
Abstract
We investigate the importance of quorum sensing in the success of house-hunting of emigrating Temnothorax ant colonies. Specifically, we show that the absence of the quorum sensing mechanism leads to failure of consensus during emigrations. We tackle this problem through the lens of distributed computing by viewing it as a natural distributed consensus algorithm. We develop an agent-based model of the house-hunting process, and use mathematical tools such as conditional probability, concentration bounds and Markov mixing time to rigorously prove the negative impact of not employing the quorum sensing mechanism on emigration outcomes. Our main result is a high probability bound for failure of consensus without quorum sensing in a two-new-nest environment, which we further extend to the general multiple-new-nest environments. We also show preliminary evidence that appropriate quorum sizes indeed help with consensus during emigrations. Our work provides theoretical foundations to analyze why Temnothorax ants evolved to utilize the quorum rule in their house-hunting process.
Access provided by Autonomous University of Puebla. Download conference paper PDF
Similar content being viewed by others

Keywords
1 Introduction
Social insect colonies are motivated to move the locations of their nesting site as a functional response to various selected forces, such as colony growth, competition, foraging efficiency, microclimate, nest deterioration, nest quality, parasitism, predation, and seasonality [18]. Through constant adaptation to a changing environment, many social insect species such as ants, termites, and bees have evolved robust algorithms to accomplish the task of collective nest relocation [32]. In this paper, we study one such algorithm observed in colonies of Temnothorax ants.

(Both photographs by S. C. Pratt.)
From [14] (Fig. 2). (a) Recruitment via tandem running in the ant of genus Temnothorax. The worker at the front is leading a tandem run, and the follower behind is about to signal its presence by tapping with its antennae on the gaster of the leader. (b) Recruitment by transport in Temnothorax ants. One worker is simply carrying another quickly to the new nestsite.
Temnothorax ant colonies have many biological constraints: individuals with limited memory and computational power, limited communication, and no central control. Despite that, colonies as a whole can reach various global goals such as nest-site selections and foraging [10]. Their remarkable collective intelligence is not only an interesting problem for biologists, but also inspiring for the computer science community. In particular, from the distributed computing perspective, the collective house-hunting behavior is closely related to the fundamental problem of consensus. Building a theoretical understanding of the key mechanisms in the house-hunting process can thus shed light on the designs of novel distributed consensus algorithms.
Colonies consist of active ants who move the remaining passive workers, the queen, and brood items (immature ants) [4, 25]. All workers are female ants. At the beginning of an emigration event, individual active ants independently search for new nest sites. If an ant finds one, she evaluates the site’s quality according to various metrics [7, 12]. Quality evaluation is relative to the old home nest [3]. If she is not satisfied with the site, she keeps searching. Otherwise if she is satisfied with the site, she returns to the home nest after some time interval that is inversely related to the new nest site quality; during this interval she might continue searching for other new potential nest sites [15, 22]. If she returns to the old nest, she recruits another active ant to the site by leading a slow tandem run from the old nest to the new site [19, 26]. This is done by the leader ant directing the follower ant along a pheromone trail (Fig. 1(a)). Upon arriving at the nest, the follower ant also evaluates the nest’s quality independently of the leader ant. Both ants then continue monitoring the quality of the nest and repeat the process of quality estimation, wait interval/continued search, and further recruitment [29].
An ant continues leading tandem runs until she perceives that the new nest’s population has exceeded a threshold, or quorum [23]. At this point, she ceases tandem runs and instead starts transporting other ants by picking one up and carrying her from the old home nest to the new nest (Fig. 1(b)). These transports are much faster than tandem runs, and they are largely directed at the passive workers and brood items, hence they serve to quickly move the entire colony to the new nest [22, 25]. The transporter rarely drops out of transporting other ants, and hence is considered fully committed to the new nest as the colony’s home [29].
Both tandem runs and transports are forms of recruitment to accelerate the emigration process, but the marginal benefits of transports in ensuring consensus remain relatively poorly understood. Previous studies have regarded the quorum sensing mechanism as a way to tune the speed-accuracy trade-off [9, 16, 17, 20, 24, 31], where a smaller quorum prompts ants to commit sooner (higher speed) to a nest that has accumulated enough population, although that nest could be inferior to another nest that is discovered later in the process (lower accuracy). However, these studies generally equate accuracy with consensus or cohesion [5, 6], when all or most ants commit to the same nest. The difference between accuracy and consensus is that the former evaluates the individuals’ ability to choose the best option in the environment, but the latter is concerned only with their ability to agree with each other. The ability to stay in a single group is not only an interesting algorithmic question, but also highly beneficial for the survival of these ant colonies [6, 9, 14, 30]. However, consensus during emigrations has been comparatively understudied. Such studies require examinations of both consensus cases and split cases, and the latter is difficult to induce experimentally. Therefore, in this paper, we conduct one of the first theoretical studies of the role of quorum sensing in emigration consensus.
At the outset, quorum sensing significantly benefits consensus because once enough ants make their choice, that choice is “locked in” and has a higher chance of becoming the final choice. This helps to ensure consensus when there are many choices and the search effort is dispersed. However, a closer look reveals that the quorum size must be carefully chosen. If the quorum size is too large, it would be very unlikely to be reached by any nest; if it is too small, multiple nests will likely reach quorum (a split), incurring significant additional costs in time and risk of exposure of the emigration [1, 2, 23]. These trade-offs pose the question of whether quorums help with consensus at all. In this paper, we aim to answer this question partially by investigating the probability of emigration consensus without the quorum sensing mechanism.
