
Abstract
The assemblage of species found at a location derives from the regional pool of species, governed by the ability of each species to colonize new habitat, and the matching of individual species to available resources, habitat, and one another. Regional species richness displays intriguing geographical patterns, best described for freshwater fishes. The number of species is inversely related to latitude, increases with drainage area and discharge of the river system, and differs among regions at the same latitude in both number of species and the traits of species present. System productivity, habitat diversity, and historical events are amongst the explanations for these patterns. At local scales, the number and relative abundance of species, their traits and functional roles, and energy pathways through food webs are useful descriptors of community structure. The processes that influence the assembly and maintenance of local communities include niche-based models that focus on the interplay between biotic interactions and abiotic forces, the habitat template model based on the association of individual species with habitat features, and disturbance models that emphasize the interplay between species interactions and variation in environmental factors that periodically reduce the abundance of some or all species in the community.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

The forces that shape community structure are those that determine which species and how many species occur together, which are common and which are rare, and the interactions amongst them. The number of coexisting species in ecological communities is a consequence of processes operating at both local and regional scales (Rosenzweig 1995; Gaston 2000). Regional diversity is determined by species formation and loss over earth history, and opportunities for dispersal. Local diversity is determined by species adaptations to the abiotic and biotic factors encountered at a site, and by each species’ dispersal ability and life history characteristics. Thus, the composition of an assemblage of species at a location is the product both of large-scale spatial patterns and the forces responsible for them, and local conditions that influence which species are best adapted to that environment, or best able to colonize it.
Community structure refers to the organization of a biological community based on numbers of individuals within different taxonomic groups and functional roles, and the underlying processes that maintain that organization. Explanations for patterns in species diversity and community structure frequently invoke niche-based models (MacArthur 1972; Chase and Leibold 2003), in which the presence and abundance of individual species is a reflection of their fit to habitat conditions and success in interspecific interactions, the subjects of Chaps. 5 and 10 respectively. In relatively stable or moderate environments, biological interactions are considered to be particularly influential in the assembly and maintenance of communities. For communities to exhibit predictable structure requires that their assembly is the outcome of non-random processes that result in repeatable patterns. This leads us to expect that the same species, in roughly the same abundances, will be found in the same locale as long as environmental conditions do not change greatly, and that similar communities should occur wherever environmental circumstances are comparable. In addition, many ecosystems experience periodic disturbances, and streams are no exception. Environmental disturbances such as floods and droughts, when sufficiently extreme or frequent, are likely to prevent biotic interactions from acting with the strength and regularity required to result in consistent community patterns. Very harsh environments or frequent disturbances may severely restrict the number of species that can survive those conditions and thus reduce diversity, whereas a moderate level of disturbance may enhance diversity by counteracting the tendency of a few superior species to become dominant.
The countering view to niche models asserts that assemblages are an unstructured sample of whichever species are able to survive and reproduce under local environmental conditions, changing as conditions change, and by chance. A more formalized version, the neutral model of Hubbell (2001, 2005), considers all species of a particular trophic level to be essentially interchangeable, and so random replacement following stochastic colonization and extinction determines the momentary composition of short-lived assemblages. Under this model, species are ecologically equivalent and substitutable, to be replaced from a regional species pool whenever a chance local extinction depletes site diversity. The recolonization of a lost population requires dispersal, and so distance, life-history traits, and other factors, such as terrain, can determine whether dispersal limits the opportunities for a particular species to re-establish.
Studies of local assemblages often assume that communities are determined solely by environmental conditions and species interactions at the local scale, without regard for such larger scale processes as dispersal, speciation, and historical biogeography. However, regional species pools and factors that influence dispersal at large spatial scales influence local diversity and assemblage structure by controlling the pool of species available to colonize a location (Ricklefs and Schluter 1993; Rosenzweig 1995). Processes occurring at large spatial scales, such as the size and composition of the regional species pool, opportunities for dispersal, and climate can influence local communities (Leibold and Chase 2017). This is especially true in local communities if species interactions are not the primary force governing community structure, including systems with frequent disturbance events such as floods and droughts, or when communities are dominated by rare species with limited dispersal capacities (Cornell and Harrison 2014).
Species sorting describes how regional diversity affects local diversity through the action of environmental factors acting at progressively smaller scales, often visualized as a hierarchical series of filters (Poff 1997). It is also important to remember that local communities are not isolated from one another. They are connected via dispersal to other local communities, forming a network or metacommunity, that in turn is influenced by spatial heterogeneity and environmental factors that are operating at different scales (Heino and Mykrä 2008; Leibold and Chase 2017). Consideration of regional diversity also reminds us that the long-term persistence of a species usually does not depend solely upon its survival in any one local community. Separate populations of a species may exhibit different trends in different locales, with the consequence that dispersal permits a long-term regularity on a larger scale that is not apparent by detailed investigation on a finer scale. Such a perspective lessens the need for equilibrium-enhancing interactions because regional processes of immigration and emigration may contribute some of the buffering against extinction that otherwise must be attributed to biotic factors.
11.1 Global and Regional Patterns in Species Diversity
Freshwater biodiversity varies at all scales, including among regions of the world, along latitudinal and environmental gradients, among ecoregions, and across habitats. As discussed in Chapter 1, freshwater habitats support more than 125,000 species of freshwater animals, comprising nearly 10% of all known species and approximately one-third of vertebrate species (Balian et al. 2008; Strayer and Dudgeon 2010). Freshwater fishes are better known than invertebrates and microorganisms, but are still far from fully catalogued, especially in tropical latitudes where the diversity of freshwater fishes is known to be much greater than at temperate latitudes (Lévêque et al. 2008). A comprehensive assessment of freshwater biodiversity is presented in a series of articles edited by Balian et al. (2008). These accounts are extremely useful for cataloguing known diversity in all major freshwater groups, providing much information on biogeographic patterns and the current status of many species.
Among freshwater organisms, fishes have been most intensively scrutinized from the perspective of identifying and attempting to explain global and regional patterns, because species-level distributions are well known for a large fraction of the fauna. The global distribution of freshwater fish species richness (number of species) is shown in Fig. 11.1 (Oberdorff et al. 2011), and interesting patterns are immediately apparent. More species occur at low (tropical) than higher latitudes. Eastern North America has more fish species than western North America or western Europe. Some regions are notably depauperate. Three main hypotheses have been invoked in the study of global patterns: species-area, species-energy, and historical events. Studies find that species richness increases with the area under consideration, with available energy and ecosystem productivity, and as a consequence of historical factors that influence speciation and dispersal. Variation in regional species richness itself is difficult to fully explain, but events in earth history that influence species formation, extinction, and differences in the ability of individual species to disperse clearly influence the regional species pool, and contemporary patterns in abundance and diversity in turn are influenced by overall productivity, habitat diversity, and biological interactions (Rosenzweig 1995; Cornell and Harrison 2014). In an analysis of freshwater fish species richness for 132 West European and North American rivers, Griffiths (2006) concluded that contemporary ecological factors provided the strongest statistical explanation of the variation in freshwater fish species richness for both continents. Historical factors, while also significant, had less explanatory power. We shall briefly consider each explanation, and because studies often infer that each plays a role and researchers usually try to deduce their relative weights, most examples provide evidence for multiple causal mechanisms.

(Reproduced from Oberdorff et al. 2011)
(a) Freshwater fish species richness for worldwide drainage basins and (b) contribution of area-related (habitat size and diversity), climate-related (climate/energy) and historical variables (climate history and geographic isolation) to explain variation in species richness
11.1.1 Species-Area Relationships
The species-area hypothesis relates to a well-established relationship between the number of species and the area of study, first established using islands of differing sizes (MacArthur and Wilson 1963, 1967). On a log-log scale, species richness (S) increases with area (A) by an exponent, z, which is empirically determined and can be compared among taxa and locations studied. The relationship typically is log-linear, according to the equation:
where S = species richness, A = area of habitat, and c and z are parameters determined from the data. The slope parameter z quantifies the rate of increase in number of species with area surveyed, and frequently falls between 0.2 and 0.4.
A number of studies have established that species richness increases log-linearly with either drainage basin area or river discharge (Fig. 11.2), including for the mollusks of North America (Sepkoski and Rex 1974), and fishes of West Africa (Hugueny 1989), South America (De Mérona et al. 2012), and Europe and North America (Oberdorff et al. 1997). Species richness is highly correlated with basin area for rivers worldwide, and stronger statistical relationships are observed when analyzed separately by continent (Amarasinghe and Welcomme 2002). The exponent of the species-area relationship always is greater in tropical than temperate regions (0.25 for Europe, 0.26 for Asia, 0.49 for Africa, 0.51 for South America), demonstrating a trend toward a more rapid increase in species richness with increasing river size at low latitudes.

On a global scale, total drainage area of the watershed and river discharge as a proxy of river size can explain a large proportion of the variation in freshwater fish species richness (Oberdorff et al. 1995). The underlying basis is still debated, but area-determined changes in rates of species formation and loss, and in habitat complexity, are possible explanations. Larger rivers may harbor greater diversity of habitat and food resources because of their size, and thus sustain a greater number of coexisting species. More habitat area may also permit larger population numbers within species, thereby reducing the probability of species extinction. Rates of speciation may be higher in larger systems due to corresponding increases in habitat heterogeneity and the occurrence of geographical barriers, such as waterfalls (Oberdorff et al. 2011). For example, higher speciation in fishes was positively related to the level of natural fragmentation caused by waterfalls in sub-drainages of the Orinoco Basin (Dias et al. 2013). Research also suggests that many of the species found in larger rivers appear to be unable to live in small streams. Longitudinal studies of riverine fishes find that addition, rather than replacement, of species characterizes the change in community assemblages along a river continuum, and a preponderance of larger species is observed downstream (e.g., Horwitz 1978; Schlosser 1987).
11.1.2 Latitudinal Diversity Gradients
An increase in numbers of species as one proceeds from high latitudes towards the tropics is one of the most general of geographic patterns of species richness (Hillebrand 2004; Kinlock et al. 2018). Several hypothesis based on the stability of tropical climates over time, their areal extent and higher biological productivity, and historic rates of speciation have been proposed to explain this pattern (Mittelbach et al. 2007; Heino 2011).
Although stronger in terrestrial than in freshwater environments, this relationship unquestionably holds for the fishes of running waters. Despite the incomplete state of taxonomic knowledge, more than 4,000 species of freshwater fish are estimated to occur in the Neotropics and approximately 3,000 in the Afrotropical region. This greatly exceeds the roughly 1,400 species found in the lakes and rivers of North America and 330 species of Europe (Lévêque et al. 2008). In an analysis of the distribution of freshwater taxa, Tisseuil et al. (2013) found greater species richness and endemism of fishes, aquatic amphibians, birds, and mammals in tropical and subtropical large drainage basins. Endemism refers to the number of species that only occur in one area and are not found elsewhere. For all taxa except fishes, variation in global distribution was related to differences in climate and productivity among regions. For fishes, drainage area and environmental heterogeneity (altitudinal range, land cover, and climate variability) were the main drivers of global diversity patterns. By analyzing the global distribution of endemic fishes, Hanly et al. (2017) found that the species richness of endemic fishes increased with lake age and area, and with river basin area, and decreased with latitude.
Explanations for the latitudinal diversity gradient (LDG) and greater diversity in the tropics frequently invoke the species-energy hypothesis, which posits that more species can be supported in areas of higher productivity. Tropical environments, with higher solar insolation and warm temperatures throughout the year, are on average more productive, resulting in greater species diversity. Species-energy relationships may also play an important role in shaping community assemblages. When considering community structure through this lens, energy availability, rather than the size of an area, is considered the dominant factor influencing species richness (Wright 1983). Greater energy (e.g., net primary productivity) in an ecosystem suggests that either there are more resources to support larger populations and so the risk of extinction is lower, or there are more resources to be used by a greater diversity of species (Hugueny et al. 2010). In analyses of global fish diversity, net terrestrial primary productivity, along with river size and flow regime, were important variables explaining gradients in diversity (Oberdorff et al. 1995; Guégan et al. 1998). Energy can also be related to climate variables such as temperature, solar radiation, and precipitation that in turn are related to the physiological tolerances of species (Turner et al. 1987; Currie 1991). In a study that included 926 basins worldwide (Fig. 11.1), area and climate-related variables explained 77% of the variation in fish diversity, although past climate and geographic isolation also appeared as important factors in shaping fish communities (Oberdorff et al. 2011). By analyzing diversity patterns of Odonata in Europe and Northern Africa, Keil et al. (2008) found that evapotranspiration, which can be used as a proxy for the energy input to an area, best explained species richness. Although some studies support the species-energy hypothesis, the mechanisms connecting energy and diversity are not clear (Hugueny et al. 2010; Heino 2011; Oberdorff et al. 2011).
The fishes of Europe and North America show a strong latitudinal diversity gradient (Fig. 11.3); however, the gradient is strong for resident but not for potadromous species (fishes that migrate within river systems), evidence that species vagility influences conformity to the LDG relationship (Griffiths 2015). Note also that these analyses extend only to 20° latitude, and so do include the greater species richness found in the tropics. The LDG also may vary with type of freshwater habitat.

