heyerdahl et al. 2006) (Fig. 6.2).

Establishment of ponderosa pine ( Pinus ponderosa ) and other conifers in montane grassland dominated by rough fescue
The nonnative invasive species of greatest concern is cheatgrass ( Bromus tectorum ), although Japanese brome (B. japonicus ) and leafy spurge ( Euphorbia esula ) are also problems in the Northern Great Plains . Distribution of cheatgrass has expanded greatly in the western half of the Northern Rockies (Ramakrishnan et al. 2006; Merrill et al. 2012), and it is likely that further expansion may be enhanced by elevated atmospheric CO2 concentrations, increased soil disturbance, and increasing spring and winter temperatures (Chambers et al. 2014; Boyte et al. 2016; Bradley et al. 2016).
Improper grazing —the mismanagement of grazing that produces detrimental effects on vegetation or soil resources—can create additional stress in some rangelands , accelerating the annual grass invasion/fire cycle, especially in some sagebrush types, the Northern Great Plains , and montane grasslands . Fortunately, most U.S. rangelands are not improperly grazed to the point of degradation (Reeves and Mitchell 2011; Reeves and Baggett 2014), a generalization that is true for most rangelands in the Northern Rockies.
6.4 Assessing the Effects of Climate Change on Rangelands
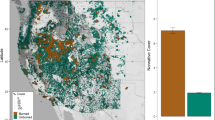
Despite the lack of focused studies on the effects of climate change on rangeland vegetation and the large uncertainty of projected climates, there are a few elements of climate change that are increasingly recognized as potential outcomes. First, projected temperature increases (Chap. 2) are expected to increase evaporative demand and pose greater overall stress (Polley et al. 2013). Projected changes in precipitation patterns and increasing potential evapotranspiration could facilitate more frequent wildfires through the combined effects of early-season plant growth and the desiccating effects of warmer summers (Morgan et al. 2008). These changes will lead to drier soils , particularly in summer when plants are physiologically active (Polley et al. 2013; Bradford et al. 2014; Palmquist et al. 2016a, b). However, winter precipitation is projected to increase 10–20% in the Northern Rockies (Chap. 2), which may compensate for increasing droughts . In addition, higher atmospheric CO2 may offset evaporative demand by increasing water use efficiency in plants. Relative to much of the rest of the United States, the Northern Rockies could experience an increase in annual net primary productivity (NPP) (Fig. 6.3), partially as a result of the likely increase in water use efficiency and increased growing season length (Reeves et al. 2014). Removal of growth limitations could result in significant changes in vegetation at higher elevations, such as the Greater Yellowstone Area . Higher NPP may seem counterintuitive because increased temperatures are associated with higher moisture stress and less favorable growing conditions. However, it is reasonable that high-elevation vegetation may experience increased production with increasing temperatures (Reeves et al. 2014), especially relatively mesic areas supporting mountain sagebrush . Increased atmospheric CO2 concentrations may modify physiological growth processes in rangeland vegetation by enhancing water use efficiency, but response may not be consistent across all vegetation (Morgan et al. 2004b, 2011; Woodward and Kelly 2008).

Mean slope of linear regression for the net primary productivity trend for low (B2), moderate (A1B), and high (A2) emission scenarios (global climate models averaged: GCGM2, HadCM3, CSIRO, MK2, MIROC3.2) (a), and standard deviation of the mean slope of linear regression of the net primary productivity trend for the same scenarios (b) (From Reeves et al. 2014)
Warmer winters and decreasing snowpack may have a significant effect on the distribution and abundance of different plant species. Minimum temperatures are expected to increase more than maximum temperatures, providing longer frost -free periods (Chap. 2). Warmer, wetter winters would favor early-season plant species and tap-rooted species that are able to access early-season soil water (Polley et al. 2013).
6.4.1 Montane Grasslands
Montane grasslands are associated with mountainous portions of the Northern Rockies, including the Palouse prairie and canyon grasslands of northern and central Idaho . Montane grasslands occur in intermountain valleys, foothills, and mountain slopes from low to relatively high elevation. They are dominated by cool-season (C3) grasses , many forbs, upland sedges, and scattered trees in some areas. Dominant species include bluebunch wheatgrass ( Pseudoroegneria spicata ), rough fescue ( Festuca campestris ), Idaho fescue (F. idahoensis ), Sandberg bluegrass ( Poa secunda ), needle-and-thread ( Hesperostipa comata ), western wheatgrass ( Pascopyrum smithii ), prairie junegrass ( Koeleria macrantha ), western needlegrass ( Achnatherum nelsonii ), and Richardson’s needlegrass (A. richardsonii ).