We start by modeling individual active ants as coupled random processes without considering the quorum sensing mechanism. Unlike in most classical distributed algorithms, the ants in our model do not receive initial input preferences, but must determine these preferences through exploration. Another difference is that our consensus requirement exempts a small portion of ants from committing to the same nest. Intuitively, we expect that the distribution of ant states converges to a limiting distribution in the long run. However, due to the probabilistic modeling, there is a non-zero probability that an emigration deviates greatly from this expectation, and this probability depends also on how many ants can be exempted by the requirement. Therefore, detailed calculations are needed to quantify the probability of deviations that satisfy the consensus requirement. Using probability tools such as conditional probability, concentration bounds and Markov mixing time, we then show that without quorum sensing, the probability of consensus is small and decays to zero exponentially fast as the colony size grows. In addition, we show preliminary evidence that appropriate quorum sizes indeed help with consensus during emigrations.
The rest of the paper is organized as follows. In Sect. 2, we present our model of individual ants, of the entire colony, and of an execution, for two-nest environments. In Sect. 3, we formally state the definition of consensus, and the metrics to measure a model’s performance in terms of consensus. In Sect. 4, we show that with a high probability, emigrations cannot eventually reach consensus without quorum sensing. In Sect. 5, we extend our results to general \(k-\)nest environments where \(k>2\). Then, in Sect. 6, we consider the addition of the quorum sensing mechanism to the emigration process in two-nest environments, and show simulation results on the quorum sizes that are sufficient for consensus.

State transition diagram for ant \(a_i\) during round \(t+1\) before/without quorum attainment. \(\alpha _1^i(t+1)\) and \(\alpha _2^i(t+1)\) are composite functions each including the probabilities of an ant taking different paths (independent discovery or tandem running) to transition out of \(n_0\) into \(n_1\) and \(n_2\), respectively.
2 Model
2.1 Timing Model and the Environment
We divide time into discrete rounds. Individual active ants are modeled as identical probabilistic finite state machines and their dynamics are coupled through recruitment actions, as described later in Sect. 2.2. Let N denote the total number of active ants in the colony. Note that passive ants, the queen, and brood items can only be transported and have no states. For ease of exposition, in the sequel, by an “ant” we mean an “active ant”. Each ant starts a round with its own state. During each round, ants can perform various state transitions and have new states, before all entering the next round at the same time. Throughout the paper, the state of an ant at round t refers to her state at the end of round t.
The environment contains the original home nest \(n_0\) and two new nests \(n_1\) and \(n_2\). The new nests \(n_1\) and \(n_2\) have qualities \(q_1\) and \(q_2\) respectively, relative to the home nest quality. For the convenience of our analysis, we let \(0<q_2<q_1\le 1\), where a higher value corresponds to a better nest. Each nest is also associated with a population that changes from round to round. We use \(x_0(t)N\), \(x_1(t)N\) and \(x_2(t)N\), where \(x_0(t) + x_1(t) + x_2(t) = 1\), to denote (active) ant populations in nest \(n_0\), \(n_1\) and \(n_2\) respectively at the end of round t. Initially, individual ants have no information on \(q_1\) and \(q_2\).
2.2 Model of Individual Ants Without Quorums
In this subsection, we describe the dynamics of an ant without quorums (a.k.a. without performing state transitions based on seeing a quorum), compactly illustrated in Fig. 2 and Eq. (1)–(6). Though these dynamics are not Markovian as the state transition of an ant is influenced by other ants during recruitments (tandem runs), we prove (in Sect. 4) that after a finite time, the state transitions of an ant become independent of the others’ states.
Individual State. The set of possible states of an ant is denoted as \({\mathcal {S}}\triangleq \left\{ n_0, n_1, n_2 \right\} \). Each state \(n_i\) refers to the ant being at nest \(n_i\), and thus in the sequel we use “in state \(n_i\)” and “in nest \(n_i\)” interchangeably. Denote the state of ant \(a_i\) at the end of round t as \(\boldsymbol{s}_{i}(t)\) with \(\boldsymbol{s}_{i}(0) = n_0\) for all \(a_i\), i.e., initially all ants locate at the home nest \(n_0\).
Transitions out of the Home Nest. In a round, an ant \(a_i\) in \(n_0\) can be recruited by following a tandem run to either \(n_1\) or \(n_2\). If \(a_i\) is not recruited, she discovers nest \(n_1\) or \(n_2\) for the first time through independent discovery with probability \(\alpha \in (0,1/2]\) for either nest and a total discovery probability of \(2\alpha \). Note that the biological meaning of the parameter \(\alpha \) is that it encodes the home nest quality - the higher the home nest quality, the less likely \(a_i\) is to search for a new nest during any round t and the smaller \(\alpha \) is. Recruitment takes priority over her performing a probabilistic state transition to either \(n_1\) or \(n_2\) through independent discovery.
Formally, at the end of round t, if ant \(a_i\) is in \(n_0\), let \(TR^i_1(t+1), TR^i_2(t+1)\) be the event that ant \(a_i\) is recruited to \(n_1\) and \(n_2\) respectively during round \(t+1\). Let \(\tau ^i_1(t+1), \tau ^i_2(t+1)\) represent their respective conditional probabilities during round \(t+1\), i.e.,
Note that for any ant \(a_i\), the two events are mutually exclusive, and \(\tau ^i_1(t+1)+\tau ^i_2(t+1) \le 1\). The exact expressions for \(\tau ^i_1(t+1)\) and \(\tau ^i_2(t+1)\) are very complex and affect the time that ant \(a_i\) transitions out of \(n_0\), which is an important milestone time for the proofs in this paper. Fortunately, we manage to circumvent calculating the exact expressions of \(\tau ^i_1(t+1)\) and \(\tau ^i_2(t+1)\) by deriving a bound on this time using a coupling argument (Proposition 2). We found that this bound was sufficient for proving our main theorem.