(Reproduced from Griffiths 2015)
Latitudinal diversity gradients for freshwater fishes in North America and Europe. Regional species richness as a function of latitude for (a) total, (b) resident, and (c) potamodromous species for Atlantic (Mississippi regions filled circles, extra-Mississippi regions open circles), Pacific (crosses) and European (triangles) realms
Whether a latitudinal diversity gradient exists for aquatic invertebrates, resulting in more species in the tropics, is uncertain because of limited information on geographic patterns of taxonomic richness at the species level. For a very large dataset of mayflies, stoneflies, and caddisflies, Vinson and Hawkins (2003) found there was no simple latitudinal gradient in local genus-level diversity, other than a decline at very high latitudes. Ephemeroptera showed three peaks in richness at 30°S, 10°S and N, and 40°N latitude, with lower values near the poles. Plecoptera peaked at 40°N and 40°S latitude. In contrast, Trichoptera showed less latitudinal variation. Diversity of Ephemeroptera was highest in the Afrotropical realm, Plecoptera in the Nearctic, and Trichoptera in Australia. Some of this variation likely is due to incomplete sampling, and some to areas of radiation and spread. In addition, the results of the study indicated that local environmental factors explained a larger proportion of the variation than historical factors related to biogeographical realms. By comparing aquatic insects sampled in 100 streams in subtropical Brazil and in Finland, Heino et al. (2018) found richness at the genus level was greater in Brazil than in Finland at both the regional and local level, although differences were greater at the regional scale. In contrast, aquatic insect abundance was greater in Finland, likely because of higher nutrient availability.
While comparison of site-scale diversity provides insight into regional patterns, it is limited in that the relationship between site and regional diversity may not be linear. To circumvent this difficulty, Pearson and Boyero (2009) first developed statistical relationships between taxonomic diversity and regional area, and then examined whether latitude explained the remaining variation (the “residuals”) for seven freshwater taxa. Diversity was greater at higher latitudes for Ephemeroptera and Plecoptera, at low latitudes for Odonata, bony fishes, and Anura, and showed no latitudinal trend for Trichoptera and Caudata. For Plecoptera, a group that has radiated primarily in cold Nearctic regions and is found mainly at higher altitudes in the tropics, a positive relationship between latitude and residuals was obtained; in contrast, a negative relationship was found for Odonata, which are highly diverse at tropical latitudes.
Several studies in Europe document that species richness of aquatic insects declines as one moves from mid to high latitudes. Iversen et al. (2016) report that species richness of Ephemeroptera, Plecoptera, and Trichoptera (EPT) fauna declines steadily along a gradient from 35° to 75o latitude. However, the decrease in richness with latitude was most evident for fauna of the headwaters, while higher richness was observed in the middle latitudes for fauna of lowland rivers. Lower species richness at high latitudes can be explained by low productivity resulting from low temperatures and solar insolation for much of the year, and also time since glaciation for colonization and adaptation by species from more temperate regions. While species richness in Ephemeroptera, Odonata, and Plecoptera was negatively related to latitude in Europe, Heino (2009) noted that highest diversity occurred in countries with high mountain ranges such as France, Germany, and Austria, and the lowest levels of diversity occurred in lowland countries such as the Netherlands, Latvia, and Denmark, suggesting that habitat heterogeneity also plays a fundamental role in shaping species diversity.
11.1.3 Historical Explanations
Changes in landforms, climate, and connectivity over earth history provide historical explanations for patterns in species diversity, as they can influence speciation and extinction events, open and close dispersal pathways, and affect time for recolonization following disruptive events such as glaciation. Comparisons of freshwater fishes of Europe and North America provide evidence that historical events, in particular recent glaciations, have left strong signatures on their respective faunas. The European fish fauna is less diverse than that of North America, and regions within North America differ greatly. All of Canada and Alaska contain some 180 species of fish, considerably fewer than the rich Mississippi basin where most of the major adaptive radiations in North America have occurred. The Tennessee and Cumberland Rivers drainage realm alone includes some 250 species of fishes (Starnes and Etnier 1986). Species richness declines from east to west across the United States (Moyle and Cech 2006), due in part to major differences in extinction rates during the Pleistocene, as well as earlier geologic influences (Smith et al. 2010). As a consequence, the western U.S. contains only about one-fourth as many fish species as does the east.
Comparison of species-area relationships for the fish faunas of Europe and North America provides further evidence that evolutionary and biogeographic history can have a profound influence on regional diversity (Fig. 11.4). Post-glacial recolonization was more restricted in Europe relative to North America because drainage divides in Europe tend to run from east to west and re-establishment from Iberia and the Adriatic was restricted by mountain ranges, therefore glaciated areas were recolonized largely from the Danube Basin. This likely limited both southward retreat and subsequent northward recolonization for the European fauna, whereas the North American fauna had a much larger refugial area free from glaciation, and easier routes for recolonization (Mahon 1984; Oberdorff et al. 1997). In both Europe and North America, the fish faunas of glaciated regions are species-poor in comparison to unglaciated regions, and contain species that are larger, more migratory, and give less parental care compared with the unglaciated regions of the Mississippi and Missouri basins (Moyle and Herbold 1987; Griffiths 2006).

(Reproduced from Oberdorff et al. 1997)
Fish species richness as a function of drainage basin area for West European and North American rivers. Lines represent best fit using a power function
At the last glacial maximum (LGM) some 18,000 years ago, sea levels were lower and tropical forests became fragmented and reduced in extent, the result of more arid climatic conditions. Contemporary fish diversity and distribution in tropical realms bears the signature of these events. In a study including tropical basins of South America, Central America, and Africa, Tedesco et al. (2005) found that present day fish diversity was greater in basins that were connected with rain forest refuges during the LGM, when compared to regions that lacked connectivity. The authors suggested that basins connected to rain forest refuges may have had greater stability in discharge than more isolated basins. In contrast, basins without connectivity most likely experienced drier conditions and greater fish extinction rates.
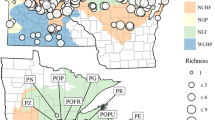
Changes in sea level during the Pleistocene due to alternating glacial advances and retreats may have influenced connectivity between drainage basins, depending on basin topography and coastal slope. Increased connectivity among currently isolated coastal river systems in ancient drainage networks (palaeo-drainages) during glaciated periods that lowered sea levels could facilitate fish dispersal among catchments. In a worldwide study analyzing the influence of historical connectivity on freshwater fishes, Dias et al. (2014) found greater diversity and lower endemism in basins that were connected during the LGM, when sea level declined by as much as 120 m, compared to those that remained isolated. These results highlight the role of dispersal processes in the present-day distribution of fishes. Similarly, past connections among coastal catchments during the LGM explained a large fraction (75%) of the genetic variation in the freshwater tetra Hollandichthys multifasciatus, endemic to streams of the Atlantic forest in southern Brazil (Thomaz et al. 2015). Present-day distributions of the 26 tetra populations sampled in this study were better explained by associations with 12 palaeo-drainages (Fig. 11.5), rather than by habitat stability, a term estimated by the permanency of Atlantic forest in the present and during the last glacial maximum.

(Reproduced from Thomaz et al. 2015)
Palaeodrainages in the Atlantic forest in southern Brazil during the last glacial maximum. The black line represents the current shoreline, and the palaeodrainages are shown in blue-green shades. Black dots represent the sampled populations
Far more ancient events in earth history also are reflected in present-day diversity patterns, as is illustrated by fish diversity gradients in the Amazon Basin. The modern Amazon was formed by the joining of its western to its eastern basins, which were isolated throughout the Miocene and only began flowing eastward 1–9 million years ago (Oberdorff et al. 2019). Present-day diversity patterns suggest that the main center of fish diversity was located in western areas, progressing eastward due to fish dispersal after the basins were united and the Amazon River assumed its modern course toward the Atlantic. This historical analysis helps to explain the unexpected finding of a decline in diversity along an upriver-downriver gradient, which is contrary to most findings but may reflect the history of the Amazon drainage network.
Comparative studies of taxonomically related fish groups clearly show that history and biogeography can influence the taxonomic and ecological diversity of a region. The Nida River in south-central Poland and the Grand River in Ontario, Canada, are two river systems that exhibit similar gradients from the headwaters downstream and occupy similar climates. Hence, they might be expected to support about the same number of species, filling roughly similar ecological roles. There are in fact many similarities (Fig. 11.6), due largely to the abundance of cyprinids in both, and comparable species in the Esocidae, Cottidae, and Gasterosteidae (Mahon 1984). Yet, there also are prominent differences, including the diversity in North America of Centrarchidae and Ictaluridae, and radiation within the genera Notropis and Etheostoma. The Grand River drainage contains more species overall, especially in smaller streams. In addition, more species in the Grand River are specialized stream dwellers, whereas the Nida includes a greater proportion of large species that are only occasional stream dwellers. Explanations for such differences always are speculative. Mahon (1984) suggests that the success of the lentic specialists (Centrarchidae and possibly Ictaluridae) in North America closed out the migratory option typical of the larger cyprinids of Europe, and favored species that formed resident populations in small streams. This may help to explain the greater North American diversity of smaller species.


(Reproduced from Mahon 1984)
Some of the fish species occupying small drainage basins (300 km2 or less) in (a) the Grand River system, southern Ontario, and (b) in the Nida River system, south-central Poland
Geographic diversity patterns of the Ephemeroptera, Plecoptera, and Trichoptera, differ among biogeographic realms. The greatest number of species of Ephemeroptera are found in the Palearctic and Nearctic regions, but higher genus diversity occurs in the Neotropical and Afrotropical realms (Barber-James et al. 2008). Plecoptera genus and species diversities are greatest in Palearctic and Nearctic realms, while Trichoptera are most diverse in the Oriental region (De Moor and Ivanov 2008; Fochetti and Tierno De Figueroa 2008). Given that the initial evolution of these taxa occurred prior to the splitting of the Pangean supercontinent (Balian et al. 2008), such disparate patterns among aquatic insects suggest either different centers of radiation or differential success in surviving subsequent environmental changes. For example, greater genus richness and endemism of Ephemeroptera in the southern hemisphere can be related to greater diversity in Gondwana than in Laurasia, greater extinction rates in the northern hemisphere, and higher climate heterogeneity in the southern hemisphere (Barber-James et al. 2008). Both recent (glaciation) and older (tectonic) geological influences are implicated by studies of the distribution of Trichoptera of the Iberian Peninsula. Consistent with other studies of Mediterranean basins, Bonada et al. (2005) found that trichopteran species composition shifts from a dominance of Palearctic species in northern basins to an increasing presence of North African and endemic species in southern basins. This reflects the combined influence of glaciations during the Quaternary that shifted northern species to more southern latitudes, and the joining of the Eurasian and African plates during the Tertiary that enabled North-African species to enter the southeast of the Iberian Peninsula.
It is evident that species-area, species-energy, and earth history all are valid explanations for large-scale patterns in species richness. From a number of analyses, Oberdorff et al. (2011) concluded that the macroecological variables of drainage area and system productivity provided the strongest explanations of global patterns in freshwater fish species richness, and history less so, with the caveat that the influence of history is more challenging to establish. Distance from a large area providing refuge from glaciation was one measure of historical influence with some explanatory power. It should be noted, however, that after river size and net primary productivity are factored out, North American rivers are still 1.7 times as rich as European ones (Oberdorff et al. 2011), and their fish assemblages differ in body form, size, and other aspects (Fig. 11.6) that no doubt have their explanations in the origin of lineages. Clearly, the distribution of species at large scale varies greatly, for reasons that we at least partly understand. We turn next to a question of much importance for the study of ecological communities: how does local species richness vary with regional species richness?
11.2 Local Patterns in Species Diversity
The most basic information regarding a local assemblage of species includes which species are present, what are their relative abundances, and can we expect to encounter the same species in the same relative abundance at the next sampling. In general, we shall see that the local assemblage is a subset of the species pool of the region. The number of species recorded is strongly influenced by sampling effort, thus requiring some standardization of methods in order to make valid comparisons. As is true in all ecological communities, a moderate number of species are common and most are rare. The most abundant species at a site tend to persist over time, unless environmental conditions change. We begin with an assessment of these patterns before proceeding to a consideration of the forces responsible.
11.2.1 Influence of the Regional Species Pool
Local richness is influenced both by regional richness and by local factors. A linear relationship between regional and local species richness suggests that local environmental conditions do not limit local richness, while an asymptotic relationship suggests that local environmental or biotic factors may ultimately limit the number of species at a site (Heino et al. 2009). From an analysis of fish species richness across a large area of the southwestern Iberian Peninsula of Europe, encompassing 436 sites distributed across 23 river basins, Filipe et al. (2010) found that local species richness (LSR) was strongly associated with regional species richness (RSR) for both native and introduced species (Fig. 11.7). Environmental factors including rainfall seasonality and stream slope also influenced local richness, but had less influence. However, considerable variation at the local scale remained unexplained, indicating a role for additional environmental variables and biological interactions. A study of the LSR–RSR relationship in stream diatoms in boreal streams in Finland found a relatively strong linear relationship between diatom species richness sampled from stream riffles, and regional species richness across multiple drainages on a 1,100 km north-south latitudinal gradient (Soininen et al. 2009). Roughly speaking, the number of diatoms collected from a single riffle ranged from a maximum of one-third to one-half of the number in the drainage system, to a minimum of 10–25%. Using data on macroinvertebrate assemblages from 705 streams sampled as part of the Swedish national lake and stream survey, Stendera and Johnson (2005) found no simple generalization between LSR and RSR. However, it did appear that maximum local species richness was usually in the range of 15–35% of regional species richness.

11.2.2 Influence of Sampling Effort
A complete survey of all the species within a local assemblage is an extremely challenging task. Such studies are unusual partly because the taxonomic knowledge of many groups is inadequate, and partly because the exhaustive compilation of a species list is rarely a priority. It is more usual to find either a detailed study of a single taxon, or an ecological investigation where the focus is on the more common species, while taxa that are difficult to identify are lumped, often at the family or genus levels. Estimated species richness at a locale is highly sensitive to sample size and the thoroughness with which all habitats are sampled. This is because many species are rare, hence often undetected, but an increasing number of rare species will be included as sampling effort increases. Two statistical approaches are commonly used to address this issue and make sample estimates more comparable across sites (Chao et al. 2014). Rarefaction uses a plot of number of species against number of samples from a random re-sampling of the data set to estimate species richness for a given sample size. Asymptotic estimators extend the species accumulation curve to derive an estimate of species richness that is relatively independent of additional sampling effort.
The importance of sample size is nicely illustrated by a study of macroinvertebrates collected from individual stones in a large reach of rapids (20–40 m wide, approximately 1 km in length) of the River Lestijoki, Finland (Kuusela 1979). The number of individuals per stone was positively correlated with the number of species per stone (Fig. 11.8a). In addition, the cumulative number of species increased with the logarithm of the cumulative number of stones sampled; that is, at a decelerating rate. The latter relationship has been reported from streams of widely different regions (Fig. 11.8b), and clearly illustrates the dependence of local species richness on sampling effort. Whether sampling included one or multiple habitats also can influence the number of species collected. For diatoms, Smucker and Vis (2011) found that a multiple habitat approach (pools, riffles, rocks, sand) resulted in higher species richness and evenness than when sampling only epilithic habitat (rocks in riffles).

