
Abstract
The diverse Potamogeton genus includes over 80 species of aquatic macrophytes that occur across a broad geographic range and have variable response to environmental conditions. This study evaluated how environmental and spatial variables structure assemblage composition and species richness of Potamogeton spp. in the US states of Minnesota and Wisconsin. Variation partitioning analysis was used to study the relative contribution of local, climate and spatial variables in explaining assemblage composition and species richness. Models were also developed for sixteen Potamogeton spp. using partial linear regression. Assemblage composition and total species richness were better explained by the pure effects of spatial and local variables as compared to the pure effects of climate variables. However, geographical structuring of variables suggested that species followed a latitudinal gradient that was strongly related to eutrophication and partially related to climate. Models for individual species were similar although some were disproportionately described by specific categories of explanatory variables. For example, invasive Potamogeton crispus was more tolerant of eutrophication than most species and was also described by a strong spatial grouping of lakes near a large urban area. These results suggest that the distribution of Potamogeton spp. is limited by species tolerances to lake variation in local and climate characteristics across spatial gradients, whereas specific species may be more limited by dispersal barriers between lakes with suitable habitat. This analysis is the first regional evaluation of factors related to the distribution of this ecologically important genus and the importance of landscape-level approaches to ecological conservation is emphasized.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Species distributions are explained by local environmental conditions, historical characteristics and biotic factors, of which species dispersal has intrigued scientists for centuries (Levin 1992). Dispersal is generally described as the movement of species from one location to another across a landscape and it strongly affects how individuals, populations and communities are organised among habitats (Logue et al. 2011; Baguette et al. 2013). A suitable habitat for the individual species occurs as a network of idealized habitat patches, varying in area, degree of isolation and quality, and is surrounded by equally unsuitable habitats (Hanski 1998). Dispersal among habitats with different quality leads to colonization of new suitable areas and extirpations of populations from unsuitable habitats. Populations which are connected in a landscape by dispersal are regarded as metapopulations (Hanski 1991; 1998). For dispersal among connected communities the relative importance of environmental characteristics and dispersal in structuring metacommunities varies (Leibold et al. 2004; Heino 2011). Species sorting is primarily influenced by niche processes, although a minimum level of dispersal is needed for species to shift among suitable habitats. In the remaining three metacommunity perspectives (i.e., mass effects, neutral theory, and patch dynamics), dispersal or dispersal limitation are the most important forces explaining variation in metacommunities. Dispersal-related spatial processes can be particularly important in freshwater ecosystems which are distinct environments surrounded by an uninhabitable terrestrial matrix. Moreover, lake species are more dependent on dispersal compared to species in rivers, which are better connected through hydrologic networks (Heino 2011). However, recent studies have suggested that dispersal limitation between lakes and rivers is more likely related to biological groupings rather than differences in habitats (Padial et al. 2014; Alahuhta et al. 2015; Heino et al. 2015). Characterising the relative effects of forcing mechanisms of species dispersal could greatly inform understanding of ecological processes in aquatic systems.
Aquatic macrophytes are important biological components of lake communities and are considered good indicators of long-term changes of freshwater ecosystems. Aquatic macrophytes respond to reduced light availability, increased sedimentation and nutrient concentrations, and hydromorphological changes, often originating from anthropogenic activities at different temporal scales (Lachavanne et al. 1992; Padial et al. 2009; Bornette and Puijalon 2011; Beck et al. 2014). Moreover, macrophytes have an essential functional role in freshwater ecosystems, as they provide habitat and shelter, breeding areas, and food resources for other aquatic and terrestrial species (Carpenter and Lodge 1986; Schmidt et al. 2005). One of the most important and diverse genera of macrophytes is Potamogeton, which includes over 80 species with wide ranges covering almost all global freshwater systems (Cook 1990). Potamogeton spp. have variable responses across environmental conditions and vary in morphology between species (Crow et al. 2000; Vestergaard and Sand-Jensen 2000; Heegard et al. 2001; Chambers et al. 2008). Most Potamogeton spp. grow in alkaline waters, but some species, like P. amplifolius, P. natans, P. epihydrus, and P. gramineus, can favour more acidic conditions (Toivonen and Huttunen 1995; Crow et al. 2000; Vestergaard and Sand-Jensen 2000). Potamogeton amplifolius, P. praelongus, and P. robbinsii are often found in deeper parts of lakes (Chambers and Kalff 1985; Crow et al. 2000). The morphology of the Potamogeton taxa varies from emergent broad-leaved plants (e.g., P. illinoensis) to submerged thin-leaved species (e.g., P. diversifolius). In addition, vegetative and reproductive morphology varies considerably across the taxon (Crow et al. 2000). Although knowledge of Potamogeton species responses to local environmental conditions is relatively well-documented, little is known about how distribution of these species is affected by dispersal across the landscape.
The influence of water quality and climate in explaining aquatic plant distributions has rarely been studied within the same work (but see Alahuhta et al. 2011; Kosten et al. 2011), whereas the effects of local environmental conditions are relatively well known. Aquatic plants, and more importantly submerged species, are strongly affected by local water quality, but climate also contributes to species distributions at broad spatial scales (Alahuhta et al. 2011; Netten et al. 2011). Moreover, aquatic plants are often investigated at small spatial scales (Jones et al. 2003; Saarneel et al. 2011) where climate has only a marginal effect on species distributions. This study examines the relative roles of local variables, climate, and geographic location in explaining the distribution of Potamogeton taxa in 214 lakes across the states of Minnesota and Wisconsin, USA. The aim was to study how assemblage composition and species richness respond to the three different groups of ecological variables (local, climate, and space), having implications for understanding dispersal mechanisms in landscape or regional contexts. The relationship between sixteen species and the three groups of explanatory variables was also examined to identify species-level trends separate from the whole community. Similar analyses have been used to examine macrophyte assemblage composition in glacial lakes (Mikulyuk et al. 2011; Beck et al. 2013; Alahuhta et al. 2013) but have not been used specifically to evaluate the diverse Potamogeton genus. As such, the results presented herein are relevant for understanding drivers of overall assemblage structure of Potamogeton taxa, including the most common species in this genus that occur in the upper Midwest United States and regions with comparable climatic and geological characteristics.
