
Abstract
The annual world production of amino acids is currently estimated at more than seven million tons and is expected to reach ten million tons by 2022. This giant market has been underpinned largely by amino acid fermentation technologies in which Corynebacterium glutamicum has played a leading role. Various genetic engineering tools and global analysis techniques for this bacterium have been developed and successfully applied with a great impact on the amino acid industry. In particular, systems biology for this bacterium is almost fully capable of predicting targets to be engineered and metabolic states that will yield maximum production, thus allowing “systems metabolic engineering ” and development of industrially competitive production strains. Additionally, whole genomes of classically derived industrial producers have been analyzed by “reverse engineering” to identify important genetic traits, enabling the establishment of new industrial processes and the creation of genetically defined producers from scratch. This “genome breeding ” strategy was first developed using C. glutamicum as a model and currently yields producers that are more efficient than classical ones. These advances in strain development technology have almost achieved the optimization of entire cellular systems as cell factories for amino acid production, as demonstrated by their ability to produce glutamate and lysine at concentrations now exceeding 150 g/L with estimated production yields towards sugar at almost 70%. This chapter describes advances in the production of amino acids by C. glutamicum and presents the latest details of the technology and strategies used for molecular strain improvement.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
- Amino acid industry
- Global market for amino acids
- Glutamate
- Lysine
- Genome breeding
- Systems metabolic engineering
- Biosensor-driven single cell screening
1 Introduction
Amino acids have a wide variety of characteristics in terms of nutritional value, taste, medicinal action, and chemical properties, and thus have many potential uses as food additives, feed supplements, pharmaceuticals, cosmetics, polymer materials, and agricultural chemicals. As each new use is developed, demand for that type of amino acid grows rapidly and is followed by the development of mass production technology for that amino acid. The annual world production of amino acids has increased year by year (Fig. 1), from 0.7 million tons in 1985 to 1.7 million tons in 1996, 3.7 million tons in 2006, and 7.0 million tons in 2016, and is expected to reach ten million tons by 2022, growing at a CAGR (Compound Annual Growth Rate) of 5.6% from 2015 to 2022 (Ikeda 2003; Hermann 2003; Ajinomoto 2007, 2016; Sanchez et al. 2018). According to a recent market research report (Research and Markets 2018) and other relevant publications (Ajinomoto 2016; Sanchez et al. 2018), the global market for amino acids is estimated to be approximately US$13 billion in 2016 and is growing at an annual rate of 7%. Figure 2 shows the estimated global markets for amino acids of different applications in 2016. The feed amino acids, namely, lysine, methionine, threonine, and tryptophan, have the largest share of the market, generating US$7.0 billion. The second largest share (US$5.3 billion) belongs to food additives, which are comprised mainly of the flavor-enhancer monosodium glutamate and the amino acids aspartate and phenylalanine, both used as ingredients in the peptide sweetener aspartyl phenylalanyl methyl ester (Aspartame).

Changes in world annual production quantities of amino acids. The numbers in the squares indicate the estimated amounts of amino acids produced (1000 metric tons)

Estimated 2016 global markets for amino acids segmented by applications
Most l-amino acids are manufactured through microbial processes, mainly through fermentation. Corynebacterium glutamicum, which plays a principal role in amino acid fermentation, is therefore highly important, as demonstrated by the increasing number of relevant research papers (Fig. 3). Figure 3 also shows the main topics in amino acid fermentation and strain development technology during the decades since such research began. Amino acid fermentation was developed primarily in Japan and has extended across East Asia and into Europe, North America, and South America. Today amino acid fermentation is a global industry. It should be noted that China has achieved a prominent presence in this field, as reflected by the drastic increase in the number of research papers relevant to amino acid production (Ma et al. 2017; Zhang et al. 2017b; Cheng et al. 2018).

History of amino acid fermentation and strain development technology in Corynebacterium glutamicum, together with the annual number of research papers relevant to this microbe
In general, commercially potent producers have been developed by the stepwise accumulation of beneficial genetic and phenotypic characteristics in one background through classical mutagenesis and/or recombinant DNA technology. Such improvements involve strains capable not only of producing amino acids at higher yields but also of producing lower quantities of by-products, as the removal of by-products dominates the costs of downstream processing (Ikeda 2003; Marienhagen and Eggeling 2008; Feng et al. 2018). The current production yields towards sugar (w/w %) can be estimated as follows: lysine hydrochloride, 60–70; glutamate, 60–70; arginine, 40–50; isoleucine, 20–30; valine, 35–40; leucine, 25–30; tryptophan, 20–30; and phenylalanine, 30–35.