(a) The number of species collected increases with the size of the sample, illustrated by Kuusela’s (1979) study of the fauna on individual stones in a large Finnish river. (b) The cumulative number of species collected increases with the logarithm of cumulative number of stones sampled: 1, River Javavankoski, Finland (Kuusela 1979); 2, Vaal River, South Africa (Chutter and Noble 1966); 3, Lytle Creek, Utah (Gaufin et al. 1956); 4, Rio Java, Costa Rica (Stout and Vandermeer 1975). Solid lines are 95% confidence limits
11.2.3 Common and Rare Species
A general observation from surveys of ecological communities is that a few species are common and most are quite rare. A collection of 52,000 insect specimens that emerged as adults from a stream flowing underneath an 11-m2 greenhouse near Schlitz, Germany, yielded a total of 148 species, but the 15 most abundant species contributed 80% of the total number of individuals (Illies 1971). Woodward et al. (2002) report a similar finding for the Broadstone Stream, UK, which is relatively species-poor due to the stream’s acidity derived from acid deposition. Demonstrating remarkable consistency over 24 years of study, a core community of eight taxa always was present, contributing 75% or more of total individuals (Fig. 11.9). A practical consequence of the tendency for a few species to dominate an assemblage is that collection of a small number of samples will include most of the common species, whereas further sampling effort will continue to produce additional species almost (but not quite) indefinitely. This underlies the relationship between sample size and local species richness (Fig. 11.8), which in turn influences the amount of sampling effort necessary to characterize a system.

(Reproduced from Woodward and Hildrew 2002)
The proportion of sampling occasions that included each taxon over 24 years of study in the Broadstone Stream, U.K. Numbers along the lower axis refer to individual species. The core community consisted of eight taxa that were always present. These included 1, Nemurella pictetii; 2, Leuctra nigra; 3, Plectrocnemia conspersa; 4, Sialis fuliginosa; 5, Pentaneurini; 6, Ceratopogonidae; 7, Heterotrissocladius marcidus/Brillia modesta; 8, Polypedilum abicorne. The pH optima for individual taxa are indicated where known
Why a few species should be more abundant, widespread, and successful and many species quite rare remains one of the great enigmas of ecology. Species that have wide regional distributions usually are locally abundant as well (Gaston and Blackburn 2000). By comparing stream insects collected at 50 headwater stream sites located in a drainage basin with data collected at 110 stream sites in 5 ecoregions in Finland, Heino (2005) found a positive and significant relationship between the local abundance of species and regional distribution. Although such a relationship can be an artifact of sampling, such as failing to detect rare species, it is likely that the explanation has an ecological basis in organism-niche relationships, dispersal abilities, and population growth rates.
Because most species are minor components of ecological webs, the completeness of the species list may not matter greatly for most ecological analyses. Nonetheless, the few exhaustive inventories are of interest because they give a sense of just how biologically diverse stream communities can be. Perhaps the most complete species lists come from long-term studies of a small German stream, the Breitenbach (Table 11.1), and of the Broadstone Stream in the UK (Schmid-Araya et al. 2002). More than one thousand invertebrate species have been collected from the Breitenbach, and because this list was compiled using aerial as well as aquatic collections it is uncertain what fraction derives from habitats other than the stream, including a small impoundment and other standing water habitats. However, it is thought to be less than one third of the invertebrate species in the stream (P. Zwick, personal communication). This compilation indicates that the greatest invertebrate diversity is located in a few groups, including several minute, interstitial phyla (Nematoda, Rotatoria, Annelida, and Platyhelminthes) and the highly diverse Diptera, especially the midge family Chironomidae. In general, it is the smallest taxa that are most diverse (Palmer et al. 1997). The Broadstone Stream is relatively depauperate because of high acidity; however, its count of 131 invertebrate species likewise reflects high representation by small taxa that are often overlooked (Schmid-Araya et al. 2002).
In summary, there are numerous factors that contribute to some areas being relatively rich in species while other areas are less so. A first level of explanation must take into account regional diversity, which is influenced by history, topography, climate, and geography; intensity of the sampling effort at both within and between-habitat levels; and local environmental conditions. There is also good reason to believe that interactions between species, which likely vary in their intensity depending upon environmental conditions, play a major role in determining local species richness. This provides the link between the topics of species diversity and community structure, and we turn now to the latter.
11.3 Local and Regional Controls of Community Assemblages
The discussion of community structure and the rules that govern community assembly has generated a rich literature in ecology. To enter into this topic, it is useful to distinguish some key ideas, keeping in mind that they are not fully independent from one another. Niche-based models focus on the interplay between biotic interactions (usually predation, herbivory, competition) and abiotic forces (primarily habitat and disturbance) that determine the suitability of a place for a particular species. The habitat template model (Southwood 1988) emphasizes the association of species with habitat features, such that individual species occur where they are best suited and more species are found where habitat conditions are most diverse. In this long-favored explanation of stream community structure, the physiological, morphological, behavioral, and life history attributes of individual species determine which will successfully colonize and maintain populations in a particular environment. Both abiotic and biotic factors can be visualized as a series of filters that determine the subset of the regional species pool that is most likely to successfully colonize and maintain populations. In contrast, explanations emphasizing stochastic variability suggest that local extinctions and dispersal are more influential than local environmental conditions in structuring communities.
Disturbance models emphasize the interplay between species interactions and variation in flow, temperature, and other environmental factors that periodically reduce the abundance of some or all species in an assemblage. Because predation, competition, and herbivory can potentially eliminate local populations, disturbance can serve as a countering force, limiting the effectiveness of strongly interacting species, and either facilitate or prevent recolonization by displaced species. A focus on disturbance seems appropriate to fluvial ecosystems because they appear to be highly variable and occasionally harsh environments. In addition, benthic invertebrates and algae are patchily distributed, and this suggests that disturbance, biotic interactions, and recolonization may combine to govern population dynamics at the local scale (Townsend 1989). Because dispersal ability varies among species, individual mobility, propensity to drift, and aerial flight ability all are important traits that may permit the persistence of weak competitors and vulnerable prey in environments where they might otherwise lose out.
11.3.1 Consistency in Assemblage Composition
Evidence that communities have predictable structure, implying primacy of habitat and niche-based explanations, has often been sought by analyzing patterns in species composition. Consistency in assemblage composition over time, and similarity in assemblage structure among locations whose environments are comparable, suggests that underlying processes, rather than randomness, govern community formation. A core community of eight taxa in the aforementioned Broadstone Stream (Fig. 11.9) was present for more than two decades of study, and species turnover generally was low. High persistence of assemblage composition and dominance by the same handful of species in the stream strongly suggests that dominant species in the locale are not simply a random sampling of a larger species pool, but instead are those whose traits allow them to be especially successful in the environmental conditions at that location. This community was persistent until the 1990s when reduction in acidity due to controls in anthropogenic emissions created conditions amenable to colonization by a large, predaceous dragonfly, Cordulegaster boltoni. Additionally, an increase pH over time supported an invasion of the stream by brown trout Salmo trutta in 2005. The abundance of invertebrates declined in response to predator introduction, and food chains lengthened with the addition of top predators, while modelling indicated that persistence and food web stability decreased. Notably, most species that were observed in the initial Broadstone studies conducted in acidic waters four decades earlier were still present. Other invertebrates typically associated with less acid conditions have not colonized the streams, suggesting that shifts in food web dynamics may be delaying community recovery (Layer et al. 2011).
The degree of community persistence appears to vary with environmental conditions. Two surveys of 27 streams in the same locale as the Broadstone Stream suggested greater persistence of taxa within cold- than warm-water streams (Townsend et al. 1987). Species persistence of fish assemblages was high and similar in two streams surveyed nine years apart, but the difference between collections was greater in the stream that exhibited higher seasonal and year-to-year variation in flow regime, maximum summer temperatures, and frequency of dewatering (Ross et al. 1985). Other studies reporting less overall persistence in assemblage structure (Grossman et al. 1982) have also attributed this to environmental variability. Based on ten years of sampling in Coweeta Creek, North Carolina, Grossman et al. (1998) concluded that environmental variability in flows rather than habitat or resource availability best explained variation in fish assemblages. Indeed, lack of assemblage persistence or of relationships between species composition and habitat variables may be a frequent finding wherever unpredictable floods and droughts introduce high temporal variation into stream assemblages (Angermeier and Schlosser 1989).
11.3.2 Local Environmental Factors and Spatial Characteristics of River Networks
Spatial comparisons of biological assemblages provide insight into the roles of community structuring mechanisms. Local communities of species that are spatially separate often are partially linked by dispersal in what is referred to as a metacommunity (Wilson 1992; Leibold et al. 2004). This framework highlights the importance of spatial processes, including spatial variability in environmental conditions, in dispersal pathways and site proximity, and in the dispersal abilities of individual species. Although each local community is the result of species sorting, in which ‘filtering’ by local environmental factors ensures that each species occurs where environmental conditions are suitable, dispersal from neighboring communities also plays a role (Heino et al. 2015). Comparisons among biological communities across space should find greatest differences when local conditions are variable and dispersal is low. Conversely, when dispersal rates are high, communities are typically more similar, especially when among-site environmental differences are modest, and species can often persist in habitats that are less favorable because of frequent recolonization (Leibold et al. 2004; Chase et al. 2005). Populations of organisms with the ability to disperse over long distances are often strongly influenced by local environmental factors, which determine whether a species can persist once it reaches a given locality. In contrast, populations of species with lesser dispersal ability are frequently limited by the relative connectivity of a system, as spatially and/or temporally heterogeneous environments may constrain their ability to colonize new habitats (Heino 2011; Padial et al. 2014).
In a study of macroinvertebrate assemblages from ten grassland streams in the Taieri River catchment on the South Island of New Zealand, Thompson and Townsend (2006) posited that if assemblage structure was explained best by dispersal, the greatest similarity among community assemblages would be in adjacent sites. However, if more distant assemblages were similar, it would potentially lend support to the argument that similar environmental conditions were more important in structuring communities. Their results indicated that dispersal was the primary factor. Assemblage similarity was best explained by spatial proximity for species with low dispersal ability, a mixed model worked best for species with moderate dispersal abilities, and neither model worked especially well for species of high dispersal ability. It is important to note that there is a tendency for ecological conditions to be spatially correlated; therefore, using comparisons of natural assemblages to test models based on species sorting by environmental conditions versus dispersal of ecologically equivalent species in structuring communities should be done with caution. However, in this study, Thompson and Townsend (2006) did not find a relationship between distance and the similarity of physical and chemical conditions for the ten streams. Thus, they attributed the observed negative relationship between spatial distance and community similarity to the distance limitations of aerial dispersal and the possible influence of chance order of arrival on community assembly.
The species of tropical floodplain rivers provide a natural experiment in community assembly because seasonal fluctuations in discharge result in repeated cycles of extirpation and recolonization of floodplain habitat at the local scale. By experimentally manipulating habitat complexity, Arrington et al. (2005) showed that species-specific differences in dispersal ability of fishes significantly affected the fish assemblage response to changes in habitat availability. Dispersal was most important to community assembly in newly formed habitat patches, whereas in older patches, abundances of individual species increasingly were influenced by habitat characteristics. Similarly, Padial et al. (2014) assessed the relative importance of environmental and spatial variables to aquatic communities of the Paraná River floodplain in Brazil. Spatial variables, primarily those related to distance between sampling sites and connectivity between sites, were more important in explaining variation in the presence of larger organisms with lower dispersal abilities, such as fish and macrophytes, while local environmental variables were more important in explaining the distribution of smaller organisms, such as phytoplankton, that had greater dispersal capacities (Fig. 11.10).

(Reproduced from Padial et al. 2014)
Difference between the contribution of environmental and spatial variables (E-S) for different biological groups ordered based on their dispersal ability, decreasing from left to right. (Phyt, phytoplankton; Peri, periphyton; Zoop, zooplankton; BMac, benthic macroinvertebrates; Fmig, migratory fish; Fsed, sedentary fish; Macr, macrophytes)
The roles of local conditions and dispersal capabilities in explaining patterns in community assemblage can vary depending on the position of a site within a river network. Headwaters are more isolated than larger streams because the overland distance between headwater streams is greater, and organisms have to move against the current to colonize upstream sites (Heino et al. 2015). Several studies have showed that local processes are more important in headwater communities than in downstream reaches. Brown and Swan (2010) found no correlation in headwater streams between similarity of macroinvertebrate communities and distance between sites (Fig. 11.11a); however, there was a significant correlation between community assemblage and environmental conditions, suggesting that species sorting rather than dispersal was the main factor influencing community structure in headwater streams (Fig. 11.11c). In contrast, in mainstem sites, similarity between communities declined with increasing distance (Fig. 11.11b), and there was a positive relationship between community assemblage and the similarity of environmental conditions (Fig. 11.11d). This suggests that community assemblage in the mainstem is influenced both by dispersal capabilities and local species sorting. However, other studies have not found clear associations in species composition between headwater and mainstem sites. The relative importance of local conditions and spatial variables to community composition differed among taxonomic groups in streams and rivers of the Danube basin in Hungary (Schmera et al. 2018). For macroinvertebrates, local environmental variables had a strong influence on community structure in streams, while both environmental conditions and spatial variables were important in communities in larger systems. For fishes and diatoms, environmental conditions and spatial characteristics influenced community assemblages in smaller streams, but did not explain variation in assemblage structure in the larger sites.