Most grasslands , particularly at lower elevations, are disturbed, fragmented, and often occupied by nonnative plant species. Improper livestock grazing , native ungulate herbivory , and nonnative plants are stressors in these grasslands . Lack of fire is also a stressor, because it can allow conifers to become established within grasslands (Arno and Gruell 1986; Heyerdahl et al. 2006). As conifer density increases with fire exclusion , grass cover declines, because most grassland species are shade intolerant. However, if fires become hotter and more frequent, there is an increased risk of mortality of native species and invasion by nonnative species (Ortega et al. 2012). As noted above, cheatgrass creates continuous fine fuels that are combustible by early summer; if fire occurs at this time, it can burn native perennial grasses before they have matured and set seed (Chambers et al. 2007; Bradley 2008). Several other nonnative species can also increase after fire, reducing native plant cover.
Nonnative plant species will probably expand in lower elevation grasslands as temperature increases, resistance to invasion decreases (Chambers et al. 2014), and disturbance increases (Bradley 2008). Drier conditions plus ungulate effects (grazing , browsing, hoof damage) may increase bare ground and possibly surface soil erosion . Low-elevation grasslands may have increasing dominance of more drought -tolerant species, such that cool-season species decline and warm-season species expand (Bachelet et al. 2001). However, elevated CO2 favors C3 grasses and enhances biomass production, whereas warming favors C4 grasses (Morgan et al. 2004a, 2007). Therefore, a warmer and drier climate may allow C4 grasses to expand westward, displacing some C3 species. In general, a warmer climate with more frequent fires will favor increasing dominance of grasslands across the landscape, in some cases displacing shrublands and conifers that are burned too frequently to regenerate successively.
6.4.2 Montane Shrubs
Montane shrubs are associated with montane and subalpine forests , occurring as large patches within forested landscapes. Rocky Mountain maple (Acer glabrum ), oceanspray (Holidiscus discolor ), tobacco brush (Ceanothus velutintis var. velutinus), Sitka alder (Alnus viridus subsp. sinuata ), thimbleberry (Rubus parviflorus ), chokecherry (Prunus virginiana ), serviceberry (Amelanchier alnifolia ), currant (Ribes spp.), snowberry ( Symphoricarpos albus ), Scouler willow ( Salix scouleriana ), and mountain ash ( Sorbus scopulina ) are common.
Montane shrubs persist in locations where disturbance kills aboveground stems, with subsequent sprouting from the root crown, rhizomes, and roots where adequate light and soil moisture are available. Fire exclusion , conifer establishment, and browsing by native and domestic animals are significant stressors. Most mesic shrubs are well adapted to frequent fire and can often compete well with associated conifers. However, even sprouting shrubs can sometimes be killed if fires are very hot and postfire weather is dry.
Drier soils and increased fire frequency may facilitate increasing dominance of more drought tolerant species such as rubber rabbitbrush ( Ericameria nauseosa ), green rabbitbrush ( Chrysothamnus viscidiflorus ), and spineless horsebrush ( Tetradymia canescens ). Nonnative plant species may also expand into these communities, particularly following fire (Bradley 2008). Some mesic shrub species (e.g., Sitka alder , Rocky Mountain maple ) may persist at higher elevations or in cooler, moister locations (e.g., north aspects, concavities).
6.4.3 Short Sagebrushes
Low sagebrush ecosystems cover only about 1% of total sagebrush habitat in the Northern Rockies, half of which is in the Western Rockies subregion. Low sagebrush sites have relatively low productivity , and are located between 1800 and 2700 m elevation in Montana and Idaho , occupying shallow claypan soils that restrict drainage and root growth . Black sagebrush is found on shallow, dry, infertile soils . Stressors include nonnative species and improper use by livestock .
Low and black sagebrush have a more limited distribution than other sagebrush species and depend on seeding for regeneration , so their distribution could be further restricted in a warmer climate, resulting in a patchier mosaic of remnant communities (West and Mooney 1972). Several traits make low sagebrush sensitive to climate change, including high mortality in the first year of growth (Shaw and Monsen 1990), which may be exacerbated if erosion increases from drought -induced reductions of plant cover. If unfavorable conditions for seeding persist following disturbance, low sagebrush may disappear from some sites, especially if annual grasses increase at the same time.
Increased fire will have negative consequences for low and black sagebrush , which are intolerant of fire and do not resprout. Fire return intervals vary considerably among communities dominated by low sagebrush . In the Greater Yellowstone Area subregion, vegetation modeling results indicate that the proportion of landscape burned will increase substantially, with the potential for fire to reach some low sagebrush communities (Sheehan et al. 2015). Increased fire activity will decrease the abundance of low sagebrush relative to other species, especially if nonnative annual grasses become more prevalent.
