
Abstract
Sagebrush steppe and lodgepole pine forests are two of the most widespread vegetation types in the western United States and they play crucial roles in the hydrologic cycle of these water-limited regions. We used a process-based ecosystem water model to characterize the potential impact of climate change and disturbance (wildfire and beetle mortality) on water cycling in adjacent sagebrush and lodgepole pine ecosystems. Despite similar climatic and topographic conditions between these ecosystems at the sites examined, lodgepole pine, and sagebrush exhibited consistent differences in water balance, notably more evaporation and drier summer soils in the sagebrush and greater transpiration and less water yield in lodgepole pine. Canopy disturbances (either fire or beetle) have dramatic impacts on water balance and availability: reducing transpiration while increasing evaporation and water yield. Results suggest that climate change may reduce snowpack, increase evaporation and transpiration, and lengthen the duration of dry soil conditions in the summer, but may have uncertain effects on drainage. Changes in the distribution of sagebrush and lodgepole pine ecosystems as a consequence of climate change and/or altered disturbance regimes will likely alter ecosystem water balance.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Climatic conditions and disturbance regimes exert dominant control over plant species distribution and ecosystem function (Harrison and Prentice 2003; Bond and others 2005). In the western US, both climate and disturbance are changing at unprecedented rates (MacDonald 2010; Seager and Vecchi 2010). There is strong evidence that reduced water availability associated with rising temperatures, increasing potential evapotranspiration, altered precipitation patterns, and declining snowpack are contributing to tree mortality and with increased drought are anticipated to shape plant communities over much of the western US in the coming century (Adams and others 2009; Allen and others 2010; Williams and others 2010; Anderegg and others 2012; Williams and others 2012). Further, wildfires and insect outbreaks are increasing in frequency and severity (Westerling and others 2006). For example, a recent mountain pine beetle outbreak is decimating lodgepole pine (Pinus contorta) stands throughout the western US and Canada (Raffa and others 2008). Although the independent effects of climate change and disturbance on ecosystem structure and function, including stand water balance, have been recognized and explored (for example, Cramer and others 2001; Gordon and Famiglietti 2004; Porporato and others 2004), relatively little is known about how changing climate and altered disturbance regimes will interact to affect ecohydrological and ecological processes that shape ecosystems (Turnbull and others 2011; Sterling and others 2012).
Plant species and vegetation-type distributions are expected to shift in response to altered climate and disturbance (Lawler and others 2009), and examination of boundaries between vegetation types can provide insight into the rates and ecological consequences of these shifts (Neilson 1993). Vegetation-type boundaries have a long history of ecological study (Yarrow and Marín 2007) but there is a growing need for these studies in the context of global environmental change (for example, Beckage and others 2008). Key aspects of boundaries are not well understood, including controls over boundary location and potential broad shifts in boundaries (Cadenasso and others 2003). In semiarid, water-limited ecosystems the impact of boundaries on ecosystem structure and function and resulting consequences for ecohydrological cycling remain uncertain. Controls over boundary locations can be conceptualized hierarchically with broad environmental conditions, often climate, dictating the rough location of a transition zone within which processes operating at fine spatial scales, such as topography, soils, disturbance history, or vegetation structure, will determine the specific location and spatial pattern of the boundary between ecosystems (Gosz 1993; Strayer and others 2003; Yarrow and Salthe 2008; Danz and others 2011).
Species distribution models suggest a potential northward and upslope movement of sagebrush (Artemisia tridentata) in response to climate change (Schlaepfer and others 2012c). In addition, lodgepole pine forests are experiencing widespread and severe tree mortality from mountain pine beetles (Raffa and others 2008). The potential for sagebrush to expand and advance, while lodgepole pine retreats following beetle mortality, underscores a potentially rapid and widespread ecological change for the western US, and highlights knowledge gaps regarding controls over lower tree line dynamics (Ettinger and others 2011). Such a transition would likely be influenced by existing vegetation, through both structural impacts and competition, implying a facilitative role for disturbance by enabling distributional shifts in ecosystems. Paleoecological evidence suggests that boundaries between sagebrush and lodgepole pine ecosystems have been responsive to moisture, but that disturbance frequency plays a dominant role in precipitating shifts (Minckley and others 2011). These recent findings emphasize the need to address potential interactions between ecohydrology and disturbance when considering dynamics between adjacent ecosystems.
The importance of ecohydrology in arid and semiarid ecosystems, and specifically the crucial influence of soil water on plant establishment, survival, and eventual ecosystem productivity and species composition, suggests that examining current and future patterns of water cycling is a useful approach to understanding the causes and consequences of ecosystem shifts. In this study, we examine ecohydrological patterns between adjacent sagebrush steppe and lodgepole pine forests (Figure 1), two of the most widespread vegetation types in the western United States (Figure 1, McArthur and Plummer 1978; West and Young 2000), each playing crucial roles in the hydrologic cycle of this water-limited region (Troendle and King 1985; Burton 1997; Seyfried and others 2005; Wilcox and others 2012). Our overall goal is to understand at fine spatial scales where the climatic and topographic controls on ecohydrology are constant and how disturbances and changing climatic conditions may impact hydrologic cycling in adjacent sagebrush and lodgepole pine stands. We characterized the vegetation and soils in these adjacent ecosystem types at the spatial scale of a stand and used the results to address two ecohydrological objectives. First, we describe patterns of soil water availability and ecosystem water balance in both ecosystems, quantifying the potential role that vegetation/soil structure plays in maintaining or even stabilizing the boundary through an influence on hydrologic cycling. We anticipate that differences between the two species in the capacity to acquire soil water (for example, sagebrush can extract soil water at lower water potential than lodgepole pine), combined with differences in canopy structure and litter abundance, will result in summer dry periods that are longer and more severe in the sagebrush ecosystems. Second, through modeling we characterize the potential impact of disturbance (insect outbreaks and/or surface or canopy-replacing wildfires) and climate change on water availability and balance, providing insight into the potential for these processes to alter the ecohydrologic differences that may sustain the boundary and thus contribute to potential distributional shifts. We anticipated that disturbances impacting the forest litter layer (for example, surface fire), will have the greatest positive impact on water yield, whereas disturbances impacting only the canopy (for example, insects) will result in the highest soil water availability. Disturbances impacting both litter and canopy (for example, canopy replacing fires) will create the driest summer soil conditions, and thus the most challenging conditions for successful lodgepole regeneration. These results will provide insight into the potential ecohydrological consequences of disturbances that are currently unfolding across much of the western US.

Adjacent sagebrush and lodgepole pine ecosystems whose boundary is characterized by abrupt changes in ecosystem composition and structure. Widespread mortality, evident in this image, is common along in the lodgepole-pine forest.