With this notation, conditioning on an ant \(a_i\) being at state \(n_0\) at time t, the probability of her transitioning to \(n_1\) in the nest round, denoted by \(\alpha ^i_1(t)\) can be expressed as
where \(\alpha \) can be formally expressed as
. It is easy to see that \(\alpha ^i_1(t+1)\) sums up the probability of her getting recruited to \(n_1\) and the probability of independent discovery of \(n_1\) in the case that she does not get recruited to either \(n_1\) or \(n_2\). Similarly, we define \(\alpha ^i_2(t+1)\) as the probability of her transitioning to \(n_2\) during round \(t+1\), i.e.,
Correspondingly,
Transitions Between New Nests
When \(\boldsymbol{s}_i(t)= n_m\) for \(m\in \{1, 2\}\), at the beginning of round \(t+1\), with probability \((1-u_m)\), ant \(a_i\) chooses to search her environment and discover the new nest she is not currently at, i.e.,
with probability \(u_m\), ant \(a_i\) tries to recruit another ant from state \(n_0\) through a tandem run and comes back to \(n_m\), i.e.,
If there is no more ant left in \(n_0\) to recruit, the leader ant \(a_i\) simply returns to nest \(n_m\) without recruiting another ant. The recruiting probability \(u_m\) is determined by the quality of new nest \(n_m\) as
where the parameter \(\lambda > 0\) represents the noise level of individual decision making to evaluate the quality of a nest \(n_m\) for \(m \in \{1,2\}\). A larger \(\lambda \) means a less noisy decision rule, and thus a higher probability of recruitment to the superior site \(n_m\). Also note that \(u_1, u_2 \in [0.5, 1] \text { and } u_1 > u_2\).
Our choice of the sigmoid function is rooted in empirical evidence. The decision making mechanism for individual ant recruitment has been shown by a number of experimental and modeling studies to be both quality-dependent [15, 21, 24, 25] and threshold-based (individuals compare the perceived nest quality to a fixed threshold) [27, 28]. The sigmoid function we chose here is thus a common choice that incorporates both dependencies into the modeling of noisy individual decision making. Intuitively, when \(n_m\) has a quality higher than that of \(n_0\)’s, \(n_m\) is the better choice and it is beneficial for ants to recruit to it. When a nest \(n_m\) is strongly superior to \(n_0\), i.e., the quality difference surpasses a threshold, the probability of an individual ant recruiting to \(n_m\) should thus be very high (close to 1 in our model). The sigmoid function is a “smooth” representation of this threshold-based rule. On the other hand, when the quality difference is small, the probability of recruitment has stronger dependencies on the quality difference. This case is modeled by a near-linear segment in the sigmoid function.
Remark 1 (Non-markovian dynamics of an individual ant)
The state \(\boldsymbol{s}_i(t)\) of any individual ant \(a_i\) during round t has dependencies on 1) her own state in the previous round \(\boldsymbol{s}_i(t-1)\), and 2) the recruitment actions of other ants.
2.3 Dynamics of the Entire Colony
We now describe what happens in an arbitrary execution, or emigration. Throughout the paper, we use “an execution” and “an emigration” interchangeably, referring to an emigration event.
Let \(\boldsymbol{s}(t) = \left\{ \boldsymbol{s}_1(t), \cdots , \boldsymbol{s}_N(t) \right\} \) for \(t=0, 1, \cdots \) denote the random process of the entire colony state, represented by a vector of dimension N that stacks the states of individual ants in the colony. Although \(\boldsymbol{s}_i(t)\) for any i is not Markovian, it is easy to see that \(\boldsymbol{s}(t)\) is a Markov chain, since for any tandem leader in round t, the choice of a follower only depends on \(\boldsymbol{s}(t-1)\) and not on any history prior to round \(t-1\). An emigration starts from round 1, with \(\boldsymbol{s}_i=n_0\) for all \(i=1, \cdots , N\). During each round, each ant not in \(n_0\) performs one state transition in random order, followed by each ant in \(n_0\) performing one state transition in random order. At the beginning of a round t, each ant has her own state \(\boldsymbol{s}_i(t-1)\) and the colony has state \(\boldsymbol{s}(t-1)\). If at the beginning of round t she is in nest \(n_1\) or \(n_2\), respectively, the population at that nest at the beginning of round t is also available to \(a_i\). During a round t, each individual ant performs one state transition according to the individual models in Sect. 2.2, which results in a transition of the colony state as well during this round. At the end of round t, each ant has a new state \(\boldsymbol{s}_i(t)\) and the colony has state \(\boldsymbol{s}(t)\). All ants then enter the next round \(t+1\) with their new states.
3 The Consensus Problem
Here we define what it means for an emigration to reach consensus. We say that an emigration has reached \(\varDelta \)-consensus (where \(\varDelta \in [0,\frac{1}{2}]\)) if there exists \(\widetilde{t}\) such that for all \(t\ge \widetilde{t}\) and a nest \(m \in \{1,2\}\), the proportion of the population at nest \(n_m\) at time t is greater than or equal to \(1-\varDelta \), i.e., \(x_m(t) \ge (1-\varDelta )\).
The metric to evaluate a model’s performance is the consensus probability C, which is the probability that an emigration reaches consensus as defined above.
Remark 2
Note that \(\varDelta \) represents the proportion of ants that can be exempted from the consensus requirement. We can see that the smaller \(\varDelta \) is (lowest value is 0), the larger \((1-\varDelta )N\) is, and hence the more ants are required for an emigration to reach consensus. In other words, the smaller \(\varDelta \) is, the more “strict” the consensus metric is and the more challenging it is for an emigration to reach consensus.