Since the year 2000, genomic and other “omics” data have accumulated for C. glutamicum, profoundly affecting strain development methods and providing a global understanding of the physiology, regulatory networks, and unknown functions of this microbe as well as the mechanisms underlying hyperproduction (Wittmann and Heinzle 2002; Ikeda and Nakagawa 2003; Kalinowski et al. 2003; Strelkov et al. 2004; Yukawa et al. 2007; Ikeda 2017; Yokota and Ikeda 2017; Becker et al. 2018). As a result, the targets of metabolic engineering have expanded beyond the core biosynthetic pathways leading to amino acids of interest into entire cellular systems including cofactor-regeneration systems, uptake and export systems, energy metabolism, global regulation, and stress responses. Such global and systematic metabolic engineering has repeatedly led to successful yield improvements for amino acid production by C. glutamicum (Eggeling and Bott 2005; Wendisch 2007; Burkovski 2008; Mitsuhashi 2014; Yokota and Ikeda 2017; Becker et al. 2018). In addition, the product spectrum of C. glutamicum has also been expanded, and metabolic engineering has been applied to the production of amino acids that formerly could not be produced effectively from glucose, such as serine, methionine, and cysteine. The present chapter describes the technologies and strategies that have been used in strain development in recent years, then reports the latest findings on rational metabolic engineering of C. glutamicum to develop efficient amino acid producers. Representative work using other bacteria, such as Escherichia coli and Pantoea ananatis , is also included for reference.
2 Recent Technologies for Strain Development
The focus in strain development technologies after the year 2000 has been directed to the development of new methodologies and tools employing genomic information, multi-omics data, bioinformatics, systems and synthetic biology, high-throughput single cell screening, and so on. These efforts have led to the development of several powerful new approaches, such as “systems metabolic engineering ”, “genome breeding ”, and “biosensor-driven single cell screening”, which has rejuvenated strain development for amino acid production. Such new approaches for strain development in C. glutamicum are highlighted here.
2.1 From Genome to Producers
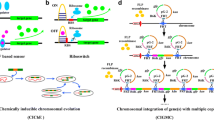
Advances in microbial genomics have dramatically transformed our approaches to strain development. Their largest benefit to this field is obviously the availability of high-throughput DNA sequencing, which has made it feasible to decode the genomes of classical industrial producers and thereby to identify important genetic traits that distinguish them from their wild-type ancestors. As a result, the conventional style of selecting improved strains by their phenotypes, formerly the standard practice in the industry, is rapidly being replaced by a new method called “genome breeding ” (Fig. 4), where desirable genotypes are systematically assembled in a wild-type genome (Ohnishi et al. 2002; Ikeda et al. 2006; Lee et al. 2012; Kim et al. 2013; Wu et al. 2015; Ma et al. 2018). The strains reconstructed from scratch through genome breeding can be more robust, give higher fermentation yields in less time, and resist stressful conditions better than classical industrial producers (Ohnishi et al. 2003). Meanwhile, microbial genomics allows in silico reconstruction of the whole metabolic map of a relevant microorganism. In fact, the genome information that is now available for a diverse variety of microorganisms has already revealed the numerous metabolic pathways that sustain their lives. This is now opening the way for the creation of a new methodology centered around redesigning a particular metabolic pathway in a desired microorganism on the model of one of the other metabolic pathways for which information is now publicly available. One example of this practice is the recent re-engineering of ‘Glutamic acid bacteria’ on the model of the unique redox metabolism seen in ‘Tooth decay bacteria’ (Takeno et al. 2010, 2016). Here, the two approaches, genome breeding and metabolic redesign , are described to show how we can take advantage of genome information to enable more efficient amino acid production.

The “genome breeding ” methodology for the creation of defined mutants that carry only beneficial mutations. This methodology starts with decoding the genomes of classical industrial producers to identify the important genetic traits that distinguish them from their wild-type ancestors (the Reverse engineering step) and progresses to systematically assembling the beneficial genetic properties in a single wild-type background (the Reengineering step)
2.1.1 Genome Breeding
Because classical strain breeding is based on random mutation and selection, we cannot eliminate the possibility that this method will introduce detrimental or unnecessary mutations into a genome. Genome breeding methodology, however, can overcome this limitation. In this program, biotechnologically useful mutations identified through the genome analysis of classical mutants are systematically introduced into the wild-type genome in a pinpointed manner (Ikeda et al. 2005), thus allowing the creation of a defined mutant that carries only useful mutations (Figs. 4 and 5). As an example, one industrial lysine producer that had undergone years of mutagenesis and screening was found to have more than 1000 mutations accumulated in its genome (Ikeda 2017). Among these, only six mutations were identified as positive mutations for lysine production ; two (hom59 and lysC311) that are located in the terminal pathway to lysine (Ohnishi et al. 2002), three (pyc458, gnd361, and mqo224) that are involved in central metabolism (Ohnishi et al. 2002, 2005; Mitsuhashi et al. 2006), and one (leuC456) that causes global induction of the amino acid-biosynthetic genes and thereby further increases production (Hayashi et al. 2006a). The assembly of these six useful mutations in a robust wild-type strain of C. glutamicum (Fig. 5a) was shown to substantially improve producer performance, resulting in a final titer of 100 g/L after 30 h of 5-L jar fermentor cultivation at a suboptimal temperature of 40 °C (Ohnishi et al. 2003; Ikeda et al. 2006).