Materials and methods
Biological surveys of Potamogeton taxa from 214 lakes were used, covering the US states of Minnesota and Wisconsin (Fig. 1). The Minnesota and Wisconsin Departments of Natural Resources (MNDNR, WDNR) have collected macrophyte data using the point intercept method beginning in the early 2000 s (Madsen 1999). All macrophyte species in each lake were surveyed in a grid design of evenly-spaced points throughout the littoral zone. Species were sampled during the growing season at each point by boat using a grapple that was sunk to the bottom and retrieved to identify species present. Early season surveys that only targeted P. crispus were not used. Data for each lake included total richness and frequency occurrence of individual species using the total number of survey points for which each Potamogeton species was found, scaled by total effort. Sampling effort was set at a point density (i.e., number of points per littoral hectare) that was sufficient to capture all but the most rare species (Mikulyuk et al. 2010; Beck et al. 2014).

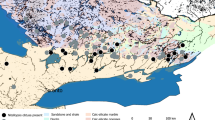
Potamogeton species richness (top) for each study lake (n = 214) and geographic centers (bottom) for each species in Minnesota and Wisconsin. County boundaries (lines) and shading for each level III ecoregion (Omernik 1987) are also shown. Ecoregions with study lakes included North Central Hardwood Forests (NCHF), Northern Glaciated Plains (NGP), Northern Lakes and Forests (NLF), and Western Cornbelt Plains (WCBP). Geographic centers for each species were estimated as the average latitude/longitude weighted by species occurrence at each lake (number of points in a lake where a species was found divided by total survey points). Point sizes for the bottom plots are the average frequency occurrences across lakes. The richness legend applies only to the top plot. PA: P. amplifolius, PC: P. crispus, PE: P. epihydrus, PF: P. foliosus, PG: P. gramineus, PI: P. illinoensis, PN: P. natans, POP: P. praelongus, POPU: P. pusillus, POR: P. richardsonii, POS: P. spirillus, PP: P. pectinatus, PR: P. robbinsii, PS: P. strictifolius, and PZ: P. zosteriformis
Study lakes occurred across a spatial gradient of land use, climate, and morphometry such that the dataset included a variety of lake types. Lakes were situated in four ecoregions of the upper Midwest United States (level III, Omernik 1987), including the Northern Lakes and Forests ecoregion in the north, the North Central Hardwood Forests ecoregion in central areas, and the Northern Glaciated Plains and Western Cornbelt Plains of southern Minnesota (Fig. 1). In general, lake productivity decreases from the south to the north across the ecoregions, whereas overall richness is generally highest in moderately productive or mesotrophic lakes in central regions of each state.
The explanatory variables used to characterize Potamogeton distributions were grouped into three categories: local, climate, and spatial. Local variables (water quality and morphometry) included alkalinity concentration (mg/L of CaCO3), colour (Pt–Co units), lake area (km2), maximum depth (m), perimeter (km), Secchi depth (m), and total phosphorus (mg/L). The water quality variables (alkalinity, colour, Secchi depth, and total phosphorus) were obtained from three sources: MNDNR division of fisheries water quality data (http://www.dnr.state.mn.us/lakefind/), the STORET database maintained by the United States Environmental Protection Agency (http://www.epa.gov/waterdata/storage-and-retrieval-and-water-quality-exchange), and the Wisconsin Lake Historical Limnological Parameters database (Papes and Vander Zanden 2010). The lake morphometry variables were obtained from the same sources or supplemented with Geographical Information System (GIS) databases (MN: https://gisdata.mn.gov/, WI: http://www.sco.wisc.edu/find-data.html). The climate variables for each lake based on geographic location included annual mean temperature (°C), maximum temperature of the warmest month (°C), minimum temperature of the coldest month (°C), annual precipitation (mm), and lake altitude (m.a.s.l.). The climate variables were derived from the Worldclim database (Hijmans et al. 2005).
Spatial variables describing variation in location were derived from the Cartesian coordinates of geographic centers of each lake (North American Datum 1983). Following similar methods in Alahuhta et al. (2013), Principal Coordinates of Neighbor Matrices (PCNM) analysis was used to deconstruct the lake locations into orthogonally and linearly uncorrelated components (Borcard and Legendre 2002). PCNM is a special form of Moran’s Eigenvector Map (MEM) functions that quantifies the spatial autocorrelation among geographic features based strictly on location and proximity between features. Significant spatial variation shown in PCNMs can indicate environmental autocorrelation, dispersal limitation, or historical effects on species distributions (Dray et al. 2012). The eigenvectors for PCNM are derived from a Euclidean distance matrix for all locations which is then truncated by the longest distance in the minimum spanning tree linking all sites on the map. Principal Coordinates Analysis of the truncated neighbor matrix produces eigenfunctions for all eigenvectors, which are in turn described by Moran’s I values that quantify spatial autocorrelation. Eigenfunctions with positive Moran’s I values were retained to describe variation among locations related to positive spatial autocorrelation. In general, this analysis can be used to describe a range of geographic patterns of spatial variation, with the first few eigenvectors describing large-scale spatial variation and the remaining describing finer-scale variation. The use of PCNM eigenvectors in statistical models provides a means to assess variation among biological communities across the landscape as explained strictly by physical location and relative to additional variables (e.g., climate or local). PCNM analysis was conducted using the PCNM package (Legendre et al. 2013) for the R statistical computing environment (R Core Team (RCT) 2015).