Methods
Site Description and Field Measurements
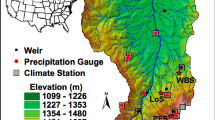
We simulated water cycling and balance at three sites in the Roosevelt National Forest in north-central Colorado (41.027N; −106.123W) (Figure 2). Each site represents a boundary between adjacent sagebrush steppe and lodgepole pine ecosystems (for example, Figure 1). The transition zone between dominance of trees versus shrubs is less than 5 m and we investigated both ecosystems for 50 m beyond the transition zone. Sites have similar mean annual temperature (2.2–2.7 °C), mean annual precipitation (440–470 mm), and elevation (2580–2640 m). At each site, trees and shrubs were sampled using the point-centered quarter method applied on two 100 m long transects oriented parallel to the boundary, located 5 and 25 m into each ecosystem-type (Mitchell 2001). Results were summarized into estimates of tree and shrub stem density, height, and standing aboveground biomass, using allometric equations for big sagebrush (Cleary and others 2008) and lodgepole pine (Pearson and others 1984), mean diameter at breast height (1.37 m) and stand basal area for the lodgepole pine ecosystem, and mean crown volume and canopy cover for the sagebrush ecosystem. Depending on tree density, 50–80 trees per field site were cored and annual rings counted, although not cross-dated, to estimate age. Herbaceous forest floor material was collected in 10, 0.25 m2 plots randomly located 25–50 m from the boundary, oven-dried to constant mass, and weighed to estimate biomass. Mineral soil samples were collected from each site, in each ecosystem in three, 5 cm diameter cores to a depth of 60 cm, partitioned into three layers (5–15, 20–30, 50–60 cm), and analyzed for texture via the Bouyoucos hydrometer method (Bouyoucos 1962).

Map of field sites (red stars) representing adjacent sagebrush (green) and lodgepone pine (black) ecosystems.
Ecohydrological Modeling Framework
Model Structure
We characterized water availability and balance using SOILWAT, a daily time step, multiple soil layer, process-based, simulation model of ecosystem water balance. SOILWAT was developed and tested in the semiarid western US shortgrass steppe (Parton 1978; Sala and others 1992; Bradford and Lauenroth 2006; Lauenroth and Bradford 2006). SOILWAT has recently been adapted for use in shrub-dominated ecosystems by: (a) incorporating an improved snow module from SWAT2K (Neitsch and others 2005), which was calibrated using SNOTEL data for the western US (Schlaepfer and others 2012b; (b) including the process of hydraulic redistribution (Ryel and others 2002); (c) modifying calculation of precipitation interception (Appendix 1); and (d) testing it against field measured data (Schlaepfer and others 2012b). Building on these previous efforts, this study is the first application of SOILWAT, encompassing all previous enhancements to SOILWAT, in a forested ecosystem. Simulating adjacent forest and sagebrush ecosystems provides a useful perspective for comparison and assessment of the forest results. SOILWAT has been used in combination with species distribution models to assess future distribution of sagebrush ecosystems (Schlaepfer and others 2012c). SOILWAT uses daily weather forcing, mean monthly relative humidity, wind speed, and cloud-cover data, monthly vegetation (live and dead biomass, litter, and active root profile) and site-specific properties of each soil layer to simulate the daily ecosystem water balance processes. Canopy conditions (biomass and seasonality) dictate potential transpiration rates and influence evaporation rates from the understory. Fluxes represented in SOILWAT include interception by vegetation and litter, evaporation of intercepted water, snow melt and loss (sublimation and wind redistribution), infiltration into the soil profile, percolation through the soil profile, bare-soil evaporation, transpiration from each soil layer, and drainage (Lauenroth and Bradford 2006; Schlaepfer and others 2012c). Hydraulic redistribution, known to occur in both sagebrush and pine species (Ryel and others 2002; Warren and others 2007), as characterized by Ryel and others (2002), is simulated in SOILWAT (further details in Schlaepfer and others 2012c). Hydraulic redistribution is only simulated for soil layers deeper than 5 cm (Wang 2011) and has the overall effect of minimizing differences in soil water content among soil layers. Outputs from SOILWAT are daily, monthly, and annual values of each water balance component (Parton 1978).
To represent forests, SOILWAT was modified to include two modest changes in model structure and appropriate parameter selection (Appendix 1). First, canopy interception in forests is calculated from leaf area index, rather than vegetation cover as utilized in the grass and shrub parameterization. Second, potential bare-soil evaporation is downregulated by the sum of litter and live biomass in the forest version, rather than by the sum of litter and total aboveground biomass in the shrub and grass version. Selection of the biomass level above which bare soil evaporation is nonexistent (illustrated in Figure A1.2) exerts important influence over soil evaporation rate and the partitioning of evaporation and transpiration. Numerous forest-specific parameters, based on field measurements, are also new in these simulations, including litter, aboveground plant biomass, and the proportion of aboveground biomass that is live, canopy height, LAI to % cover conversion coefficient, albedo and partitioning of evapotranspiration into bare-soil evaporation and transpiration (Appendix 1). The minimum soil water potential at which roots can extract water was set at −3.9 MPa for sagebrush (Kolb and Sperry 1999a; Kolb and Sperry 1999b) and −2.0 MPa for lodgepole pine (Fetcher 1976). Although this parameter influences how dry soils will become during the dry season, especially for deeper layers, it has relatively little impact on total annual water balance because the soil water available for transpiration is minimal at soil water potential lower than −2.0 MPa. Other modifications impacting both the lodgepole pine and sagebrush vegetation formulations include: (1) linear interpolation of monthly inputs to estimate daily values for calculations, (2) estimation of evaporative demand, based on PET, is now decreased for each canopy stratum by the amount evaporated from all strata above, (3) scaling transpiration estimates to the proportion of canopy above the snowpack when snow is present. SOILWAT estimates have been validated against field measurements for both sagebrush (Appendix 3) and forest (Appendix 4) ecosystems.
Scenarios and Analyses
We Addressed Two Main Objectives with These Modeling Efforts
Objective 1
To characterize differences in ecosystem water balance and soil water availability between the sagebrush and lodgepole ecosystems. To do this, we ran SOILWAT at each field site using 1/8-degree gridded weather data from the years 1981 to 2010 (Maurer and others 2002), which we scaled down to the site-level using 4-km gridded mean monthly PRISM climate data (Daly and others 2002; http://www.prism.oregonstate.edu/), and the National Climatic Data Center (2005) and soil conditions based on field measurements (Appendix 2). At each site, identical weather inputs were used for both the sagebrush and lodgepole pine simulations, ensuring that differences in model output are a consequence of either differences in soil conditions or vegetation structure and function represented in the model. Daily SOILWAT output over the 30-year period was averaged by day of year to produce seasonal trajectories for: (a) canopy and litter interception and subsequent evaporation, (b) infiltration into the soil profile, (c) evaporation from the soil, (d) soil water potential (in each of three layers), (e) transpiration, and (f) drainage out of the soil profile. In addition, total annual sums were calculated and averaged over the 30-year period for interception and subsequent evaporation, infiltration, soil evaporation, transpiration, and drainage. To characterize the relationship between daily transpiration and daily vapor pressure deficit (VPD) we calculated VPD as the difference between mean daily saturated vapor pressure (SVP) and nighttime SVP, as described in Yoder and others (2005).
