
Abstract
Brazil’s coastline contains landscape features that are favorable for mangrove development. By revisiting the 1990 paper entitled “Variability of Mangroves Along the Brazilian Coast,” we review aspects of the notion of variability, considering coasts among the most dynamic places on the planet, shifting from a theoretical to an empiric approach. In the previous work, hereafter referred to as the 1990 paper, we suggested mangroves developed in seven out of eight coastal segments inspired by the coastal environmental settings’ framework. We expand the previous work using concepts of complex systems theory to illuminate contemporaneous coastal management issues related to multiple spatial and temporal scales. Here, we suggest that these eight segments occur within three major process domains: Northernmost Domain, Central Deltaic Coast Domain, and Cabo Frio do Laguna Domain, where different factors work together to form geoecological characteristics that are unique, and irreplaceable if lost.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
1 Introduction
Considering the high dynamism of coastal systems, the advances in the concepts of complex systems theory to illuminate contemporaneous coastal management issues related to multiple spatial and temporal scales, and the recent knowledge produced on Brazilian mangroves and saltmarshes in this chapter, we revisit and expand the contribution presented by Schaeffer-Novelli et al. (1990) (hereafter referred to as the 1990 paper) to understand mangrove and salt marsh patterns and processes along the Brazilian coast and to discuss the originally proposed macroecological concepts.
The 1990 paper was written at a time when interest in mangroves on a global scale was beginning to blossom triggered mainly by a series of outstanding publications, that is, Chapman’s (1975) book on mangrove biogeography, Walsh’s (1974) seminal work on mangrove zonation, and the concept of “outwelling” by Odum (1971) and Odum and Heald (1975). Seminal papers such as The Ecology of Mangroves by Lugo and Snedaker (1974) were shifting attention from vegetation to ecological processes. Other major contributions were the UNESCO’s release of the world’s most thorough mangrove bibliographic survey since 1614 (Rollet 1981), and the Handbook for Mangrove Area Management (Hamilton and Snedaker 1984). The Association of Marine Science Researchers (ALICMAR) was created in 1974 and provided the first social-technical platform for sharing ecological knowledge in the Americas. At about the same time, the Organization of American States (OEA) and UNESCO’s Regional Office for Latin America subsidized the participation of researchers in various meetings in Hawaii, United States (1974), and Cali, Colombia (1978). Simultaneously, several academic institutions initiated local activities supported by national research organizations, which also sponsored academic exchanges, planting the seed of mangrove ecology on increasingly fertile ground.
Thus, the 1990 paper was an outgrowth of the convergence of attention from multiple Brazilian organizations being directed at coastal systems and the emerging recognition that ecological knowledge would contribute significantly to administer systems that were being recognized for their ecologic importance for coastal fisheries, whereas just a decade earlier, they had been misjudged as useless wastelands suitable for reclamation – a mindset that had prevailed since colonial times and was firmly entrenched in society. What was emerging since the 1960s and early 1970s (Odum 1969; Odum 1970) was the realization that perhaps these lands were not wastelands after all, and that they merited to be managed more rationally, in terms of what today is termed an ecosystem services-based approach (Gregory and Goudie 2011). Changing deeply seated mindsets requires considerable efforts, such as increasing research levels for understanding the behavior of these systems and building a cadre of human resources that could increase societal awareness to promote their conservation rather than their reclamation and transformation.
The 1990 paper adopted a mesoscale (landscape) perspective for convenience. Landscapes are the most tangible ecological criteria and remain an appropriate level of observation for broad-scale management as well as for focusing on smaller scales. In this respect, we propose a refinement of the mesoscale classification scheme used in the 1990 paper and argue that the application of the coastal environmental setting (CES) framework (Thom 1982; Woodroffe 1992; Twilley et al. 2018;) improves our capacity to appropriately understand and scale mangroves’ macroecological attributes and responses to natural and anthropogenic stressors at larger spatial and temporal scales.
In addition, later in this chapter, we discuss an approach we call dynamic framing as complementary for adopting a landscape perspective coherent with nature’s investment and endurance strategy. However, of critical importance is that the mesoscale landscape approach merges human social systems and geomorphic systems into a unitary system, and recognizes multiple interactive and intercausal scales, geomorphological, social, and ecological processes (Huggett 1995) that are interdependent and vital for sustainability.
2 Scales and Variability in Mangrove Macroecology
The Brazilian coast is characterized by mangrove forests along most of its 10,959.52 km length (IBGE 2016), between the latitudes 04°20′12″N and 33°45′07″S. The measured length of any coast is a function of the scale and resolution of the measurement (Mandelbrot 1983), so it is not surprising that various length estimates exist in the literature (see Chap. 1 for more details). On a global scale, Brazil’s coastline length ranks 12th (IBGE 2016), but shelters the second-largest mangrove area cover in the world (FAO 2007; Giri et al. 2011; Hamilton and Casey 2016). This suggests that from a broad perspective, this coast contains landscape features that are particularly favorable for mangrove development. Because a broad view subdues detail, processes, and structures, it is not surprising that a closer look often reveals unexpected variability at different levels.
Reality is complex and stratified into characteristic scales, dynamics, and patterns. These tend to be bundled into discrete scales of interaction (Rowe 1961; Simon 1962). Variability is the result of complexity; the diversity of components compounded by the spatiotemporal diversity of factors influences landscape responses and development at various scales, leaving distinct signatures that reveal dominant influences. The interaction between factor regimes and scales results in relatively distinct landforms. Thus, the original paper intended to examine zonality, that is, Brazilian coastal patterns, in terms of features and regional environments. The outcome was a proposed division of the coast into segments within which similar broad climatic, geomorphologic, and oceanographic features and comparable management needs are found. Mangroves were accounted as mostly azonal perhaps because of the dominance of local (site) factors in influencing development. Complexity defies any attempt of classifying any coast where the diversity of landscape elements is high and where these elements and forcing functions act in combination and interact in complex ways.
In the context of revisiting the 1990 paper, we find it desirable to review some aspects of the notion of variability, considering that coasts are the most dynamic places on the planet. It is misleading to consider coastal features as static or perceive variability as problematic. Variability is a manifestation of complexity and although it presents obstacles to generalizing and identifying clear-cut patterns in nature, it is part of it and is present at all scales driven by external and internal factors such as self-organization. Variability paradoxically entails the iterative power of order, of system-level responses that eventually can lead to adaptive change and the capture of environmental energies to gradually perform increasingly more complex geoecologic work that makes more complete use of all available energies.
Furthermore, categorizations are based on generalizations and variability tends to obscure categories. In fact, all categorization schemes are simplifications and ignore variability at some scale. In nature, absolute categories do not exist, because categories exist only as (human) ideas, whereas reality is a continuum; change, variability, and transformation are pervasive but until very recently we have perceived reality in terms of static components and neglected processes and change. A shift taking place in ecology is the increasing adoption of a process-based perspective. The relationships between ecological processes and spatiotemporal patterns on a variety of scales are one of the most relevant research topics for most unresolved questions in ecology. Even climate was accounted as constant until very recently. What is pertinent in this appreciation is that temporal variability is an important area of concern, because the temporal scope of human observation is often very limited when considering the extended endurance of many geomorphic features. Focusing on the narrow spatiotemporal window of human experience inevitably provides a “keyhole” or partial view of coastal systems that ignoring its limitations can undermine management efforts no matter how well-intended they might be. In general, ecological events have a characteristic frequency and a corresponding spatial scale, and an ecological study of the landscape must conform to these scales (Turner 1998; Blackburn and Gaston 2002).