4 Failure of Consensus in Two-Nest Environments
In this section, we explore colony emigration behavior only with individual transition rules and tandem runs defined above (i.e., without quorum sensing). Equivalently, we consider the case where the quorum size is N, so that the quorum sensing mechanism never has any effect. We show an upper bound on the consensus probability C for a given \(\varDelta \) and colony size N. This upper bound decreases to 0 exponentially fast as \(N\rightarrow \infty \).
Next we introduce two quantities, denoted by H and \(\pi ^*\), that will be used in the statement of our main result. It is easy to see from Eq. (4) and (5) that if an ant \(a_i\) jumps out of the home nest \(n_0\) at some time, then from that time onward, the state transition of \(a_i\) becomes Markovian and is governed by the following transition matrix
The transition in H is also illustrated in Fig. 4. It can also be seen (which we will formally show later) that the state of each ant has an identical limiting distribution, denoted by \(\pi ^* \triangleq \frac{1}{2-u_1-u_2}\left[ 1-u_2, 1-u_1 \right] \in {\mathbb {R}}^2\), with support on \(\{n_1, n_2\}\) only.
Theorem 1
For any \(\varDelta \in [0, 1-\pi ^*(n_1)]\), let \(\epsilon _0 = \frac{1-\pi ^*(n_1)-\varDelta }{2} > 0\). Then it holds that
for any \(t > \left( \frac{1}{\ln (1-2\alpha )} + \frac{1}{\ln (1-R(H))} \right) \ln \frac{\epsilon _0}{2}\), where \(R(H) = 2 -u_1-u_2\) is Dobrushin’s coefficient of ergodicity ([11, Chapter 6.2]) of H.
Remark 3
Theorem 1 is stated for \(n_1\). A similar result holds for \(n_2\). Theorem 1 says that for any t greater than \(\left( \frac{1}{\ln \beta } + \frac{1}{\ln (1-R(H))} \right) \ln \frac{\epsilon _0}{2}\), the probability of \(x_1(t)\) reaching \((1-\varDelta )\) is upper bounded by \(2\exp \left( -\frac{\epsilon _0^2N}{2} \right) \). Thus, the total consensus probability C for the given \(\varDelta \) is upper bounded by \(4\exp \left( -\frac{\epsilon _0^2N}{2} \right) \), which decreases to 0 exponentially fast as N increases. It is worth noting that real ant colonies often need \(\varDelta \) to be very small or even zero for survival. From the theorem expression, we can see that the smaller \(\varDelta \) is, i.e., the more stringent the consensus, the lower is the upper bound of the consensus probability. Therefore, Theorem 1 implies that extra mechanisms, such as the quorum rule are necessary to help the emigration reach consensus.
Later in Sect. 5, we also show that the proofs in this section and related results can easily extend to environments with multiple nests.

Flowchart of the proofs.
4.1 Analysis of Main Result
Despite the fact that the dynamics of the entire ant colony is a Markov chain, analyzing this Markov chain is highly non-trivial because the state is quite involved and the state space is huge – it contains all the possible partitions of ants into three groups, with each group representing one nest as the state of an individual ant. In this section we analytically show that despite the fact that the emigration behaviors of individual ants are interactive, the dynamics of any individual ant are independent of other ants shortly after she leaves the original home nests either through discovery or through recruitment. Moreover, we show that this independence manifests itself in a non-trivial way after a few rounds – suggesting that a large portion of ants quickly rely only on individual intelligence. Then we show that this independence is harmful to realizing social cohesion.
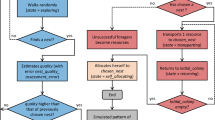
Several intermediate results are derived in proving Theorem 1. The connections of the supporting lemmas and corollaries with respect to Theorem 1 are shown in Fig. 3. Please note that due to space constraints, we show the proof details of only Theorem 1 in this paper. Those of all other intermediate results can be found in [33].
Definition 1
For each \(i \in [N]\), define random variable \(T_i^1 \triangleq \inf \{t: \, \boldsymbol{s}_i(t)\not =n_0\}\) as the first round at the beginning of which ant \(a_i\) has transitioned out of the \(n_0\) state in any arbitrary execution of the emigration.
Remark 4
It can be shown that \(T_i^1\) is finite with probability 1 [33]. It follows immediately from Definition 1 that \( \mathbb {P}\left\{ \boldsymbol{s}_i(t) = n_0 \mid t \ge T_i^1 \right\} = 0\) for any ant \(a_i\).
It turns out that ant \(a_i\)’s state transitions become independent of other ants after \(T_i^1\), the time that \(a_i\) leaves \(n_0\), formally stated in the following proposition.
Proposition 1
For every \(i,j \in [N], i\not =j\) and every \(t > T_i^1\), the state transitions of ant \(a_i\) are independent from \(a_j\), i.e.,
where \(s_1, s_2, s_1^\prime \in {\mathcal {S}}\) and \(s_1^\prime \not = n_0\).
The next proposition is devoted to showing that after a few rounds, many ants have left the home nest \(n_0\). Consider N random indicator variables \({\mathbbm {1}{\left\{ {T_i^1> t}\right\} }}\) for any t, each variable taking values in the \(\{0, 1\}\). Using stochastic dominance and Hoeffding’s inequality [13], we show a high probability upper bound on the number of ants still in \(n_0\) at round t. Here stochastic dominance is used to tackle the challenges caused by the dependency among the N indicator random variables.