Statistical analysis
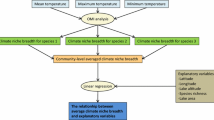
The variation partitioning procedure provided by the varpart function in the vegan package for R (Oksanen et al. 2015) was used to evaluate effects of local, climate, and spatial variables on the Potamogeton data. Partial redundancy analysis (pRDA) was used to evaluate variation in the assemblage composition across lakes (i.e., counts of occurrence of all Potamogeton species by lake) and partial linear regression (pLR) was used to evaluate variation in total species richness (log-transformed) of Potamogeton spp. and frequency occurrence of sixteen individual species. Frequency occurrence data were arcsine-square root-transformed to use linear multivariate methods and Hellinger-transformed to minimize the effect of zeros (i.e., species absence, Legendre and Gallagher 2001). The most important explanatory variables among each category (local, climate, spatial) were selected prior to variation partitioning to reduce model complexity and to avoid type I error. Variable selection followed a standard forward selection technique with stopping criteria if the inclusion of an additional variable in a model produced a probability value less than the selected alpha-level. The significance of a variable was based on a Monte Carlo permutation test of the model residuals following the default procedure (999 permutations, α = 0.05) in the forward.sel function in the packfor package for R (Dray et al. 2013). The selected variables in each category were used to create combined models with all categories for variation partitioning. Total variation of the response (assemblage composition, total richness, occurrence of individual species) was decomposed into separate fractions of: (1) pure local, (2) pure climate, (3) pure spatial, (4) shared local/climate, (5) shared local/spatial, (6) shared climate/spatial, (7) shared all categories, and (8) unexplained (1-total explained) (Fig. 2, Anderson and Gribble 1998). Detailed information on the variation partitioning process is given in Legendre et al. (2005) and Borcard et al. (2011).

Conceptual representation of variation partitioning analyses used to estimate explained variance of Potamogeton assemblage composition, total richness, and frequency occurrence of individual species. Categories of explanatory variables for each lake included local, climate, and spatial information. Partial redundancy analysis and partial linear regression models were used to explain total variation (area covered by all circles), pure variation (non-overlapped local, climate, or spatial areas), shared variation (overlapping areas between categories or all three), and unexplained (total area–circle area). Additional details are in Legendre et al. (2005) and Borcard et al. (2011)
The variation partitioning was based on adjusted R2 which provided unbiased estimates of the explained variation of the modelled response variables by individual and combined categories of explanatory variables (Fig. 2, Peres-Neto et al. 2006). The number of explanatory variables is also taken into account in the adjusted R2 values, which allows the different models to be compared to one another (Blanchet et al. 2008). The use of adjusted R2 values often results in a decreased percentage of explained variation, that is, it generates a considerable amount of unexplained variation due to the high degree of stochasticity in species distributions (Capers et al. 2010; Alahuhta and Heino 2013).
Results
Potamogeton distribution and lake characteristics
Abundance and distribution of Potamogeton spp. varied considerably across the study region (Fig. 1; Table 1). Species richness among lakes was generally highest in the North Central Hardwood Forests and Northern Lakes and Forests Ecoregions. The most abundant species among ecoregions was P. pectinatus, except in the Northern Lakes and Forest ecoregion which was dominated by P. zosteriformis (Table 1). Lake characteristics also varied with distinct differences among the local and climate categories (Table 2). In general, local and climate variables were more strongly related to longitudinal and latitudinal gradients as compared to non-monotonic differences between ecoregions and states.
Spatial groupings of lakes through analysis of geographic coordinates with PCNM produced 38 axes with positive Moran’s I scores significantly different than zero (α = 0.05). As described below, selection of spatial axes during initial model development selected some axes more often than others (Fig. 3). Broad-scale variation in spatial groupings described by the first and second axes characterized variation in lake locations in central Minnesota, with the first axis describing a grouping in the Northern Lakes and Forests ecoregion and the second describing a grouping in the North Central Hardwood Forests ecoregion (Fig. 3). The fifth axis included central groupings similar to the first and second axes but also included a Wisconsin grouping in the north–central region of the state. Spatial variation described by the ninth axis was less clear and appeared to describe spatial groupings not captured by the other three axes.

Scores for selected axes from principal components of neighbouring matrices. The axes shown were the four most frequently retained during variable selection of models to explain Potamogeton distribution. Points for each lake are scaled relative to axis scores to show spatial autocorrelation from geographic location. The Moran’s eigenvectors (top) are a combined measure of spatial autocorrelation described by each axis. See Fig. 1 for the ecoregion labels
Assemblage composition and species richness
Variation partitioning with pRDA and pLR for assemblage composition and total Potamogeton richness, respectively, indicated that local, climate, and spatial variables explained a total of 32 and 46 % of the variation in each (Fig. 4; Table 3). For assemblage composition, initial variable selection by category in order of decreasing importance (Table 4, based on adjusted R2 for each variable, p < 0.05 for all) was alkalinity, Secchi depth, total phosphorus, and lake area for local variables; maximum temperature, precipitation, lake altitude, mean temperature, and minimum temperature for climate variables; and twenty-one spatial axes from PCNM (axes one and two were most important). Variation of assemblage composition explained by category (Fig. 4; Table 3) indicated that the pure effect of spatial variables (5.9 %) exceeded the pure effects of climate (0.6 %) and local variables (3.4 %). The joint effects of variable categories suggested that climate and space had the largest shared variation (5.9 %), whereas variation in assemblage composition explained by all three categories was 12.4 %. For total richness, significant local variables were maximum lake depth, Secchi depth, and total phosphorus (Table 4). Linear models for the parameter estimates for richness indicated a positive association with lake depth (p < 0.0001) and Secchi depth (p < 0.05), whereas a negative association was observed with total phosphorus (p < 0.0001). For the climate variables, a negative association with minimum temperature (p < 0.0001) and a positive association with precipitation (p < 0.05) were observed with richness. Ten of the thirty-eight spatial variables were selected for richness. Variation of total richness explained by each category indicated that the pure effects of spatial variables (12.6 %) were largest, although pure local effects explained a comparable amount of the variation (11.8 %) (Fig. 4; Table 3). Likewise, the joint effects (two-way and all three categories) were smaller compared to the pure effect of local or spatial variables, although shared variation between all three categories was 10.9 %.