Objective 2
To assess the impact of disturbance and climate change on water balance and availability. To do this, we ran SOILWAT for each site with individual and combined alterations to vegetation/soil structure and climatic conditions. For the forest ecosystem, four types of disturbance were simulated: (1) a moderate insect outbreak, implemented as a 50% reduction in canopy biomass; (2) a severe insect outbreak, implemented as complete tree mortality; (3) a surface fire, implemented as complete loss of the forest floor litter; and (4) a canopy-replacing fire, implemented as both tree mortality and forest floor loss. For the sagebrush ecosystem, a single fire disturbance was examined, implemented as complete shrub and grass mortality and litter loss. Our simulations did not account for effects of secondary succession, but rather describe immediate responses to disturbances. Climate change was incorporated by simulating anticipated climatic conditions for the years 2070–2100 (90 years later than the “current” simulations used for objective 1), for SRES emission scenario families B1 and A2 (Nakicenovic and Swart 2000) obtained from downscaled ensemble predictions (Maurer and others 2007). Although both scenarios predict consistently higher temperatures for these sites and slightly higher precipitation, the magnitude and seasonal patterns of changes differ between scenarios (Figure 3). Daily outputs were summarized into mean seasonal patterns and annual sums as described for objective 1.

Climate averaged among the three field sites from 1980 to 2010 (“Recent”; solid line) and for 2070–2100 under the A2 SRES scenario (“A2”; dotted line) and the B2 SRES scenario (“B1”; dashed line). Mean annual precipitation (mm) and temperature (°C) for each scenario are shown in upper right and left, respectively.
Results
Vegetation and Soil Conditions
Vegetation in the sagebrush ecosystem was dominated by big sagebrush (A. tridentata ssp. vaseyana) with 10,782 individuals ha−1 (SD among sites = 6241). Average crown volume and height were 0.05 m3 (SD = 0.04) and 42 cm (SD = 12), respectively. Shrub canopy cover averaged 14% (SD = 6.7) and aboveground biomass in the sagebrush ecosystem was 2.4 Mg ha−1 (SD = 1.3). Vegetation in the lodgepole ecosystem consisted almost entirely of lodgepole pine (Pinus contorta), with 1130 stems ha−1 (SD among sites = 641). Average tree DBH and height were 16.9 cm (SD among sites = 0.35) and 9.5 m (SD = 1.6). Stand basal area was 32 m2 ha−1 (SD = 17) and aboveground biomass was 113.4 Mg ha−1 (SD = 60.7). Mean tree age was 105 years (SD of 39 years) across the three field sites. Many of the trees at these sites are dead or dying from a mountain pine beetle (Dendroctonus ponderosae) outbreak that is common within lodgepole pine forests across the western US (Raffa and others 2008). The percentage of trees displaying visible signs of mortality in summer 2011 averaged 47% and ranged between 45 and 49%; additional mortality likely occurred following these surveys.
Litter depth in the sagebrush ecosystem was 0.5 cm (SD = 0.3), compared to 2.9 cm (SD = 0.3) in the lodgepole pine ecosystem. Similarly, sagebrush litter mass averaged 4.8 Mg ha−1 (SD = 1.8), compared to 19.3 Mg ha−1 (SD = 4.7) in lodgepole pine. Mineral soils at all sites are sandy loams. Soil texture differences are modest, with no consistent differences or anticipated impacts on water availability or ecosystem water balance (Appendix 2).
Water Availability and Balance in Adjacent Ecosystems (Objective 1)
Seasonal Water Cycling and Availability
Because climatic conditions were similar between vegetation types, many seasonal patterns of water cycling and availability were similar between the sagebrush and lodgepole pine ecosystems. The combination of melting snowpack and high-spring precipitation generated clear spring peaks in water infiltration into the soil and drainage in both ecosystems (Figure 4). In addition, both ecosystems displayed the highest soil water potential at all depths during the spring, in contrast to summer, when water potential is lowest. Also transpiration and evaporation from canopy/litter in both ecosystems were highest during summer, with interception losses continuing into the fall.

Average seasonal patterns of soil water potential and water balance components for sagebrush (left panels) and lodgepole pine (right panels). Top panels show soil water potential at 3 depths (black, red and blue lines corresponding to left Y axis) and infiltration into the soil profile (light blue line corresponding to right Y axis). Bottom panels show evaporation from interception (“surface evaporation”), soil evaporation and transpiration (grey, brown and green lines, respectively, corresponding to left Y axis) and drainage (purple line corresponding to right Y axis). Shaded area around lines is the standard deviation of 30-year mean seasonal patterns among the three field sites.
Consistent with our expectations about the ecohydrological role of vegetation and ecosystem structure, our modeling results suggest three consistent differences between sagebrush and lodgepole pine ecosystems. First, at these sites where maximum temperature can be above 0°C for any day of the year, lodgepole pine foliage above the snowpack results in transpiration during the winter (Figure 4), albeit at extremely low rates. Because winter transpiration occurs under a relatively stable snowpack that is not adding substantial water to the soil profile, soil water is slowly depleted, and soil water potential gradually declines, whereas soil water potential in the sagebrush ecosystem remains high throughout the winter. Second, the substantial layer of litter on the forest floor in the lodgepole pine ecosystem inhibits evaporation from the soil; in combination with the tree foliage, the litter layer greatly reduced soil evaporation. By contrast, the much thinner litter layer and lower foliage mass in the sagebrush ecosystem allowed consistent soil evaporation during summer and fall. Lastly, model results suggest that the period of dry soils during summer and early fall was both longer and more severe in the sagebrush ecosystem. Mean seasonal soil water potential patterns suggest similar timing in fall soil water recharge between sagebrush and lodgepole pine. However, the modeled sagebrush soil dried out much earlier in the summer, sagebrush soil water potential dropped below −1 MPa at roughly day 170, compared to day 200 in lodgepole pine. The notably lower minimum summer soil water potential in sagebrush compared to lodgepole pine emerges as a consequence of sagebrush’s ability to extract and transpire soil water at lower water potential than lodgepole (Figure 5).

Relationship between daily transpiration and average daily vapor pressure deficit for sagebrush and lodgepole ecosystems during the summer (days 150–210) across a range of soil water potential for all three sites under ambient climatic conditions in the absence of disturbance. Colors indicate the soil water potential of the wettest layer; small points are individual days, large points are smoothed (loess) relationships for each level of soil water potential (gray points are days when soil water potential was less than −1.9 MPa.
Annual Water Balance
When summed over the year, these modeled seasonal patterns suggest interesting differences in water balance between undisturbed sagebrush and lodgepole pine under ambient conditions (Table 1). Although annual canopy interception and sublimation were similar (60–66 and 117–121 mm y−1; see Appendix 1 for description of interception algorithms), litter interception in lodgepole pine was four times higher than sagebrush (61 vs. 15 mm y−1). Consequently, more water enters the soil profile in sagebrush ecosystems (255 vs. 199 mm y−1 in lodgepole pine) but the fate of water after entering the soil profile was quite different between the ecosystems. Notably, soil evaporation (not including litter) averaged 75 mm y−1 in sagebrush soils, compared to essentially 0 mm y−1 in the lodgepole pine soils. Although modeled transpiration was higher in lodgepole pine (189 vs. 148 mm y−1 in sagebrush), modeled drainage was lower in lodgepole pine (10 vs. 32 mm y−1 in sagebrush).