Proposition 2
Let \(\beta \triangleq 1-2\alpha \). For \(t\ge 1\) and any number \(d \in [0,1]\), it holds that
i.e., with a probability of at least \((1-\exp \left( -2Nd^2 \right) )\), the number of ants staying at home nest beyond time t is at most \(N\left( \beta ^t+d \right) \).
Corollary 1
For any given \(\epsilon \in (0, 1)\), for any \(t\ge \log _{\beta }(\frac{\epsilon }{2})\), it holds that
In other words, with a probability of at least \((1-\exp \left( -N\epsilon ^2/2 \right) )\), at most \(\epsilon N\) ants remain in the home nest \(n_0\) after round \( \log _{\beta }(\frac{\epsilon }{2})\).

State transition diagram for individual ants after they leave \(n_0\), before/without quorum attainment.
Next, we show that every ant \(a_i\) has an identical limiting distribution. Towards this, we first show that every ant \(a_i\) that has transitioned out of \(n_0\) has the same limiting distribution. Furthermore, we show that all ants eventually transition out of \(n_0\) and thus all ants share the same limiting distribution. The proof of Lemma 1 uses the quantity Q(t), defined as
which is a random variable representing the set of ants that have transitioned out of \(n_0\) by the end of round t, in an arbitrary emigration. Q(t) is thus a function of an execution. It is easy to see that w.r.t. this emigration, \(Q(t-1)\subseteq Q(t)\) for any \(t\ge 1\).
Lemma 1
For each \(a_i\), its limiting distribution, denoted by \(\pi _i\), is well-defined, and can be expressed as
For ease of exposition, we define \(\pi ^* = \pi _i\). From Lemma 1 it can be seen that the probability ratio \(\frac{\pi ^*(n_1)}{\pi ^*(n_2)} = \exp \left( \lambda (q_1-q_2) \right) \) is very sensitive to the nest quality gap \((q_1-q_2)\) and \(\lambda \).
It turns out that for t large enough, any ant that has transitioned out of \(n_0\) has state distributions “close” to the stationary distribution \(\pi ^*\), formally stated next.
Lemma 2
For any ant \(a_i\), let \(\pi _{i,t}\) denote the probability distribution of her state over the possible states depicted in Fig. 4 at time \(t \ge T_i^1\). Then for any number of rounds \(\ell >0\), it holds that
Using Lemma 2, the following corollary immediately follows:
Corollary 2
Fix any \(\delta \in (0, 1)\). For any ant \(a_i\) and \(t > T_i^1 +\ell \), where \(\ell \triangleq \log _{\left( 1-R(H) \right) } \frac{\delta }{2}\), it holds that
Combined with Corollary 1, we are now ready to prove Theorem 1.
Proof of Theorem 1
We first give the intuition and a proof sketch to show an upper bound on the probability of the population at \(n_1\) being higher than a certain number \(C_0\), for t large enough.
We break down the problem into two cases. In the first case, by a certain milestone-round \(k_1\), the number of ants that have transitioned out of \(n_0\) is low. In the second case, that number is high. Now, by applying concentration bounds, we show that the first case has a low probability. We thus subsequently focus on analyzing the second case which has a high probability. From Corollary 2 we know that after a certain number \(k_2\) of rounds, most of the ants that have left \(n_0\) will have distributions that are very close to the limiting distribution \(\pi ^*\). Thus, at any round \(t \ge k_1+k_2\), with high probability, the proportion of ants in \(n_1\) is also close to \(\pi ^*(n_1)\) among ants that have left \(n_0\) (at most N ants). In other words, after \(k_1+k_2\) rounds the probability of \(n_1\)’s population being much higher than \(\pi ^*(n_1)N\) should be quite low. Summing up the bounds for the first and second cases gives us an overall upper bound on this probability, proving the theorem.
For ease of exposition, let \(B_i(t) = {\mathbbm {1}{\left\{ {\boldsymbol{s}_i(t) = n_1}\right\} }}\) for each \(i\in [N]\) and \(t\ge 0\). Let \(C_0\) be an arbitrary positive number, \(C_0 \in [0, N]\). Let \(C_1 = (1-\epsilon _0)N\). Recall that \(\beta = 1-2\alpha \).
We bound the two terms in the RHS of Eq. (10) separately.
Bounding the 1st term: For any \(t\ge \log _{\beta } \frac{\epsilon _0}{2}\), we have
where the last inequality follows from Corollary 1.
Bounding the 2nd Term: Note that
It is easy to see that
In addition, we have
Conditioning on \(\left| Q(\log _{\beta } \frac{\epsilon _0}{2})) \right| \ge (1-\epsilon _0)N\), from Corollary 2, we know that for each \(a_i\in Q(\log _{\beta } \frac{\epsilon _0}{2})\), for any \(t > \log _{\beta } \frac{\epsilon _0}{2} + \ell \), where \(\ell = \log _{\left( 1-R(H) \right) } \frac{\epsilon _0}{2}\), it holds that \(\pi _{i,t}(n_1) \le \pi ^*(n_1) +\epsilon _0\). Hence we get
Thus,
Combining Eq. (11) and (12), we conclude that
Combining the probability bounds on the first and second terms of Thoerem 1, we have
proving Theorem 1.
5 Extension: Failure of Consensus in More-Nest Environments
Both the results on asymptotic independence and its negative impact presented in Sect. 4 can be extended to the general k-new-nest environments where \(k>2\). On a high level, the necessary additions to the individual transition model (Fig. 2) are: 1) a new state for each new nest, each similar to the \(n_1\) and \(n_2\), 2) all new nests can exchange ants with each other, and 3) all new nests can receive ants from \(n_0\) through recruitment or discovery. The model for timing, environment, and execution of the whole colony remain the same as the two-nest case, where \(n_1\) has the highest quality. After adjusting quantities H and \(\pi ^*\), one can derive results similar to Theorem 1: without quorum sensing, the probability of consensus can be arbitrarily low. We detail these changes below in this section.