The Brazilian coast has been divided into different segments by several authors for different purposes, highlighting certain features and processes. The complexity of responses from a complex system approach is obviously overwhelming. In our 1990 paper, the purpose was to highlight geographic variability in the context of settings for mangrove establishment and development. That paper did not intend to describe causal factors or system dynamics but was limited to describing patterns of mangrove structure along the coast in very broad terms. To do that we conveniently divided the coast into eight broad segments oriented to mangrove abiotic drivers (Fig. 3.1) such as climatological (temperature, precipitation, and potential evapotranspiration), hydrographic (river order rank), and oceanographic (tidal amplitude), including the resulting mangrove development (structure; see Table 3.1). That method was summarized by Schaeffer-Novelli et al. (2016).

Coastal segments proposed in Schaeffer-Novelli et al. (1990), divided by dots: I – Cape Orange (04°30′N) to Cape Norte (01°40′N), II – Cape Norte to Ponta Curuçá (00°36′S), III – Ponta Curuçá to Ponta Mangues Secos (02°15′S), IV – Ponta Mangues Secos to Cape Calcanhar (05°08′S), V – Cape Calcanhar to Recôncavo Baiano (13°00′S), VI – Recôncavo Baiano to Cabo Frio (23°00′S), VII – Cabo Frio to Torres (29°20′S), and VIII – Torres to Chuí (22°35′S)
Within seven out of the eight coastal segments (Table 3.1), mangroves occupy landforms that bear the signature of past legacies of dominant formative processes (Rovai et al. 2018) (Fig. 3.2). Geographic partitioning is a common tool for supporting spatial reasoning for deriving qualitative inferences from broad categories. Here, we strengthen the original mesoscale approach by incorporating the coastal geomorphology variability within each segment as originally proposed in the 1990 paper, following the elements that have been presented in Chaps. 1 and 2.

Coastal segments proposed in Schaeffer-Novelli et al. (1990) highlighting dominant environmental forcing’s and climate-driven threats. Mangroves are present in segments I–VII, salt marshes are predominant at segment VIII. (a) TP-mean annual temperature, (b) PT-mean annual precipitation, (c) PET-mean annual potential evapotranspiration, (d) TR-mean tidal range, and (e) River order rank. Sources: TP and PT data from Hijmans et al. (2005), PET from Title and Bemmels (2018), TR from Vestbo et al. (2018), and river order rank from Patterson and Kelso (2018)
3 The Coastal Environmental Setting (CES) Framework
Some fifty years ago, Bruce Thom proposed a framework based on ecogeomorphology to explain ecological regularities linked to different Coastal Environmental Setting (CESs). This is the framework used in the 1990 paper. These ideas were further developed by Rovai et al. (2018) and applied to multiple-scale ecological models to explain global variations in the mangrove ecosystem’s properties. Incorporating ecogeomorphic forcings into predictive models has helped to advance hypotheses that improve our understanding and capacity to foresee the effects of global changes in these ecosystems.
Ecology has made great strides since the 1980s when the 1990 paper was conceived; new notions, tools, and concepts have been developed and are taking increasingly important roles in expanding observational windows in quality and scope, furthering interpretation, and reinterpretation of data and previous analyses. These new tools and notions have interacted catalytically to broaden ecological knowledge greatly in time and space. Understanding and dissemination of knowledge now have achieved global scales and we now can speak of global or macroecology as a discrete research field. The beneficiary community has expanded as well, and now includes scientists, educators, resource managers, and large stakeholder communities. New tools have also become available such as remote sensing instruments including global positioning systems (GPS) and inexpensive portable sensors. Increasing computational power and progressively easier access to distant places and real-time communications among researchers has propitiated a revolution in ecology that is still taking place and continues at an increasing pace. Furthermore, there are now more universities, scientists, and engineers than ever before in history. More importantly, complex environmental issues are part of the public sphere or social spaces nowadays. Thus, there is an increased demand for scientific communication to nonprofessionals to promote greater public understanding and engagement by educated constituencies.
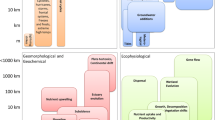
The 1990 paper’s perspective remains relevant as an appropriate level of observation for revealing mesoscale order as a starting point for a triadic approach that pays attention to events that take place at other levels: the focal level, the next higher level, and the level immediately below (Salthe 1985). Here we will demonstrate that the CES frameworkFootnote 1 provides the most obvious and tangible ground for improvement in spatial resolution, accessing constraints, and moving toward higher fidelity scales. All ecological processes and structures are multiscales (Allen and Hoekstra 1992). We provide a reanalysis of mangrove structural (biomass) and functional (primary productivity, carbon sequestration) attributes discussed in the 1990 paper. We used global compilations on climatic and oceanographic variables to predict mangrove ecological traits at a continental scale, expanding models proposed in our original 1990 paper from a conceptual to an empiric perspective. Particularly, we explored how the relative contribution of rivers, tidal range, along with regional climate, shapes distinct CES, reflected in substrate conditions to which plants respond (Thom 1982; Woodroffe 1992; Twilley et al. 2018) (Fig. 3.3). Distinct CES, for example, deltas, estuaries, and lagoons, are formed by the relative contribution of geophysical variables (e.g., river discharge, tidal amplitude, wave energy). Along with regional climatic drivers, these geophysical forcings constrain carbon partitioning among ecosystem compartments (soil, above- and belowground biomass). CES types include large rivers, small deltas (grouped as deltaic by Thom 1982), tidal systems (estuaries, bedrock as defined by Thom 1982), lagoons (including composite settings as defined by Thom 1982), carbonate coastal settings, and arheic or dry coastlines.

Coastal ecogeomorphology conceptual framework, showing how bidirectional fluxes between abiotic and biotic components control nutrient stoichiometry and carbon storage in mangroves. CES types: I – large rivers; II – small deltas; III – tidal systems; IV – lagoons; V – carbonate coastal settings; and VI – arheic, or dry coastlines. PET: potential evapotranspiration; C:N:P: carbon-to-nitrogen-to-phosphorus ratio. Adapted from Twilley et al. (2018)
The following brief, yet comprehensive overview on dominant global types of CES was originally summarized by Rovai et al. (2018). However, we suggest consultation of the original sources (Thom 1982; Woodroffe 1992) for additional information. One of the major factors defining the different CES is sediment source (i.e., river-borne), which represents a combination of geophysical processes and local geology influencing mangrove dynamics (Thom 1982; Woodroffe 1992; Woodroffe 2002).
The CES framework provides an alternative to the latitude gradient paradigm, and its use has advanced our capacity to predict mangrove ecological attributes such as aboveground biomass (Rovai et al. 2016), litterfall production (Ribeiro et al. 2019), and soil organic carbon (Rovai et al. 2018) at larger scales with a high confidence level. This is particularly useful for coastlines that lack such information. Here, we focus on the variability of mangrove aboveground biomass (AGB), litterfall (NPPL, or Net Primary Productivity Litterfall), and soil organic carbon (SOC) along the Brazilian coastline, as these ecosystem attributes constitute the largest long-term (>100 years), perennial carbon pools in mangrove forests. However, a pressing need remains for generating estimates of belowground biomass (roots) and productivity as these are significant components of ecosystem-level C stock and budget, respectively. CESs provide an ecological/terrain conceptual unit for management that is easily geographically defined.
4 Aboveground Biomass
Previous attempts to predict continental-scale mangrove aboveground biomass (AGB) include latitude (Saenger and Snedaker 1993; Twilley et al. 1992) and climate-based models (Hutchison et al. 2014). Although latitude-based models can indirectly encompass critical climatic and geophysical variables, their individual contribution to explain AGB value spatial patterns is unknown, since their explanatory power is not explicitly weighted in the statistical analysis. Although a climatic modeling approach explicitly includes climate variables such as temperature and precipitation to explain mangrove AGB at the global scale (Hutchison et al. 2014), this analysis is limited not only by the number of climatic variables included in the model but also by the lack of other environmental variables that directly influence mangrove structural and functional properties at regional and local scales (Twilley 1995; Twilley and Rivera-Monroy 2009).