Disturbance and Climate Impacts (Objective 2)
Disturbance Impacts
Our simulations suggest that disturbance can dramatically influence water cycling, and that the impacts are dependent on disturbance type. Moderate beetle mortality reduced canopy interception from 66 to 52 mm y−1 and slightly increased litter interception (7 mm y−1 higher) and infiltration (17 mm y−1 higher; Table 1). Although transpiration in the moderate beetle scenario decreased transpiration by 17 mm y−1, drainage increased by about 29 mm y−1 (Appendix 5), and soils were substantially wetter than the undisturbed forest scenario, although not as wet as the severe beetle mortality scenario (Figure 5). By comparison, severe beetle mortality in the lodgepole pine forest eliminated canopy interception, increased litter interception slightly, and increased infiltration (Table 1). However, transpiration was eliminated and bare soil evaporation was still somewhat inhibited (Appendix 5) by the intact litter layer, resulting in consistently wet soils at all depths (Figure 6) and a substantial increase in drainage (from 10 to 214 mm y−1).

Impact of climate change scenarios and disturbance treatments on mean seasonal patterns of soil water potential at 3 depths (black, red and blue lines corresponding to left Y axis) and infiltration into the soil profile (light blue line corresponding to right Y axis). Columns are climate scenarios and rows are disturbance treatments.
Surface fire scenarios eliminated the litter layer and litter interception, which increased infiltration (Table 1). Although modeled soil evaporation also increased following litter removal (from 0 to 46 mm y−1), the increase was smaller than the increase in infiltration (~60 mm y−1). As a consequence of initially higher soil water, surface fire increased transpiration slightly. Following surface fire, elevated soil evaporation and sustained transpiration rapidly depleted soil water, resulting in a longer and more severe summer dry period, despite greater infiltration (Figure 6).
The modeled ecohydrological consequences of severe fire (removing both litter and live vegetation) were consistent between sagebrush and lodgepole ecosystems because the only remaining differences between the ecosystems were subtle and inconsistent such as slight soil texture differences. Interception by dead standing boles would likely cause differences, but was not represented in these simulations. In both ecosystems, the severe fire scenario eliminated interception and increased infiltration as all precipitation entered the soil profile (Table 1). Severe fire also eliminated transpiration while dramatically increasing bare soil evaporation to roughly 220 mm y−1 and drainage to over 100 mm y−1. The elevated soil evaporation, along with the lack of hydraulic redistribution of water by roots, resulted in extremely dry surface soils for a prolonged period during the summer and consistently wet soils in the deep layer that was not impacted by evaporation (Figure 6).
Climate Change Impacts
The altered temperature and precipitation patterns of the A2 and B1 scenarios (Figure 3) impacted both the seasonal timing of water fluxes as well as annual water balance. Warmer temperatures resulted in less precipitation occurring as snow as well as earlier spring snowmelt, which created a smaller spring peak in infiltration and drainage in both ecosystems (Figure 6; Appendix 5). In addition, the increased proportion of precipitation occurring as rain, in combination with modest increases in overall precipitation, elevated overall interception (sum of canopy and litter) by approximately 15 mm y−1 in sagebrush and 30 mm y−1 in lodgepole pine (Table 1). Despite higher interception, modeled water infiltration into the soil increased in both climate change scenarios because sublimation decreased substantially. Soil evaporation increased by about 30 mm y−1 in the sagebrush ecosystem under both scenarios, but remained negligible in the lodgepole pine ecosystem.
The consequences of climate change for modeled drainage and transpiration varied between climate scenarios and ecosystems. Under the hotter and slightly drier A2 scenario, transpiration decreased around 12 mm y−1 in sagebrush whereas drainage increased by about 6 mm y−1. In lodgepole pine under the A2 scenario, transpiration increased by roughly 10 mm y−1 while drainage remained unchanged. Under the B1 scenario (not as warm as A2 but wetter than both A2 and ambient), sagebrush transpiration increased slightly and drainage increased by around 15 mm y−1, whereas in lodgepole pine, transpiration increased by approximately 30 mm y−1 and drainage increased only slightly. Seasonal soil water dynamics responded to the earlier snowmelt, longer warm season and higher potential evapotranspiration rates that emerge from the hotter temperatures in the climate change scenarios. As a result, the summer periods of dry soils in sagebrush and lodgepole ecosystems were both longer and drier under the two climate change scenarios (Figure 6). In addition, warmer temperatures under both scenarios caused periodic winter snowmelt, providing water recharge to the soil profile that largely offset the winter transpiration losses in lodgepole pine and maintained higher soil water potential throughout the winter in lodgepole pine soils.
The combined impact of climate change and disturbance on modeled water balance in these ecosystems were generally additive. The notable exception occurred as a consequence of disturbance completely eliminating a water balance component. For example, canopy fire eliminated transpiration regardless of the climate scenario. Because climate change and disturbance often impact water balance in the same direction, many water balance components, including the length and severity of dry summer soils, infiltration, soil evaporation, transpiration, and drainage, displayed the largest change in both seasonal and annual water balance (Figure 6; Table 1; Appendix 5), when climate change and disturbance were combined.
Discussion
Complete ecohydrological datasets with long-term measurements of all ecosystem water balance components are scarce for any ecosystem, especially in western coniferous forests and shrublands (Jackson and others 2009a). As a result, our understanding of the relationships among vegetation, soils, climate, and water balance remains limited, hampering efforts to assess the impact of changing climate on water cycling, availability, utilization, or overall yield (Wilcox 2010). Our modeling approach directly addresses these challenging questions and is strengthened by comparisons with previous research.
We have previously shown that water cycling and balance estimates from SOILWAT for sagebrush ecosystems are consistent with results from empirical and other modeling studies (Appendix 3; see also appendices from Schlaepfer and others 2012c). However, the results presented here are the first application of SOILWAT to a forested ecosystem. Comparison of modeled water balance and availability for lodgepole pine forests compare favorably to field measurements of soil water and transpiration from a lodgepole pine location near our sites (Appendix 4). Our 30-year average transpiration results suggest a strong summer peak of roughly 1.5 mm day−1 followed by a relatively rapid decline. Spittlehouse (2002) estimated that daily transpiration in lodgepole pine forests ranged between 0.5 and 1.5 mm day−1 before mid August and between roughly 0.2 and 1.0 mm day−1 between mid August and October. In a mixed stand of lodgepole pine and subalpine fir, Pataki and others (2000) estimated maximum daily transpiration at 3–3.5 mm day−1. Knight and others (1981) estimated daily maximum transpiration for lodgepole pine forests at a slightly higher elevation to be 3.3 mm. Our estimates of maximum daily transpiration values were lower than either Pataki and others (2000) or Knight and others (1981), perhaps because of our plot locations near lower tree line, where water availability is presumably more limiting. Critically, SOILWAT characterization of the relationship between transpiration and daily VPD (Figure 5) are very similar to results presented in Pataki and others (2000) for lodgepole and Naithani and others (2012) for sagebrush, although comparisons are complicated by the daily (including nighttime) representation of VPD in SOILWAT.