State transition diagram for ant \(a_i\) during round \(t+1\) in a k-nest environment before/without quorum attainment. Probabilities \(\alpha _l^i(t+1), l \in [1,k]\) are composite functions each including the probabilities of an ant taking different paths (independent discovery or tandem running) to transition out of \(n_0\) into \(n_l\). Compared to Fig. 2, this figure shows the addition of one more new nest \(n_3\); any more new nests can be added in the same way.
Figure 5 shows the transition diagram for an individual ant before/without her seeing a quorum at any nest, and Eq. (13)–(16) define transition probabilities among the four states. Similar to the two-nest case, \( \mathbb {P}\left\{ TR_l^i(t+1) \right\} = \tau _l^i(t+1)\) for each \(l \in [1,k]\) is defined as the probability of the event that ant \(a_i\) transitions from \(n_0\) to \(n_l\) during round \(t+1\) by following a tandem run. Figure 5 displays only the addition of a third new nest, \(n_3\), and any more new nest can be added in the same way. The addition of \(n_3\) requires that during round t, an ant at \(n_0\) transitions to \(n_3\) with probability \(\alpha _3(t)\); an ant at \(n_3\) stays in \(n_3\) with probability \(u_3\); and an ant at a new nest l transitions to any other new nest \(m \not = l\) with transition probability \(\frac{1-u_l}{k-1}\).
where
The two quantities used in the main theorem for a k-nest environment, H and \(\pi ^*\), are also different, as shown below.
-
H, a \(k\times k\) transition matrix of an arbitrary ant \(a_i\)’s state \(\boldsymbol{s}_i\) after she transitions out of \(n_0\), as specified in Eq. (17).
-
\(\pi ^* \in {\mathbb {R}}^k\), a vector representing the limiting distribution of an arbitrary ant \(a_i\) (Eq. (18)). The l-th element is the limiting distribution of state \(n_l\), for \(l \in [1,k]\).
Solving the equation system \(\pi _i = \pi _iH\), we also obtain that
Main Theorem for k-Nests
Theorem 2
For any \(\varDelta \in [0, 1-\pi ^*(n_1)]\), let \(\epsilon _0 = \frac{1-\pi ^*(n_1)-\varDelta }{2} > 0\). Then it holds that
for any \(t > \left( \frac{1}{\ln (1-k\alpha )} + \frac{1}{\ln (1-R(H))} \right) \ln \frac{\epsilon _0}{2}\), where R(H) is Dobrushin’s coefficient of ergodicity ([11, Chapter 6.2]) of H.
Remark 5
Theorem 2 is stated for \(n_1\). A similar result holds for \(n_l\) for \(l > 1\). Like in the two-nest case, Theorem 2 again implies that extra mechanisms, such as the quorum rule are necessary to help the emigration reach consensus.

State transition diagram for individual ants committed to \(n_1\) and \(n_2\), on the left and right, respectively.
6 Consensus with Quorum Sensing in Two-Nest Environments
An important work in progress is analyzing the probability of consensus when the quorum rule is in effect. In this section, we show as a work in progress the addition of the quorum sensing mechanism to our model, and our current results on the quorum sizes that are sufficient for consensus of average emigrations in two-nest environments.

State transition diagram for ant \(a_i\) during round \(t+1\) with the quorum sensing mechanism. She first starts transitioning according to the left part of the figure, identical to Fig. 2. Then once she sees a quorum at either \(n_1\) and \(n_2\) (but not both), she commits to that nest and can only stay in that nest, as shown on the right part of the figure, identical to Fig. 6.
Note that the dynamics shown in Fig. 2 are also accurate here before an ant \(a_i\) sees a quorum for the first time at either nest. Thus, she starts her transitions according to Fig. 2 before seeing any quorum. The evaluation of whether \(n_m\) has reached quorum happens whenever \(a_i\) is in \(n_m\) at the beginning of a round t. Before she performs any transitions during round t, she compares the nest population to a quorum size, if \(a_i\) has not yet seen a quorum at \(n_m\) (or at any other nest). Once she detects that the population is at least as high as the quorum size, she becomes “committed” to \(n_m\). After that, she no longer monitors the nest’s population. We model an ant’s commitment by disallowing her to transition out of \(n_m\). This means she has to perform a transport action and stay in the \(n_m\) state at any round after \(n_m\)’s quorum attainment. As a result, once a nest reaches the quorum, it never drops out of the quorum and every ant that transitions to that nest gets “stuck” in that nest. We thus model a “committed” ant with a separate Markov chain that essentially only has one possible state, as shown in Fig. 6 and Eq. (19)–(22). For a committed ant \(a_i\), let \(n_m\) be the nest that she is committed to where \(m \in \{1,2\}\). Then the other new nest she is not committed to is \(n_{3-m}\).
Individual Model With Quorums. We show the full model in Fig. 7. The addition of transporting as a possible recruitment method thus has two impacts in the full model:
-
An ant \(a_i\) in \(n_0\) can get recruited by being transported to either \(n_1\) or \(n_2\), in addition to following a tandem run.