(Reproduced from Brown and Swan 2010)
Relationships between community similarity (Bray-Curtis) and geographic distance among sites for benthic macroinvertebrates across 52 sites in (a) headwaters and (b) mainstems. Relationships between community similarity and environmental similarity for the 52 sites in (c) headwaters and (d) mainstems
Seasonal variation also can affect the relative influence of local environmental conditions on the composition of aquatic communities along a river network. In northern Sweden, Göthe et al. (2013) found that local variables including water chemistry and frequency of low flows acted as strong filters at upstream sites in all seasons, but were important at downstream sites only during autumn, suggesting that seasonal changes in dispersal can create temporal variation in community assemblages. Comparing the effects of local (e.g., flow regime and habitat characteristics) and regional (e.g., distance among sites) factors in dryland streams of Arizona, US, Cañedo-Argüelles et al. (2015) found that local variables were most important in explaining community assemblages. This is unsurprising, as dryland systems are characterized by distinct seasonality in flow and spatial heterogeneity in environmental conditions, including in both perennial and intermittent reaches. Similar to many of the aforementioned studies, the effect of local environmental conditions on community assemblage varied among taxa with different dispersal capacities. The distributions of invertebrates with low dispersal capacities were most influenced by local habitat conditions, while strong dispersers were present in all suitable habitats. Interestingly, the presence of taxa with intermediate dispersal capacities was related to topographic distance, suggesting that these species can escape from most harsh conditions, but do not have the ability to reach all suitable habitats.
How dispersal capabilities and spatial relationships of the river system influence community assemblages is governed by the size of a river network and connectivity within the network (Heino et al. 2015). Depending on connectivity within a river network and dispersal mode of organisms, stream systems can differ in the importance of dispersal in overcoming spatial fragmentation of populations and communities (Tonkin et al. 2018). In low connectivity systems, such as arid-land streams that can be highly fragmented by episodic drying, the river network will play a smaller role as a dispersal route than in systems with continuous connectivity by water (Fig. 11.12). Variation among organisms in dispersal mode also influences how species distributions interact with connectivity and spatial characteristics of a watershed. Freshwater fishes can only disperse along aquatic corridors, but many macroinvertebrates and amphibians are only restricted to aquatic environments for part of their development. In headwater streams of the Central Amazon, Stegmann et al. (2019) found that distance to large-river floodplain systems had a greater influence on fish assemblages than distance among sites or local conditions. Streams located closer to large rivers showed greater taxonomic and functional richness, suggesting that floodplains can influence upstream assemblages, probably through increased dispersal during flood pulses and higher habitat availability during the dry season. In contrast, overland connectivity influences the structure of aquatic invertebrates, as Razeng et al. (2016) found for streams in central Australia. Dispersal through valleys was more important in structuring invertebrate communities than dispersal based on distance between sites or through waterways, likely related to the arid conditions of the study sites. Compared to other studies conducted in arid lands, local environmental conditions had little influence on the community structure in this example, probably because the study sites were relatively homogenous.

(Reproduced from Tonkin et al. 2018)
Conceptual model of the explanatory power of river network, degree of connectivity, and dispersal mode to local community composition via overall dispersal. Taxa illustrating differences in dispersal mode: (a) diving beetle, Boreonectes aequinoctialis; (b) giant waterbug, Abedus herberti; (c) desert sucker, Catostomus clarki; (d) dragonfly, Ophiogomphus occidentis; (e) crayfish, Pacifastacus leniusculus; (f) rainbow trout, Oncorhynchus mykiss
Understanding the relative influence of the factors governing community structure can provide insight to management options (Datry et al. 2015). For example, when local environmental variables are most important in shaping local communities, conservation efforts may be more effective if they are focused on local habitat conservation. In contrast, in systems where dispersal is the main factor regulating the distribution of species and community assemblage, efforts to maintain connectivity within a river network and to enhance refuges or areas of the stream that are relatively buffered from disturbance during times of limited connectivity should be prioritized.
11.3.3 The Habitat Template and Species Traits
Habitat template theory places particular emphasis on the matching of habitat requirements of individual species to the abiotic and biotic conditions of a locale (Southwood 1988; Townsend and Hildrew 1994). Increasingly, such efforts examine the traits of species with the expectation that attributes such as size, body shape, lifespan, and mode of dispersal will help us understand why certain species succeed where others do not, and may also provide clues regarding the environmental factors that are responsible for structuring communities. A conceptual elaboration of this approach connects the regional to the local species pool through a hierarchical series of filters that determines the likelihood that a particular subset of colonists will be successful at a locale (Fig. 11.13). As initially developed by Tonn et al. (1990) and Poff (1997), and further elaborated by Heino (2009) and Cornell and Harrison (2014), the local assemblage of species is filtered from a regional species pool first through dispersal barriers, which are influenced by factors such as topography and climate, but also by differences in species’ dispersal capabilities; and secondly by environmental filters that exist at a hierarchy of scales, such as flow and temperature regimes, and macro- and microhabitat variables. We expect systems that experience frequent disturbance and recolonization to be strongly influenced by the species pool, whereas those more strongly structured by species interactions may be less so.

(Reproduced from Poff 1997)
The number and types of species at a site reflect the influence of regional diversity and species traits (trophic, habitat, life history, etc.) that allow them to pass through multiple biotic and abiotic filters at hierarchical spatial scales. The species found within a particular microhabitat possess traits suitable for prevailing watershed/basin, valley bottom/stream reach, and channel unit and habitat conditions
The ability of individual species to reach a location and then persist there in the face of whatever environmental conditions and biological interactions a species encounters will be determined by many aspects of its morphology, physiology, life history, and dispersal ability. There are referred to as species traits, and biological assemblages increasingly are being viewed from the perspective of species traits rather than taxonomic identity (Poff et al. 2006; Menezes et al. 2010). Using a trait-based approach allows scientists to more efficiently make comparisons of species assemblages among regions because traits, rather than species, may be shared more commonly among sites (Verberk et al. 2013). Suites of traits have been defined for a variety of taxonomic groups, including plants, algae, invertebrates, and fishes (Statzner and Bêche 2010). Over the past few decades, traits have been applied in the study of community assembly (Poff 1997; Lamouroux et al. 2004), stream health and biomonitoring (Menezes et al. 2010; Statzner and Bêche 2010), community and ecosystem function (Naeem and Wright 2003; Tolonen et al. 2017), and in predicting species invasions (Olden et al. 2006; Statzner et al. 2008).
Traits are typically divided into biological and ecological categories (Vieira et al. 2006). Biological traits traditionally include morphological, physiological, and life history attributes, while ecological traits are related to habitat preferences. However, suites of traits are different for different taxonomic groups. In fishes, traits typically reflect life-history strategies (e.g., longevity, migration), body size, reproductive attributes, trophic habits, habitat preference, and tolerance to physicochemical variables (Frimpong and Angermeier 2010). In macroinvertebrates, biological traits typically include attributes such as maximum body size, number of reproduction cycles and descendants per year, method of reproduction, method of dispersal, resistant life stages, respiration mode, locomotion mode, diet, and feeding habits. Ecological traits for macroinvertebrates are frequently related to habitat requirements such as altitude, substrate, lateral and longitudinal distribution, and to physicochemical tolerance (e.g., salinity, temperature, and pH among other variables) (Schmera et al. 2017). Each trait typically includes several subcategories or modalities (Table 11.2) that are assigned specific codes and values that are used for empirical analyses. Specialists have generated databases of traits for fishes and macroinvertebrates that have facilitated the use of species traits to address ecological and management questions (Macroinvetebrates: Usseglio-Polatera et al. 2000; Poff et al. 2006; Tomanova et al. 2006; Statzner et al. 2007; Fishes: Noble et al. 2007; Frimpong and Angermeier 2009). However, further work is still needed to expand these data bases to other genera and regions (Statzner and Bêche 2010). Verberk et al. (2013) emphasize that traits do not occur in isolation, nor does a single trait define the relationship between a species and the environment. Rather, the combination of traits within a species govern the adaptive response of an organism to environmental conditions. Thus, environmental conditions do not act on single traits, but rather on a species, which is an assemblage of traits.
Efforts to demonstrate that species traits match with environmental variables have met with at least moderate success. To test the hypothesis that functional organization of fish communities is related to hydrological variability, Poff and Allan (1995) described habitat, trophic, morphological, and tolerance characteristics using six categories of species traits for each of the 106 fish species present at 34 sites in Wisconsin and Minnesota. Two ecologically-defined assemblages were identified, associated with either hydrologically variable streams (high variation in daily flows, moderate frequency of spates) or hydrologically stable streams (high predictability of daily flows, stable baseflow conditions). Fish assemblages from variable sites had generalized feeding strategies, were associated with silt and general substrate categories, were characterized by slow-velocity species with headwater affinities, and were tolerant of sedimentation. These findings indicate that hydrologic regime acts as an environmental filter, supporting theoretical predictions that variable habitats should harbor resource generalists whereas stable habitats should include a higher proportion of specialist species.
The hierarchical filter model implies that associations between traits of the species assemblage and habitat variables should exist at multiple spatial scales. Lamouroux et al. (2004) assessed correlations between traits and environmental variables across spatial scales within two river basins in France, after first summarizing the functional composition of invertebrate communities using 60 categories of 12 biological traits (Table 11.2). Roughly half of the tested relationships between traits and environmental variables were significant at the microhabitat scale, and about one-fourth of the tests were significant at the reach scale. Although a number of invertebrate traits differed between basins, this was not attributable to between-basin habitat differences. In this example, filters at the microhabitat scale clearly were most influential on the suite of community traits. Using 11 ecological traits and 11 biological traits, Usseglio-Polatera et al. (2000) identified gradients in body size, reproductive rate, and feeding ecology within 472 European macroinvertebrate taxa. Because they were able to aggregate taxa into groups with similar traits, the authors speculated that improved resolution of habitat affinities or response to pollution might be attained using a subset of species sharing a similar suite of traits, rather than using the entire assemblage.
Any consideration of species traits must recognize that some traits tend to co-occur and others may rarely, if ever be found, in the same species. For example, large body size, long lifespan, and low reproductive potential form one common suite of attributes; small body size, short lifespan, and high reproductive potential another. These are frequently referred to as slow and fast, or K and r species, respectively (Begon et al. 2006). Mixtures between these two suites of attributes are rare, suggesting a trade-off exists between two alternate life styles, or these two suites of traits may represent two ends of a spectrum.
Three life history strategies for fishes in tropical regions and in North America have been proposed: (i) opportunistic (characterized by small body size, earlier maturation, high reproductive effort, low fecundity, and low investment in individual offspring), (ii) periodic (characterized by large body size, late maturation, moderate reproductive effort, high fecundity, and low investment per offspring), and (iii) equilibrium (characterized by variable body size, moderate to long life span, low reproductive effort, low batch fecundity, and high investment per offspring). In this model, the r strategy is divided into opportunistic and periodic strategies and the equilibrium strategy is related to K species (Winemiller and Rose 1992; Winemiller 2005). This model has been applied to investigate the relationship between fishes and environmental variables, species distribution, the role of local and regional factors on fish assemblages, and the impact of hydrologic alteration in rivers (Hoeinghaus et al. 2007; Frimpong and Angermeier 2010).
The relative contribution of local environmental conditions and spatial characteristics of a river network on community structure, described earlier for species, has also been investigated using species traits. Using both taxonomic and functional approaches, Hoeinghaus et al. (2007) found that the taxonomic make-up of fish assemblages in Texas, US, was related to species’ geographic distributions and historical processes, and linked primarily to dispersal, suggesting that regional factors strongly influenced local fish assemblages. However, functional analyses based on trophic and life-history traits indicated that local factors such as habitat stability and predation pressure can have equal influence on local assemblage composition. The functional approach was able to distinguish more stable, deepwater sites dominated by piscivores from sites with more variable habitat where opportunistic groups were dominant. This study suggests that trait analysis may be a more effective approach than species identity in describing relationships between assemblages and the environment. Similar results were obtain by Logez et al. (2013) for fishes in rivers of Europe, where temperature and physical structure (e.g., slope, benthic sediment structure) were the primary environmental variables that explained trait distribution in the community, indicating that similar environmental conditions resulted in similar trait distributions.
Additional studies have found trait-based analyses to provide insight into the relative influence of environmental and spatial factors. An analysis of species and trait composition of macrophytes, macroinvertebrates, and fishes in headwaters and larger streams in Denmark found that environmental variables were more strongly associated with variation in traits than with taxonomic composition of fishes and macroinvertebrates (Göthe et al. 2017). Larger scale spatial factors had a stronger influence on macrophyte and fish communities, likely related to their lower dispersal capacity compared with macroinvertebrates. In the River Teno drainage in Finland and Norway, Tolonen et al. (2016) also found that trait composition of macroinvertebrate communities was more strongly related to local environmental variables than was species composition, indicating that local processes may act as filters to combinations of traits.
Several studies have compared the performance of species traits and taxonomic composition in detecting anthropogenic impacts on riverine systems. An investigation of the impacts of agriculture on macroinvertebrate communities in streams of New Zealand was able to distinguish sites based on land use practices from analyses of both traits and taxonomy. However, differences among the land use categories were more pronounced using traits, such as number of reproductive cycles, life duration of adults, egg laying modes, and parental-care behavior (Dolédec et al. 2006). Similarly, trait analysis was informative in a study of the responses of macroinvertebrates to a gradient of human disturbance in the Han River, the largest tributary of the Yangtze River in China (Li et al. 2019). Multivoltinism, fast development, short life span, and small body size were dominant at more disturbed sites, while univoltine species, characterized by slower development, larger body size, and longer life span, were more abundant at reference sites. Interestingly, species with strong flying dispersal capacity were found at more disturbed sites, most likely because this trait facilitates dispersal and recolonization. Burrowers and sprawlers were also more abundant at more disturbed sites, possibly due to their ability to cope with increased siltation. Both trait and taxonomic composition differed with extent of human disturbance, but environmental conditions explained a larger proportion of trait than taxonomic variation, as suggested by the habitat template model. Further application of trait analysis to the study of anthropogenic impacts will benefit from identification of additional traits that more precisely indicate the causes of changes to lotic communities (Statzner and Bêche 2010; Verberk et al. 2013).
Although assemblage diversity traditionally has been evaluated based on the number and relative abundance of species or other taxonomic unit, recent attention has turned to the diversity of organisms’ functional traits. Functional diversity and richness are determined by converting an assemblage of species into an assemblage of traits, most commonly using feeding habits, body size, substrate preferences, and locomotion. Functional diversity is then calculated from the relative abundance of organisms possessing those traits (Schmera et al. 2017).
Recent evidence suggests that functional diversity can reveal differences among assemblages more effectively than analyses based on species richness and abundance (Cadotte et al. 2011; Gagic et al. 2015). For example, Ephemeroptera, Plecoptera, and Trichoptera exhibited no differences in taxon richness between land use categories in savanna streams of the Cerrado, Brazil (Castro et al. 2018). However, functional richness and functional dispersion (measures of functional diversity) were higher in the less disturbed site, where Shannon’s index of species diversity was also greater (Fig. 11.14). This reduction in functional diversity indicates that environmental conditions in the most disturbed habitats could support only a limited number of functional roles, consistent with the habitat template hypothesis. In other cases, however, species richness and functional measures of diversity can show similar patterns in the same system. Pease et al. (2012) observed the expected increase in species richness and functional diversity of fishes from the headwater to the lower reaches in the Grijalva River, Mexico, likely related to greater food and habitat resources in the lowland reaches.