Relative to other sagebrush species, low and black sagebrush have limited adaptive capacity . Black sagebrush hybridizes with silver sagebrush , and sprouting is thought to be a heritable trait in crosses between non-sprouting and sprouting sagebrushes (McArthur 1994). However, silver sagebrush distributions are typically disjunct with those of low and black sagebrush, so acquisition of sprouting traits is unlikely. The relatively low productivity of low sagebrush sites may also limit adaptive capacity , especially if other risk factors are present.
6.4.4 Sprouting Sagebrush Species
Significant areas of threetip and silver sagebrush shrublands have been converted to agricultural lands. Those that remain are often used for livestock grazing because of the palatable herbaceous undergrowth . Rangelands with improper grazing typically have a large amount of bare ground, low vigor of native herbaceous species, and presence of nonnative plant species. Improper livestock grazing can cause loss of topsoil if vegetation cover and density decline and bare ground increases (Sheatch and Carlson 1998).
Both species can sprout from the root crown following top kill from fire (Bunting et al. 1987); silver sagebrush is a vigorous sprouter (Rupp et al. 1997), whereas threetip sagebrush is less vigorous (Bunting et al. 1987; Akinsoji 1988). Both species occur on mesic sites, where threetip sagebrush is often associated with mountain big sagebrush , and silver sagebrush occupies riparian benches or moist toe slopes. Although these species sprout, increased fire frequency and severity (particularly in threetip communities) may facilitate dominance by fire-adapted shrubs, herbs, and nonnative species . More spring and winter precipitation would promote establishment and early seed set in nonnative annual grasses , providing a competitive advantage over native perennial grasses (Bradley 2008) and creating fine fuels that can burn sagebrush and native grasses before they have matured and set seed (Chambers and Pellant 2008).
Historical fire return intervals for both species are relatively short, with threetip sagebrush cover returning to preburn levels 30–40 years after fire (Barrington et al. 1988). All three subspecies of silver sagebrush sprout after fire, and along with threetip, also occur on more mesic sites. If high-severity burns are more frequent in a warmer climate, they may not only cause mortality, but create unfavorable conditions for postfire regeneration (from sprouting or seed), and provide invasive species with a competitive edge. Understory composition may shift to more xeric grassland species (e.g., bluebunch wheatgrass , needle-and-thread ) that are better adapted to drier conditions. In a warmer climate, both sagebrush species may persist in sites that retain sufficient moisture (e.g., higher elevation, north aspect, concavities).
6.4.5 Wyoming Big Sagebrush and Basin Big Sagebrush
Distribution of Wyoming big sagebrush is patchy in Montana and more evenly distributed in the Eastern Rockies and Grassland subregions. Stressors to both sagebrush communities include improper livestock grazing , native ungulate herbivory , and nonnative invasive plants . Loss of topsoil can occur if vegetation cover and density decline and bare ground increases (e.g., with ungulate impacts). Wyoming big sagebrush habitat coincides with oil and gas development , especially on the eastern edge of its distribution. The Grassland and Greater Yellowstone Area subregions contain the largest extent of big sagebrush, with basin big sagebrush dominant in the Western Rockies subregion.
Big sagebrush ecosystems have been subjected to many stressors: oil and gas development (Knick et al. 2003), big sagebrush removal to increase livestock forage , plant pathogens and insects , improper grazing (Davies et al. 2011), nonnative invasive species (Davies et al. 2011), and altered disturbance regimes (Balch et al. 2013). These stressors, especially oil and gas development , cause habitat loss and fragmentation (Doherty et al. 2008; Walston et al. 2009), creating barriers to plant dispersal and degrading habitat for sage-grouse and other wildlife species (Rowland et al. 2006). Improper use by livestock alters structure and composition of big sagebrush communities and increases the probability of nonnative annual grass invasion (Cooper et al. 2007; Davies et al. 2011), especially cheatgrass (Brooks et al. 2004; Balch et al. 2013). Cheatgrass invasion poses a continued and heightened threat to big sagebrush ecosystems in the future, because its biomass production and fire frequency are projected to increase in response to rising temperature and CO2 levels (Ziska et al. 2005). Field brome ( Bromus arvensis ) can also negatively affect big sagebrush, because it establishes after fires that kill big sagebrush (Cooper et al. 2007).
Amount and timing of precipitation control seedling establishment of big sagebrush at low elevation, whereas minimum temperature and snow depth control germination and survival at high elevation (Poore et al. 2009; Nelson et al. 2014). If drought increases in the future, germination and survival of big sagebrush seedlings may decrease (Schlaepfer et al. 2014a, b). Drought and increased summer temperature can also affect survival and growth of mature big sagebrush plants (Poore et al. 2009), as well as perennial grasses and forbs. In addition, big sagebrush seeds have low viability after 2 years (Wijayratne and Pyke 2009, 2012), are poorly dispersed (Young et al. 1989), and are episodically produced (Young et al. 1989). Big sagebrush is a poor competitor relative to associated herbaceous species (Schlaepfer et al. 2014a), and because it is killed by fire, postfire recovery may be challenging if wildfires are more frequent in the future.