-
an ant \(a_i\) in either state \(n_1\) or \(n_2\) choosing to stay in the same state tries to recruit another ant from state \(n_0\) through a tandem run if the quorum is not reached (Fig. 2, Eq. (5)), or through a transport otherwise (Fig. 6, Eq. (19)). Whether the recruitment is successful or not still has no effect on \(a_i\)’s own state transitions during this round. Otherwise, if she does not recruit, she searches her environment and discovers the new nest she is not currently at (Eq. (4)).
It still holds that during any given round t, if an ant \(a_i\) at \(n_0\) does not get recruited, her transitions are Markovian and independent (Fig. 2, Fig. 6). The whole colony dynamics are the same as shown in Sect. 2.3 and the whole colony state retains its Markovian properties.

3D plots demonstrating quorum sizes that are sufficient for consensus, when \(\alpha \le \frac{1}{3}\). Views from two angles.
Current Work: In our work in progress, through theoretical analysis and simulation work, we are striving to derive quorum sizes that are sufficient for consensus. Our preliminary results in Fig. 8 show such quorum sizes for two-nest environments. In these emigrations, we enforce that \(\varDelta =0\) to model the most challenging requirement of consensus. In Fig. 8, for the full ranges of \(u_1\) and \(u_2\) in the frequent case that \(\alpha < \frac{1}{3}\), we visualize the quorum sizes (QS) in the range [0.25, 0.5] that are expected to lead to consensus. The desirable values of the quorum size show general consistency with experimental findings of the observed quorum size employed by Temnothorax ant colonies [8, 22]. However, we are still working on deriving the mathematical expressions for quorum sizes that are sufficient for consensus, as well as on extending these results to k-nest environments (\(k>2\)). We plan to show all related details of this effort in a follow-up manuscript.
7 Discussion and Future Work
In this paper, we used analytical tools to show that without quorum sensing, the collective nest site selection process by Temnothorax ants has a limited probability to reach consensus. And this probability can be arbitrarily low for a colony size arbitrarily large. Conversely, we obtain a high probability bound for failure of consensus. Without quorum sensing, the only form of recruitment, tandem runs, does speed up the emigration process, but our results show that emigrations would still have a high probability of splitting among multiple new sites, imposing significant risks to the colony’s survival. We first analyze a model of a two-new-nest environment, and then extend our results to environments with more nests. Our results provide insights into the importance of extra mechanisms, such as the quorum sensing mechanism, for emigrations to reach consensus in an unpredictable environment with multiple nests.
In this paper we also provided a preview of an important work in progress investigating how different quorum sizes influence emigration outcomes if quorum sensing is involved, in two-nest environments. Further extensions in this direction are to apply similar analytical methods to the general environment with the addition of quorum sensing to gain insights on how the number of nests and their qualities might influence the desirable values for the quorum size, with the goal to avoid splits, or to ensure consensus, or with an objective involving a specific degree or probability of consensus.
Additionally, another future work direction is to make our model more bio-plausible. Specifically, our model does not consider the very small probability that committed ants “drop out” of the nest they are committed to, and go back to searching. Adding this into the model could make it biologically more realistic.
Finally, one more way to strengthen our theoretical results is by adding a time bound metric to our consensus problem. Our current consensus metric, the consensus probability C, only requires that at least \((1-\varDelta )N\) ants keep staying at either \(n_1\) or \(n_2\) after a finite number of rounds. By adding a time bound metric as well, we would be able to better characterize the consensus probability (even if lower than a given C) of an emigration by a certain time t.
References
Doering, G.N., Pratt, S.C.: Queen location and nest site preference influence colony reunification by the ant temnothorax rugatulus. Insectes Soc. 63(4), 585–591 (2016). https://doi.org/10.1007/s00040-016-0503-1
Doering, G.N., Pratt, S.C.: Symmetry breaking and pivotal individuals during the reunification of ant colonies. J. Exp. Biol. 222(5), jeb194019 (2019)
Doran, C., Newham, Z.F., Phillips, B.B., Franks, N.R.: Commitment time depends on both current and target nest value in temnothorax albipennis ant colonies. Behav. Ecol. Sociobiol. 69(7), 1183–1190 (2015). https://doi.org/10.1007/s00265-015-1932-y
Dornhaus, A., Holley, J.A., Pook, V.G., Worswick, G., Franks, N.R.: Why do not all workers work? Colony size and workload during emigrations in the ant temnothorax albipennis. Behav. Ecol. Sociobiol. 63(1), 43–51 (2008)
Franks, N.R., Dechaume-Moncharmont, F.X., Hanmore, E., Reynolds, J.K.: Speed versus accuracy in decision-making ants: expediting politics and policy implementation. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 364(1518), 845–852 (2009)