Reengineering of a lysine producer (a) and an arginine and citrulline producer (b). In the case of the new lysine producer, six beneficial mutations identified from the genome of a classical lysine producer were assembled in a single wild-type background (Ikeda et al. 2006), while the new arginine and citrulline producer was created by assembly of three positive mutations derived from three different lines of classical producers (Ikeda et al. 2009)
The usefulness of the genome breeding approach has been also demonstrated in the production of arginine and citrulline (Ikeda et al. 2009, 2010a). In this case, the assembly of three positive mutations (argB26, ΔargR, and argB31) derived from three different lines of classical producers in a single wild-type background (Fig. 5b) has led to the new strain RBid, characterized by dramatically increased productivity of arginine and citrulline compared with the best classical producer, A-27, even at the suboptimal temperature of 38 °C (Fig. 6). In this approach, not only identification of beneficial mutations but also screening for the specific host that will give the best performance is an important consideration because the wild-type background can have a significant impact on the ultimate outcome (Ohnishi and Ikeda 2006; Ikeda et al. 2009). The host strain into which the three mutations were incorporated was ATCC 13032, which was identified through screening from among several C. glutamicum wild-type strains as the strain with the highest potential for industrial arginine/citrulline production at elevated temperatures (Ikeda et al. 2009). If another wild-type strain had been used as the host, the result would have been unsatisfactory.

Fermentation kinetics of the newly developed strain RBid at 38 °C in 5-L jar fermentor cultivation. For comparison, the profiles of the best classical producer A-27 when cultured under its optimal 30 °C conditions are shown as controls. Open circles arginine and citrulline production by strain RBid, closed circles growth of strain RBid, open squares arginine and citrulline production by strain A-27, closed squares growth of strain A-27
2.1.2 Metabolic Redesign
Progress in genomics has made it possible to construct an organism’s entire metabolic map in silico. Diverse metabolic pathways of approximately 3500 bacterial species have already been constructed and are available at the Kyoto Encyclopedia of Genes and Genomes (http://www.kegg.jp/kegg-bin/get_htext?htext=br08601_map00010.keg&hier=5) (Kanehisa et al. 2014). Most well-known bacteria possess the complete pentose phosphate pathway which enables them to generate NADPH as a reducing power. In some unique microorganisms, however, the database has revealed a defective pentose phosphate pathway. One such microorganism is Streptococcus mutans, a tooth decay bacterium reportedly harboring an unusual glycolytic pathway that can generate NADPH at the step of glyceraldehyde 3-phosphate dehydrogenase (Fig. 7). Based on these findings, an attempt has been made to recreate the S. mutans-type NADPH-generating glycolytic pathway in C. glutamicum (Takeno et al. 2010, 2016). In this study, endogenous NAD-dependent glyceraldehyde 3-phosphate dehydrogenase (GapA) of C. glutamicum was replaced with nonphosphorylating NADP-dependent glyceraldehyde 3-phosphate dehydrogenase (GapN) of S. mutans (Fig. 8). Unfortunately, the resulting strain (RE2) exhibited severely retarded growth, probably because the engineering attempt had favored the generation of reducing power while theoretically restricting ATP generation. The strategy for solving this problem was to use GapA together with GapN in the early growth phase where more ATP is required for growth, and thereafter to shift the combination-type glycolytic pathway to one that depends only on GapN for production in the subsequent growth phase (Fig. 8). To achieve this, the gene for GapA was expressed under the myo-inositol-inducible promoter of iolT1 encoding a myo-inositol transporter. In strain RE2Aiol which was thus engineered, a well-balanced use of GapA and GapN has led to both improved growth and high-level lysine production . Moreover, it has been demonstrated that blockade of the oxidative pentose phosphate pathway through a defect in glucose 6-phosphate dehydrogenase did not significantly affect lysine production in the engineered strain (Fig. 8), while a drastic decrease in lysine production was observed for the reference strain (Takeno et al. 2016). This study was the first to demonstrate efficient lysine production independent of the oxidative pentose phosphate pathway.

Glycolysis and the pentose phosphate pathway in C. glutamicum ATCC 13032 and S. mutans UA159. The metabolic pathways are extracted from the KEGG pathway database (Kanehisa et al. 2014). Each enzyme is presented as an EC number. GapA NAD-dependent glyceraldehyde 3-phosphate dehydrogenase from C. glutamicum, GapC NAD-dependent glyceraldehyde 3-phosphate dehydrogenase from S. mutans, GapN nonphosphorylating NADP-dependent glyceraldehyde 3-phosphate dehydrogenase

Engineering of C. glutamicum with the S. mutans-type NADPH-generating glycolytic pathway for lysine production . The control strain C. glutamicum ΔgapB has an NADH-generating glycolytic pathway (left). Replacement of endogenous GapA with S. mutans GapN created an NADPH-generating glycolytic pathway, leading to increased lysine production but retarded fermentation (middle). Further engineering to supplementarily express GapA only in the early growth phase resulted in efficient lysine production independent of the oxidative pentose phosphate pathway (right)
2.2 Systems Metabolic Engineering
The cumulative body of knowledge on cellular metabolism and physiological properties of amino acid-producing microorganisms was combined with “omics” technologies and computational methods, including metabolic flux profiling and in silico modeling, to facilitate metabolic engineering in a systematic and global manner (Dai and Nielsen 2015; Hirasawa and Shimizu 2016; Ma et al. 2017; Lee and Wendisch 2017; Zhang et al. 2017b; Becker et al. 2018). This “systems metabolic engineering ” approach is particularly useful for predicting a combination of genetic modifications that would lead to the theoretically best flux scenario for amino acid production. Several applications of this have recently led to the successful creation of efficient amino acid producers in both C. glutamicum and E. coli.