Fractions of variation explained by local, climate, and spatial variables for Potamegeton assemblage composition, richness, and individual species. The top plot shows total explained variance (sum of pure and shared effects), the middle shows pure variation explained by each variable category, and the bottom shows shared fractions of variation for different combinations of the variable categories. See Fig. 2 for a conceptual representation of the fractions of variation
Species level
The combined dataset for Minnesota and Wisconsin included 25 Potamogeton spp., of which only sixteen were sufficiently abundant to create regression models for variation partitioning. Species present but not modelled included P. alpinus, P. bicupulatus, P. filiformis, P. nodosus, P. oakesianus, P. obtusifolius, and P. vaseyi (n = 1–7 lakes). For the remaining species, total explained variance ranged from 66.4 % (P. pectinatus) to 14.7 % (P. richardsonii) with an average of 31.7 % for all species (Fig. 4; Table 3). For the pure effects of each variable category, local effects ranged from 11.0 % (P. illinoensis) to <1 % explained variance (P. pusillus, P. strictifoloius, P. zosteriformis), climate effects ranged from 2.3 % (P. epihydrus) to <1 % (numerous spp.), and spatial effects ranged from 19.9 % (P. crispus) to 2.5 % (P. amplifolius). Within the joint effects, explained variance that was shared between categories was generally lowest for local plus climate effects (average 0.4 % for all species), whereas climate plus space and local plus space had similar averages (both ~4 %). Average shared explained variance among all three categories was relatively large (10.2 %), suggesting species response to shared effects was better explained by covariation among all explanatory variables rather than shared effects between pairwise categories.
An evaluation of variables within each category provided additional information on drivers of species occurrence (Table 4; Figs. 5, 6, 7). For local variables, alkalinity, Secchi depth, and total phosphorus were most commonly selected from forward selection (Table 4). Species that included these variables typically showed negative associations with alkalinity and total phosphorus and positive associations with Secchi depth. These trends were reversed for some species, most notably for P. crispus and P. pectinatus. For climate variables, lake altitude and precipitation were most commonly selected. Species associations with climate variables did not show any regular patterns with the exception of temperature variables, such that species were most often negatively associated with temperature. However, P. crispus and P. pectinatus had positive associations with maximum temperature. Selection of spatial variables typically did not include more than a few axes, suggesting variation by location was explained primarily by the first few spatial axes. However, some species had models with a relatively high number of axes, including P. crispus, P. pectinatus, P. praelongus, and P. robbinsii.



Redundancy analysis biplots for Potamogeton spp. relative to spatial variables from principal coordinates of neighbouring matrices (PCNM). The PCNM axes used in the figure were the top ten most frequent axes that were retained after variable selection for individual species models. See Fig. 1 for species abbreviations
Discussion
Assemblage composition and species richness
Of the pure fractions of variation, assemblage composition and species richness of Potamogeton spp. were most strongly explained by spatial variables (Table 3). The relatively high explained variation due to pure spatial effects was expected given known relationships between diversity and dispersal patterns across a landscape. However, a more compelling explanation for observed patterns of species distributions is the geographical structuring of local and climate variables across space. The models included many important environmental variables that structure macrophyte distributions (Vestergaard and Sand-Jensen 2000; Jones et al. 2003; Mikulyuk et al. 2011; Alahuhta et al. 2013; Beck et al. 2013) and the notable difference in the pure effects of local and climate variables relative to the shared effects with spatial variables provided evidence that geographic variation in environmental variables was important. For assemblage composition, the joint effects of climate and space were equally high compared to the highest pure effects of individual categories. Similarly, the joint effects of climate and space for total richness were comparable to those for assemblage composition, although the pure effects of space and local variables were higher compared to assemblage composition. This finding indicates that climate variables are geographically structured, related to the decreasing gradient with latitude in climate (Beck et al. 2013; Alahuhta 2015). The shared effects of all three variable groups combined were also high for assemblage composition and species richness, indicating that their individual influences cannot be statistically distinguished. Similar results were found by Beck et al. (2013) where a majority of variation of a macrophyte-based index of biotic integrity for Minnesota was explained by shared effects of environmental and anthropogenic variables, rather than the pure effects of each. Mikulyuk et al. (2011) also found that spatial variables contributed a relatively high amount of explained variation for macrophyte communities in Wisconsin. They suggested that habitat limitation related to strong latitudinal gradients may explain the strong effects of spatial variables. Although, many Potamogeton spp. favour more nutrient- and alkaline-rich waters, the geographic centers were located in the middle of Minnesota and northern Wisconsin where lakes are more mesotrophic (Fig. 1). Thus, Potamogeton assemblage composition and species richness is defined by habitat suitability related to nutrient levels, where excessive phosphorus concentrations reduce suitable habitat in the southern region of the study area.