Our estimates for overall water balance are also consistent with previous work by Knight and others (1985), who modeled water balance across a range of lodgepole pine forests that are higher and wetter than our sites. They estimated higher total annual interception (240 mm vs. our estimate of 130 mm), transpiration (250 mm vs. our estimate of 190 mm) and drainage (130 mm vs. our estimate of 10 mm.) Based on data from the same sites, Knight and others suggested that drainage is tightly linked to snowmelt—as it is in our simulations. Kaye and others (1999) estimated 28 to 67 mm y−1 of drainage from ponderosa pine forests in northern Arizona that receive 577 mm y−1 of precipitation, compared to our estimate of 10 mm from sites with 448 mm of annual precipitation. Although our lower drainage estimates may be a result of the drier conditions at our lower tree line sites compared to Knight and others (1985) or Kaye and others (1999), SOILWAT does not currently include preferential flow paths, which may result in decreased infiltration to deep soil layers and potential decreased drainage (van der Heijden and others 2013). Kaufman (1985) estimated that annual transpiration in high-elevation lodgepole pine forests ranged from around 150 mm y−1 at 22 m2 ha−1 basal area to roughly 250 mm y−1 at 35 m2 ha−1 basal area. By comparison, our annual transpiration estimates under ambient conditions for undisturbed lodgepole pine were 189 mm y−1 with 32 m2 ha−1 of basal area.
Our results imply that the differences in species composition and vegetation structure observed between these adjacent ecosystems impact ecohydrological function and create predictable differences in water cycling between sagebrush and lodgepole pine ecosystems despite the lack of difference in either climate or topographic influences across ecosystems. Although patterns of canopy interception show relatively small differences, the substantial forest litter layer in lodgepole pine minimizes soil evaporation and moderates soil moisture levels during the summer. As a result, lodgepole pine may lose slightly less water to drainage than sagebrush, and somewhat more to annual transpiration. Because drainage in both sagebrush and lodgepole pine occurs almost exclusively in spring as soil water is recharged by snowmelt, the lower drainage from the lodgepole pine ecosystem may be due in part to low, but non-zero (average ~0.1 mm day−1), winter transpiration that decreases spring soil moisture content and lowers the probability of soil saturation leading to drainage. Transpiration while snowpack exists has been observed in previous studies (Knight and others 1985; Day and others 1989; Monson and others 2005) and may be even more pronounced at our sites as warmer winters may promote cold-season transpiration. In our simulations, transpiration as a percentage of actual evapotranspiration averaged 46% in lodgepole pine (similar to the 42% observed by Viville and others 1993), compared to only 36% in sagebrush.
Because soil texture is similar among our sites (Appendix 2), these ecohydrological differences were driven primarily by differences in litter and vegetation. The minimal effect of soil differences is apparent in comparisons of severe fire scenarios in sagebrush versus lodgepole pine (Table 1; Figure 6; Appendix 5). However, the role of structural differences in reinforcing and/or maintaining the abrupt boundary between lodgepole and sagebrush that typifies the fine spatial scale characteristics of this boundary remains unclear. On one hand, the extremely dry sagebrush soils in summer—influenced by both vegetation and litter structure as well as the minimum water potential at which sagebrush can extract water from the soil—(see “Methods” section) may inhibit successful lodgepole pine germination and establishment into the sagebrush. Further, these modeled soil conditions may drive a positive feedback to the boundary and contribute to its abrupt nature. However, lodgepole pine regeneration is most successful following stand-replacing disturbances that remove the litter layer and expose mineral soil (Lotan and Critchfield 1990), and our simulations indicate that this combination of disturbances results in forest soil water availability patterns similar to the sagebrush ecosystem. Although successful lodgepole pine regeneration may maintain the boundary because of differential snow capture or microclimatic consequences of post-disturbance standing and downed woody material in the forest, neither process is incorporated in our simulations. An additional source of variation not explicitly represented in these simulations is partial tree mortality, and resulting outcomes for stand-level tree age structure, that often occurs in forest pest outbreaks (Samman and Logan 2000; Logan and others 2003). Simulations of partial mortality (not shown) displayed water balance patterns intermediate between the simulations of complete mortality and intact stands, suggesting that initial ecohydrological response to beetle-induced mortality will be similarly bounded by the scenarios presented here and may diverge into conditions not represented in these simulations as stand age structure progresses following the disturbance.
Snow interception and subsequent sublimation in lodgepole pine canopies can reduce snow water equivalent (SWE) by 20–25% compared to clearcut areas, although standing dead trees, representative of a recent insect outbreak, have more moderate effects on SWE (Boon 2012), and canopy interception in sagebrush ecosystems is likely lower. In addition, the lower snow interception rates in forests experiencing insect mortality may be largely offset by higher snow sublimation rates from the snowpack, resulting in relatively little overall change in spring water additions from snow as a consequence of mortality (Biederman and others 2012). Thus, although our results may slightly overestimate water delivered to the soil in lodgepole pine forests, with a resulting increase in our estimates of lodgepole pine transpiration and/or drainage, the magnitude of these overestimates are likely small. Regardless of these uncertainties, our results highlight the central role of winter precipitation and snowpack for recharging soil water in sagebrush and lodgepole pine ecosystems, which is consistent with previous dendrochronological and physiological studies showing a strong relationships between tree growth and winter precipitation in western forests (Fritts 1966; Stahle and others 2009; Kerhoulas and others 2013)
As anticipated, the ecohydrological differences between sagebrush and lodgepole pine suggest that shifts in the distribution of these ecosystems at broad spatial scales would alter regional water balance, independent of changes in climate itself. In particular, the greater drainage in the sagebrush ecosystem, which is consistent with observations of ecohydrological impacts from woody plant encroachment (Huxman and others 2005), underscores that widespread transition to sagebrush could increase groundwater recharge and/or streamflow. Our results suggest that disturbances impacting vegetation or litter may have substantial ecohydrological consequences. Consistent with some previous work, these simulations suggest that forest disturbance increases soil-water content (Zou and others 2008; Dore and others 2012), decreases evapotranspiration and increases water yield (Adams and others 2011; Edburg and others 2012; Mikkelson and others 2013).
The climatic changes captured in the A2 and B1 emission scenarios were sufficient to alter modeled water balance for both ecosystems. The potential impact of climate change-driven alterations to water cycling and plant water availability is a growing area of concern (Newman and others 2006), especially as evidence emerges that changes in climate are already affecting the ecohydrology of arid and semiarid ecosystems (Barnett and others 2008; Cayan and others 2010). Predictions of earlier and less peaked seasonal timing of snowmelt and infiltration, increased overall interception and soil evaporation, higher transpiration in forests, and drier summer soils are consistent across emission scenarios, and are consistent with other studies (Wetherald and Manabe 2002; Gerten and others 2007; Molotch and others 2009; Schlaepfer and others 2012a). Climate change consequences for infiltration and drainage, as well as transpiration in sagebrush, differ between A2 and B1 scenarios, and point to high uncertainty in estimates of future water yield from semiarid ecosystems. In fact, previous work indicates both increases, (Wetherald and Manabe 2002) and decreases (Cayan and others 2010).