Becker et al. (2011) used flux model analysis with a genome-scale metabolic model to predict the target steps to be modified for optimum lysine production by C. glutamicum. Ultimately, this analysis identified twelve stages of modifications to a wild-type genome resulting in the lysine hyper-producer LYH-12 (Fig. 9), which can achieve a final titer of 120 g/L with a conversion yield of 55% on glucose after 30 h of 5-L jar fermentor cultivation at 30 °C. Among the twelve modifications were six (introduction of the lysC311 and hom59 mutations, duplication of the ddh and lysA genes, and overexpression of the lysC and dapB genes under a strong promoter) that cause increased flux through the lysine biosynthetic pathway, three (introduction of the pyc458 mutation, overexpression of the pyc gene under a strong promoter, and deletion of the pck gene) that cause increased flux towards oxaloacetate through anaplerotic carboxylation, two (overexpression of the fbp gene and the zwf-opcA-tkt-tal operon under strong promoters) that cause increased flux through the pentose phosphate pathway for NADPH supply, and one (replacement of the start codon ATG by the rare GTG in the icd gene) that causes reduced flux through the TCA cycle and thereby increases the availability of oxaloacetate.

Schematic diagram of genetic modifications to a wild-type genome leading to the lysine hyper-producer C. glutamicum LYS-12
Lee et al. (2007) and Park et al. (2007a) reported the strategies for systems metabolic engineering of E. coli for the production of threonine and valine, respectively. For threonine production, the target genes to be engineered were identified through transcriptome profiling and in silico flux response analysis, ultimately resulting in construction of a defined threonine hyper-producer that can achieve a final titer of 82.4 g/L with a conversion yield of 39.3% on glucose after 50 h of fed-batch culture (Lee et al. 2007). For valine production, likewise, in silico gene knockout simulation identified three target genes to be disrupted , leading to the design of an efficient valine producer (Park et al. 2007a).
2.3 Biosensor-Driven Single Cell Screening
Since rational strain improvement generally depends on known genetic information, relying on this approach alone can cause researchers to miss unknowns, which are often difficult to predict. For this reason, the classical approach consisting of multiple rounds of random mutation and screening is still significant for strain improvement. The classical approach also offers opportunities to find novel information applicable to the rational approach. Yet screening almost always requires the cultivation and subsequent productivity analysis of individual mutants, which requires considerable time, labor, and cost. Biosensor-driven single cell screening is a technique that allows researchers to overcome such disadvantages and thereby accelerate strain improvement. The major technique that has been applied to C. glutamicum is based on the principle that transcriptional regulators (TRs) activate expression of their target gene fused to a reporter gene eyfp (encoding enhanced yellow fluorescent protein, eYFP) in response to intracellular concentration of a specific metabolite. This setup enables translation of the intracellular metabolite concentration into a fluorescent output and the high-throughput screening of single cells via fluorescence-activated cell sorting (FACS). The representative achievements have been performed using the homologous TRs: LysG and Lrp (Fig. 10).