Big sagebrush is projected to expand in northeast and north-central Montana , where climate may be sufficiently cool and moist (Schrag et al. 2011), and decrease in the Western Rockies and northwest Greater Yellowstone Area subregions, primarily from summer drought (Schlaepfer et al. 2012). Projected shifts in community composition and productivity in big sagebrush ecosystems remain uncertain. If drought increases, native herbaceous plant diversity and cover may be reduced. In non-drought years, higher temperatures and increased CO2 may increase biomass production (Reeves et al. 2014), fire frequency , and herbaceous biomass at the expense of fire-intolerant big sagebrush.
Although lower soil water availability may pose a long-term stress for big sagebrush ecosystems, extended periods of sustained drought are required to cause mortality (Kolb and Sperry 1999). Big sagebrush should have some capacity to adapt to climate change. The species occurs over a large geographic area with diverse topography, soils, and climate, suggesting that it can persist in a broad range of ecological conditions. In addition, various subspecies of big sagebrush hybridize and have a high level of polyploidy, providing capacity to undergo selection and adapt to variable climate (e.g., Poore et al. 2009).
6.4.6 Mountain Big Sagebrush
Some areas of mountain big sagebrush shrublands have been converted to agricultural lands, and those that remain are used for domestic livestock grazing , primarily because of the palatable herbaceous undergrowth . Those that have had improper grazing typically have bare ground and low vigor of native herbaceous species, and as a result, nonnative plant species are often present. Improper livestock grazing, native ungulate herbivory , and nonnative invasive plants are the primary stressors. Fire exclusion is also a stressor, facilitating conifer establishment and decline of grass cover.
Mountain big sagebrush is killed by fire. If wildfire frequency and severity increase, community composition will shift to dominance by fire-adapted shrub, herbaceous, and nonnative species . Increased spring and winter precipitation may facilitate nonnative annual grasses (especially cheatgrass ) establishment, although this is less likely in cooler locations compared to lower elevation Wyoming and basin big sagebrush . Concerns about cheatgrass and other nonnative species have been discussed above, although some sagebrush communities may be less susceptible to cheatgrass invasion following fire (Lavin et al. 2013).
Mountain big sagebrush is not fire adapted, and may decline in cover and density or become extirpated in response to warmer temperatures and increased fire frequency and severity. Historical fire return intervals in mountain big sagebrush were a few decades, compared to Wyoming big sagebrush (>100 years) (Heyerdahl et al. 2006; Lesica et al. 2007). Mountain big sagebrush regenerates from seeds, with full recovery 15–40 years after fire (Bunting et al. 1987). Because the sagebrush seed bank is minimal, if fires burn large areas and there are no live, seed-bearing sagebrush nearby, there may be a conversion to grassland . In addition, nonnative species may expand into these areas or increase in abundance (Bradley 2008).
Mountain big sagebrush occurs at higher elevations, typically on more productive cooler, mesic sites that are less susceptible to nonnative species . If these sites become warmer and drier, herbaceous understory composition could shift to more drought -tolerant species, including cheatgrass (Chambers et al. 2014). Mountain big sagebrush may be able to persist and expand into cooler locations (higher elevation, north aspects, concavities, deeper soils ). Native grassland species that are more tolerant of warmer, drier conditions (e.g., bluebunch wheatgrass , needle-and-thread ) may also be able to persist in the understory.
6.4.7 Northern Great Plains
Grasslands extend across the northern Great Plains , from the foothill grasslands along the east slope of the northern and central Rocky Mountains in Montana to the Red River basin in eastern North Dakota . Annual precipitation increases from west to east, with a concurrent transition from shortgrass prairie to northern mixed grass prairie to tallgrass prairie . Shortgrass prairie is characterized by grama (Bouteloua spp. )/needlegrass/wheatgrass and a mix of C3 and C4 plant species. Northern mixed grass prairie is characterized by wheatgrass/needlegrass in the west and wheatgrass/bluestem (Andropogon spp. )/needlegrass in the east, and a mix of C3 and C4 plant species. Tallgrass prairie is characterized by bluestem and a dominance of C4 grasses, although C3 grass species are also present.