A literature review to assemble a global dataset containing information on published mangrove AGB and forest structure data is summarized in a review by Rovai et al. (2016). The inclusion of other geophysical variables in the climatic-geophysical model significantly improves AGB estimates at the latitudinal scale as demonstrated for the neotropics. As in the conceptual model proposed in the 1990 paper, the review by Rovai et al. (2016) shows that at continental scales, higher tidal amplitudes contributed to high forest biomass associated with warm temperatures, abundant rainfall, and low potential evapotranspiration (Figs. 3.1a–c and 3.3a). For the Brazilian coast, this model corroborates the mangrove forest structural development described for each segment proposed in the 1990 paper (see Chaps. 4 and 6), with higher AGB values predicted for low latitude, deltaic and macrotidal coastlines (Segments I–III, Fig. 3.1; Table 3.1), and lower values along increasingly austral latitudes, tide- and wave-dominated, or dry coastlines (Segments IV and VII, Table 3.1; Fig. 3.4a).

Predicted mangrove aboveground biomass (a) (AGB in Mg ha–1), litterfall productivity (b) (NPPL in Mg ha–1 year–1), and soil organic carbon density (c) (SOC in mg cm–3) in Brazilian mangroves. Histograms depict the frequency of modeled values for each mangrove attribute. AGB data extracted from Rovai et al. (2016), NPPL, from Ribeiro et al. (2019), and SOC from Rovai et al. (2018)
Mangrove AGB values in Brazil range from 25.3 to 284.8 Mg ha–1 (mean = 95.8 Mg ha–1), within the range estimated for the neotropics (16.6–627.0 Mg ha–1, mean = 88.7 Mg ha–1) (Rovai et al. 2016). Using a biomass-to-carbon conversion factor of 0.475 (Hamilton and Friess 2018) and a mangrove forest cover of 7675 km2 (Hamilton and Casey 2016), the total C stored in mangroves’ AGB in Brazil is estimated at 0.04 PgC, which corresponds to 7.3% of global C stocks in mangrove AGB (Rovai et al. 2016).
Rovai et al. (2016) show that CES represents a major determinant on mangrove wetland development, configuration, and realized maximum biomass, particularly considering the diversity of mangrove geoecological settings and associated dynamics (Thom 1982; Woodroffe 1992; Twilley 1995). This energy signature is strongly influenced by the local tidal range and river discharge, critical geophysical variables explaining a significant percentage of the AGB total variance (Rovai et al. 2016). Indeed, tidal amplitude, a component of the hydroperiod regime in coastal regions, significantly influenced mangrove structural development. Higher tidal amplitude promotes nutrient exchange and aeration of soil layers, which reduces sulfide production and accumulation, allowing higher growth rates and forest development (Lugo and Snedaker 1974; Castañeda-Moya et al. 2013).
5 Net Primary Productivity – Litterfall (NPPL)
Ribeiro et al. (2019) provided the first model that accounts for continental-scale variability in mangrove Net Primary Productivity [Litterfall] (NPPL) in response to climatic and geophysical variables combined. Their results advance the current understating of mangrove NPPL variability across latitudinal and longitudinal gradients, considering that previous studies did not account for the role of geophysical forces in driving large-scale NPPL variability. Instead, correlations were usually performed using absolute variation in latitude degrees as a predictor of mangrove primary productivity (e.g., Twilley et al. 1992; Saenger and Snedaker 1993; Bouillon et al. 2008).
The model by Ribeiro et al. (2019) addresses a core question in mangrove macroecology, clarifying the role of factors that control mangrove NPPL at larger spatial scales. The authors show that mangrove NPPL is controlled by a combination of climatic (temperature and precipitation) and geophysical forces, such as tidal range. Here we used the model results by Ribeiro et al. (2019) for the neotropics to estimate NPPL for Brazilian mangroves (Fig. 3.4b). The predicted NPPL values for Brazilian mangroves ranged from 3.79 to 16.97 Mg ha–1 year–1 (mean = 10.92 Mg ha–1 year–1), and the range reported for the neotropics is 1.66–28.81 Mg ha–1 year–1 (mean = 10.25 Mg ha–1 year–1) (see Ribeiro et al. 2019 for details). Using a biomass-to-carbon conversion factor of 0.475 (see Hamilton and Friess 2018), the predicted mean NPPL for Brazilian mangroves corresponds to 5.5 Mg C ha–1 year–1. Using Hamilton and Casey’s (2016) estimative for Brazilian mangrove forest cover of 7675 km2, the annual rate of C removed from the atmosphere by mangrove NPPL in the country is estimated at 4 Tg C, which corresponds to 30% of total NPPL in the neotropics (Ribeiro et al. 2019).
Higher NPPL rates were predicted for mangrove forests influenced by large river systems, such as along the Amazon River coastline. These patterns of high NPPL rates predicted for river-dominated coastlines are consistent with observed values reported for other deltaic coastal settings in the neotropics such as in the San Juan River delta (Colombia), Orinoco River delta (Venezuela), and Essequibo River (Guyana) (see Ribeiro et al. 2019 for details). These regions with high NPPL are located in tropical regions subjected to low annual variability in temperature, high rates of rainfall (>2000 mm year–1) (Hijmans et al. 2005), and macrotidal regimes (Carrère et al. 2012). Conversely, the low rates of NPPL in Brazil were predicted for mangroves subjected to lower winter temperatures, reduced tidal amplitude (i.e., Segment VII, Table 3.1; Fig. 3.2), as well as reduced annual precipitation and reduced river discharge (Segment IV, Table 3.1; Fig. 3.2), which altogether constrain high primary productivity and forest development.
Ribeiro et al. (2019) showed that the interaction between precipitation and temperature accounted for most of the variability in mangrove NPPL across the neotropics. Temperature and precipitation regimes have long been described as important drivers of mangrove NPPL (Pool et al. 1975; Twilley 1995; Day et al. 1996; Feher et al. 2017). Temperature affects plants’ vital processes from photosynthesis and respiration to reproductive success and carbon storage (Duke 1990; Lovelock 2008). Similarly, rainfall also influences mangrove growth and primary production (Day et al. 1996; Twilley et al. 1997; Agraz-Hernández et al. 2015). Lower primary production has been reported for mangrove forests along dry coastlines, whereas the highest NPPL rates were related to areas with rainfall regimes over 2000 mm year−1 (Hernández and Mullen 1975; Félix-Pico et al. 2006; Lema and Polanía 2007). The synergism between temperature and precipitation regimes plays a major role in determining mangrove development and distribution (Spalding et al. 2010; Osland et al. 2016; Feher et al. 2017).
The results in Ribeiro et al. (2019) also highlighted the role of tidal regimes in mangrove NPPL variability at larger scales. These findings support previous studies that show a strong positive influence of tidal amplitudes in primary production (Cintrón and Schaeffer-Novelli 1981; Alongi 2002). Tides are an energy subsidy to mangroves’ primary production (Odum et al. 1982) and as this energy increases, so is the amount of organic matter exchanged between mangroves and adjacent environments (Twilley et al. 1986, 1992). Periodic tidal inundation promotes nutrient exchange and soil aeration, which reduces the accumulation of toxic substances (e.g., sulfides) and enhances forest development (Lugo and Snedaker 1974; Castañeda-Moya et al. 2013). In addition, earlier studies have shown tides to be a major driver of carbon allocation between above- and belowground compartments in mangrove forests. For instance, higher tides are frequently associated with well-developed mangrove forest stands (Cintrón and Schaeffer-Novelli 1981; Twilley 1995; Rovai et al. 2016). Conversely, mangrove root biomass was found to be higher in sites subjected to infrequent inundation (Castañeda-Moya et al. 2011; Adame et al. 2017). Similarly, higher soil organic carbon stocks have been negatively correlated with tides (Rovai et al. 2018). Also, the tidal amplitude is an important component of hydroperiod influencing mangrove species zonation (Crase et al. 2013) as well as the vertical range of suitable environment for mangrove establishment (Hutchings and Saenger 1987).