Biosensors using transcriptional regulators LysG (a) and Lrp (b). The biosensors are based on the principle that transcriptional regulators activate the target gene fused to a reporter gene eyfp in response to intracellular concentration of each effector metabolite. Eventually, increased intracellular concentration of the metabolite is reflected as increased fluorescence, which enables screening of single cells by fluorescence-activated cell sorting (FACS). (a) LysG activates expression of the lysE gene in the presence of increased levels of the effectors lysine, arginine, or histidine. (b) Lrp activates expression of the brnFE operon in the presence of increased levels of valine, leucine, isoleucine, or methionine
In response to cytosolic concentrations of basic amino acids such as lysine, arginine, or histidine, LysG activates expression of the lysE gene encoding an exporter for lysine and arginine (Bellmann et al. 2001). A plasmid containing the eyfp gene under the control of the promoter of the lysE gene enables the translation of the intracellular lysine concentration into a fluorescent signal (Binder et al. 2012). A mutant library consisting of 7 × 106 cells obtained from wild-type C. glutamicum ATCC 13032 carrying this plasmid was subjected to FACS analysis, which resulted in the isolation of 185 lysine-producing mutants. It should be noted that increased intracellular concentration of lysine correlates with its increased concentration in the culture supernatant. Whole-genome sequencing of the mutants revealed novel mutations in the murE gene encoding UDP-N-acetylmuramoyl-l-alanyl-d-glutamate-2,6-diaminopimelate ligase that utilizes d, l-diaminopimelate as a substrate in the lysine synthetic pathway. One of these mutations, mutation murE G81E, was introduced into the genome of the defined lysine producer C. glutamicum DM1933 (Blombach et al. 2009b) where it increased lysine production by approximately 1.3-fold on glucose (Binder et al. 2012). The LysG-based biosensor has also been utilized to isolate less-feedback variants of key enzymes in lysine, arginine, and histidine biosynthesis (Schendzielorz et al. 2014). This technique was performed on a mutant library consisting of approximately 2.2 × 106 cells of the C. glutamicum ΔargR strain carrying a plasmid containing a randomly mutated argB gene encoding N-acetyl-l-glutamate kinase. Analysis of 96 selected positive cells revealed the presence of ArgB variants. The argB gene, which was modified based on information on these variants, encoded a feedback-resistant enzyme to inhibition by arginine and allowed the ΔargR strain to produce 35 mM arginine on glucose (Schendzielorz et al. 2014). A similar approach has identified feedback-resistant variants of aspartate kinase (LysC) and ATP phosphoribosyl transferase (HisG), which are key enzymes in lysine and histidine biosynthesis, respectively, and revealed that these mutations lead to the production of the respective amino acids by the wild-type strain (Schendzielorz et al. 2014).
Another representative example of biosensor-driven single cell screening is based on the transcriptional regulator Lrp. Lrp activates expression of the brnFE operon encoding the export system for valine, leucine, isoleucine, and methionine in response to intracellular concentration of these amino acids (Lange et al. 2012). By a principle similar to that of the LysG-based biosensor, the Lrp-based biosensor led to the isolation of five mutants that produced different amounts of valine, leucine, and isoleucine from a random mutant library of wild-type ATCC 13032 (Mustafi et al. 2012). The Lrp-based approach has also been utilized for adaptive laboratory evolution for valine production . An evolved consortium originating from the C. glutamicum ΔaceE strain and having gone through five iterative evolution steps exhibited a higher growth rate, up to 25% increased valine production, and three- to four-fold decreased production of the by-product alanine. Genome sequencing of a single isolate revealed a loss-of-function mutation in the ureD gene encoding the urease accessory protein, and introduction of this mutation into the non-evolved ΔaceE strain resulted in an increase in valine production up to two-fold (Mahr et al. 2015).
Very recently, TR-based biosensors coupled with FACS for C. glutamicum have been expanded to detection of shikimic acid, serine, and cAMP. The shikimic acid biosensor is based on the transcriptional regulator ShiR (Kubota et al. 2015). This was utilized in the screening of a ribosome binding site (RBS) library for the tktA gene encoding transketolase, an enzyme that catalyzes the formation of erythrose-4-phosphate, a precursor of shikimic acid, and in the identification of a useful RBS sequence for improved shikimic acid production (Liu et al. 2018a). In contrast, the serine biosensor employs the transcriptional regulator NCgl0581 (Binder et al. 2012). This system was used to screen the random mutant library of a serine-producing C. glutamicum strain that was obtained through a combination of random mutagenesis and rational engineering, which resulted in the isolation of a mutant with 1.3-fold higher serine production than its parental strain (Zhang et al. 2018e). The mutant produced 35 g/L of serine with a conversion yield of 35% on sucrose after 120 h of batch culture (Zhang et al. 2018e). Although all of the TRs described above bind with the corresponding effectors and thereafter can activate expression of the reporter gene, the cAMP sensor is composed of the transcriptional regulator GlxR and a promoter repressed by cAMP-bound GlxR. This biosensor has been used successfully to separate cells with different cAMP levels (Schulte et al. 2017).
3 Current Status of Amino Acid Production
Recently, various genetic engineering tools and global analysis techniques for C. glutamicum as well as high-throughput genomic analysis technologies have been successfully applied and have contributed both to the understanding of the molecular mechanisms underlying high-level production and to the development of more advanced production strains of this microbe. At the same time, the new approaches to strain development such as genome breeding and systems metabolic engineering have allowed the creation of nearly optimal genetically defined and industrially competitive producers from scratch. Here, the current status of production of various amino acids by C. glutamicum is highlighted, with a special focus on the amino acids whose production methods have been significantly advanced in the 2000s. These include glutamate, lysine, arginine, citrulline, ornithine, the branched-chain amino acids valine, leucine, and isoleucine, and the sulfur-containing amino acids methionine, S-adenosyl-methionine, and cysteine. Tryptophan is also included because the scale of its market has been growing rapidly. Production technology aimed at other industrially important amino acids such as serine, alanine, and threonine has been omitted because it was discussed in the first edition of this book and/or other publications (Ikeda 2003, Willis et al. 2005; Sprenger 2007; Rieping and Hermann 2007; Dong et al. 2011; Ikeda and Takeno 2013; Yokota and Ikeda 2017).