Our simulations provide preliminary insights into the potential combined consequences of both disturbance and climate change, a topic of regional importance (Wilcox 2010) but associated with very limited observations. The combination of climate change and disturbance had the largest impact on water balance, suggesting that alterations to ecosystem structure resulting from these multi-factor scenarios may cross thresholds in water availability that more profoundly impact function than anticipated from single-factor scenarios. For example, the longer and more severe dry summer soils in lodgepole pine under climate change, especially under the A2 scenario, created summer soil water conditions more similar to sagebrush ecosystems, highlighting a potentially important driver of future transition from lodgepole pine to sagebrush forecast by species distribution models (Schlaepfer and others 2012c). In contrast, established individuals of sagebrush or lodgepole pine may be able to enhance long-term productivity if they can withstand longer, and more severe, summer drought conditions to capitalize on the longer growing seasons and potentially more water available for transpiration, associated with future climate. In addition, nutrient losses following disturbance will impact nutrient availability and may exert important control over lodgepole pine recovery from disturbances (Knight and others 1985).
The climate change treatments we examined were structured to represent changes in mean monthly climate, not increases in the magnitude of weather variation or increases in the frequency of extreme weather events that may accompany climate change (CCSP 2008). Although climate change will presumably alter the location of conditions that define the broad transition zone between lodgepole pine and sagebrush, ecosystem structure legacies may inhibit boundary shifts in the absence of disturbance, at least during the lifetime of established trees. For example, the combination of warmer temperatures and removal of the litter and canopy resulted in extremely dry summer soils (Figure 6). As disturbances and extreme drought become more frequent (Westerling and others 2006; Bigler and others 2007; Cayan and others 2010; Williams and others 2012; Anderegg and others 2013), the probability of tree or shrub regeneration will increasingly determine the distribution of these ecosystems (Jackson and others 2009b).
References
Adams HD, Guardiola-Claramonte M, Barron-Gafford GA, Villegas JC, Breshears DD, Zou CB, Troch PA, Huxman TE. 2009. Temperature sensitivity of drought-induced tree mortality portends increased regional die-off under global-change-type drought. Proc Natl Acad Sci USA 106:7063–6.
Adams HD, Luce CH, Breshears DD, Allen CD, Weiler M, Hale VC, Smith AMS, Huxman TE. 2011. Ecohydrological consequences of drought- and infestation-triggered tree die-off: insights and hypotheses. Ecohydrology 5(2):145–59.
Allen CD, Macalady AK, Chenchouni H, Bachelet D, McDowell N, Vennetier M, Kitzberger T, Rigling A, Breshears DD, Hogg EH, Gonzalez P, Fensham R, Zhang Z, Castro J, Demidova N, Lim J-H, Allard G, Running SW, Semerci A, Cobb N. 2010. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For Ecol Manag 259:660–84.
Anderegg WRL, Berry JA, Smith DD, Sperry JS, Anderegg LDL, Field CB. 2012. The roles of hydraulic and carbon stress in a widespread climate-induced forest die-off. Proc Natl Acad Sci 109:233–7.
Anderegg WRL, Plavcová L, Anderegg LDL, Hacke UG, Berry JA, Field CB. 2013. Drought’s legacy: multiyear hydraulic deterioration underlies widespread aspen forest die-off and portends increased future risk. Glob Change Biol 19:1188–96.
Barnett TP, Pierce DW, Hidalgo HG, Bonfils C, Santer BD, Das T, Bala G, Wood AW, Nozawa T, Mirin AA, Cayan DR, Dettinger MD. 2008. Human-induced changes in the hydrology of the Western United States. Science 319:1080–3.
Beckage B, Osborne B, Gavin DG, Pucko C, Siccama T, Perkins T. 2008. A rapid upward shift of a forest ecotone during 40 years of warming in the Green Mountains of Vermont. Proc Natl Acad Sci 105:4197–202.
Biederman JA, Brooks PD, Harpold AA, Gochis DJ, Gutmann E, Reed DE, Pendall E, Ewers BE. 2012. Multiscale observations of snow accumulation and peak snowpack following widespread, insect-induced lodgepole pine mortality. Ecohydrology. doi:10.1002/eco.1342.
Bigler C, Gavin DG, Gunning C, Veblen TT. 2007. Drought induces lagged tree mortality in a subalpine forest in the Rocky Mountains. Oikos 116:1983–94.
Bond WJ, Woodward FI, Midgley GF. 2005. The global distribution of ecosystems in a world without fire. New Phytol 165:525–38.
Boon S. 2012. Snow accumulation following forest disturbance. Ecohydrology 5:279–85.
Bouyoucos GJ. 1962. Hydrometer method improved for making particle size analyses of soils. Agron J 54:464–5.
Bradford JB, Lauenroth WK. 2006. Controls over invasion of Bromus tectorum: the importance of climate, soil, disturbance and seed availability. J Veg Sci 17:693–704.
Burton TA. 1997. Effects of basin-scale timber harvest on water yield and peak streamflow. J Am Water Res Assoc 33:1187–96.
Cadenasso ML, Pickett STA, Weathers KC, Bell SS, Benning TL, Carreiro MM, Dawson TE. 2003. An interdisciplinary and synthetic approach to ecological boundaries. Bioscience 53:717–22.
Cayan DR, Das T, Pierce DW, Barnett TP, Tyree M, Gershunov A. 2010. Future dryness in the southwest US and the hydrology of the early 21st century drought. Proc Natl Acad Sci 107(50):21271–6.
CCSP. 2008. A report by the U.S. Climate Change Science Program and the Subcommittee on Global Change Research. In: Karl TR, Meehl GA, Miller CD, Hassol SJ, Waple AM, Murray WL, Eds. Weather and climate extremes in a changing climate. Regions of focus: North America, Hawaii, Caribbean, and U.S. Pacific Islands. Washington, DC: Department of Commerce, NOAA’s National Climatic Data Center.
Cleary MB, Pendall E, Ewers BE. 2008. Testing sagebrush allometric relationships across three fire chronosequences in Wyoming, USA. J Arid Environ 72:285–301.
Cramer W, Bondeau A, Woodward FI, Prentice IC, Betts RA, Brovkin V, Cox PM, Fisher V, Foley JA, Friend AD, Kucharik C, Lomas MR, Ramankutty N, Sitch S, Smith B, White A, Young-Molling C. 2001. Global response of terrestrial ecosystem structure and function to CO2 and climate change: results from six dynamic global vegetation models. Glob Change Biol 7:357–73.
Daly C, Gibson WP, Taylor GH, Johnson GL, Pasteris P. 2002. A knowledge-based approach to the statistical mapping of climate. Clim Res 22:99–113.
Danz NP, Reich PB, Frelich LE, Niemi GJ. 2011. Vegetation controls vary across space and spatial scale in a historic grassland-forest biome boundary. Ecography 34:402–14.
Day TA, DeLucia EH, Smith WK. 1989. Influence of cold soil and snowcover on photosynthesis and leaf conductance in two Rocky Mountain conifers. Oecologia 80:546–52.
Dore S, Montes-Helu M, Hart SC, Hungate BA, Koch GW, Moon JB, Finkral AJ, Kolb TE. 2012. Recovery of ponderosa pine ecosystem carbon and water fluxes from thinning and stand-replacing fire. Glob Change Biol 18:3171–85.
Edburg SL, Hicke JA, Brooks PD, Pendall EG, Ewers BE, Norton U, Gochis D, Gutmann ED, Meddens AJH. 2012. Cascading impacts of bark beetle-caused tree mortality on coupled biogeophysical and biogeochemical processes. Front Ecol Environ 10:416–24.