Franks, N.R., Dornhaus, A., Fitzsimmons, J.P., Stevens, M.: Speed versus accuracy in collective decision making. Proc. R. Soc. Lond. Ser. B: Biol. Sci. 270(1532), 2457–2463 (2003)
Franks, N.R., Mallon, E.B., Bray, H.E., Hamilton, M.J., Mischler, T.C.: Strategies for choosing between alternatives with different attributes: exemplified by house-hunting ants. Anim. Behav. 65(1), 215–223 (2003)
Franks, N.R., Stuttard, J.P., Doran, C., Esposito, J.C., Master, M.C., Sendova-Franks, A.B., Masuda, N., Britton, N.F.: How ants use quorum sensing to estimate the average quality of a fluctuating resource. Sci. Rep. 5(1), 11890 (2015)
Franks, N., et al.: Speed-cohesion trade-offs in collective decision making in ants and the concept of precision in animal behaviour. Anim. Behav. 85(6), 1233–1244 (2013)
Gordon, D.M.: The ecology of collective behavior in ants. Ann. Rev. Entomol. 64(1), 35–50 (2019). pMID: 30256667
Hajek, B.: Random Processes for Engineers. Cambridge University Press, Cambridge (2015)
Healey, C.I.M., Pratt, S.C.: The effect of prior experience on nest site evaluation by the ant temnothorax curvispinosus. Anim. Behav. 76, 893–899 (2008)
Hoeffding, W.: Probability inequalities for sums of bounded random variables. J. Am. Stat. Assoc. 58(301), 13–30 (1963)
Johnstone, R.A., Dall, S.R.X., Franks, N.R., Pratt, S.C., Mallon, E.B., Britton, N.F., Sumpter, D.J.T.: Information flow, opinion polling and collective intelligence in house-hunting social insects. Philos. Trans. R. Soc. Lond. Ser. B: Biol. Sci. 357(1427), 1567–1583 (2002)
Mallon, E., Pratt, S., Franks, N.: Individual and collective decision-making during nest site selection by the ant leptothorax albipennis. Behav. Ecol. Sociobiol. 50(4), 352–359 (2001). https://doi.org/10.1007/s002650100377
Marshall, J.A.R., Bogacz, R., Dornhaus, A., Planqué, R., Kovacs, T., Franks, N.R.: On optimal decision-making in brains and social insect colonies. J. R. Soc. Interface 6(40), 1065–1074 (2009)
Marshall, J.A., Dornhaus, A., Franks, N.R., Kovacs, T.: Noise, cost and speed-accuracy trade-offs: decision-making in a decentralized system. J. R. Soc. Interface 3(7), 243–254 (2006)
McGlynn, T.P.: The ecology of nest movement in social insects. Ann. Rev. Entomol. 57(1), 291–308 (2012). pMID: 21910641
Moglich, M.H.J.: Social organization of nest emigration in leptothorax (hym., form.) (1978)
Planqué, R., Dornhaus, A., Franks, N.R., Kovacs, T., Marshall, J.A.R.: Weighting waiting in collective decision-making. Behav. Ecol. Sociobiol. 61(3), 347–356 (2007). https://doi.org/10.1007/s00265-006-0263-4
Pratt, S.C.: Behavioral mechanisms of collective nest-site choice by the ant temnothorax curvispinosus. Insectes Soc. 52(4), 383–392 (2005). https://doi.org/10.1007/s00040-005-0823-z
Pratt, S., Mallon, E., Sumpter, D., et al.: Quorum sensing, recruitment, and collective decision-making during colony emigration by the ant leptothorax albipennis. Behav. Ecol. Sociobiol. 52(2), 117–127 (2002). https://doi.org/10.1007/s00265-002-0487-x
Pratt, S.C.: Quorum sensing by encounter rates in the ant Temnothorax albipennis. Behav. Ecol. 16(2), 488–496 (2005)
Pratt, S.C., Sumpter, D.J.T.: A tunable algorithm for collective decision-making. Proc. Natl. Acad. Sci. 103(43), 15906–15910 (2006)
Pratt, S.C., Sumpter, D.J., Mallon, E.B., Franks, N.R.: An agent-based model of collective nest choice by the ant temnothorax albipennis. Anim. Behav. 70(5), 1023–1036 (2005)
Richardson, T.O., Sleeman, P.A., Mcnamara, J.M., Houston, A.I., Franks, N.R.: Teaching with evaluation in ants. Curr. Biol. 17(17), 1520–1526 (2007)
Robinson, E.J.H., Franks, N.R., Ellis, S., Okuda, S., Marshall, J.A.R.: A simple threshold rule is sufficient to explain sophisticated collective decision-making. PLOS ONE 6(5), 1–11 (2011)
Robinson, E.J., Smith, F.D., Sullivan, K.M., Franks, N.R.: Do ants make direct comparisons? Proc. R. Soc. B: Biol. Sci. 276(1667), 2635–2641 (2009)
Sasaki, T., Colling, B., Sonnenschein, A., Boggess, M.M., Pratt, S.C.: Flexibility of collective decision making during house hunting in temnothorax ants. Behav. Ecol. Sociobiol. 69(5), 707–714 (2015). https://doi.org/10.1007/s00265-015-1882-4
Stroeymeyt, N., Giurfa, M., Franks, N.R.: Improving decision speed, accuracy and group cohesion through early information gathering in house-hunting ants. PLOS ONE 5(9), 1–10 (2010)
Sumpter, D.J., Pratt, S.C.: Quorum responses and consensus decision making. Philos. Trans. R. Soc. B: Biol. Sci. 364(1518), 743–753 (2009)
Visscher, P.K.: Group decision making in nest-site selection among social insects. Ann. Rev. Entomol. 52(1), 255–275 (2007). pMID: 16968203
Zhao, J., Su, L., Lynch, N.: (2021). https://github.com/snowbabyjia/QuorumSensingConsensus
Acknowledgements
J. Zhao and N. Lynch are supported by NSF Awards CCF-2003830, CCF-1461559 and CCF-0939370.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2021 Springer Nature Switzerland AG
About this paper

Cite this paper
Zhao, J., Su, L., Lynch, N. (2021). Lack of Quorum Sensing Leads to Failure of Consensus in Temnothorax Ant Emigration. In: Johnen, C., Schiller, E.M., Schmid, S. (eds) Stabilization, Safety, and Security of Distributed Systems. SSS 2021. Lecture Notes in Computer Science(), vol 13046. Springer, Cham. https://doi.org/10.1007/978-3-030-91081-5_14
Download citation
DOI: https://doi.org/10.1007/978-3-030-91081-5_14
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-91080-8
Online ISBN: 978-3-030-91081-5
eBook Packages: Computer ScienceComputer Science (R0)