3.1 Glutamate
Since the discovery of C. glutamicum as a producer of the food flavoring monosodium glutamate, commercial production of glutamate has been conducted using this microbe exclusively. The industrial glutamate titer is assumed to exceed 150 g/L (Sanchez et al. 2018) with an estimated production yield towards sugar of almost 70%. The global demand for monosodium glutamate amounted to over three million tons in 2016 (Ajinomoto 2016; Sanchez et al. 2018) and is expected to surpass four million tons by 2023 (Global market insights 2016). Glutamate production by C. glutamicum is induced by biotin limitation or by treatment with certain fatty acid ester surfactants or with β-lactam antibiotics such as penicillin. Although induction treatment is the core technology involved in industrial glutamate production processes, the molecular basis of the induction of glutamate secretion was long unknown. In recent years, however, a valuable insight into the secretion mechanism has been gained with the identification of the NCgl1221 gene product as a glutamate exporter (Nakamura et al. 2007). An intriguing finding is that only a specific point mutation in the NCgl1221gene resulted in glutamate secretion without any induction treatments. It has also been shown that amplification of the wild-type NCgl1221gene increases glutamate secretion while its disruption substantially abolishes secretion accompanied by an increase in the intracellular glutamate pool under the induction conditions mentioned above. The gene in question encodes the YggB protein, which was originally described as a putative mechanosensitive channel (Nottebrock et al. 2003). Later electrophysiological studies using an E. coli or Bacillus subtilis strain devoid of mechanosensitive channels indicated that the NCgl1221 gene product actually possesses the activity of a mechanosensitive channel (Börngen et al. 2010; Hashimoto et al. 2010). It has also been shown that glutamate excretion through the channel was mediated by passive diffusion (Hashimoto et al. 2012), while carrier-mediated glutamate secretion by C. glutamicum was shown to be energy-dependent (Gutmann et al. 1992). Based on this possible function as a mechanosensitive channel, the following mechanism has been proposed: the induction conditions, such as biotin limitation and penicillin treatment, alter membrane tension by inhibiting lipid or peptidoglycan synthesis. This triggers conformational changes in the NCgl1221 gene product, which in turn enables the protein to export glutamate (Fig. 11).

Possible mechanisms triggering glutamate overproduction under the induction conditions in C. glutamicum. In this model, proteins AccBC and DtsR form the biotin-dependent acetyl-CoA carboxylase complex required for fatty acid biosynthesis; this biotin-enzyme complex is thought to be the primary target of biotin limitation and surfactant addition
Very recently, a different type of mechanosensitive channel was identified as a second glutamate exporter in some limited strains of C. glutamicum (Wang et al. 2018b). Although the channel MscCG2 shares only 23% identity with the NCgl1221 gene product, both channels have an important common feature: they need to be activated for glutamate production by the alteration of membrane tension, which is triggered by biotin limitation or penicillin treatment. This observation raises the question of how the new model may be congruent with the accepted notion that a decrease in the activity of the 2-oxoglutarate dehydrogenase complex (ODHC) is crucial for glutamate production (Shingu and Terui 1971; Kawahara et al. 1997; Kimura 2003; Asakura et al. 2007; Kim et al. 2009a, b). Although the new model seems to explain the basics of the mechanism underlying the induction of glutamate secretion, it is probably not sufficient to explain the entire process of glutamate production by C. glutamicum. Recently, a possible connection at a molecular level has been uncovered between ODHC activity and glutamate production (Fig. 11). A novel 15 kDa protein known as OdhI was identified as a regulator of ODHC (Niebisch et al. 2006). The unphosphorylated form of OdhI binds to the OdhA protein, one of the subunits of ODHC, and inhibits ODHC activity. This inhibition can be prevented by the PknG-catalyzed phosphorylation of OdhI. A phospho-serine/threonine protein phosphatase responsible for dephosphorylation of OdhI has also been identified (Schultz et al. 2007). Interestingly, disruption of the odhI gene was shown to abolish glutamate production even under the induction conditions (Schultz et al. 2007), suggesting a close relationship between the regulator protein and the reduction of ODHC activity that occurs during glutamate production. It is also worth noting that proteome analyses have revealed a significant increase in the OdhI protein upon penicillin treatment, which has become a conventional industrial method of triggering glutamate production (Kim et al. 2009a, b). These findings have confirmed the existence of a connection between ODHC activity and glutamate production, but it should be noted that an ODHC-activity-reducing metabolic change alone is not sufficient to induce glutamate production (Kim et al. 2009a, b).
Taken together, the evidence to date suggests a link between the induction treatments, such as biotin limitation and penicillin treatment, and glutamate production . In our proposed mechanism, the induction treatments enhance the synthesis of the regulator protein OdhI in its unphosphorylated form and thereby inhibit ODHC activity. This causes a metabolic shift at the branch point of 2-oxoglutarate, which channels carbon toward glutamate. Intracellularly-accumulated glutamate is then secreted into the medium via the NCgl1221 gene product YggB which has been activated in response to altered membrane tension (Fig. 11). Questions for the future include why and how the OdhI protein is overexpressed in response to the induction treatment and what conditions are required for the phosphorylation and dephosphorylation of OdhI.