(Reproduced from Castro et al. 2018)
Difference among three categories of land use pressure in (a) genus richness, (b) functional richness, (c) Shannon diversity, and (d) functional dispersion. Medians (lines), quartiles (box end), and maximum and minimum values (whiskers) are represented. Significant differences among the categories are indicated with different letters
11.4 Disturbance, Diversity, and Community Structure
Hurricanes, fires, floods, and droughts are well-known examples of extreme disturbances that episodically cause high mortality to populations of many species in forest, grassland, coral reef, river, and other ecosystems. Because species vary in their resistance to disturbance, as well as rates of recolonization and recovery, disturbance can ameliorate strong biological interactions and help to maintain populations of species that might otherwise be eliminated by their consumers or competitors. Streamflow is both the most obvious and the most readily quantified disturbance variable in fluvial ecosystems, and flow variability is evident at all scales from the turbulence around a stone to the occasional extremes of major floods and droughts. Thus, small-scale disturbance might act almost continuously, whereas larger disturbances occur less frequently. Seasonal temperature extremes, pathogen outbreaks, sediment pulses from bank failure, and the arrival of a novel species each might constitute a disturbance to a particular system.
Two major axes of environmental variation that affect stream community structure are environmental harshness and disturbance frequency. These axes are independent, as one environment might be uniformly harsh, a second subject to frequent and extreme disturbances (Peckarsky 1983). Both act to restrict the abundance and diversity of species that are found at a location, but they differ in that perennially harsh environments such as those that are very cold or highly acid have an on-going effect, whereas disturbance implies an alternation with periods of more benign conditions. Thus harsh environments would be expected to contain relatively few species and experience less species replacement, whereas frequently disturbed environments would be expected to contain more species and exhibit higher species turnover. Because strong biotic interactions including competition and predation can reduce species diversity, it may be that an intermediate level of disturbance promotes diversity by ameliorating species effects on one another (Connell 1978; Ward and Stanford 1983).
Because the stream benthos is subject to ongoing turbulence and disturbance, it is possible that strong biological interactions may be frequently interrupted, resulting in a continuously shifting mosaic of habitat conditions and species colonization and replacement. This is the model of patch dynamics (Townsend 1989), and it is consistent with the patchy distribution of algae and invertebrates on the streambed (Downes et al. 1998). A fluctuating environment combined with continual dispersal and colonization permits more species to co-occur than would be true if conditions exhibited greater constancy. It also confers some regularity to pattern, because environmental circumstances are predictable in the aggregate even though they are unpredictable for any given place and time.
Individual species will differ in their vulnerability to a particular disturbance event, reflecting many aspects of morphology, behavior, and lifestyle; thus the impact of the same disturbance may differ among species (Lytle and Poff 2004). In addition, between-habitat differences occur as a consequence of bed and substrate characteristics, influencing the availability of refuges within the substrate (Matthaei et al. 1999) and resulting in microhabitats where disturbance is less pronounced (Lancaster et al. 1991; Matthaei et al. 2000).
Several authors have highlighted the importance of characterizing disturbances in terms of type, intensity, duration, frequency, and spatial extent when studying their effects on ecosystems, as it will aid in comparing events within and among systems (Lake 2000; Winemiller et al. 2010). Peckarsky et al. (2014) describes several methods to assess streambed disturbance in headwater streams, including photographs of the stream bed, hydrological indices, Shields number, and visual characterization of channel stability.
11.4.1 The Influence of Extreme Events
Changes in local and seasonal abundances of the stream biota in response to flow variation received frequent mention in previous chapters. Periphyton, benthic macroinvertebrates, and fishes all can be strongly influenced by fluctuations in flow. In piedmont rivers of the Venezuelan Andes, benthic invertebrates are subject to frequent flash floods during the rainy season and droughts during the dry season. Total macroinvertebrate abundance exhibited a strong negative relation with average monthly rainfall, used as a surrogate of flow because no stream gauges were available (Flecker and Feifarek 1994). Numbers recovered during flood-free periods due to colonization and recruitment, resulting in a strong positive relationship between insect abundances and time elapsed since the last storm (Fig. 11.15). In the Glenfinish River in Ireland, a catastrophic flood (a 1-in-50-year event) that occurred in August, 1986, produced a 70% decline in taxon richness and reduced densities to 5% of pre-flood levels. Interestingly, responses to the flood differed among taxa. The smallest and more abundant chironomids were less affected than Ephemeroptera, Plecoptera, and Trichoptera, which needed 3–5 years to recover. Insects with fast life cycles (r-strategy) recovered faster than slow life cycle, large-bodied species (K-strategy), which in turn showed faster recovery than mollusks and crustaceans (Woodward et al. 2016).

(Reproduced from Flecker and Feifarek 1994)
The number of aquatic insects in a river of the Andean foothills subject to a pronounced dry season and frequent floods during the rainy season. Time elapsed since the most recent rainfall event >25 mm is used in lieu of stormflow data
Disturbance has been shown to have dramatic, ecosystem-wide effects. Examples include those where floods and droughts are of sufficient magnitude that habitat is disrupted and many of the organisms present are displaced, comparative studies of rivers basins with different disturbance regimes, and instances where disturbance most severely affects a species that is a strong interactor within a community. Sycamore Creek, Arizona, is a desert stream subject to extreme flash floods that might occur a few times annually. Fisher et al. (1982) describe the recovery of the system following a late summer flood, until another flood some 60 days later re-started the sequence. Biomass of algae and invertebrates were reduced by nearly 100%, but recovery occurred quickly, particularly by the algae, which initially was dominated by diatoms and later by filamentous green and blue-green algae (Fig. 11.16). Macroinvertebrate recovery also was rapid, but slower by several weeks than the algal recovery. Nutrient uptake and community metabolism changed over community succession, and this highly productive system began to export surplus primary production. The Sycamore Creek example nicely demonstrates the effects of a disturbance that occurs with unpredictable regularity, and so ecosystem dynamics can only be understood within the cycle of disturbance and recovery. Periodic drought obviously can severely disrupt stream ecosystems as well. Based on an evaluation of resilience and recovery of Sycamore Creek to a number of spates and droughts, Boulton et al. (1992) concluded that droughts had the greater impact.

(Reproduced from Fisher et al. 1982)
Temporal succession of the biota in Sycamore Creek, Arizona, following flooding. (a) Percent cover of algal patch types; (b) Mean invertebrate numbers and biomass after flooding. Values are means and 95% confidence intervals
Extreme drying can have profound impacts on aquatic communities. Bogan et al. (2015) described temporal shifts in community composition in intermittent, arid-land streams of the Madrean Sky Islands in Arizona, US, during seasonal drying. In these headwater systems, flow is generally intermittent and streams can remain dry for more than 9 months of the year; however, some streams are perennial, and contain macroinvertebrate taxa that are rare in the intermittent streams. As droughts begin, lateral connectivity with riparian areas is lost. This is followed by loss of longitudinal connectivity as flow ceases, leaving only isolated pools (Fig. 11.17). In response, most lotic species disappear, and only relatively tolerant Coleoptera, Hemiptera, and Trichoptera remain. The new habitat is subsequently colonized by other coleopterans and hemipterans that are not typically encountered under flowing conditions. As the drought persists, vertical connectivity is lost and invertebrate richness decreases markedly. Such declines are even evident in the drought-tolerant groups. Some invertebrates may initially migrate to hyporheic habitats, but as this refuge dries, only organisms that have evolved resting stages can remain. As flow returns to the stream, only 10–12 species of resistant taxa typically are present, but recovery is rapid, reaching abundance levels similar to perennial streams in 8–10 weeks. Recovery continues as strong aerial dispersers colonize streams. These species are drought-tolerant taxa that remained dominant in perennial pools during the drought. Total recovery of the macroinvertebrate assemblage is typically observed in streams that are near dry season refuges (e.g. headwater seeps and springs, perennial pools, flowing reaches) that maintained flow for at least 4–5 months.

(Reproduced from Bogan et al. 2015)
Changes in aquatic invertebrate species richness in streams of Madrean Sky Island as flow decreased (solid line) and recovered (dashed line). A–D: thresholds during the drying period. E: recovery by taxa resistant to drought. F: recovery via aerial recolonisation of resilient taxa. G: full recovery via multiple resistance and resilience (instream and overland) pathways. Processes contributing to species loss and recovery are indicated at the top of the figure
Notably, these patterns were observed during predictable seasonal droughts in conditions under which the macroinvertebrate community has evolved. Extreme droughts can convert perennial to intermittent systems, and researchers have documented changes in expected species composition. In particular, large predators were lost from many habitats. However, macroinvertebrate densities increased and species richness remained similar when compared to typical seasonal drying (Bogan and Lytle 2011). Collectively, these results indicate that drought severity and habitat isolation can influence local and regional invertebrate richness (Fig. 11.18). The greatest number of species were observed in streams that experienced mild and more predictable droughts and had greater connectivity to perennial refuges, while only the most resistant species are found in more isolated streams under severe drought conditions (Bogan et al. 2015).