Historically, frequent wildfire maintained grassland dominance, particularly in the eastern Great Plains . Starting in the late nineteenth century, settlement altered fire regimes by reducing fire frequency and changing the seasonality of fire. The predominant land use and land cover changed from grasslands to crop agriculture and domestic livestock production, affecting the continuity of fuels and fire spread. Reduced fire has encouraged woody plant encroachment, especially in the eastern Great Plains (Morgan et al. 2008). Invasive grass and forb species have reduced the diversity of native grasslands, with increased noxious weeds such as leafy spurge , Kentucky bluegrass ( Poa pratensis ), Japanese brome , and cheatgrass . Energy development and associated infrastructure fragments grassland locally, and roads and vehicles help spread nonnative species .
Soil water availability affects plant species distribution and abundance, productivity , and associated social and economic systems of the northern Great Plains . Interactions of temperature, precipitation, topography, soil, and ambient CO2 with plant physiological mechanisms will influence how grasslands respond to climate change (Chen et al. 1996; Bachman et al. 2010; Morgan et al. 2011). Slope, aspect, insolation, and soil water holding capacity will modify these effects locally. Landscape variability in available soil water will result in uneven patterns of vegetation change and productivity . Elevated CO2 may counter the effects of higher temperatures and evaporative demand by increasing water use efficiency of some plant species, especially C3 grasses (Morgan et al. 2011), although most nonnative invasive species are C3 plants, and expansion of nonnatives would be a negative outcome (Morgan et al. 2008).
The adaptive capacity of Great Plains grasslands was demonstrated in the Central Plains during the 1930s and 1950s droughts (Weaver 1968). There was a shift in C4 grasses, in which big bluestem ( Andropogon gerardii ) and little bluestem ( Schizachyrium scoparium ) were replaced by the shortgrass species blue grama ( Bouteloua gracili ) and buffalograss ( Bouteloua dactyloides ). Shifts from tallgrass prairie to mixed grass prairie were also documented with an increase in two C3 plants, western wheatgrass and needlegrass. This shift was later reversed during the higher precipitation period of the 1940s, indicating significant resilience of Great Plains grasslands to drought.
6.5 Adapting Rangeland Vegetation and Management to Climate Change
Rangeland vegetation in the northern Rockies will be affected by altered fire regimes , increased drought , and increased establishment of nonnative invasive species in a changing climate. Effects of climate change will compound existing stressors caused by human activities. Therefore, adaptation options for rangeland vegetation are focused on increasing the resilience of rangeland ecosystems, primarily through nonnative invasive species control and prevention.
Ecologically based invasive plant management (EBIPM ) (Krueger-Mangold et al. 2006; Sheley et al. 2006) provides a framework for controlling nonnative species in rangelands . EBIPM focuses on strategies to repair damaged ecological processes that facilitate establishment of nonnatives (James et al. 2010). For example, prescribed fire treatments can be used where fire regimes have been altered, and seeding of desired natives can be done where seed availability and dispersal of natives is low.
Another adaptation strategy is to increase proactive management actions to prevent establishment of nonnative species . Early detection/rapid response (EDRR) is commonly used to prevent nonnative species establishment. Other tactics include implementing weed-free policies, conducting outreach to educate employees and the public about nonnatives (e.g., teach people to clean their boots), and developing weed management areas that are collaboratively managed by multiple agencies, non-governmental organizations, and the public.
Grazing management will be important in maintaining and increasing resilience of rangelands to climate change. A warmer climate will lead to altered availability of forage , requiring some reconsideration of grazing strategies. For example, reducing grazing in July and August may encourage growth of desired perennials in degraded systems. Livestock grazing can also be managed through development of site-specific, within-season triggers and end-point indicators that would inform livestock movement guides and allow for the maintenance and enhancement of plant health.
A changing climate has led to a decline of pollinators in some rangeland systems and may lead to phenological mismatches between pollinators and host plants. Pollinator declines may negatively affect the health of grasslands in the Northern Rockies, so encouraging native pollinators may help sustain these systems. Tactics that promote native pollinators include revegetation with native species, appropriate herbicide and insecticide use, and education. Implementing long-term monitoring of pollinators can help identify where treatments should be prioritized.
Existing stressors in montane shrublands include fire exclusion and conifer establishment, browsing by native and domestic ungulates , and insects and disease . Warmer temperatures and drier conditions may lead to an increase in high-severity fires that can cause extirpation of characteristic species and local soil erosion . Adaptation tactics include implementing fuel reduction projects such as brush cutting, slashing, mastication, and targeted browsing. Reestablishing appropriate fire regimes may help maintain these shrublands and increase their resilience to a warmer climate. EDRR and EBIPM can be used to control nonnatives and to maintain adequate shrub cover, vigor, and species richness. Educating specialists on ecology and disturbances affecting shrublands, effects of repeated burns, reforestation needs, and reporting on weeds will also help maintain these systems.
References
Akinsoji, A. (1988). Postfire vegetation dynamics in a sagebrush steppe in southeastern Idaho, USA. Vegetatio, 78, 151–155.