Ettinger AK, Ford KR, HilleRisLambers J. 2011. Climate determines upper, but not lower, altitudinal range limits of Pacific Northwest conifers. Ecology 92:1323–31.
Fetcher N. 1976. Patterns of leaf resistance to lodgepole pine transpiration in Wyoming. Ecology 57:339–45.
Fritts HC. 1966. Growth-rings of trees: their correlation with climate. Science 154:973–9.
Gerten D, Schaphoff S, Lucht W. 2007. Potential future changes in water limitations of the terrestrial biosphere. Clim Change 80:277–99.
Gordon WS, Famiglietti JS. 2004. Response of the water balance to climate change in the United States over the 20th and 21st centuries: results from the VEMAP Phase 2 model intercomparisons. Glob Biogeochem Cycles 18:1030.
Gosz JR. 1993. Ecotone Hierarchies. Ecol Appl 3:370–6.
Harrison SP, Prentice CI. 2003. Climate and CO2 controls on global vegetation distribution at the last glacial maximum: analysis based on palaeovegetation data, biome modelling and palaeoclimate simulations. Glob Change Biol 9:983–1004.
Huxman TE, Wilcox BP, Breshears DD, Scott RL, Snyder KA, Small EE, Hultine K, Pockman WT, Jackson RB. 2005. Ecohydrological implications of woody plant encroachment. Ecology 86:308–19.
Jackson RB, Jobbágy EG, Nosetto MD. 2009a. Ecohydrology in a human-dominated landscape. Ecohydrology 2:383–9.
Jackson ST, Betancourt JL, Booth RK, Gray ST. 2009b. Ecology and the ratchet of events: climate variability, niche dimensions, and species distributions. Proc Natl Acad Sci USA 106:19685–92.
Kaufmann MR. 1985. Annual transpiration in subalpine forests: large differences among four tree species. For Ecol Manag 13:235–46.
Kaye JP, Hart SC, Cobb RC, Stone JE. 1999. Water and nutrient outflow following the ecological restoration of a ponderosa pine–bunchgrass ecosystem. Restor Ecol 7:252–61.
Kerhoulas LP, Kolb TE, Koch GW. 2013. Tree size, stand density, and the source of water used across seasons by ponderosa pine in northern Arizona. For Ecol Manag 289:425–33.
Knight DH, Fahey TJ, Running SW. 1985. Water and nutrient outflow from contrasting lodgepole pine forests in Wyoming. Ecol Monogr 55:29–48.
Knight DH, Fahey TJ, Running SW, Harrison AT, Wallace LL. 1981. Transpiration from 100-yr-old lodgepole pine forests estimated with whole-tree potometers. Ecology 62:717–26.
Kolb KJ, Sperry JS. 1999a. Transport constraints on water use by the Great Basin shrub, Artemisia tridentata. Plant Cell Environ 22:925–35.
Kolb KJ, Sperry JS. 1999b. Differences in drought adaptation between subspecies of sagebrush (Artemisia tridentata). Ecology 80:2373–84.
Lauenroth WK, Bradford JB. 2006. Ecohydrology and the partitioning AET between transpiration and evaporation in a semiarid steppe. Ecosystems 9:756–67.
Lawler JJ, Shafer SL, White D, Kareiva P, Maurer EP, Blaustein AR, Bartlein PJ. 2009. Projected climate-induced faunal change in the Western Hemisphere. Ecology 90:588–97.
Logan JA, Régnière J, Powell JA. 2003. Assessing the impacts of global warming on forest pest dynamics. Frontiers Ecol Environ 1:130–7.
Lotan JE, Critchfield WB. 1990. Lodgepole Pine. In: Burns RM, Honkala BH, Eds. Silvics of North America, USDA Forest Service Agricultural Handbook, Vol. 1. Washington, DC: USDA. p 654.
MacDonald GM. 2010. Water, climate change, and sustainability in the southwest. Proc Natl Acad Sci 107:21256–62.
Maurer EP, Brekke L, Pruitt T, Duffy PB. 2007. Fine-resolution climate projections enhance regional climate change impact studies. EOS Trans AGU 88:504.
Maurer EP, Wood AW, Adam JC, Lettenmaier DP, Nijssen B. 2002. A long-term hydrologically based dataset of land surface fluxes and states for the conterminous United States. J Clim 15:3237–51.
McArthur ED, Plummer AP. 1978. Biogeography and management of native western shrubs: a case study, section tridentatae of Artemisia. Great Basin Nat Mem 2:229–43.
Mikkelson KM, Maxwell RM, Ferguson I, Stednick JD, McCray JE, Sharp JO. 2013. Mountain pine beetle infestation impacts: modeling water and energy budgets at the hill-slope scale. Ecohydrology 6:64–72.
Minckley TA, Shriver RK, Shuman B. 2011. Resilience and regime change in a southern Rocky Mountain ecosystem during the past 17,000 years. Ecol Monogr 82:49–68.
Mitchell K. 2001. Quantitative analysis by the point-centered quarter method. Geneva: Hobart and William Smith Colleges.
Molotch NP, Brooks PD, Burns SP, Litvak M, Monson RK, McConnell JR, Musselman K. 2009. Ecohydrological controls on snowmelt partitioning in mixed-conifer sub-alpine forests. Ecohydrology 2:129–42.
Monson RK, Sparks JP, Rosenstiel TN, Scott-Denton LE, Huxman TE, Harley PC, Turnipseed AA, Burns SP, Backlund B, Hu J. 2005. Climatic influences on net ecosystem CO2 exchange during the transition from wintertime carbon source to springtime carbon sink in a high-elevation, subalpine forest. Oecologia 146:130–47.
Naithani KJ, Ewers BE, Pendall E. 2012. Sap flux-scaled transpiration and stomatal conductance response to soil and atmospheric drought in a semi-arid sagebrush ecosystem. J Hydrol 464–465:176–85.
Nakicenovic N, Swart R, Eds. 2000. Emissions scenarios: a special report of working group III of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press.
National Climatic Data Center. 2005. Climate maps of the United States. http://cdo.ncdc.noaa.gov/cgi-bin/climaps/climaps.pl. Accessed May 2010.
Neilson RP. 1993. Transient ecotone response to climatic change: some conceptual and modelling approaches. Ecol Appl 3:385–95.
Neitsch S, Arnold J, Kiniry J, Williams J. 2005. Soil and water assessment tool (SWAT) theoretical documentation. version, 2005. Temple, TX: Blackland Research Center, Texas Agricultural Experiment Station.
Newman BD, Wilcox BP, Archer SR, Breshears DD, Dahm CN, Duffy CJ, McDowell NG, Phillips FM, Scanlon BR, Vivoni ER. 2006. Ecohydrology of water-limited environments: a scientific vision. Water Resour Res 42:06302.
Parton WJ. 1978. Abiotic section of ELM. In: Innis GS, Ed. Grassland simulation model. New York: Springer-Verlag. p 31–53.
Pataki DE, Oren R, Smith WK. 2000. Sap flux of co-occurring species in a western subalpine forest during seasonal soil drought. Ecology 81:2557–66.
Pearson JA, Fahey TJ, Knight DH. 1984. Biomass and leaf-area in contrasting lodgepole pine forests. Can J For Res 14:259–65.