Acetylome and succinylome analyses of glutamate-producing C. glutamicum have suggested that protein acetylation and succinylation are involved in glutamate production through the post-translational control of key enzymes such as phosphoenolpyruvate carboxylase (PPC) and ODHC (Mizuno et al. 2016). It has been shown that acetylation of PPC at lysine 653 caused decreased enzymatic activity, resulting in reduced glutamate production (Nagano-Shoji et al. 2017). On the contrary, deacetylation of the lysine residue has been suggested to improve glutamate production through activation of PPC because the increase in PPC activity during glutamate production was canceled by the defect of the deacetylases (Nagano-Shoji et al. 2017).
Continuous efforts have been made not only to understand glutamate production but also to improve the process. In addition to the general approaches, in which metabolic fluxes are directed into glutamate (Kimura 2003; Sato et al. 2008; Sawada et al. 2010), an innovative metabolic design allowing an increased maximum theoretical yield has recently been reported (Chinen et al. 2007). Glutamate biosynthesis from glucose in C. glutamicum is inevitably associated with the release of CO2 in the pyruvate dehydrogenase reaction, but the creation of a novel metabolic route by installing the phosphoketolase pathway of Bifidobacterium animalis allowed the CO2-releasing pyruvate dehydrogenase reaction to be bypassed via acetyl phosphate, and thereby led to increased glutamate production coupled with the suppression of CO2 emission. On the other hand, expression of the Vitreoscilla hemoglobin gene vgb under a tac promoter in a wild-type C. glutamicum strain has been shown to increase glutamate production in both shake-flask and fermentor cultivations (Liu et al. 2008), probably due to the enhancement of respiration by the hemoglobin (Webster 1987; Kallio et al. 1994; Zhang et al. 2007). Very recently, a Chinese group has demonstrated efficient glutamate production in the biotin-excessive corn stover hydrolysate (lignocellulose biomass). To achieve this, an industrial strain of C. glutamicum was improved by two genetic modifications, that is, (1) truncating C-terminal amino acid residue of the mechanosensitive channel NCgl1221, leading to activation of glutamate secretion without any induction treatments, and (2) engineering odhA ribosome-binding site, leading to decreased ODHC activity. The engineered strain XW6 achieved a final titer of 65.2 g/L with a yield of 63% after 48 h of 3-L jar fermentor cultivation using corn stover hydrolysate as the feedstock (Wen and Bao 2019).
Aside from C. glutamicum, Ajinomoto isolated the gram-negative acid-tolerant bacterium Pantoea ananatis AJ13355 to generate a glutamate producer (Hara et al. 2012). This producer was shown to allow glutamate fermentation to be conducted under acidic conditions (pH 3–5) where the solubility of glutamate is low, leading to a new process called “glutamate crystallization fermentation ” (Izui et al. 2006, Usuda et al. 2017). This new type of fermentation is considered to reduce the amounts of alkali (e.g., ammonia) and acid (e.g., sulfuric acid or hydrochloric acid) during the fermentation and subsequent purification steps, respectively, and also the amounts of by-product salts such as ammonium sulfate. This means that the new process is expected to decrease not only production costs but also the burden on the environment, thus indicating its potential as a sustainable production process.
3.2 Lysine
Lysine, an essential amino acid for animals, has significant commercial value as a feed additive to promote the growth of animals including swine and poultry, and thus is the second-ranking amino acid after glutamate in terms of worldwide annual production. Lysine is also used as a fish feed additive because it is generally the first limiting essential amino acid in many protein sources used in fish feeds (Hua 2013). The scale of the lysine market has been estimated at approximately 2.3 million tons in 2016 (Ajinomoto 2016) and is still growing at annual rates of around 10% (Ikeda 2017). As the scale of production has increased, lysine prices per kilogram have dropped to around US$1.4 (Ajinomoto 2016), fluctuating between US$1.2 and 2.5 over the past decade (Ajinomoto 2016; Eggeling and Bott 2015; Ikeda 2017), depending largely on competition from natural lysine sources such as soybean meal and sardine. The main suppliers are CJ CheilJedang (South Korea), Global Bio-Chem Technology Group (China), Ajinomoto (Japan), Archer Daniels Midland (USA), and Evonik Industries (Germany), among others (Eggeling and Bott 2015). Major commercial plants are located in the respective corn belts in China, North America, Brazil, Indonesia, and Russia. Because of the growing market for lysine, exhaustive studies have been undertaken in an attempt to engineer the metabolism of C. glutamicum for lysine production (Ikeda 2017). These studies have resulted in several effective strategies for rational strain improvement, including engineering of terminal pathways (Shiio and Miyajima 1969; Sano and Shiio 1971; Kase and Nakayama 1974), central metabolism (Petersen et al. 2001; Peters-Wendisch et al. 2001; Riedel et al. 2001; Shiio et al. 1984; Becker et al. 2009; Chen et al. 2014; Blombach et al. 2007b; Radmacher and Eggeling 2007; van Ooyen et al. 2012; Mitsuhashi et al. 2006), NADPH-regeneration systems (Marx et al. 2003; Becker et al. 2005, 2007; Ohnishi et al. 2005; Kiefer et al. 2004; Moon et al. 2005; Takeno et al. 2010, 2016; Komati Reddy et al. 2015; Bommareddy et al. 2014; Kabus et al. 2007a; Xu et al. 2014c, 2018b, c), export systems (Burkovski and Krämer 2002; Vrljić et al. 1996), glucose uptake systems (Ikeda et al. 2010b, 2011, 2015; Lindner et al. 2011a, b; Ikeda 2012), energy metabolism (Bott and Niebisch 2003; Kabus et al. 2007b), and global regulation (Burkovski 2008; Brockmann-Gretza and Kalinowski 2006; Krömer et al. 2004, 2008; Hayashi et al. 2006a, b). In addition, recent genome-based and systems-level approaches such as genome breeding (Ohnishi et al. 2002; Ikeda et al. 2006) and systems metabolic engineering (Becker et al. 2011) have led to lysine producers with superior production performance in terms of yield, titer, and productivity, as described earlier in this chapter.