Although not selected as a significant term in the model by Ribeiro et al. (2019), potential evapotranspiration (PET) has been acknowledged as one of the major climatic factors determining the distribution of life zones on Earth (Holdridge 1967). PET represents the amount of water that could potentially be used by plants, but it is transferred back to the atmosphere through evaporation, thus, being an important regulator of forest water balance (Holdridge 1967). The interaction between PET and precipitation is especially important for mangroves, due to soil water content and salinity balance (Clough 1992; Wolanski et al. 1992). Indeed, PET has been shown to play a major role in the continental-scale variability of aboveground biomass and soil organic carbon stocks in mangroves (Rovai et al. 2016, 2018).
In equatorial climates, where temperatures are constantly high throughout the year, precipitation rates are moderate to high and the ratio between precipitation and PET is low, so mangrove forests can allocate more energy to their aboveground biomass and thus are better developed (Schaeffer-Novelli et al. 1990; Clough 1992). Where PET exceeds rainfall, the water deficit leads to decreased soil moisture, and consequently higher soil salinities, water stress on mangrove trees, and restricted forest development (Schaeffer-Novelli et al. 1990; Day et al. 1996; Castañeda-Moya et al. 2006). Moreover, the upper limit of distribution and survival of particular mangrove species is very often determined by soil salinity and soil water content, which are regulated by the conjunction of PET, rainfall, and tidal amplitude (Wolanski et al. 1992; Castañeda-Moya et al. 2006).
Furthermore, the influence of river discharge on mangrove ecosystems functioning is also indubitable. Nevertheless, excessive freshwater discharges act as a constraint by promoting competition by glicophytes that limits mangrove colonization. This is true in the Amazon estuary as well as south of Laguna (28°30′S) where freshwater habitats prevail displacing mangroves and favor freshwater marsh development. Overall, rivers are responsible for most of the freshwater input in mangroves, acting as a source of nutrients (phosphorus) and decreasing interstitial salinity (Pool et al. 1975; Castañeda-Moya et al. 2013). Riverine mangroves are characterized by optimal structural growth, with high values of aboveground biomass and NPPL resulting from high nutrient availability, abundant freshwater drainage, and reduced soil salinity levels, which are controlled by river discharge (Cintrón et al. 1978; Castañeda-Moya et al. 2006). River discharge is particularly important in dry (or arheic) coastlines such as in Northeast Brazil (see Chap. 1). In these dry climates, evapotranspiration exceeds the moisture supplied by precipitation, and river discharge becomes an important source of freshwater that controls salinity within limits that are not stressful for mangrove survival, forming extensive hypersaline flats (or “apicuns”).
The apicum (in singular) is a spatial-temporal ecogeomorphic feature of the mangrove ecosystem; it is a morphoclimatic hydrosere, a dynamic feature of the high intertidal zone, and technically a high salt marsh feature. The high salt marsh is influenced by precipitation, runoff, or seepage (Costa and Davy 1992; Hadlich et al. 2010). In dry coastlines with minor river discharge, massive mangrove diebacks can occur triggered by inland droughts, multidecadal fluctuations in sea level such as the 18.6-year Metonic Cycle (Munk et al. 2002), reductions in rainfall, and abnormally high air temperatures (Duke et al. 2017; Lovelock et al. 2017). During these events, only mangroves fringing estuary channels and upstream riverine stands remained healthy and mostly intact (Duke et al. 2017).
6 Soil Organic Carbon (SOC) Stocks
In the present work, we include a new mangrove ecological feature not covered in the 1990 paper, the continental-scale variability of Soil Organic Carbon (SOC) stocks in response to climatic and geophysical drivers. Mangroves have long been recognized for their potential role as a significant global carbon sink that may mitigate atmospheric CO2 enrichment (Twilley et al. 1992). They were recently recognized as the most carbon-dense forests in the tropics (Donato et al. 2011), culminating in an increase in research papers reporting mostly on local and regional carbon stocks. Few studies have attempted to deliver global mangrove carbon budgets (Chmura et al. 2003; Bouillon et al. 2008). Only recently have specific models been developed to account for global variation in mangrove SOC stocks (Jardine and Siikamäki 2014; Atwood et al. 2017; Rovai et al. 2018). Attention has been driven to SOC stocks, because most of the carbon in mangroves ecosystems is stored in this compartment (Twilley et al. 1992; Hamilton and Friess 2018), where it remains stable for much longer compared to AGB.
Rovai et al. (2018) demonstrated how local and regional estimates of SOC linked to CES can render a more realistic spatial representation of global mangrove SOC stocks. They combined 107 published and unpublished studies conducted worldwide to yield a dataset consisting of depth-integrated (top meter) mangrove SOC density values, reporting on 551 sites from 43 countries. In contrast to previous studies (e.g., Jardine and Siikamäki 2014; Atwood et al. 2017), this dataset included exclusively soil profiles that were at least 0.3 m in depth (which were then normalized to a depth of 1 m), and mangrove SOC density values obtained from elemental analyses or chemical determination (i.e., wet oxidation). Rovai et al. (2018) showed that the diversity of CESs can contribute to the global integration of complex geomorphological, geophysical, and climatic responses that explain the contribution of mangroves to global carbon sequestration. Their approach improved our capacity to predict the global contribution of coastal systems such as mangroves to carbon dynamics in the Earth system. Although their global mangrove SOC budget estimate was similar to early ones, for example, 2.3 PgC (Rovai et al. 2018) and 2.6 PgC (Atwood et al. 2017), they showed that mangrove SOC stocks vary markedly across different types of CESs, increasing from river- to tide/wave-dominated to carbonate coastlines. For example, a global estimate, recently provided by Atwood et al. (2017), used a country-level mean mangrove SOC stock of 283 Mg C ha–1 based on values from 48 countries to extrapolate global patterns for the remaining 57 countries that lack data on mangrove SOC. Results in the study by Rovai et al. (2018) indicate that for those countries, many of which comprise mostly carbonate CESs, the global mean reference value of mangrove SOC stocks suggested by Atwood et al. (2017) is about 50% lower than values based on distinct CESs. Moreover, their analysis showed that the CES framework has the potential to resolve unexpected patterns observed between carbonate and river-dominated coastal landforms identified in former global mangrove SOC budgets (Jardine and Siikamäki 2014). They showed that mangrove SOC stocks have been underestimated by up to 44% (a difference equivalent of roughly 200 MgC ha−1) and overestimated by up to 86% (around 400 MgC ha−1) in carbonate and deltaic settings, respectively, likely due to the omission of geomorphological and geophysical drivers in accounting for the large-scale variability of mangrove SOC stocks.
Here we used Rovai et al. (2018) results to compute estimates of SOC for Brazilian mangroves (Fig. 3.4c). Lower SOC density values were predicted for deltaic and macrotidal (Segments I–III, Table 3.1; Fig. 3.2) and arid (Segment IV, Table 3.1; Fig. 3.2) CESs. Higher SOC values were consistent along tide- and wave-dominated coastlines (Segments V–VII, Table 3.1; Fig. 3.2). Mangrove SOC stocks in the soil top meter in Brazil ranged from 72.1 to 388.3 MgC ha–1 (mean = 240.4 MgC ha–1), within the global range of 33.8 to 464.1 MgC ha–1 (mean = 296.6 MgC ha–1) (Rovai et al. 2018). Using the mangrove forest area of 7675 km2 (Hamilton and Casey 2016), the total carbon stored in mangroves soils in Brazil is estimated at 0.15 PgC, which corresponds to 6.5% of global SOC stocks in contrast to the 9.3% suggested earlier (Hamilton and Friess 2018).