Allen, C. R., Gunderson, L., & Johnson, A. R. (2005). The use of discontinuities and functional groups to assess relative resilience in complex systems. Ecosystems, 8, 958–966.
Arno, S., & Gruell, G. (1986). Douglas-fir encroachment into mountain grasslands in southwestern Montana. Journal of Range Management, 39, 272–276.
Bachelet, D., Neilson, R. P., Lenihan, J. M., & Drapek, R. J. (2001). Climate change effects on vegetation distribution and carbon budget in the United States. Ecosystems, 4, 164–185.
Bachman, S., Heisler-White, J. L., Pendall, E., et al. (2010). Elevated carbon dioxide alters impacts of precipitation pulses on ecosystem photosynthesis and respiration in a semi-arid grassland. Oecologia, 162, 791–802.
Balch, J. K., Bradley, B. A., D’Antonio, C. M., & Gómez-Dans, J. (2013). Introduced annual grass increases regional fire activity across the arid western USA (1980–2009). Global Change Biology, 19, 173–183.
Barrington, M., Bunting, S., & Wright, G. (1988). A fire management plan for Craters of the Moon National Monument, Cooperative Agreement CA-9000-8-0005. Moscow: University of Idaho, Range Resources Department.
Boyte, S. P., Wylie, B. K., & Major, D. J. (2016). Cheatgrass percent cover change: Comparing recent estimates to climate change driven predictions in the Northern Great Basin. Rangeland Ecology and Management, 69, 265–279.
Bradford, J., Schlaepfer, D., & Lauenroth, W. (2014). Ecohydrology of adjacent sagebrush and lodgepole pine ecosystems: The consequences of climate change and disturbance. Ecosystems, 17, 590–605.
Bradley, B. A. (2008). Regional analysis of the impacts of climate change on cheatgrass invasion shows potential risk and opportunity. Global Change Biology, 14, 1–13.
Bradley, B. A., Curtis, C. A., & Chambers, J. C. (2016). Bromus response to climate and projected changes with climate change. In M. J. Germino (Ed.), Exotic brome-grasses in arid and semiarid ecosystems of the Western US. Cham: Springer International Publishing.
Brooks, M. L., D’Antonio, C. M., Richardson, D. M., et al. (2004). Effects of invasive alien plants on fire regimes. Bio Science, 54, 677–688.
Bunting, S. C., Kilgore, B. M., & Bushey, C. L. (1987). Guidelines for prescribed burning sagebrush-grass rangelands in the northern Great Basin, General Technical Report INT-231. Ogden: U.S. Forest Service, Intermountain Research Station.
Chambers, J. C., & Pellant, M. (2008). Climate change impacts on northwestern and intermountain United States rangelands. Rangelands, 30, 29–33.
Chambers, J. C., Roundy, B. A., Blank, R. R., et al. (2007). What makes Great Basin sagebrush ecosystems invasible to Bromus tectorum? Ecological Monographs, 77, 117–145.
Chambers, J. C., Bradley, B. A., Brown, C. A., et al. (2014). Resilience to stress and disturbance, and resistance to Bromus tectorum L. invasion in the cold desert shrublands of western North America. Ecosystems, 17, 360–375.
Chen, D., Hunt, H. W., & Morgan, J. A. (1996). Responses of a C3 and C4 perennial grass to CO2 enrichment and climate change: Comparison between model predictions and experimental data. Ecological Modelling, 87, 11–27.
Cooper, S. V., Lesica, P., & Kudray, G. M. (2007). Postfire recovery of Wyoming big sagebrush shrub-steppe in central and southeast Montana. Helena: Bureau of Land Management, State Office, Montana Natural Heritage Program.
Davies, K. W., Boyd, C. S., Beck, J. L., et al. (2011). Saving the sagebrush sea: An ecosystem conservation plan for big sagebrush plant communities. Biological Conservation, 144, 2573–2584.
Doherty, K. E., Naugle, D. E., Walker, B. L., & Graham, J. M. (2008). Greater sage-grouse winter habitat selection and energy development. Journal of Wildlife Management, 72, 187–195.
Folke, C., Carpenter, S., Walker, B., et al. (2004). Regime shifts, resilience, and biodiversity in ecosystem management. Annual Review of Ecology, Evolution, and Systematics, 33, 557–581.
Heyerdahl, E. K., Miller, R. F., & Parson, R. A. (2006). History of fire and Douglas-fir establishment in a savanna and sagebrush-grassland mosaic, southwestern Montana, USA. Forest Ecology and Management, 230, 107–118.
Holling, C. S. (1973). Resilience and stability in ecological systems. Annual Review of Ecology and Systematics, 4, 1–23.