Pure effects of local environmental variables also explained a significant but smaller amount of variation for assemblage composition and species richness as compared to pure spatial effects (Table 3). All local variables were significantly related to Potamogeton assemblage composition, whereas only maximum depth and total phosphorus were correlated with species richness (Table 4). However, the variation in total richness that was explained by depth and phosphorus was much higher than the pure effects of all local variables on assemblage composition. The positive association between species richness and maximum depth is likely related to habitat availability, as habitat heterogeneity increases with an increasing depth gradient (Tolonen et al. 2001; Alahuhta et al. 2013). Increasing lake depth has also been associated with increases in biotic integrity of macrophytes in glacial lakes (Beck et al. 2010). Increasing lake depth and species richness could also be linked to effects of wave exposure. Macrophytes in deeper lakes are less susceptible to uprooting from wave action during high wind events (Riis and Hawes 2003). Turbidity in shallow lakes is also influenced by wind, which could indirectly limit macrophyte growth by light scattering. The negative influence of total phosphorus on species richness further emphasizes that the nutrient status of many lakes exceeds levels for sustaining Potamogeton spp. Similar results related to latitudinal gradients in water quality have been found for species richness of all submerged macrophytes in Midwest glacial lakes of the United States (Beck et al. 2014; Alahuhta 2015). Interestingly, neither alkalinity nor Secchi depth were related to Potamogeton species richness, although both of these variables can affect growth patterns of submerged vascular plants through carbon and light availability, respectively (Chambers and Kalff 1985; Madsen et al. 1996). Moreover, lake area was not associated with richness, which is contrary to established relationships between the two (e.g., MacArther and Wilson 1967). Lake depth is correlated with lake size for the study lakes and post hoc comparisons showed that depth and size were both positively correlated with richness, with the former having a stronger association. The variable selection procedure used in the analysis identified the most parsimonious model that maximized explanatory power and minimized redundancy among variables. Although lake area is related to richness, it was likely not selected given the greater increase in explained variability with maximum depth.
Lastly, individual climate variables were also correlated to the distribution of Potamogeton communities. Previous studies have similarly reported that local water quality and morphometric conditions are often more important than climate for aquatic macrophytes at spatial scales similar to our analysis (Santamaría 2002; Chappuis et al. 2014; Alahuhta 2015). For assemblage composition, all individual climate variables were significant for Potamogeton spp., whereas species richness was only positively correlated with annual precipitation and negatively with minimum temperature of the coldest month (Table 4). Higher annual precipitation has been correlated to higher nutrients and suspended solids that are transported from terrestrial sources during intense rainfalls (Cobbaert et al. 2014). However, this contradicts our results that showed a negative association between species richness and total phosphorous. Annual precipitation can function as a proxy for water-induced dispersal as plant propagules are more easily transported through stream networks during high flows (Riis 2008), which may explain the positive association observed from the model. The increase in Potamogeton species richness with decreasing winter temperature was unexpected given previous descriptions of temperature and aquatic plant richness (e.g., Pip 1989). However, Pip (1989) argue that temperature in itself is a poor predictor and the relationship with richness is likely related to interactions with other variables that influence macrophyte distribution. The relationship between temperature and richness may have also been poorly described with a linear model as the response is not monotonic across the gradient (i.e., species maxima at moderate temperatures). For example, Beck et al. (2014) used additive models to describe non-linear relationships between macrophyte indicators of community health and climate variables. Species richness showed a distinct modal response to increasing growing degree days measured at each lake. Therefore, we argue that macrophyte communities in the northern region of our study area are in fact limited by climate despite a positive association of richness with increasing minimum temperatures. Harsh winter conditions are known to restrict macrophyte growth through thick ice cover limiting the availability of carbon, oxygen, and light, freezing bottom sediments, or increasing ice erosion (Lind et al. 2014). Alahuhta (2015) and Johnson et al. (2010) have described a similar gradient in species richness of all macrophyte taxa in the Midwest United States.
Species level
Similar conclusions about the effects of local, climate, and spatial variables for assemblage composition and total richness can be generalized for individual species. Overall, the pure effects of spatial and local variables were much larger than climate effects and the combined effects of variable categories were generally larger than the pure effects for any given species (Table 3). The latter conclusion was particularly true for effects shared between all categories, local plus space, and climate plus space. However, some differences between species were observed that potentially explains geographic variation in the relative distributions of each. For example, the total explained variation of separate models for P. pectinatus, P. pusillus, and P. crispus exceeded the total explained variation for assemblage composition and total richness. The geographic centers of each species in relation to the spatial gradients among the explanatory variables provided a potential explanation for the relatively high amount of explained variation of the models. Both P. crispus and P. pectinatus were centered more closely than the other species to the southern area of Minnesota where lakes are more nutrient-rich, alkaline, and warmer. For P. pectinatus, local variables were also geographically patterned, likely in the southern lakes. Both species are commonly regarded as tolerant of eutrophic conditions (Beck et al. 2010) and relatively intolerant of harsh climate conditions.
Valley and Heiskary (2012) provide evidence that P. crispus in Minnesota may be light-limited during harsh winters by thicker snow depth on frozen lakes. The relationship between P. crispus and snow cover is important for understanding the competitive advantages of this introduced species. Unlike native Potamogeton spp., seasonal growth of curly-leaf pondweed begins before ice-off from turions in the sediment that were deposited by mature plants the year prior. Early growth provides an advantage over native species that begin growth later in the spring. Therefore, light limitation from heavy snow cover can reduce growth of curly-leaf pondweed early in the season and release native species from competitive pressures. As such, geographic climate variation and the positive association with maximum temperature suggests that P. crispus is likely restricted in the northern lakes by climate. However, future expansion north may be mediated by warmer winter conditions associated with climate change as the snow depth on frozen lakes may change.