7 Advancing the CES Framework: Challenges for Mangrove Macroecologists
Tremendous advances have been made recently in terms of mapping the global mangrove forest cover. The two most recent mangrove forest cover estimates range from nearly 82,000 (Hamilton and Casey 2016) to 132,000 km2 (Giri et al. 2011). This difference of approximately 40% in mangrove forest cover is due to different methodologies used to classify mangrove occurrence within each degree-cell. While the database in Hamilton and Casey (2016) (CGMFC-21)Footnote 2 estimates the percent cover for each degree-cell within a mangrove forest, the earlier database in Giri et al. (2011) (MFW)Footnote 3 uses a presence approach. Despite these methodological aspects, both CGMFC-21 and MFW databases have a very high resolution of approximately 900 m2 (30 × 30 m at the equator).
The parameters on which we based most of the discussion in this chapter (that is, AGB, NPPL, and SOC) were conveyed using the mangrove forest cover provided by the CGMFC-21 database but adjusted to a much lower fidelity (approximately 625 km2 or 25 × 25 km at the equator) than the original spatial resolution. As pointed out in the original sources, we based our analyses on Rovai et al. (2016 and 2018) and Ribeiro et al. (2019).
There are essentially two main reasons that may be preventing the development of robust higher-resolution large-scale mapping of mangrove ecological attributes. First, the attempt to balance the loss of information during the trade-off process of down- and upscaling data with different resolutions (Blackburn and Gaston 2002). Indeed, recent efforts in macroecology strived to consolidate a database of environmental variables that are thought to be relevant to species’ ecology and geographic distribution at a reasonable spatial resolution (0.08333° or approximately 8.3 km at the equator) (Title and Bemmels 2018). Even the WorldClim database (Hijmans et al. 2005), which has over 3000 citations, has resolutions that range from 1 to 340 km2. In this respect, some of the predictors used in the analyses we present here have a coarse native resolution, such as river discharge (0.5°). Thus, it is reasonable to work with an intermediary cell size (e.g., 0.25°) that is spatially representative of most CES domains, which the modeling framework is based on. Second, although the integration of information on mangrove typology based on local hydrology and topography (e.g., fringe vs. interior sites) would potentially allow for more robust local and global estimates, most papers in which the analyses presented here are based on do not include accurate information on hydroperiod. Accordingly, the spatial resolution of most global compilations on marine and terrestrial environmental variables (Title and Bemmels 2018) does not reflect the variability compatible with neither the CMFGC-21 nor the MFW database native resolution.
In order to perform a multiscale spatial analysis, both dependent and independent variables would have to be available at differing resolutions. Moreover, the set of environmental variables would have to hold ecological meaning across different spatial scales, which is unlikely as variables that control SOC formation in coastal wetlands differ at different scales (check Holmquist et al. 2018; Osland et al. 2018; and Rovai et al. 2018). While the scale-dependent issues discussed here are perhaps one of the major challenges mangrove ecologists will face when upscaling ecological traits from site-level observations, the CES framework resolved much of the dramatic difference in mangrove SOC estimates, particularly in terms of spatial variability with mangrove soil properties following close the energetic signature of distinct coastline types (Rovai et al. 2016, 2018; Twilley et al. 2018; Ribeiro et al. 2019).
8 CES Restrict the Atlantic South American Mangrove Limit
Laguna is an interesting threshold and is currently considered the southernmost limit of mangroves in Brazil (Cintrón and Schaeffer-Novelli 1981; Soares et al. 2012). However, it is attention-grabbing, because suitable habitats further south in the country seem to be present (Ximenes et al. 2018), yet mangroves as an ecosystem stop abruptly at Laguna (28°48′S). The mean sea surface temperatures here vary from 18.2 °C in summer to 16 °C in winter (Ximenes et al. 2018).
In a biogeographical terminus, this is a particularly interesting transitional zone, because it appears not only to be a limit to a species but to an ecosystem; at this geographic location, a regime or phase shift takes place. The discontinuity apparent in Laguna is a topic of great relevance to climate change research and the understanding of the future of mangroves in the region. Rather than mangrove expansion, the region may have been experiencing a contraction due to increasing freshwater dominance that might have resulted in a freshwater barrier blockage that now limits further mangrove expansion to the south beyond Laguna (Cintrón-Molero and Schaeffer-Novelli 2019) (Map 16).
Furthermore, because of potential conflict with agricultural land use in the Patos Lagoon region, it is likely that manmade attempts to restrict saline intrusions could further limit mangrove expansion to the south in the near future. Below Laguna is the 620 km coastal tract of Rio Grande do Sul State (Map 17), which encompasses South America America’s longest barrier structure, running almost uninterruptedly except near Cassino and Tramandaí inlets; the former is the inlet to the Patos Lagoon. Both are permanent openings due to the high freshwater discharges of the coastal lagoons behind the barrier.
Patos Lagoon’s extensive marshes are dominated by the genera Spartina, Juncus, Cyperus, Typha, Scirpus, Paspalum, and Sesuvium (Delaney 1962), which prevail in an eminently freshwater environment promoted by abundant rainfall water (P ≈ 1500 mm year−1), reduced potential evapotranspiration, high percolation rates, seepage, river flows, and microtidal regime (Hijmans et al. 2005; Carrère et al. 2013; Cohen et al. 2013). These occupy the biogeographic changeover zone that extends to northeastern Argentina (Costa and Davy 1992). The coast of the Rio Grande do Sul State, south of 34°S, is well known to receive rain throughout the year, including the passage of Mesoscale Convective Systems (MCS) (Houze Jr 2004), severe frontal systems as well as sporadic severe hail and frost events. The larger continental landmass at 10–25°S is conducive to the development of deep convective activity fed by Amazon moisture transport by a low-level jet into the area. This makes this area the most active MCS region in the world (Nesbitt and Zipser 2003).
Mangroves are documented to persist at the latitude of 38°45′S at Westport, Australia, where the mean annual atmospheric temperature is 18 °C and the coldest is 17 °C and where humid subtropical (Cfa) and maritime (Cfb) climate prevails (Peel et al. 2007). At Corner Inlet, Australia, they are found at 38°54′S. So, the abrupt phase shift at Laguna (Brazil) is a prominent feature that merits further and more detailed attention in the context of environmental change prediction. In any case, climate change is perhaps one of the most active research areas in present times, and southeastern Brazil and mangrove ecosystem dynamics offer fruitful research possibilities that would lead to understanding how climate influences coastal vegetation.
The southern domain is an area where planetary, regional, and local processes interact but where it is realistic to locate instrumentation to provide local-level data recordings and frequent site-level vegetation and interstitial salinity monitoring. This is a region where active climatological research is taking place and where climatology is of great interest because of its impact on agriculture and the local economy. This research is bound to help support new hypotheses about the distribution and abrupt limit of mangrove occurrence in this coastal segment.
9 Dynamic Framing and the Three Coastal Domains
The segments described, in the 1990 paper, are embedded within three broad domains that span the whole coast; they remain relevant to serve as guiding posts for versatile back-and-forth shifting of observation scales, an approach we have designated as dynamic framing. The three domains we identified are (Fig. 3.5)
-
The Northernmost Domain is highly moisture- and tide-subsidized and extends from the Guyanas and Amapá (Brazil, Cape Orange, Oiapoque River) to Cape São Roque).