James, J. J., Smith, B. S., Vasquez, E. A., & Sheley, R. L. (2010). Principles for ecologically based invasive plant management. Invasive Plant Science and Management, 3, 229–239.
Knick, S. T., Dobkin, D. S., Rotenberry, J. T., et al. (2003). Teetering on the edge or too late? Conservation and research issues for avifauna or sagebrush habitats. The Condor, 105, 611–634.
Kolb, K. J., & Sperry, J. S. (1999). Differences in drought adaptation between subspecies of sagebrush (Artemisia tridentata). Ecology, 80, 2373–2384.
Krueger-Mangold, J. M., Sheley, R. L., & Svejcar, T. J. (2006). Toward ecologically-based invasive plant management on rangeland. Weed Science, 54, 597–605.
Lavin, M., Brummer, T., Quire, J., et al. (2013). Physical disturbance shapes vascular plant diversity more profoundly than fire in the sagebrush steppe of southeastern Idaho, U.S.A. Ecology and Evolution, 3, 1626–1641.
Lesica, P., Cooper, S. V., & Kudray, G. (2007). Recovery of big sagebrush following fire in southwest Montana. Rangeland Ecology and Management, 60, 261–269.
Lund, G. H. (2007). Accounting for the worlds rangelands. Rangelands, 29, 3–10.
McArthur, E. D. (1994). Ecology, distribution, and values of sagebrush within the Intermountain region. In: S. B. Monsen, & S. G. Kitchen (Comps.), Proceedings—Ecology and management of annual rangelands (General Technical Report INT-GTR-313, pp. 347–351). Ogden: U.S. Forest Service, Intermountain Research Station.
Merrill, K. R., Meyer, S. E., & Coleman, C. E. (2012). Population genetic analysis of Bromus tectorum (Poaceae) indicates recent range expansion may be facilitated by specialist genotypes. American Journal of Botany, 99, 529–537.
Morgan, J. A., Mosier, A. R., Milchunas, D. G., et al. (2004a). CO2 enhances productivity, alters species composition, and reduces digestibility of short grass steppe vegetation. Ecological Applications, 14, 208–219.
Morgan, J. A., Pataki, D. E., Körner, C., et al. (2004b). Water relations in grassland and desert ecosystems exposed to elevated atmospheric CO2. Oecologia, 140, 11–25.
Morgan, J. A., Milchunas, D. G., LeCain, D. R., et al. (2007). Carbon dioxide enrichment alters plant community structure and accelerates shrub growth in the short grass steppe. Proceedings of the National Academy of Sciences, USA, 104, 14724–14729.
Morgan, J. A., Derner, J. D., Milchunas, D. G., & Pendall, E. (2008). Management implications of global change for Great Plains rangelands. Rangelands, 30, 18–22.
Morgan, J. A., LeCain, D. R., Pendall, E., et al. (2011). C4 grasses prosper as carbon dioxide eliminates desiccation in warmed semi-arid grassland. Nature, 476, 202–206.
Nelson, Z. J., Weisberg, P. J., & Kitchen, S. G. (2014). Influence of climate and environment on postfire recovery of mountain big sagebrush. International Journal of Wildland Fire, 23, 131–142.
Ortega, Y., Pearson, D. E., Waller, L. P., et al. (2012). Population-level compensation impedes biological control of an invasive forb and indirect release of a native grass. Ecology, 93, 783–792.
Palmquist, K. A., Schlaepfer, D. R., Bradford, J. B., & Lauenroth, W. K. (2016a). Spatial and ecological variation in dryland ecohydrological responses to climate change: Implications for management. Ecosphere, 7, e01590.
Palmquist, K. A., Schlaepfer, D. R., Bradford, J. B., & Lauenroth, W. K. (2016b). Mid-latitude shrub steppe plant communities: Climate change consequences for soil water resources. Ecology, 97, 2342–2354.
Polley, H. W., Briske, D. D., Morgan, J. A., et al. (2013). Climate change and north American rangelands: Trends, projections, and implications. Rangeland Ecology and Management, 66, 493–511.
Poore, R. E., Lamanna, C. A., Ebersole, J. J., & Enquist, B. J. (2009). Controls on radial growth of mountain big sagebrush and implications for climate change. Western North American Naturalist, 69, 556–562.
Ramakrishnan, A. P., Meyer, S. E., Fairbanks, D. J., & Coleman, C. E. (2006). Ecological significance of microsatellite variation in western north American populations of Bromus tectorum. Plant Species Biology, 21, 61–73.
Reeves, M. C., & Baggett, L. S. (2014). A remote sensing protocol for identifying rangelands with degraded productive capacity. Ecological Indicators, 43, 172–182.