(Reproduced from Bogan et al. 2015)
Conceptual model describing the interactive effects of drought severity (based on drying intensity and duration) and habitat isolation (distance to nearest perennial refuge) on stream invertebrate species richness. A: Richness increased by the colonization of lentic taxa during part of the year under moderate drought disturbance. B–D: Richness decreased due to high drought severity or isolation, or a combination of both. E: in highly isolated sites under high drought severity, only highly resistant or resilient taxa remain
11.4.2 Disturbance Frequency and Biotic Responses
Flow variation has been shown to mediate species interactions. Throughout most of Arizona, the introduced mosquitofish Gambusia affinis has replaced a native poeciliid, the Sonoran topminnow Poecilopsis occidentalis, largely through predation on juveniles (Meffe 1984). In mountainous regions subject to extreme flash floods, however, long-term coexistence of the two species results from the native fish’s superior ability to avoid downstream displacement. Hydropsychids (Trichoptera) and simuliids (Diptera) exhibit a similar interaction seasonally in coastal Californian streams (Hemphill and Cooper 1983). On hard substrates in fast-flowing sections, black fly larvae are more abundant in spring and early summer, while caddis larvae predominate thereafter. Winters of high discharge lead to greater numbers of simuliids, and winters of low flow lead to higher densities of hydropsychids. By scrubbing substrate surfaces with a brush, Hemphill and Cooper showed that disturbance benefited simuliids because they were the more rapid colonizers, whereas caddis larvae were superior at monopolizing space on rock surfaces. As time passed since the last disturbance, hydropsychid larvae gradually replaced simuliids due to their aggressive defense of net sites.
Because flow variation is such a pervasive feature of the fluvial environment, many organisms show adaptation to resist or minimize its effects (Lytle and Poff 2004). Timing of life cycle events such as egg-laying or emergence can be effective when flow extremes have a degree of predictability. The emergence of young rainbow trout from spawning gravel in spring appears to be an example of synchronizing a sensitive life cycle stage with the long-term average dynamics of the flow regime. The rainbow trout Oncorhyncus mykiss, one of the most widely introduced species world-wide, succeeds in new environments where the flow regime matches its native range and fails in other environments where it does not, apparently because floods harm trout fry (Fausch et al. 2001). Position shifts to protected areas are a common behavioral response to high flows, such as the relocation of fishes from the thalweg to the floodplain during a flood event in an Illinois river (Schwartz and Herricks 2005). In arid-land streams of the southwestern US prone to flash flooding, giant waterbugs (Belostomatidae) crawl out of streams in advance of the flood, using rainfall as the cue (Lytle 1999). Morphological adaptations including streamlining and other adaptations to minimize drag were described in Sect. 5.1.2.
Any pattern of disturbance that recurs with some long-term regularity is referred to as a disturbance regime. Riseng et al. (2004) characterized disturbance in 97 Midwestern US streams that differed in their frequency of spates and droughts using several measures of low and high flow occurrence, substrate movement, and summer water temperature (which can reach stressful levels during low flow periods). In locations where scouring floods reduced consumer biomass, algal biomass was strongly influenced by nutrient supply, whereas in more stable streams grazers depressed algal biomass regardless of nutrient concentration. The different disturbance regimes identified in this study were clearly a product of regional patterns in geology and precipitation, and so exhibited a degree of spatial predictability. In some California rivers, a grazing caddis larva is abundant during flood-free periods and escapes most predation due to its large size and robust case, but is highly vulnerable to high flows and rolling stones during floods (Wootton et al. 1996). During flood-free periods and in dammed rivers, the main energy pathway is from algae to the caddis; however, after flooding occurs in unregulated river segments, more energy flows to smaller grazers and then to young steelhead trout. In essence, disturbance regime acts as a switch, causing one of two possible food web configurations to become dominant.
Disturbance frequency can influence the responses of entire aquatic communities. Many studies have focused on the effects of floods; however, more recent work has been focused on low flows and drought in response to climate change and the intensification of anthropogenic water withdrawals. Using stream mesocosms, Ledger et al. (2008) investigated the effects of low flow frequency on periphyton by conducting short dewatering disturbances (6 days) in high frequency (33-day) and low frequency (99-day) cycles. In the controls, Gongrosira incrustans, a crustose green alga, was dominant and was accompanied by small unicellular diatoms (Rhoicosphenia) and blue-greens (Phormidium). With dewatering events, Gongrosira decreased in abundance due to its vulnerability to drying. When the frequency of dewatering was low, Gongrosira was able to partially recover, but under high frequency disturbances the species only grew in small patches, leaving space available to other algae, particularly mat-forming diatoms. Contrasting traits between Gongrosira and the diatoms likely explain the effects of disturbance on the algal community. Gongrosira has the ability to gain and retain space through crust formation, but cannot resist dry conditions, while diatoms can take advantage of available substrate space through rapid growth, but do not have the traits to retain space when competing with other species.
The effects of low water events can be species- and habitat-specific. Haghkerdar et al. (2019) experimentally manipulated the number of daily disturbances in stream mesocosms by simulating the effects of flood disturbance on substrate. The richness of macroinvertebrate taxa declined as disturbance frequency increased. The abundance of Chironomidae, the dominant group of organisms, was not affected by disturbance frequency, but the abundance of the remaining populations decreased as disturbance increased, suggesting that disturbance effects can vary among taxa. Walters and Post (2011) studied the effects of low flows on benthic communities by diverting between 40 and 80% of the water from three streams in the western US. Total biomass of aquatic insects declined with decreasing flows, with greater effects observed in riffles than pools. Some feeding groups such as collectors and scrapers were most affected, but family-level richness did not vary with altered flows.
In tropical headwater streams of Bolivia that experienced a range of environmental conditions related to altitude, climate, and flow regime, environmental variables were considered to be more important than dispersal capabilities in shaping communities (Datry et al. 2016). However, streams that experienced moderate environmental conditions were more influenced by dispersal. Under extreme environmental conditions, neither species sorting nor dispersal explained community structure, suggesting that neutral processes may shape communities in more extreme environments. Datry et al. (2015) speculate that under the dynamic conditions experienced by many streams and rivers, the relative contribution of species sorting and dispersal can vary as the system recovers from a disturbance. Shortly after disturbance, dispersal is an important process that enhances the arrival of organisms, but after a patch has been colonized, processes related to species sorting can shape local communities.
As previously mentioned, the effects of disturbance on communities can be mitigated by the presence of refuges, or regions of the stream buffered from a disturbance. Refuges from one disturbance event sometimes provide protection from multiple kinds of disturbance, but this is not always true. For instance, examples of refuges from increased flow include pools, floodplains, areas downstream of large boulders, and interstitial areas between rocks. Refuges from drought can be found in perennial pools, the hyporheic zone, and leaf litter accumulations. Organisms that persist in refuges after a disturbance can play a fundamental role in the persistence of populations as they can colonize habitat patches in the stream where organisms were lost due to disturbance (Lake 2000).
It is important to note that organisms can also be the source of disturbance, altering the physical, chemical, and biological structure of streams. Such species are considered ecological engineers, and their ability to influence habitat structure has the potential to affect the abundance and distribution of other species (Jones et al. 1994). The grazing fish Parodon apolineri has been classified as an ecological engineer, as it creates observable grazing scars and a patchy distribution of algae and sediments on stone surfaces. By quantifying the spatial pattern of scars in a neotropical stream and by experimentally manipulating densities of Parodon, Flecker and Taylor (2004) demonstrated that grazing enhanced habitat heterogeneity, although with time or at higher densities grazing tended to reduce algae and sediments to a more uniform condition of low abundance. However, the hypothesized correspondence between habitat heterogeneity and richness of the algal and invertebrate assemblages was not observed, perhaps because the timescale at which those taxa respond was slower than the loss and renewal dynamics generated by grazing Parodon. Sockeye salmon Oncorhynchus nerka can affect stream benthic communities through their seasonal spawning migrations. In streams of Alaska, the biomass of algae and insects tends to increase during the spring, but then decreases by 75–85% during salmon nest excavation, and also post spawning, when salmon disturb the streambed to maintain the nest free of sediments (Moore and Schindler 2008). After salmon die, algae quickly reach pre-spawning levels, but invertebrate recovery is slower. Anthropogenic activities have altered the distribution of some species of ecosystem engineers through the introduction of non-native species. Beavers (Castor canadensis), notorious for their engineering capabilities, were introduced into the Cape Horn Biosphere Reserve in southern Chile in the mid-1940s (Anderson and Rosemond 2007). They have subsequently altered riparian and in-stream diversity through changes in hydrogeomorphic and biogeochemical processes.
11.5 Summary
The number of coexisting species in ecological communities is a consequence of processes operating at both local and regional scales. The assemblage of species at a location are a sub-set of the species in the region, which are themselves a product of species formation, loss, and dispersal over earth history. The local assemblage is also the result of species sorting that is influenced by local environmental conditions and other species present, and by each species’ dispersal ability and life history characteristics.
Patterns in the regional species richness of freshwater organisms are best known for fishes, whose global distribution reveals a number of intriguing patterns. The number of species of freshwater fishes is inversely related to latitude, increases with drainage area and discharge of the river system, and differs among regions at the same latitude in both number of species and the body plans and traits of species present. Greater productivity, more habitat diversity, and more habitat area are amongst the explanations for these patterns. Historical changes in landforms, climate, and connectivity provide further insight into patterns in species diversity, as they can influence speciation and extinction events, open and close dispersal pathways, and affect time for recolonization following disruptive events such as glaciation. Comparisons of freshwater fishes of Europe and North America provide evidence that historical events, in particular recent glaciations, have left strong signatures on their respective faunas. Post-glacial recolonization was more restricted in Europe relative to North America because drainage divides in Europe tend to run from east to west, and re-establishment from Iberia and the Adriatic was restricted by mountain ranges. In contrast, the North American fauna had a much larger refugial area in unglaciated regions of the Mississippi and Missouri basins, and easier routes for recolonization. Present-day patterns in freshwater fish diversity in tropical rivers also bear the signature of past glacial events. Lower sea levels during the last glacial maximum allowed for connections at their lower termini between some now-isolated river systems, and greater aridity resulted in differential isolation among rivers during periods of tropical forest fragmentation. It appears that macroecological variables of drainage area and system productivity provide the strongest explanations of global patterns in freshwater fish species richness, and history less so, with the caveat that the influence of history is more challenging to establish. Although the historical biogeography of aquatic insects is less well known, diversity in major insect groups likewise shows continental-scale patterns such that number of species and genera differ among biogeographical regions.
Local species richness varies in proportion to regional species richness, indicating that site surveys are likely to find more species in areas with a larger species pool. A comprehensive survey of all species at a site is a challenging task, especially for invertebrates and the smallest organisms, due to incomplete taxonomic knowledge and the fact that most species are rare and escape detection in all but the most extensive sampling. As a consequence, most invertebrate studies focus on the more common species in the assemblage. Community structure refers to the organization of a biological community based on numbers of individuals within different taxonomic groups and functional roles, and the underlying processes that maintain that organization. Niche-based explanations for patterns in species diversity and community structure focus on the fit of individual species and their traits to habitat conditions, and the influence of interspecific interactions. However, environmental disturbances such as floods and droughts, when sufficiently extreme or frequent, are likely to prevent biotic interactions from acting with the strength and regularity required to result in consistent community patterns. In these circumstances, species assemblages are likely to be composed of those species best adapted to environmental extremes and those most capable of dispersal and recolonization of disrupted environments. Studies of species assemblages in streams frequently explore the relative importance of these two contrasting explanations.
All organisms have some capacity to disperse, and local assemblages most likely are connected to other, neighboring assemblages by occasional or frequent movements between them. Differences in dispersal modes, as when fish are limited to connecting waterways but insects are capable of aerial dispersal, and degree of physical connectivity, influence whether each local assemblage is structured more by local environmental conditions or more by proximity to neighbors.
Species traits such as size, body shape, lifespan, and mode of dispersal are increasingly explored as an alternative to species identity to help us understand why certain species succeed where others do not. This perspective intersects well with a conceptual framework in which the local assemblage of species is filtered from a regional species pool first through dispersal barriers, which are influenced by factors such as topography and climate, but also by differences in species’ dispersal capabilities; and secondly by environmental filters that exist at a hierarchy of scales, such as flow and temperature regimes, and macro- and microhabitat variables. Supporting evidence exists in trait-based analyses that provide insight into the relative influence of environmental and spatial factors in determining the composition of local assemblages.