Porporato A, Daly E, Rodriguez , ÄêIturbe I. 2004. Soil water balance and ecosystem response to climate change. Am Nat 164:625–32.
Raffa KF, Aukema BH, Bentz BJ, Carroll AL, Hicke JA, Turner MG, Romme WH. 2008. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: the dynamics of bark beetle eruptions. Bioscience 58:501–17.
Ryel R, Caldwell M, Yoder C, Or D, Leffler A. 2002. Hydraulic redistribution in a stand of Artemisia tridentata: evaluation of benefits to transpiration assessed with a simulation model. Oecologia 130:173–84.
Sala OE, Lauenroth WK, Parton WJ. 1992. Long-term soil-water dynamics in the shortgrass steppe. Ecology 73:1175–81.
Samman S, Logan J. 2000. Assessment and response to bark beetle outbreaks in the Rocky Mountain area: Report to Congress from Forest Health Protection, Washington Office, Forest Service, U.S. Department of Agriculture. General Technical Report RMRS-GTR-62. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, 46 p.
Schlaepfer DR, Lauenroth WK, Bradford JB. 2012a. Consequences of declining snow accumulation for water balance of mid-latitude dry regions. Glob Change Biol 18:1988–97.
Schlaepfer DR, Lauenroth WK, Bradford JB. 2012b. Ecohydrological niche of sagebrush ecosystems. Ecohydrology 5:453–66.
Schlaepfer DR, Lauenroth WK, Bradford JB. 2012c. Effects of ecohydrological variables on current and future ranges, local suitability patterns, and model accuracy in big sagebrush. Ecography 35:374–84.
Seager R, Vecchi GA. 2010. Greenhouse warming and the 21st century hydroclimate of southwestern North America. Proc Natl Acad Sci 107:21277–82.
Seyfried MS, Schwinning S, Walvoord MA, Pockman WT, Newman BD, Jackson RB, Phillips EM. 2005. Ecohydrological control of deep drainage in arid and semiarid regions. Ecology 86:277–87.
Spittlehouse DL. 2002. Sap flow and transpiration of old lodgepole pine trees. . Proceedings 25th Conference on Agricultural and Forest Meteorology, 20–24 May 2002, Norfolk, Virginia. American Meteorological Society, Boston, MA.
Stahle DW, Cleaveland MK, Grissino-Mayer HD, Griffin RD, Fye FK, Therrell MD, Burnette DJ, Meko DM, Villanueva Diaz J. 2009. Cool- and warm-season precipitation reconstructions over Western New Mexico. J Clim 22:3729–50.
Sterling SM, Ducharne A, Polcher J. 2012. The impact of global land-cover change on the terrestrial water cycle. Nature Clim Change 3(4):385–90.
Strayer DL, Power ME, Fagan WF, Pickett STA, Belnap J. 2003. A classification of ecological boundaries. Bioscience 53:723–9.
Troendle CA, King RM. 1985. The effect of timber harvest on the Fool Creek Watershed, 30 years later. Water Resour Res 21:1915–22.
Turnbull L, Wilcox BP, Belnap J, Ravi S, D’Odorico P, Childers D, Gwenzi W, Okin G, Wainwright J, Caylor KK, Sankey T. 2011. Understanding the role of ecohydrological feedbacks in ecosystem state change in drylands. Ecohydrology 5:174–83.
van der Heijden G, Legout A, Pollier B, Bréchet C, Ranger J, Dambrine E. 2013. Tracing and modeling preferential flow in a forest soil—potential impact on nutrient leaching. Geoderma 195–196:12–22.
Viville D, Biron P, Granier A, Dambrine E, Probst A. 1993. Interception in a mountainous declining spruce stand in the Strengbach catchment. J Hydrol 144:273–82.
Wang G. 2011. Assessing the potential hydrological impacts of hydraulic redistribution in Amazonia using a numerical modeling approach. Water Resour Res 47:W02528.
Warren JM, Meinzer FC, Brooks JR, Domec JC, Coulombe R. 2007. Hydraulic redistribution of soil water in two old-growth coniferous forests: quantifying patterns and controls. New Phytol 173:753–65.
West NE, Young JA. 2000. Intermountain valleys and lower mountain slopes. In: Barbour MG, Billings WD, Eds. North American terrestrial vegetation. Cambridge: Cambridge University Press. p 255–84.
Westerling AL, Hidalgo HG, Cayan DR, Swetnam TW. 2006. Warming and earlier spring increase western U.S. forest wildfire activity. Science 313:940–3.
Wetherald RT, Manabe S. 2002. Simulation of hydrologic changes associated with global warming. J Geophys Res 107:4379.
Wilcox BP. 2010. Transformative ecosystem change and ecohydrology: ushering in a new era for watershed management. Ecohydrology 3:126–30.
Wilcox BP, Seyfried MS, Breshears DD, McDonnell JJ. 2012. Ecohydrologic connections and complexities in drylands: new perspectives for understanding transformative landscape change. Ecohydrology 5:143–4.
Williams AP, Allen CD, Millar CI, Swetnam TW, Michaelsen J, Still CJ, Leavitt SW. 2010. Forest responses to increasing aridity and warmth in the southwestern United States. Proc Natl Acad Sci 107(50):21289–94.
Williams PA, Allen CD, Macalady AK, Griffin D, Woodhouse CA, Meko DM, Swetnam TW, Rauscher SA, Seager R, Grissino-Mayer HD, Dean JS, Cook ER, Gangodagamage C, Cai M, McDowell NG. 2012. Temperature as a potent driver of regional forest drought stress and tree mortality. Nat Clim Change 3(3):292–7.
Yarrow MM, Marín VH. 2007. Toward conceptual cohesiveness: a historical analysis of the theory and utility of ecological boundaries and transition zones. Ecosystems 10:462–76.
Yarrow MM, Salthe SN. 2008. Ecological boundaries in the context of hierarchy theory. Biosystems 92:233–44.
Yoder R, Odhiambo L, Wright W. 2005. Effects of vapor–pressure deficit and net-irradiance calculation methods on accuracy of standardized penman–monteith equation in a humid climate. J Irrigation Drainage Eng 131:228–37.
Zou CB, Breshears DD, Newman BD, Wilcox BP, Gard MO, Rich PM. 2008. Soil water dynamics under low- versus high-ponderosa pine tree density: ecohydrological functioning and restoration implications. Ecohydrology 1:309–15.
Acknowledgments
Eddy covariance and soil moisture data from Chimney Park presented in Appendix 4 were generously provided by Elise Pendall and Brent Ewers, processed by David Reed (all University of Wyoming), with funding support from McIntire-Stennis, Wyoming Water Development Commission, US Geological Survey and National Science Foundation through Emerging Topics in Biogeochemical Cycles. Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the US Government.
Author information
Authors and Affiliations
Corresponding author
Additional information
Author Contributions
JB, DS, and WL conceived of and designed the study; JB and DS performed research and analyzed the data; JB, DS, and WL wrote the paper.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Bradford, J.B., Schlaepfer, D.R. & Lauenroth, W.K. Ecohydrology of Adjacent Sagebrush and Lodgepole Pine Ecosystems: The Consequences of Climate Change and Disturbance. Ecosystems 17, 590–605 (2014). https://doi.org/10.1007/s10021-013-9745-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-013-9745-1