Curly-leaf pondweed is the only invasive species in the genus that occurs in the study region and was likely introduced to the region in the early 1900 s (Valley and Heiskary 2012). The species occurs in over 700 lakes in the region, although its abundance varies. P. crispus often dominates macrophyte communities in shallow, turbid-water lakes in southern Minnesota and is a nuisance species that affects recreation in heavily-used lakes near urban centers. As such, the largest pure effect for P. crispus was attributed to spatial variables, suggesting that lake location or proximity was an important factor explaining distribution. Interestingly, the second spatial axis from PCNM had the strongest correlation to the distribution of P. crispus. This axis explained spatial correlation among lakes in the North Central Hardwood Forests ecoregion of central Minnesota, which is also near a large metropolitan area (Fig. 3). Species transport between lakes with heavy recreational use in urban areas is a well-known vector of exotic species invasion (e.g., Miro and Ventura 2013). The importance of spatial groupings among lakes for P. crispus provides further support of human-aided transport as a means of dispersal in the Midwest United States.
Other species also had disproportionately large amounts of variation explained by specific categories of the explanatory variables. As noted above, P. pusillus had a large amount of total explained variation compared to the remaining species. Although most of this variation was from shared effects, a large percentage of pure variation was explained by spatial variables. The geographic center of P. pusillus was located in northern Wisconsin and was well-described by the ninth spatial axis that characterized a lake grouping in that region (Figs. 3, 7). Additionally, the influence of climate (pure fraction and geographically-structured climate effect) was large for P. pusillus relative to the remaining species. The climate variables of lake altitude and precipitation were positively correlated with the distribution of P. pusillus, which suggests that the species is more commonly found in lakes that are higher in the hydrologic network and that receive more precipitation. These lakes likely have variable water levels as they fill from precipitation and drain more quickly than lakes lower in the watershed. Some studies have suggested that P. pusillus is an early colonizer of habitats affected by hydrological alteration (Boedeltje et al. 2001; Van Geest et al. 2005), which may explain the strong association of the species with the climate variables. Local variables also explained a disproportionate amount of pure effects for other species, particularly P. amplifolius and P. illinoensis. Both species were correlated with Secchi depth and lower total phosphorus, suggesting relative intolerance of eutrophic conditions. Both species were related to alkalinity, although P. amplifolius had a negative association and P. illinoensis had a positive association. These associations are supported by geographical structuring of local variables (local/space) and especially a strong correlation with alkalinity. Geographic centers of each species in Fig. 1 show P. amplifolius further east from P. illinoensis, coincident with a decrease in alkalinity from southern and western Minnesota to northern Wisconsin.
Conclusions
This study provided a unique decomposition of factors that influence the distribution of Potamogeton spp. in glacial lakes of the Midwest United States. To our knowledge, no studies have evaluated the contributing factors of local, spatial, and climate variables on the distribution of this species-rich and morphologically-diverse genus. Variation partitioning analyses revealed that assemblage composition and total species richness were best explained by spatial groupings of the study lakes, particularly lake groups along a strong latitudinal gradient. Further evaluation suggested that the pure effects of spatial variables potentially described dispersal limitations as lakes closer in space were more similar in species composition. More importantly, shared variation between spatial groupings and environmental factors described limitations in habitat suitability related to eutrophication in southern lakes such that most Potamogeton spp. in the analysis were unable to colonize high-nutrient lakes. An additional, but minor, confounding effect described a potential climate limitation along the latitudinal gradient such that most species were unable to colonize northern regions of the study area. Accordingly, the geographic centers shown in Fig. 1 represent a tradeoff in habitat suitability related to geographical structuring of environmental variables. Models for individual species generally supported the results from the community models, with the exception of some species that were disproportionately described by specific categories of variables. For example, the invasive species P. crispus was strongly related to both eutrophication and spatial variables. This suggests a higher tolerance to elevated nutrient levels and a mechanism for dispersal between lakes, respectively, to provide an explanation for the invasive spread of the species in the region. Overall, these results provide support that the latitudinal gradient is partially based on climatic differences, whereas land-use changes along this gradient have further affected water quality in the southern parts of the states. A similar gradient that has been steepened by anthropogenic activities has been reported for wetland plant species in the Great Lakes region (Johnson et al. 2010).
Overall, this analysis provides an argument that the management and conservation of this important genus could focus on drivers of assemblage composition that are spatially aggregated across the landscape, in addition to traditional management efforts that focus on local characteristics related to eutrophication. Management efforts for individual species could be similar but dependent on lake characteristics and their variation across the landscape relative to the species of interest. Dispersal limitation also has relevance for restoration efforts such that connectivity between lakes should be sufficient for colonization provided that habitat is suitable. Planting native species in suitable habitats may have minimal lasting effect if lakes are separated by large distances across the landscape. Finally, considering the integration of Potamogeton spp. within the larger community of macrophytes and other biota may also be important for developing a landscape-level understanding of factors that drive variation in the entire community, having implications for regional efforts of lake management and conservation.