The role of disturbance in local community structure has been the focus of much study in lotic ecosystems, especially with regard to flood and drought events. Local conditions can be perennially harsh, as when temperature, pH, or some other environmental variable is continually extreme, or can alternate between moderate and extreme condition, as with episodically recurring floods and droughts. This is seen in desert streams, when scoured communities accrue biomass following a flood, only to be re-set when the next flood occurs; and in tropical streams during the rainy season, when insect numbers correspond to days since the last substantial rain event. Experimental manipulation of the frequency of de-watering shows how species traits determine patterns of recovery and species replacement. Differential dispersal ability among species and connectivity within a river network also are important explanatory variables in the response of stream communities to hydrologic disturbance. Finally, not all disturbance is abiotic, as organisms also are disturbance agents. This is seen when the nest-building activities of large salmon runs disturb the stream bed, and in some tropical streams where very abundant sediment-feeding fishes create a patchwork of feeding scars on benthic substrates.
References
Amarasinghe US, Welcomme RL (2002) An analysis of fish species richness in natural lakes. Environ Biol Fishes 65:327–339. https://doi.org/10.1023/A:1020558820327
Anderson CB, Rosemond AD (2007) Ecosystem engineering by invasive exotic beavers reduces in-stream diversity and enhances ecosystem function in Cape Horn, Chile. Oecologia 154:141–153. https://doi.org/10.1007/s00442-007-0757-4
Angermeier PL, Schlosser IJ (1989) Species-area relationships for stream fishes. Ecology 70:1450–1462. https://doi.org/10.2307/1938204
Arrington DA, Winemiller KO, Layman CA (2005) Community assembly at the patch scale in a species rich tropical river. Oecologia 144:157–167. https://doi.org/10.1007/s00442-005-0014-7
Balian EV, Leveque C, Segers H, Martens K (2008) Freshwater animal diversity assessment. Hydrobiologia 595:1–637. https://doi.org/10.1007/978-1-4020-8259-7
Barber-James HM, Gattolliat JL, Sartori M, Hubbard MD (2008) Global diversity of mayflies (Ephemeroptera, Insecta) in freshwater. Hydrobiologia 595:339–350
Begon M, Harper JL, Townsend CR (2006) Ecology: individuals, populations and communities, 4th edn. Blackwell Publishing, Oxford
Bogan MT, Boersma KS, Lytle DA (2015) Resistance and resilience of invertebrate communities to seasonal and supraseasonal drought in arid-land headwater streams. Freshw Biol 60:2547–2558. https://doi.org/10.1111/fwb.12522
Bogan MT, Lytle DA (2011) Severe drought drives novel community trajectories in desert stream pools. Freshw Biol 56:2070–2081. https://doi.org/10.1111/j.1365-2427.2011.02638.x
Bonada N, Zamora-Muñoz C, Rieradevall M, Prat N (2005) Ecological and historical filters constraining spatial caddisfly distribution in Mediterranean rivers. Freshw Biol 50:781–797. https://doi.org/10.1111/j.1365-2427.2005.01357.x
Boulton AJ, Peterson CG, Grimm NB, Fisher SG (1992) Stability of an aquatic macroinvertebrate community in a multiyear hydrologic disturbance regime. Ecology 73:2192–2207. https://doi.org/10.2307/1941467
Brown BL, Swan CM (2010) Dendritic network structure constrains metacommunity properties in riverine ecosystems. J Anim Ecol 79:571–580. https://doi.org/10.1111/j.1365-2656.2010.01668.x
Cadotte MW, Carscadden K, Mirotchnick N (2011) Beyond species: functional diversity and the maintenance of ecological processes and services. J Appl Ecol 48:1079–1087. https://doi.org/10.1111/j.1365-2664.2011.02048.x
Cañedo-Argüelles M, Boersma KS, Bogan MT et al (2015) Dispersal strength determines meta-community structure in a dendritic riverine network. J Biogeogr 42:778–790. https://doi.org/10.1111/jbi.12457
de Castro DMP, Dolédec S, Callisto M (2018) Land cover disturbance homogenizes aquatic insect functional structure in neotropical savanna streams. Ecol Indic 84:573–582. https://doi.org/10.1016/j.ecolind.2017.09.030
Chao A, Gotelli NJ, Hsieh TC et al (2014) Rarefaction and extrapolation with Hill numbers: a framework for sampling and estimation in species diversity studies. Ecol Monogr 84:45–67. https://doi.org/10.1890/13-0133.1
Chase JM, Amarasekare P, Cottenie K et al (2005) Competing theories for competitive metacommunities. In: Holyoak M, Leibold MA, Holt RD (eds) Metacommunities: spatial dynamics and ecological communities. The University of Chicago Press, Chicago, pp 335–354
Chase JM, Leibold MA (2003) Ecological niches: linking classical and contemporary approaches. University of Chicago Press, Chicago
Chutter FM, Noble R (1966) The reliability of method of sampling stream invertebrates. Arch Für Hydrobiol 62:95–103
Connell JH (1978) Diversity in tropical rain forests and coral reefs. Science 199:1302–1310. https://doi.org/10.1126/science.199.4335.1302
Cornell HV, Harrison SP (2014) What are species pools and when are they important? Annu Rev Ecol Evol Syst 45:45–67. https://doi.org/10.1146/annurev-ecolsys-120213-091759
Currie DJ (1991) Energy and large-scale patterns of animal- and plant-species richness. Am Nat 137:27–49
Datry T, Bonada N, Heino J (2015) Towards understanding the organisation of metacommunities in highly dynamic ecological systems. Oikos 125:149–159. https://doi.org/10.1111/oik.02922
Datry T, Melo AS, Moya N et al (2016) Metacommunity patterns across three Neotropical catchments with varying environmental harshness. Freshw Biol 61:277–292. https://doi.org/10.1111/fwb.12702
De Mérona B, Tejerina-Garro FL, Vigouroux R (2012) Fish-habitat relationships in French Guiana rivers: a review. Cybium 36:7–15
De Moor FC, Ivanov VD (2008) Global diversity of caddisflies (Trichoptera: Insecta) in freshwater. Hydrobiologia 595:393–407
Dias MS, Cornu JF, Oberdorff T et al (2013) Natural fragmentation in river networks as a driver of speciation for freshwater fishes. Ecography (Cop) 36:683–689. https://doi.org/10.1111/j.1600-0587.2012.07724.x
Dias MS, Oberdorff T, Hugueny B et al (2014) Global imprint of historical connectivity on freshwater fish biodiversity. Ecol Lett 17:1130–1140. https://doi.org/10.1111/ele.12319
Dolédec S, Phillips N, Scarsbrook M et al (2006) Comparison of structural and functional approaches to determining landuse effects on grassland stream invertebrate communities. J North Am Benthol Soc 25:44–60. https://doi.org/10.1899/0887-3593(2006)25[44:COSAFA]2.0.CO;2
Downes BJ, Lake PS, Schreiber ESG, Glaister A (1998) Habitat structure and regulation of local species diversity in a stony, upland stream. Ecol Monogr 68:237–257. https://doi.org/10.1890/0012-9615(1998)068[0237:HSAROL]2.0.CO;2
Fausch KD, Taniguchi Y, Nakano S et al (2001) Flood disturbance regimes influence rainbow trout invasion success among five holarctic regions. Ecol Appl 11:1438–1455. https://doi.org/10.1890/1051-0761(2001)011[1438:FDRIRT]2.0.CO;2
Filipe AF, Magalhães MF, Collares-Pereira MJ (2010) Native and introduced fish species richness in Mediterranean streams: the role of multiple landscape influences. Divers Distrib 16:773–785. https://doi.org/10.1111/j.1472-4642.2010.00678.x
Fisher SG, Gray LJ, Grimm NB, Busch DE (1982) Temporal succession in a desert stream ecosystem following flash flooding. Ecol Monogr 52:93–110. https://doi.org/10.2307/2937346
Flecker AS, Feifarek B (1994) Disturbance and the temporal variability of invertebrate assemblages in two Andean streams. Freshw Biol 31:131–142. https://doi.org/10.1111/j.1365-2427.1994.tb00847.x
Flecker AS, Taylor BW (2004) Tropical fishes as biological bulldozers: density effects on resource heterogeneity and species diversity. Ecology 85:2267–2278. https://doi.org/10.1890/03-0194
Fochetti R, Tierno De Figueroa JM (2008) Global diversity of stoneflies (Plecoptera; Insecta) in freshwater. Hydrobiologia 595:365–377. https://doi.org/10.1007/s10750-007-9031-3
Frimpong EA, Angermeier PL (2009) FishTraits: a Database of ecological and life-history traits of freshwater fishes of the United States. Fisheries 34:487–495
Frimpong EA, Angermeier PL (2010) Trait-based approaches in the analysis of stream fish communities. Am Fish Soc Symp 73:109–136
Gagic V, Bartomeus I, Jonsson T et al (2015) Functional identity and diversity of animals predict ecosystem functioning better than species-based indices. Proc R Soc B Biol Sci 282:20142620. https://doi.org/10.1098/rspb.2014.2620
Gaston KJ (2000) Global patterns in biodiversity. Nature 405:220–227. https://doi.org/10.1038/35012228
Gaston KJ, Blackburn TM (2000) Pattern and processes in macroecology. Blackwell Scientific, Oxford
Gaufin AR, Harris EK, Walter J (1956) A statistical evaluation of stream bottom sampling data obtained from three standard samplers. Ecology 37:643–648
Göthe E, Angeler DG, Sandin L (2013) Metacommunity structure in a small boreal stream network. J Anim Ecol 82:449–458. https://doi.org/10.1111/1365-2656.12004
Göthe E, Baattrup-Pedersen A, Wiberg-Larsen P et al (2017) Environmental and spatial controls of taxonomic versus trait composition of stream biota. Freshw Biol 62:397–413. https://doi.org/10.1111/fwb.12875
Griffiths D (2006) Pattern and process in the ecological biogeography of European freshwater fish. J Anim Ecol 75:734–751. https://doi.org/10.1111/j.1365-2656.2006.01094.x
Griffiths D (2015) Connectivity and vagility determine spatial richness gradients and diversification of freshwater fish in North America and Europe. Biol J Linn Soc 116:773–786. https://doi.org/10.1111/bij.12638
Grossman GD, Moyle PB, Whitaker JO (1982) Stochasticity in structural and functional characteristics of an Indiana stream fish assemblage: a test of community theory. Am Nat 120:423–454. https://doi.org/10.1086/284004
Grossman GD, Ratajczak RE, Crawford M, Freeman MC (1998) Assemblage organization in stream fishes: effects of environmental variation and interspecific interactions. Ecol Monogr 68:395–420. https://doi.org/10.1890/0012-9615(1998)068[0395:AOISFE]2.0.CO;2
Guégan JF, Lek S, Oberdorff T (1998) Energy availability and habitat heterogeneity predict global riverine fish diversity. Nature 391:382–384. https://doi.org/10.1038/34899
Haghkerdar JM, McLachlan JR, Ireland A, Greig HS (2019) Repeat disturbances have cumulative impacts on stream communities. Ecol Evol 9:2898–2906. https://doi.org/10.1002/ece3.4968
Hanly PJ, Mittelbach GG, Schemske DW (2017) Speciation and the latitudinal diversity gradient: insights from the global distribution of endemic fish. Am Nat 189:604–615. https://doi.org/10.1086/691535
Heino J (2011) A macroecological perspective of diversity patterns in the freshwater realm. Freshw Biol 56:1703–1722. https://doi.org/10.1111/j.1365-2427.2011.02610.x
Heino J (2009) Biodiversity of aquatic insects: spatial gradients and environmental correlates of assemblage-level measures at large scales. Freshw Rev 2:1–29. https://doi.org/10.1608/FRJ-2.1.1
Heino J (2005) Positive relationship between regional distribution and local abundance in stream insects: a consequence of niche breadth or niche position? Ecography (Cop) 28:345–354. https://doi.org/10.1111/j.0906-7590.2005.04151.x
Heino J, Melo AS, Jyrkänkallio-Mikkola J et al (2018) Subtropical streams harbour higher genus richness and lower abundance of insects compared to boreal streams, but scale matters. J Biogeogr 45:1983–1993. https://doi.org/10.1111/jbi.13400
Heino J, Melo AS, Siqueira T et al (2015) Metacommunity organisation, spatial extent and dispersal in aquatic systems: patterns, processes and prospects. Freshw Biol 60:845–869. https://doi.org/10.1111/fwb.12533
Heino J, Mykrä H (2008) Control of stream insect assemblages: roles of spatial configuration and local environmental factors. Ecol Entomol 33:614–622. https://doi.org/10.1111/j.1365-2311.2008.01012.x
Heino J, Virkkala R, Toivonen H (2009) Climate change and freshwater biodiversity: detected patterns, future trends and adaptations in northern regions. Biol Rev 84:39–54. https://doi.org/10.1111/j.1469-185X.2008.00060.x
Hemphill N, Cooper SD (1983) The effect of physical disturbance on the relative abundances of two filter-feeding insects in a small stream. Oecologia 58:378–382. https://doi.org/10.1007/BF00385239
Hillebrand H (2004) On the generality of the latitudinal diversity gradient. Am Nat 163:192–211. https://doi.org/10.1086/381004
Hoeinghaus DJ, Winemiller KO, Birnbaum JS (2007) Local and regional determinants of stream fish assemblage structure: inferences based on taxonomic vs. functional groups. J Biogeogr 34:324–338. https://doi.org/10.1111/j.1365-2699.2006.01587.x
Horwitz RJ (1978) Temporal variability patterns and the distributional patterns of stream fishes. Ecol Monogr 48:07–321. https://doi.org/10.2307/2937233
Hubbell SP (2005) Neutral theory in community ecology and the hypothesis of functional equivalence. Funct Ecol 19:166–172. https://doi.org/10.1111/j.0269-8463.2005.00965.x
Hubbell SP (2001) The unified neutral theory of biodiversity and biogeography. Princeton University Press, Princeton
Hugueny B (1989) West African rivers as biogeographic islands: species richness of fish communities. Oecologia 79:236–243. https://doi.org/10.1007/BF00388483
Hugueny B, Oberdorff T, Tedescco PA (2010) Community ecology of river fishes: a large-scale perspective. Am Fish Soc Symp 73:29–62
Illies J (1971) Emergenz 1969 im Breitenbach. Arch Für Hydrobiol 69:14–59
Iversen LL, Jacobsen D, Sand-Jensen K (2016) Are latitudinal richness gradients in European freshwater species only structured according to dispersal and time? Ecography (Cop) 39:1247–1249. https://doi.org/10.1111/ecog.02183
Jones CG, Lawton JH, Shachak M (1994) Organisms as ecosystem engineers. Oikos 69:373–386. https://doi.org/10.2307/3545850
Keil P, Simova I, Hawkins BA (2008) Water-energy and the geographical species richness pattern of European and North African dragonflies (Odonata). Insect Conserv Divers 1:142–150. https://doi.org/10.1111/j.1752-4598.2008.00019.x
Kinlock NL, Prowant L, Herstoff EM et al (2018) Explaining global variation in the latitudinal diversity gradient: meta-analysis confirms known patterns and uncovers new ones. Glob Ecol Biogeogr 27:125–141. https://doi.org/10.1111/geb.12665
Kuusela K (1979) Early summer ecology and community structure of the macrozoobenthos on stones in the Javajankoski Rapids on the River Lestijoki, Finland. Acta Univ Ouluensis (Ser A, No. 87, Oulu, Finland)
Lake PS (2000) Disturbance, patchiness, and diversity in streams. J North Am Benthol Soc 19:573–592. https://doi.org/10.2307/1468118
Lamouroux N, Dolédec S, Gayraud S (2004) Biological traits of stream macroinvertebrate communities: effects of microhabitat, reach, and basin filters. J North Am Benthol Soc 23:449–466. https://doi.org/10.1899/0887-3593(2004)023%3c0449:BTOSMC%3e2.0.CO;2
Lancaster J, Hildrew AG, Townsend CR (1991) Invertebrate predation on patchy and mobile prey in streams. J Anim Ecol 60:625–641. https://doi.org/10.2307/5302
Layer K, Hildrew AG, Jenkins GB et al (2011) Long-term dynamics of a well-characterised food web. Four decades of acidification and recovery in the Broadstone Stream model system. Adv Ecol Res 44:69–117. https://doi.org/10.1016/B978-0-12-374794-5.00002-X
Ledger ME, Harris RML, Armitage PD, Milner AM (2008) Disturbance frequency influences patch dynamics in stream benthic algal communities. Oecologia 155:809–819. https://doi.org/10.1007/s00442-007-0950-5
Leibold MA, Chase JM (2017) Metacommunity ecology. Princeton University Press, Princeton
Leibold MA, Holyoak M, Mouquet N et al (2004) The metacommunity concept: a framework for multi-scale community ecology. Ecol Lett 7:601–613. https://doi.org/10.1111/j.1461-0248.2004.00608.x
Lévêque C, Oberdorff T, Paugy D et al (2008) Global diversity of fish (Pisces) in freshwater. Hydrobiologia 595:545–567