-
The Central Deltaic Coast Domain extends from below Cape São Roque to Cabo Frio, as a domain characterized by warm temperatures but strong lateral constraints due to high levels of wave/energy.
-
The Cabo Frio to Laguna Domain, largely below the Tropic of Capricorn and increasingly influenced by cold frontal systems and the convective activity of the South Atlantic Convergence Zone. This portion of the Brazilian coast is periodically and strongly influenced by local, regional (South American Monsoon System, Robertson et al. 2005), and global forcings (e.g., ENSO).

Northernmost, Central Deltaic, and Cabo Frio to Laguna Coastal Domains
An apparent paradox by which muddy coasts act simultaneously as outwelling sources of biological organic matter while being geological sinks is resolved by recognizing a dialectical perspective between scales. In the short term, outwelling is notable and characteristic but over long temporal scales, deposition and accumulation prevail. This suggests that the CES scale integrates equilibrial and nonequilibrial dynamics at the scale of the whole system.
10 Final Remarks
It is misleading to consider coastal features as static or perceive variability as a disturbing feature. Variability is representative of complexity and although it presents obstacles to generalizing and identifying clear-cut patterns in nature, it is part of it and is present at all scales driven by external and internal factors, especially climate and self-organization. The emerging awareness about mangrove systems in sequestering carbon emissions and their contribution to climate regulation increases the relevance of continued research for education, developing robust conservation policy and for suggesting future research grounded in the emerging field of complexity science.
Notes
- 1.
The term Coastal Environmental Settings (CESs) refer to a typology of mangrove-occurring localities that share certain composed by geophysical, geomorphic, and biologic characteristics.
- 2.
CGMFC-21 (project): Continuous Global Mangrove Forest Cover for the Twenty-first Century.
- 3.
MFW (dataset): Mangrove Forest Cover Loss dataset.
References
Adame MF, Cherian S, Reef R, Stewart-Koster B (2017) Mangrove root biomass and the uncertainty of belowground carbon estimations. Forest Ecol Manag 403:52–60
Agraz-Hernández CM, Keb CAC, Iriarte-Vivar S, Venegas GP, Serratos BV, Sáenz JO (2015) Phenological variation of Rhizophora mangle and ground water chemistry associated to changes of the precipitation. Hydrobiologia 25(1):49–61
Allen TFH, Hoekstra TW (1992) Toward a unified ecology. Columbia University Press, New York
Alongi DM (2002) Present state and future of the world’s mangrove forests. Environ Conserv 29(3):331–349
Atwood TB, Connolly RM, Almahasheer H, Carnell PE, Duarte CM, Lewis CJE, Irigoien X, Kelleway JJ, Lavery PS, Macreadie PI, Serrano O, Sanders CJ, Santos I, Steven ADL, Lovelock CE (2017) Global patterns in mangrove soil carbon stocks and losses. Nat Clim Change 7(7):523–528
Blackburn TM, Gaston KJ (2002) Scale in macroecology. Glob Ecol Biogeogr 11(3):185–189
Bouillon S, Borges AV, Castañeda-Moya E, Diele K, Dittmar T, Duke NC, Kristensen E, Lee SY, Marchand C, Middelburg JJ, Rivera-Monroy VH, Smith TJ III, Twilley RR (2008) Mangrove production and carbon sinks: a revision of global budget estimates. Glob Biogeochem Cycles 22:2
Carrère L, Lyard F, Cancet M, Guillot A, Roblou L (2012) FES 2012: a new global tidal model taking advantage of nearly 20 years of altimetry. In: Ouwehand L (ed) 20 years of progress in radar altimetry, Venice, 24–29 September 2013
Carrère L, Lyard F, Cancet M, Guillot A, Roblou L (2013) FES 2012: a new global tidal model taking advantage of nearly 20 years of altimetry. In: Proceedings of ‘20 years of progress in radar altimetry’, 24–29 September 2012, Venice, Italy
Castañeda-Moya E, Rivera-Monroy VH, Twilley RR (2006) Mangrove zonation in the dry life zone of the Gulf of Fonseca, Honduras. Estuar Coast 29(5):751–764
Castañeda-Moya E, Twilley RR, Rivera-Monroy VH, Marx BD, Coronado-Molina C, Ewe SML (2011) Patterns of root dynamics in mangrove forests along environmental gradients in the Florida Coastal Everglades, USA. Ecosystems 14(7):1178–1195
Castañeda-Moya E, Twilley RR, Rivera-Monroy VH (2013) Allocation of biomass and net primary productivity of mangrove forests along environmental gradients in the Florida Coastal Everglades, USA. For Ecol Manag 307:226–241
Chapman VJ (1975) Mangrove vegetation. J. Cramer, Vaduz
Chmura GL, Anisfeld SC, Cahoon DR, Lynch JC (2003) Global carbon sequestration in tidal, saline wetland soils. Glob Biogeochem Cycle 17(4):22–1–22–12
Cintrón G, Schaeffer-Novelli Y (1981) Los manglares de la costa brasileña: revisión preliminar de la literatura. In: Informe Técnico preparado para la Oficina Regional de Ciencia y Tecnología para América Latina y el Caribe de Unesco y la Universidad Federal de Santa Catarina (ed/rev: Abuchahla GMO). UNESCO, p 47. http://www.producao.usp.br/handle/BDPI/43826
Cintrón G, Lugo AE, Pool DJ, Morris G (1978) Mangroves of arid environments in Puerto Rico and adjacent islands. Biotropica 10(2):110–121
Cintrón-Molero G, Schaeffer-Novelli Y (2019) The role of atmospheric-tropospheric rivers in partitioning coastal habitats and limiting the poleward expansion of mangroves along the southeast coast of Brazil. Int J Hydrol 3(2):92–94
Clough BF (1992) Primary productivity and growth of mangrove forests. In: Robertson AI, Alongi DM (eds) Tropical mangrove ecosystems. American Geophysical Union, Washington, DC, pp 225–249
Cohen S, Kettner AJ, Syvitski JPM, Fekete BM (2013) WBMsed, a distributed global-scale riverine sediment flux model: model description and validation. Comput Geosci 53:80–93
Costa CSB, Davy AJ (1992) Coastal Saltmarsh communities of Latin America. In: Seeliger U (ed) Coastal plant communities of Latin America. Academic, San Diego, pp 179–199
Crase B, Liedloff A, Vesk PA, Burgman MA, Wintle BA (2013) Hydroperiod is the main driver of the spatial pattern of dominance in mangrove communities. Glob Ecol Biogeogr 22:806–817
Day JW Jr, Coronado-Molina C, Vera-Herrera FR, Twilley R, Rivera-Monroy VH, Alvarez-Guillen H, Day R, Conner W (1996) A 7 year record of above-ground net primary production in a southeastern Mexican mangrove forest. Aquat Bot 55:39–60
Delaney PJV (1962) Quaternary geologic history of the coastal plain of Rio Grande do Sul, Brazil. S Am Coast Stu Techn Rep 10(A):1–63
Donato DC, Kauffman JB, Murdiyarso D, Kurnianto S, Stidham M, Kanninen M (2011) Mangroves among the most carbon-rich forests in the tropics. Nat Geosci 4(5):293–297
Duke NC (1990) Phenological trends with latitude in the mangrove tree Avicennia marina. J Ecol 78(1):113–133