References
Alahuhta J (2015) Geographic patterns of lake macrophyte communities and species richness at regional scales. J Veg Sci 26:564–575
Alahuhta J, Heino J (2013) Spatial extent, regional specificity and metacommunity structuring in lake macrophytes. J Biogeogr 40:1572–1582
Alahuhta J, Heino J, Luoto M (2011) Climate change and the future distributions of aquatic macrophytes across boreal catchments. J Biogeogr 38:383–393
Alahuhta J, Kanninen A, Hellsten S, Vuori KM, Kuoppala M, Hämäläinen H (2013) Environmental and spatial correlates of community composition, richness and status of boreal lake macrophytes. Ecol Indic 32:172–181
Alahuhta J, Rääpysjärvi J, Hellsten S, Kuoppala M, Aroviita J (2015) Species sorting drives variation of boreal lake and river macrophyte communities. Commun Ecol 16:76–85
Anderson MJ, Gribble NA (1998) Partitioning the variation among spatial, temporal and environmental components in a multivariate data set. Aust J Ecol 23:158–167
Baguette M, Blanchet S, Legrand D, Stevens VM, Turlure C (2013) Individual dispersal, landscape connectivity and ecological networks. Biol Rev 88:310–326
Beck MW, Hatch LK, Vondracek B, Valley RD (2010) Development of a macrophyte-based index of biotic integrity for Minnesota lakes. Ecol Indic 10:968–979
Beck MW, Vondracek B, Hatch LK (2013) Environmental clustering of lakes to evaluate performance of a macrophyte index of biotic integrity. Aquat Bot 108:16–25
Beck MW, Tomcko CM, Valley RD, Staples DF (2014) Analysis of macrophyte indicator variation as a function of sampling, temporal, and stressor effects. Ecol Indic 46:323–335
Blanchet FG, Legendre P, Borcard D (2008) Forward selection of explanatory variables. Ecography 89:2623–2632
Boedeltje G, Smolders AJP, Roelofs JGM, Van Groenendael JM (2001) Constructed shallow zones along navigation canals: vegetation establishment and change in relation to environmental characteristics. Aquat Conserv 11(6):453–471
Borcard D, Legendre P (2002) All-scale spatial analysis of ecological data by means of principal coordinates of neighbour matrices. Ecol Model 153:51–68
Borcard D, Gillet F, Legendre P (2011) Numerical ecology with R. Springer, New York
Bornette G, Puijalon S (2011) Response of aquatic plants to abiotic factors: a review. Aquat Sci 73:1–14
Capers RS, Selsky R, Bugbee GJ (2010) The relative importance of local conditions and regional processes in structuring aquatic plant communities. Freshw Biol 55:952–966
Carpenter SR, Lodge DM (1986) Effects of submersed macrophytes on ecosystem processes. Aquat Bot 26:341–370
Chambers PA, Kalff J (1985) Depth distribution and biomass of submerged aquatic macrophyte communities in relation to Secchi depth. J Fish Aquat Sci 42:701–709
Chambers PA, Lacoul P, Murphy KJ, Thomaz SM (2008) Global diversity of aquatic macrophytes in freshwater. Hydrobiologia 595:9–26
Chappuis E, Gacia E, Ballesteros E (2014) Environmental factors explaining the distribution and diversity of vascular aquatic macrophytes in a highly heterogeneous Mediterranean region. Aquat Bot 113:72–82
Cobbaert D, Wong A, Bayley SE (2014) Precipitation-induced alternative regime switches in shallow lakes of the Boreal Plains (Alberta, Canada). Ecosystems 17(3):535–549
Cook CD (1990) Aquatic plant book. The Hague, Netherlands
Crow G, Hellquist C, Fassett N (2000) Aquatic and wetland plants of Northeastern North America, volume 2, Angiosperms: Monocotyledons [e-book]. University of Wisconsin Press, Ipswich
Dray S, Pélissier R, Couteron P, Fortin MJ, Legendre P, Peres-Neto PR, Bellier E, Bivand R, Blanchet FG, De Cáceres M, Dufour AB, Heegaard E, Jombart T, Munoz F, Oksanen J, Thioulouse J, Wagner HH (2012) Community ecology in the age of multivariate multiscale spatial analysis. Ecol Monogr 82:257–275
Dray S, Legendre P, Blanchet G (2013) packfor: Forward selection with permutation (Canoco p. 46). R package version 0.0-8/r109. https://R-Forge.R-project.org/projects/sedar/. Accessed Dec 2015
Hanski I (1991) Metapopulation dynamics: brief history and conceptual domain. Biol J Linn Soc 42:3–16
Hanski I (1998) Metapopulation dynamics. Nature 396:41–49
Heegard E, Birks HH, Gibson CE, Smith SJ, Wolfe-Murphy S (2001) Species-environmental relationships of aquatic macrophytes in Northern Ireland. Aquat Bot 70:175–223
Heino J (2011) A macroecological perspective of diversity patterns in the freshwater realm. Freshw Biol 56:1703–1722
Heino J, Soininen J, Alahuhta J, Lappalainen J, Virtanen R (2015) A comparative analysis of metacommunity types in the freshwater realm. Ecol Evol 5:1525–1537
Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A (2005) Very high resolution interpolated climate surfaces for global land areas. Int J Climatol 25:1965–1978
Johnson CA, Zedler JB, Tulbure MG (2010) Latitudinal gradient of floristic condition among Great Lakes coastal wetlands. J Gt Lakes Res 36:772–779
Jones JI, Li W, Maberly SC (2003) Area, altitude and aquatic plant diversity. Ecography 26:411–420
Kosten S, Jeppesen E, Huszar VLM, Mazzeo N, van Nes EH, Peeters ETHM, Scheffer M (2011) Ambiguous climate impacts on competition between submerged macrophytes and phytoplankton in shallow lakes. Freshw Biol 56:1540–1553
Lachavanne JB, Perfetta J, Juge R (1992) Influence of water eutrophication on the macrophytic vegetation of Lake Lugano. Aquat Sci 54:351–363
Legendre P, Gallagher ED (2001) Ecologically meaningful transformations for ordination of species data. Oecologia 129:271–280
Legendre P, Borcard D, Peres-Neto PR (2005) Analyzing beta diversity: partitioning the spatial variation of community composition data. Ecol Monogr 75:435–450
Legendre P, Borcard D, Blanchet FG, Dray S (2013) PCNM: NEM spatial eigenfuction and principal coordinate analyses. R package version 2.1-2/r109. https://www.R-project.org/. Accessed Dec 2015