Reeves, M. C., & Mitchell, J. E. (2011). Extent of coterminous U.S. rangelands: Quantifying implications of differing agency perspectives. Rangeland Ecology and Management, 64, 1–12.
Reeves, M., Moreno, A., Bagne, K., & Running, S. W. (2014). Estimating the effects of climate change on net primary production of US rangelands. Climatic Change, 126, 429–442.
Rowland, M. M., Wisdom, M. J., Spring, L. H., & Meinke, C. W. (2006). Greater sage-grouse as an umbrella species for sagebrush-associated vertebrates. Biological Conservation, 129, 323–335.
Rupp, L., Roger, K., Jerrian, E., & William, V. (1997). Shearing and growth of five intermountain native shrub species. Journal of Environmental Horticulture, 15, 123–125.
Schlaepfer, D. R., Lauenroth, W. K., & Bradford, J. B. (2012). Effects of ecohydrological variables on current and future ranges, local suitability patterns, and model accuracy in big sagebrush. Ecography, 35, 374–384.
Schlaepfer, D. R., Lauenroth, W. K., & Bradford, J. B. (2014a). Modeling regeneration responses of big sagebrush (Artemisia tridentata) to abiotic conditions. Ecological Modelling, 286, 66–77.
Schlaepfer, D. R., Lauenroth, W. K., & Bradford, J. B. (2014b). Natural regeneration processes in big sagebrush (Artemisia tridentata). Rangeland Ecology and Management, 67, 344–357.
Schrag, A., Konrad, S., Miller, B., et al. (2011). Climate-change impacts on sagebrush habitat and West Nile virus transmission risk and conservation implications for greater sage-grouse. Geo Journal, 76, 561–575.
Shaw, N. L., & Monsen, S. B. (1990). Use of sagebrush for improvement of wildlife habitat. In H. G. Fisser (Ed.), Wyoming shrublands: aspen, sagebrush and wildlife management. Proceedings, 17th Wyoming shrub ecology workshop (pp. 19–35). Laramie: University of Wyoming, Department of Range Management.
Sheatch, G. W., & Carlson, W. T. (1998). Impact of cattle treading on hill land. 1. Soil damage patterns and pasture status. New Zealand Journal of Agricultural Research, 41, 271–278.
Sheehan, T., Bachelet, D., & Ferschweiler, K. (2015). Projected major fire and vegetation changes in the Pacific Northwest of the conterminous United States under selected CMIP5 climate futures. Ecological Modelling, 317, 16–29.
Sheley, R. L., Mangold, J. M., & Anderson, J. L. (2006). Potential for successional theory to guide restoration of invasive-plant-dominated rangeland. Ecological Monographs, 76, 365–379.
Walston, L. J., Cantwell, B. L., & Krummel, J. R. (2009). Quantifying spatiotemporal changes in a sagebrush ecosystem in relation to energy development. Ecography, 32, 943–952.
Weaver, J. E. (1968). Prairie plants and their environment: A fifty-year study in the Midwest. Lincoln: University of Nebraska Press.
West, M., & Mooney, H. A. (1972). Photosynthetic characteristics of three species of sagebrush as related to their distribution patterns in the White Mountains of California. American Midland Naturalist, 88, 479–484.
Wijayratne, U. C., & Pyke, D. A. (2009). Investigating seed longevity of big sagebrush (Artemisia tridentata), Open-file Report 2009-1146. Reston: U.S. Geological Survey.
Wijayratne, U. C., & Pyke, D. A. (2012). Burial increases seed longevity of two Artemisia tridentata (Asteraceae) subspecies. American Journal of Botany, 99, 438–447.
Woodward, F. I., & Kelly, C. K. (2008). Responses of global plant diversity capacity to changes in carbon dioxide concentration and climate. Ecological Letters, 11, 1229–1237.
Young, J. A., Evans, R. A., & Palmquist, D. E. (1989). Big sagebrush (Artemisia tridentata) seed production. Weed Science, 37, 47–53.
Ziska, L. H., Reeves, J. B., & Blank, B. (2005). The impact of recent increases in atmospheric CO2 on biomass production and vegetative retention of Cheatgrass (Bromus tectorum): Implications for fire disturbance. Global Change Biology, 11, 1325–1332.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2018 Springer International Publishing AG
About this chapter
Cite this chapter
Reeves, M.C. et al. (2018). Effects of Climate Change on Rangeland Vegetation in the Northern Rockies. In: Halofsky, J., Peterson, D. (eds) Climate Change and Rocky Mountain Ecosystems. Advances in Global Change Research, vol 63. Springer, Cham. https://doi.org/10.1007/978-3-319-56928-4_6
Download citation
DOI: https://doi.org/10.1007/978-3-319-56928-4_6
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-56927-7
Online ISBN: 978-3-319-56928-4
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)