Li Z, Wang J, Liu Z et al (2019) Different responses of taxonomic and functional structures of stream macroinvertebrate communities to local stressors and regional factors in a subtropical biodiversity hotspot. Sci Total Environ 655:1288–1300. https://doi.org/10.1016/j.scitotenv.2018.11.222
Logez M, Bady P, Melcher A, Pont D (2013) A continental-scale analysis of fish assemblage functional structure in European rivers. Ecography (Cop) 36:080–091. https://doi.org/10.1111/j.1600-0587.2012.07447.x
Lytle DA (1999) Use of rainfall cues by Abedus herberti (Hemiptera: Belostomatidae): a mechanism for avoiding flash floods. J Insect Behav 12:1–12
Lytle DA, Poff NLR (2004) Adaptation to natural flow regimes. Trends Ecol Evol 19:94–100. https://doi.org/10.1016/j.tree.2003.10.002
MacArthur RH (1972) Geographical ecology: patterns in the distribution of species. Harper and Row, New York
MacArthur RH, Wilson EO (1963) An equilibrium theory of insular zoogeography. Evolution (N Y) 17:373–387
MacArthur RH, Wilson EO (1967) The theory of island biogeography. Princeton University Press, Princeton
Mahon R (1984) Divergent structure in fish taxocenes of north temperate streams. Can J Fish Aquat Sci 41:330–350. https://doi.org/10.1139/f84-037
Matthaei CD, Arbuckle CJ, Townsend CR (2000) Stable surface stones as refugia for invertebrates during disturbance in a New Zealand stream. J North Am Benthol Soc 19:82–93. https://doi.org/10.2307/1468283
Matthaei CD, Peacock KA, Townsend CR (1999) Scour and fill patterns in a New Zealand stream and potential implications for invertebrate refugia. Freshw Biol 42:41–57. https://doi.org/10.1046/j.1365-2427.1999.00456.x
Meffe GK (1984) Effects of abiotic disturbance of coexistence of predator-prey fish species. Ecology 65:1525–1534. https://doi.org/10.2307/1939132
Menezes S, Baird DJ, Soares AMVM (2010) Beyond taxonomy: a review of macroinvertebrate trait-based community descriptors as tools for freshwater biomonitoring. J Appl Ecol 47:711–719. https://doi.org/10.1111/j.1365-2664.2010.01819.x
Mittelbach GG, Schemske DW, Cornell HV et al (2007) Evolution and the latitudinal diversity gradient: speciation, extinction and biogeography. Ecol Lett 10:315–331. https://doi.org/10.1111/j.1461-0248.2007.01020.x
Moore JW, Schindler DE (2008) Biotic disturbance and benthic community dynamics in salmon-bearing streams. J Anim Ecol 77:275–284. https://doi.org/10.1111/j.1365-2656.2007.01336.x
Moyle PB, Cech JJJ (2006) Fishes: an introduction to ichthyology. Prentice-Hall, Englewood Cliffs, NJ
Moyle PB, Herbold B (1987) Life-history patterns and community structure in stream fishes of western North America: comparisons with eastern North America and Europe. In: Matthews WJ, Heins DC (eds) Community and evolutionary ecology of North American stream fishes. University of Oklahoma Press, Norman
Naeem S, Wright JP (2003) Disentangling biodiversity effects on ecosystem functioning: deriving solutions to a seemingly insurmountable problem. Ecol Lett, 567–579
Noble RAA, Cowx IG, Goffaux D, Kestemont P (2007) Assessing the health of European rivers using functional ecological guilds of fish communities: standardising species classification and approaches to metric selection. Fish Manag Ecol 14:381–392. https://doi.org/10.1111/j.1365-2400.2007.00575.x
Oberdorff T, Dias MS, Jézéquel C et al (2019) Unexpected fish diversity gradients in the Amazon basin. Sci Adv 5:1–10. https://doi.org/10.1126/sciadv.aav8681
Oberdorff T, Guégan J-F, Hugueny B (1995) Global scale patterns of fish species richness in rivers. Ecography (Cop) 18:345–352
Oberdorff T, Hugueny B, Guégan J-F (1997) Is there an influence of historical events on contemporary fish species richness in rivers? J Biogeogr 24:461–467
Oberdorff T, Tedesco PA, Hugueny B et al (2011) Global and regional patterns in riverine fish species richness: a review. Int J Ecol 2011:967631. https://doi.org/10.1155/2011/967631
Olden JD, Leroy Poff N, Bestgen KR (2006) Life-history strategies predict fish invasions and extirpations in the Colorado River Basin. Ecol Monogr 76:25–40. https://doi.org/10.1890/05-0330
Padial AA, Ceschin F, Declerck SAJ et al (2014) Dispersal ability determines the role of environmental, spatial and temporal drivers of metacommunity structure. PLoS One 9:1–8. https://doi.org/10.1371/journal.pone.0111227
Palmer MA, Covich AP, Finlay BJ et al (1997) Biodiversity and ecosystem processes in freshwater sediments. Ambio 26:571–577. https://doi.org/10.2307/4314671
Pearson RG, Boyero L (2009) Gradients in regional diversity of freshwater taxa. J North Am Benthol Soc 28:504–514. https://doi.org/10.1899/08-118.1
Pease AA, González-Díaz AA, Rodiles-Hernández R, Winemiller KO (2012) Functional diversity and trait-environment relationships of stream fish assemblages in a large tropical catchment. Freshw Biol 57:1060–1075. https://doi.org/10.1111/j.1365-2427.2012.02768.x
Peckarsky BL (1983) Biotic interactions or abiotic limitations? A model of lotic community structure. In: Fontaine TI, Bartell S (eds) Dynamics of lotic ecosystems. Ann Arbor Science, Ann Arbor, MI, pp 303–323
Peckarsky BL, McIntosh AR, Horn SC et al (2014) Characterizing disturbance regimes of mountain streams. Freshw Sci 33:716–730. https://doi.org/10.1086/677215
Poff NL (1997) Landscape filters and species traits: towards mechanistic understanding and prediction in stream ecology. J North Am Benthol Soc 16:391–409. https://doi.org/10.2307/1468026
Poff NL, Allan JD (1995) Functional organization of stream fish assemblages in relation to hydrological variability. Ecology 76:606–627. https://doi.org/10.2307/1941217
Poff NL, Olden JD, Vieira NKM et al (2006) Functional trait niches of North American lotic insects: traits-based ecological applications in light of phylogenetic relationships. J North Am Benthol Soc 25:730–755
Razeng E, Morán-Ordóñez A, Brim Box J et al (2016) A potential role for overland dispersal in shaping aquatic invertebrate communities in arid regions. Freshw Biol 61:745–757. https://doi.org/10.1111/fwb.12744
Ricklefs RE, Schluter D (1993) Species diversity in ecological communities. Oxford University Press, Oxford
Riseng CM, Wiley MJ, Stevenson RJ (2004) Hydrologic disturbance and nutrient effects on benthic community structure in midwestern US streams: a covariance structure analysis. J North Am Benthol Soc 23:309–326. https://doi.org/10.1899/0887-3593(2004)023%3c0309:HDANEO%3e2.0.CO;2
Rosenzweig ML (1995) Species diversity in space and time. Cambridge University Press, Cambridge
Ross ST, Matthews WJ, Echelle AA (1985) Persistence of stream fish assemblages: effects of environmental change. Am Nat 122:583–601. https://doi.org/10.1086/284393
Schlosser IJ (1987) The role of predation in age- and size-related habitat use by stream fishes. Ecology 68:651–659. https://doi.org/10.2307/1938470
Schmera D, Árva D, Boda P et al (2018) Does isolation influence the relative role of environmental and dispersal-related processes in stream networks? An empirical test of the network position hypothesis using multiple taxa. Freshw Biol 63:74–85. https://doi.org/10.1111/fwb.12973
Schmera D, Heino J, Podani J et al (2017) Functional diversity: a review of methodology and current knowledge in freshwater macroinvertebrate research. Hydrobiologia 787:27–44. https://doi.org/10.1007/s10750-016-2974-5
Schmid-Araya JM, Hildrew AG, Robertson A et al (2002) The importance of meiofauna in food webs: evidence from an acid stream. Ecology 83:1271–1285
Schwartz JS, Herricks EE (2005) Fish use of stage-specific fluvial habitats as refuge patches during a flood in a low-gradient Illinois stream. Can J Fish Aquat Sci 62:1540–1552. https://doi.org/10.1139/f05-060
Sepkoski JJ, Rex MA (1974) Distribution of freshwater mussels: coastal rivers as biogeographic islands. Syst Zool 23:165–188. https://doi.org/10.2307/2412130
Smith GR, Badgley C, Eiting TP, Larson PS (2010) Species diversity gradients in relation to geological history in North American freshwater fishes. Evol Ecol Res 12:693–726
Smucker NJ, Vis ML (2011) Contributions of habitat sampling and alkalinity to diatom diversity and distributional patterns in streams: implications for conservation. Biodivers Conserv 20:643–661. https://doi.org/10.1007/s10531-010-9972-0
Soininen J, Heino J, Kokocinski M, Muotka T (2009) Local-regional diversity relationship varies with spatial scale in lotic diatoms. J Biogeogr 36:720–727. https://doi.org/10.1111/j.1365-2699.2008.02034.x
Southwood TRE (1988) Tactics, strategies and templets. Oikos 52:3–18. https://doi.org/10.2307/3565974
Starnes W, Etnier D (1986) Drainage evolution and fish biogeography of the Tennessee and Cumberland Rivers drainage realm. In: Hocutt CH, Wiley E (eds) The zoogeography of North American Freshwater Fishes. Wiley Interscience, New York, pp 325–361
Statzner B, Bêche LA (2010) Can biological invertebrate traits resolve effects of multiple stressors on running water ecosystems? Freshw Biol 55:80–119. https://doi.org/10.1111/j.1365-2427.2009.02369.x
Statzner B, Bonada N, Dolédec S (2008) Biological attributes discriminating invasive from native European stream macroinvertebrates. Biol Invasions 10:517–530. https://doi.org/10.1007/s10530-007-9148-3
Statzner B, Bonada N, Dolédec S (2007) Conservation of taxonomic and biological trait diversity of European stream macroinvertebrate communities: a case for a collective public database. Biodivers Conserv 16:3609–3632. https://doi.org/10.1007/s10531-007-9150-1
Stegmann LF, Leitão RP, Zuanon J, Magnusson WE (2019) Distance to large rivers affects fish diversity patterns in highly dynamic streams of Central Amazonia. PLoS One 14:1–17. https://doi.org/10.1371/journal.pone.0223880
Stendera SES, Johnson RK (2005) Additive partitioning of aquatic invertebrate species diversity across multiple spatial scales. Freshw Biol 50:1360–1375. https://doi.org/10.1111/j.1365-2427.2005.01403.x
Stout J, Vandermeer J (1975) Comparison of species richness for stream-inhabiting insects in tropical and mid-latitude streams. Am Nat 109:263–280. https://doi.org/10.1086/282996
Strayer DL, Dudgeon D (2010) Freshwater biodiversity conservation: recent progress and future challenges. J North Am Benthol Soc 29:344–358. https://doi.org/10.1899/08-171.1
Tedesco PA, Oberdorff T, Lasso CA et al (2005) Evidence of history in explaining diversity patterns in tropical riverine fish. J Biogeogr 32:1899–1907. https://doi.org/10.1111/j.1365-2699.2005.01345.x
Thomaz AT, Malabarba LR, Bonatto SL, Knowles LL (2015) Testing the effect of palaeodrainages versus habitat stability on genetic divergence in riverine systems: study of a Neotropical fish of the Brazilian coastal Atlantic Forest. J Biogeogr 42:2389–2401. https://doi.org/10.1111/jbi.12597
Thompson R, Townsend C (2006) A truce with neutral theory: local deterministic factors, species traits and dispersal limitation together determine patterns of diversity in stream invertebrates. J Anim Ecol 75:476–484. https://doi.org/10.1111/j.1365-2656.2006.01068.x
Tisseuil C, Cornu JF, Beauchard O et al (2013) Global diversity patterns and cross-taxa convergence in freshwater systems. J Anim Ecol 82:365–376. https://doi.org/10.1111/1365-2656.12018
Tolonen KE, Leinonen K, Marttila H et al (2017) Environmental predictability of taxonomic and functional community composition in high-latitude streams. Freshw Biol 62:1–16. https://doi.org/10.1111/fwb.12832
Tolonen KE, Tokola L, Grönroos M et al (2016) Hierarchical decomposition of trait patterns of macroinvertebrate communities in subarctic streams. Freshw Sci 35:1032–1048. https://doi.org/10.1086/687966
Tomanova S, Goitia E, Helešic J (2006) Trophic levels and functional feeding groups of macroinvertebrates in neotropical streams. Hydrobiologia 556:251–264. https://doi.org/10.1007/s10750-005-1255-5
Tonkin JD, Altermatt F, Finn DS et al (2018) The role of dispersal in river network metacommunities: patterns, processes, and pathways. Freshw Biol 63:141–163. https://doi.org/10.1111/fwb.13037
Tonn WM, Magnuson JJ, Rask M, Toivonen J (1990) Intercontinental comparison of small-lake fish assemblages: the balance between local and regional processes. Am Nat 136:345–375. https://doi.org/10.1086/285102
Townsend CR (1989) The patch dynamics concept of stream community ecology. J North Am Benthol Soc 8:36–50. https://doi.org/10.2307/1467400
Townsend CR, Hildrew AG (1994) Species traits in relation to habitat templet for river systems. Freshw Biol 31:265–276. https://doi.org/10.1111/j.1365-2427.1994.tb01740.x
Townsend CR, Hildrew AG, Schofield K (1987) Persistence of stream invertebrate communities in relation to environmental variability. J Anim Ecol 56:597–613. https://doi.org/10.2307/5071
Turner JRG, Gatehouse CM, Corey CA (1987) Does solar energy control organic diversity? Butterflies, moths and the British climate. Oikos 48:195–205. https://doi.org/10.2307/3565855
Usseglio-Polatera P, Bournaud M, Richoux P, Tachet H (2000) Biological and ecological traits of benthic freshwater macroinvertebrates: relationships and definition of groups with similar traits. Freshw Biol 43:175–205
Verberk WCEP, Van Noordwijk CGE, Hildrew AG (2013) Delivering on a promise: integrating species traits to transform descriptive community ecology into a predictive science. Freshw Sci 32:531–547. https://doi.org/10.1899/12-092.1
Vieira NKM, Poff NL, Carlisle DM et al (2006) A Database of lotic invertebrate traits for North America. U.S. Geological Survey Data Series 187
Vinson MR, Hawkins CP (2003) Broad-scale geographical patterns in local stream insect genera richness. Ecography (Cop) 26:751–767. https://doi.org/10.1111/j.0906-7590.2003.03397.x
Walters AW, Post DM (2011) How low can you go? Impacts of a low-flow disturbance on aquatic insect communities. Ecol Appl 21:163–174. https://doi.org/10.1890/09-2323.1
Ward JV, Stanford JA (1983) The serial discontinuity concept of lotic ecosystems. In: Fontaine TI, Bartell JM (eds) Dynamics of lotic systems. Ann Arbor Science, Ann Arbor, MI, pp 29–42
Wilson DS (1992) Complex interactions in metacommunities, with implications for biodiversity and higher levels of selection. Ecology 73:1984–2000
Winemiller KO (2005) Life history strategies, population regulation, and implications for fisheries management. Can J Fish Aquat Sci 62:872–885. https://doi.org/10.1139/f05-040
Winemiller KO, Flecker AS, Hoeinghaus DJ (2010) Patch dynamics and environmental heterogeneity in lotic ecosystems. J North Am Benthol Soc 29:84–99. https://doi.org/10.1899/08-048.1
Winemiller KO, Rose KA (1992) Patterns of life-history diversification in North American fishes: implications for population regulation. Can J Fish Aquat Sci 49:2196–2218. https://doi.org/10.1139/f92-242
Woodward G, Bonada N, Brown LE et al (2016) The effects of climatic fluctuations and extreme events on running water ecosystems. Philos Trans R Soc B Biol Sci 371:20150274. https://doi.org/10.1098/rstb.2015.0274
Woodward G, Hildrew AG (2002) Food web structure in riverine landscapes. Freshw Biol 47:777–798. https://doi.org/10.1046/j.1365-2427.2002.00908.x
Woodward G, Jones JI, Hildrew AG (2002) Community persistence in Broadstone Stream (U.K.) over three decades. Freshw Biol 47:1419–1435. https://doi.org/10.1046/j.1365-2427.2002.00872.x
Wootton JT, Parker MS, Power ME (1996) Effects of disturbance on river food webs. Science 273:1558–1561. https://doi.org/10.1126/science.273.5281.1558
Wright DH (1983) Species-energy theory: an extension of species-area theory. Oikos 41:496–506. https://doi.org/10.2307/3544109
Zwick P (1992) Stream habitat fragmentation-a threat to biodiversity. Biodivers Conserv 1:80–97. https://doi.org/10.1007/BF00731036
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Copyright information
© 2021 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Allan, J.D., Castillo, M.M., Capps, K.A. (2021). Lotic Communities. In: Stream Ecology . Springer, Cham. https://doi.org/10.1007/978-3-030-61286-3_11
Download citation
DOI: https://doi.org/10.1007/978-3-030-61286-3_11
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-61285-6
Online ISBN: 978-3-030-61286-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)