Leibold MA, Holyoak M, Mouquet N, Amarasekare P, Chase JM, Hoopes MF, Holt RD, Shurin JB, Law R, Tilman D, Loreau M, Gonzalez A (2004) The metacommunity concept: a framework for multi-scale community ecology. Ecol Lett 7:601–613
Levin SA (1992) The problem of pattern and scale in ecology. Ecology 73:1943–1967
Lind L, Nilsson C, Polvi LE, Weber C (2014) The role of ice dynamics in shaping vegetation in flowing waters. Biol Rev 89:791–804
Logue JB, Mouquet N, Peter H, Hillebrand H, The Metacommunity working group (2011) Empirical approaches to metacommunities: a review and comparison with theory. Trends Ecol Evol 26:482–491
MacArther RH, Wilson EO (1967) The theory of island biogeography. Princeton University Press, New Jersey
Madsen JD (1999) Point intercept and line intercept methods for aquatic plant management. Technical Report TN APCRP-M1-02, APCRP Technical notes collection. U.S. Army Engineer Center, Vicksburg
Madsen TV, Maberly SC, Bowes G (1996) Photosynthetic acclimation of submersed angiosperms to CO2 and HCO3 −. Aquat Bot 53:15–30
Mikulyuk A, Hauxwell J, Rasmussen P, Knight S, Wagner KI, Nault ME, Ridgely D (2010) Testing a methodology for assessing plant communities in temperate inland lakes. Lake Reserv Manag 26(1):54–62
Mikulyuk A, Sharma S, Van Egeren S, Erdmann E, Nault ME, Hauxwell J (2011) The relative role of environmental, spatial, and land-use patterns in explaining aquatic macrophyte community composition. Can J Fish Aquat Sci 68:1778–1789
Miro A, Ventura M (2013) Historical use, fishing management and lake characteristics explain the presence of non-native trout in Pyrenean lakes: Implications for conservation. Biol Conserv 167:17–24
Netten JJC, van Zuidam J, Kosten S, Peeters ETHM (2011) Differential response to climatic variation of free-floating and submerged macrophytes in ditches. Freshw Biol 56:1761–1768
Oksanen J, Blanchet FG, Kindt R, Legendre P, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Henry M, Stevens H, Wagner H (2015) Vegan: Community Ecology Package. R package version2.3-2. https://CRAN.R-project.org/package=vegan. Accessed Dec 2015
Omernik JM (1987) Ecoregions of the conterminous United States. Ann Assoc Am Geogr 77:118–125
Padial AA, Carvalho P, Thomaz SM, Boschilia SM, Rodrigues RB, Kobayashi JT (2009) The role of an extreme flood disturbance on macrophyte assemblages in a Neotropical floodplain. Aquat Sci 71(4):389–398
Padial AA, Ceschin F, Declerck SAJ, De Meester L, Bonecker CC, Lansac-Tôha FA, Rodrigues L, Rodrigues LC, Train S, Velho LFM, Bini LM (2014) Dispersal ability determines the role of environmental, spatial and temporal drivers of metacommunity structure. PLoS One 9(10):e111227
Papes M, Vander Zanden J (2010) Wisconsin lake historical limnological parameters 1925–2009. Long Term Ecological Research Network. doi:10.6073/pasta/66320ff8063706f6b3ee83a0ef3ef439. Accessed 15 Feb 2015
Peres-Neto PR, Legendre P, Dray S, Borcard D (2006) Variation partitioning of species data matrices: estimation and comparison of fractions. Ecology 87:2614–2625
Pip E (1989) Water temperature and freshwater macrophyte distribution. Aquat Bot 34:367–373
R Core Team (RCT) (2015) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. https://www.R-project.org/. Accessed Dec 2015
Riis T (2008) Dispersal and colonisation of plants in lowland streams: success rates and bottlenecks. Hydrobiologia 596:341–351
Riis T, Hawes I (2003) Effect of wave exposure on vegetation abundance, richness, and depth distribution of shallow plants in a New Zealand lake. Freshw Biol 48:75–87
Saarneel JM, Soons MB, Geurts JJM, Beltman B, Verhoeven JTA (2011) Multiple effects of land-use changes impede the colonization of open water in fen ponds. J Veg Sci 22:551–563
Santamaría L (2002) Why are most aquatic plants widely distributed? Dispersal, clonal growth, and small-scale heterogeneity in a stressful environment. Acta Oecol 23(3):137–154
Schmidt MH, Lefebvre G, Poulin B, Tscharntke T (2005) Reed cutting affects arthropod communities, potentially reducing food for passerines. Biol Conserv 121:157–166
Toivonen H, Huttunen P (1995) Aquatic macrophytes and ecological gradients in 57 small lakes in southern Finland. Aquat Bot 51:197–221
Tolonen KT, Hamalainen H, Holopainen IJ, Karjalainen J (2001) Influences of habitat type and environmental variables on littoral macroinvertebrate communities in a large lake system. Arch Hydrobiol 152(1):39–67
Valley RD, Heiskary S (2012) Short-term declines in curly-leaf pondweed in Minnesota: potential influences of snowfall. Lake Reserv Manag 28(4):338–345
Van Geest GJ, Wolters H, Roozen FCJM, Coops H, Roijackers RMM, Buijse AD, Scheffer M (2005) Water-level fluctuations affect macrophyte richness in floodplain lakes. Hydrobiologia 539:239–248
Vestergaard O, Sand-Jensen K (2000) Aquatic macrophyte richness in Danish lakes in relation to alkalinity, transparency, and lake area. Can J Fish Aquat Sci 57:2022–2031
Acknowledgments
We thank two anonymous reviewers for comments that have improved the manuscript. We thank Jani Heino for his development ideas in the early stage of the work. We also acknowledge the extensive efforts of area managers and field crews from the Minnesota and Wisconsin Departments of Natural Resources for obtaining the data used in our analyses. Work performed for this manuscript by MW Beck is not affiliated with the US Environmental Protection Agency.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Beck, M.W., Alahuhta, J. Ecological determinants of Potamogeton taxa in glacial lakes: assemblage composition, species richness, and species-level approach. Aquat Sci 79, 427–441 (2017). https://doi.org/10.1007/s00027-016-0508-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00027-016-0508-x