Duke NC, Kovacs JM, Griffiths AD, Preece L, Hill DJE, van Oosterzee P, Mackenzie J, Morning HS, Burrows D (2017) Large-scale dieback of mangroves in Australia’s Gulf of Carpentaria: a severe ecosystem response, coincidental with an unusually extreme weather event. Mar Freshwater Res 68(10):1816–1829
Feher LC, Osland MJ, Griffith KT, Grace JB, Howard RJ, Stagg CL, Enwright NM, Kraus KW, Gabler CA, Day RH, Rogers K (2017) Linear and nonlinear effects of temperature and precipitation on ecosystem properties in tidal saline wetlands. Ecosphere 8(10):e01956
Félix-Pico E, Holguín-Quiñones O, Hernández-Herrera A, Flores-Verdugo F (2006) Mangrove primary production at El Conchalito Estuary in La Paz Bay (Baja California Sur, Mexico). Cienc Mar 32(1A):53–63
Fisheries and Aquaculture Department – FAO (2007) The world’s mangroves 1980–2005. FAO Romefos
Giri CE, Ochieng E, Tieszen LL, Zhu Z, Singh A, Loveland T, Masek J, Duke NC (2011) Status and distribution of mangrove forests of the world using earth observation satellite data. Glob Ecol Biogeogr 20:154–159
Gregory KJ, Goudie AS (2011) The Sage handbook of geomorphology. Sage, Los Angeles
Hadlich GM, Celino JJ, Ucha JM (2010) Diferenciação físico-química entre apicuns, manguezais e encostas na Baía de Todos os Santos, Nordeste do Brasil. Geociências 29(4):633–641
Hamilton SE, Casey D (2016) Creation of a high spatio-temporal resolution global database of continuous mangrove forest cover for the 21st century (CGMFC-21). Glob Ecol Biogeogr 25(6):729–738
Hamilton SE, Friess DA (2018) Global carbon stocks and potential emissions due to mangrove deforestation from 2000 to 2012. Nat Clim Change 8(3):240–244
Hamilton SE, Snedaker SC (eds) (1984) Handbook for mangrove area management. Environment and Policy Institute East-West Center, International Union for the Conservation of Nature and Natural Resources & United Nations Educational, Scientific and Cultural Organization
Hernández A, Mullen K (1975) Produtividad primaria neta en un manglar del Pacífico Colombiano. In: Memorias del Seminario Sobre El Pacífico Colombiano. Universidad del Valle, Cali
Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A (2005) Very high-resolution interpolated climate surfaces for global land areas. Int J Climatol 25:1965–1978
Holdridge LR (1967) Life zone ecology. Tropical Science Center, San José
Holmquist JR, Windham-Myers L, Bliss N, Crooks S, Morris JT, Megonigal JP, Troxler T, Weller D, Callaway J, Drexler J, Ferner MC, Gonneea ME, Kroeger KD, Schile-Beers L, Woo I, Buffington K, Breithaupt J, Boyd BM, Brown LN, Dix N, Hice L, Horton BP, MacDonald GM, Moyer RP, Reay W, Shaw T, Smith E, Smoak JM, Sommerfield C, Thorne K, Velinksy D, Watson E, Grimes KW, Woodrey M (2018) Accuracy and precision of tidal wetland soil carbon mapping in the conterminous United States. Sci Rep-UK 8(1):9478
Houze RA Jr (2004) Mesoscale convective systems. Rev Geophys 42(RG4003)
Huggett RJ (1995) Geoecology: an evolutionary approach. Routledge, New York City
Hutchings P, Saenger P (1987) Ecology of mangroves. University of Queensland Press, St Lucia
Hutchison J, Manica A, Swetnam R, Balmford A, Spalding M (2014) Predicting global patterns in mangrove forest biomass. Conserv Lett 7(3):233–240
Instituto Brasileiro de Geografia e Estatistica – IBGE (2016) Caracterização do território: posição e extensão. In: Anuário Estatístico do Brasil, vol 1. IBGE, Rio de Janeiro
Jardine SL, Siikamäki JV (2014) A global predictive model of carbon in mangrove soils. Environ Res Lett 9(10):104013
Lema LF, Polanía J (2007) Estructura y dinámica del manglar del delta del río Ranchería. Caribe colombiano. Rev Biol Trop 55(1):11–21
Lovelock CE (2008) Soil respiration and belowground carbon allocation in mangrove forests. Ecosystems 11(2):342–354
Lovelock CE, Feller IC, Reef R, Hickey S, Ball MC (2017) Mangrove dieback during fluctuating sea levels. Sci Rep-UK 7(1):1–8
Lugo AE, Snedaker SC (1974) The ecology of mangroves. Annu Rev Ecol Syst 5(1):39–64
Mandelbrot BB (1983) The fractal geometry of nature, 1st edn. WH Freeman, New York City
Munk W, Dzieciuch M, Jayne S (2002) Millennial climate variability: is there a tidal connection? J Clim 15:370–385
Nesbitt SW, Zipser EJ (2003) The diurnal cycle of rainfall and convective intensity according to three years of TRMM measurements. AMS J Clim 16:1456–1475
Odum EP (1969) The strategy of ecosystem development. Science 164:262–270
Odum WE (1970) Pathways of energy flow in a South Florida Estuary. PhD dissertation, University of Miami
Odum WE, Heald EJ (1975) Mangrove forests and aquatic productivity. In: An introduction to land-water interactions. Springer, Berlin/Heidelberg, pp 129–136
Odum WE, McIvor CC, Smith TJ III (1982) The ecology of the mangroves of South Florida: a community profile. US Fish and Wildlife Service – Office of Biological Services, Washington, DC
Osland MJ, Enwright NM, Day RH, Gabler CA, Stagg CL, Grace JB (2016) Beyond just sea-level rise: considering macroclimatic drivers within coastal wetland vulnerability assessments to climate change. Glob Change Biol 22:1–11
Osland MJ, Gabler CA, Grace JB, Day RH, McCoy ML, McLeod JL, From AS, Enwright NM, Feher LC, Stagg CL, Hartley SB (2018) Climate and plant control on soil organic matter in coastal wetlands. Glob Change Biol 24(11):5361–5379
Patterson T, Kelso NV (2018) Natural earth. https://www.naturalearthdata.com/downloads/10m-physical-vectors/10m-rivers-lake-centerlines/
Peel MC, Finlayson BL, McMahon TA (2007) Updated world map of the Köppen-Geiger climate classification. Hydrol Earth Syst Sci 5:439–473
Pool DJ, Lugo AE, Snedaker SC (1975) Litter production in mangrove forests of southern Florida and Puerto Rico. In: Proceedings of the international symposium on biology and management of mangroves, Honolulu, 8–11 October 1974
Ribeiro RA, Rovai AS, Twilley RR, Castañeda-Moya E (2019) Spatial variability of mangrove primary productivity in the neotropics. Ecosphere 10(8):e02841
Robertson AW, Mechoso CR, Ropelewski CF, Grimm AM (2005) The American monsoon systems. In: Chang C-P, Wang B, Lau N-CG (eds) The global monsoon system: research and forecast. Secretariat of the World Meteorological Organization, Geneva, pp 197–206
Rovai AS, Riul P, Twilley RR, Castañeda-Moya E, Rivera-Monroy VH, Williams AA, Simard M, Cifuentes-Jara M, Lewis RR, Crooks S, Horta PA, Schaeffer-Novelli Y, Cintrón G, Pozo-Cajas PPR (2016) Scaling mangrove aboveground biomass from site-level to continental-scale. Glob Ecol Biogeogr 25(3):286–298
Rovai AS, Twilley RR, Castañeda-Moya E, Riul P, Cifuentes-Jara M, Manrow-Villalobos M, Horta PA, Simonassi JC, Fonseca AL, Pagliosa PR (2018) Global controls on carbon storage in mangrove soils. Nat Clim Change 8(6):534–538
Rowe JS (1961) The level-of-integration concept and ecology. Ecology 42(2):420–427
Saenger P, Snedaker SC (1993) Pantropical trends in mangrove above-ground biomass and annual litterfall. Oecologia 96(3):293–299
Schaeffer-Novelli Y (1999) Situação atual do grupo de ecossistemas: manguezal, marisma e apicum, incluindo os principais vetores de pressão e as perspectivas para sua conservação e usos sustentável. Avaliação e ações prioritárias para a conservação da biodiversidade da zona costeira e marinha. Projeto de Conservação e Utilização Sustentável da Diversidade Biológica Brasileira – PROBIO. Report BDT_mangue-1999
Schaeffer-Novelli Y, Cintrón-Molero G, Adaime RR, Camargo TM (1990) Variability of mangrove ecosystems along the Brazilian coast. Estuaries 13(2):204–218
Schaeffer-Novelli Y, Soriano-Sierra EJ, Vale CC, Bernini E, Rovai AS, Pinheiro MAA, Schmidt AJ, Almeida R, Coelho-Jr C, Menghini RP, Martinez DI, Abuchahla GMO, Cunha-Lignon M, Charlier-Sarubo S, Shirazawa-Freitas J, Cintrón-Molero G (2016) Climate changes in mangrove forests and salt marshes. Braz J Oceanogr 64(sp2):37–52
Simon HA (1962) The architecture of complexity. Proc Am Phils Soc 106(6):467–482
Soares MLG, Estrada GCD, Fernandez V, Tognella MMP (2012) Southern limit of the Western South Atlantic mangroves: assessment of the potential effects of global warming from a biogeographical perspective. Estuar Coast Shelf S 101:44–53
Spalding M, Kainuma M, Collins L (2010) The world Atlas of mangroves. Earthscan, London
Thom BG (1982) Mangrove ecology – a geomorphological perspective. In: Clough BF (ed) Mangrove ecosystems in Australia: structure, function and management. Australian National University Press, Canberra, pp 3–17
Title PO, Bemmels JB (2018) ENVIREM: an expanded set of bioclimatic and topographic variables increases flexibility and improves performance of ecological niche modeling. Ecography 41(2):291–307
Turner MG (1998) Landscape ecology. In: Dodson SI, Allen TFH, Carpenter SR, Ives AR, Jeanne RL, Kichell JF, Langston NE, Turner MG (eds) Ecology. Oxford University Press, New York/Oxford, pp 77–122
Twilley RR (1995) Properties of mangrove ecosystems related to the energy signature of coastal environments. In: Hall CAS (ed) Maximum power: the ideas and applications of H. T. Odum. University Press of Colorado, Denver, pp 43–62
Twilley RR, Rivera-Monroy VH (2009) Ecogeomorphic models of nutrient biogeochemistry for mangrove wetlands. In: Perillo GME, Wolanski E, Cahoon DR, Brinson MM (eds) Coastal wetlands: an integrated ecosystem approach. Elsevier BV, Dordrecht, pp 641–683
Twilley RR, Lugo AE, Patterson-Zucca C (1986) Litter production and turnover in basin mangrove forests in southwest Florida. Ecology 67(3):670–683
Twilley RR, Chen R, Hargis T (1992) Carbon sinks in mangroves and their implications to carbon budget of tropical coastal ecosystems. Water Air Soil Poll 64:265–288
Twilley RR, Pozo M, Garcia VH, Rivera-Monroy VH, Zambrano R, Bodero A (1997) Litter dynamics in riverine mangrove forests in the Guayas river estuary, Ecuador. Oecologia 111(1):109–122
Twilley RR, Rovai AS, Riul P (2018) Coastal morphology explains global blue carbon distributions. Front Ecol Environ 16(9):1–6
Vestbo S, Obst M, Fernandez FJQ, Intanai I, Funch P (2018) Present and potential future distributions of Asian Horseshoe crabs determine areas for conservation. Front Mar Sci 5:164
Walsh GE (1974) Mangroves: a review. In: Reimhold RJ, Queen WH (eds) Ecology of halophytes. Academic, New York, pp 51–174
Wolanski E, Mazda Y, Ridd P (1992) Mangrove hydrodynamics. In: Tropical mangrove ecosystems. American Geophysical Union, Washington, DC, pp 43–62
Woodroffe CD (1992) Mangrove sediments and geomorphology. In: Robertson AI, Alongi DM (eds) Tropical mangrove ecosystems. American Geophysical Union, Washington, DC, pp 7–41
Woodroffe CD (2002) Coasts: form, process and evolution. Cambridge University Press, Cambridge
Ximenes AC, Ponsoni L, Lira CF, Koedam N, Dahdouh-Guebas F (2018) Does sea surface temperature contribute to determining range limits and expansion of mangroves in Eastern South America (Brazil)? Remote Sens-Basel 10:1787
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
1 Electronic Supplementary Materials
Map 1
Amapá State, Brazil: Mangrove, salt flat, and salt marsh areas. (Sources indicated in the legend) (DOCX 1609 kb)
Map 2
Coastline of Pará State, Brazil: Mangrove, salt flat, and salt marsh areas (DOCX 1239 kb)
Map 3
Coastline of Maranhão State, Brazil: Mangrove, salt flat, and salt marsh areas (DOCX 1984 kb)
Map 4
Coastline of Piauí State, Brazil: Mangrove, salt flat, and salt marsh areas (DOCX 1118 kb)
Map 5
Coastline of Ceará State, Brazil: Mangrove, salt flat, and salt marsh areas (DOCX 1740 kb)
Map 6
Coastline of Rio Grande do Norte State, Brazil: Mangrove, salt flat, and salt marsh areas (DOCX 1413 kb)
Map 7
Coastline of Paraíba State, Brazil: Mangrove, salt flat, and salt marsh areas (DOCX 1366 kb)
Map 8
Coastline of Pernambuco State, Brazil: Mangrove, salt flat, and salt marsh areas (DOCX 1623 kb)
Map 9
Coastline of Alagoas State, Brazil: Mangrove, salt flat, and salt marsh areas (DOCX 1524 kb)
Map 10
Coastline of Sergipe State, Brazil: Mangrove, salt flat, and salt marsh areas (DOCX 1634 kb)
Map 11
Coastline of Bahia State, Brazil: Mangrove, salt flat, and salt marsh areas (DOCX 2064 kb)
Map 12
Coastline of Espírito Santo State, Brazil: Mangrove, salt flat, and salt marsh areas (DOCX 2067 kb)
Map 13
Coastline of Rio de Janeiro State, Brazil: Mangrove, salt flat, and salt marsh areas (DOCX 2031 kb)
Map 14
Coastline of São Paulo State, Brazil: Mangrove, salt flat, and salt marsh areas (DOCX 1704 kb)
Map 15
Coastline of Paraná State, Brazil: Mangrove, salt flat, and salt marsh areas (DOCX 1889 kb)
Map 16
Coastline of Santa Catarina State, Brazil: Mangrove, salt flat, and salt marsh areas (DOCX 1868 kb)
Map 17
Coastline of Rio Grande do Sul State, Brazil: Mangrove, salt flat, and salt marsh areas (DOCX 2130 kb)
Map 18
Fernando de Noronha, Pernambuco State, Brazil: Sueste mangrove forest indicated in green (DOCX 192 kb)
Rights and permissions
Copyright information
© 2023 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Cintrón-Molero, G. et al. (2023). Variability of Mangroves Along the Brazilian Coast: Revisiting. In: Schaeffer-Novelli, Y., Abuchahla, G.M.d.O., Cintrón-Molero, G. (eds) Brazilian Mangroves and Salt Marshes. Brazilian Marine Biodiversity . Springer, Cham. https://doi.org/10.1007/978-3-031-13486-9_3
Download citation
DOI: https://doi.org/10.1007/978-3-031-13486-9_3
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-13485-2
Online ISBN: 978-3-031-13486-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)