A genome-scale model of the C. glutamicum metabolic network has been constructed based on the annotated genome, available literature, and various “omic” data (Kjeldsen and Nielsen 2009). The constructed metabolic model consists of 446 reactions and 411 metabolites; the predicted metabolic fluxes during lysine production and growth under various conditions are highly consistent with experimental values. The ability to predict the metabolic state associated with maximum production yield can be used to guide strain engineering. This strategy has been proven through the rational design of high lysine-producing strains of C. glutamicum (Krömer et al. 2004; Becker et al. 2005; Wendisch et al. 2006).
Recently, Chinese groups have dramatically improved lysine productive performance of C. glutamicum by rational metabolic engineering (Xu et al. 2013, 2014a, b, c, 2018a, b, c). Seventeen stages of modification to a wild-type genome, aimed at increasing the carbon flow into the lysine-biosynthetic pathway, have led to the lysine hyper-producer Lys5-8, which achieved a final lysine titer of 130.82 g/L (163.52 g/L as lysine hydrochloride) with a conversion yield of 47.06% on glucose after 48 h of 7-L jar fermentor cultivation (Xu et al. 2014b). Very recently, different lines of lysine hyper-producers have been rationally constructed from a classical lysine producer. These include strains JL-6 ΔdapB::Ec-dapB C115G, G116C (Xu et al. 2018b) and JL-69Ptac-Mgdh (Xu et al. 2018a). The typical strategy, used to create the former strain, was to lower dependency on NADPH during lysine biosynthesis. For this purpose, the NADPH-dependent DapB gene was replaced with an E. coli gene encoding a mutant type of NADH-dependent DapB to switch its nucleotide-cofactor specificities from NADPH to NADH, leading to an increase in final lysine titer from 82.6 to 117.3 g/L, an increase in conversion yield on sugar from 35 to 44%, and an increase in productivity from 2.07 to 2.93 g/L/h in 5-L jar fermentor cultivation. The latter strain, in contrast, was created by optimizing the carbon flux through the TCA cycle to balance cell growth and availability of oxaloacetate and glutamate for lysine biosynthesis from the viewpoint of maximizing precursor supply. Rational engineering of the phosphoenolpyruvate-pyruvate-oxaloacetate node and the TCA cycle, as well as the suitable feeding of biotin, has resulted in a final lysine titer of 181.5 g/L with a conversion yield of 64.6% on sugar after 48 h (productivity of 3.78 g/L/h) in 5-L jar fermentor cultivation. On the other hand, a classically derived lysine-producing mutant of C. glutamicum was improved by engineering glucose uptake systems. Adequate expression of the bypasses for glucose uptake (IolT1, IolT2, and PpgK), in addition to the native phosphoenolpyruvate-dependent sugar phosphotransferase system (PTSGlc), in the strain has resulted in the lysine hyper-producer ZL-92, which achieved a final lysine titer of 201.6 g/L with a conversion yield of 65% on glucose after 40 h (productivity of 5.04 g/L/h) in 5-L jar fermentor cultivation (Xu et al. 2019). An outline of the metabolic engineering strategies by which these lysine hyper-producers were generated is schematically shown in Fig. 12.

Outline of metabolic engineering disclosed by the Chinese groups for lysine production
The main feedstocks for lysine production by C. glutamicum are sugars from agricultural crops, such as cane molasses, beet molasses, and starch hydrolysates (glucose) from corn and cassava, but it is becoming necessary to engineer the use of alternative raw materials, in particular, materials that do not compete with human food or energy sources. To reduce the environmental impact of lysine production, C. glutamicum strains have been constructed that can utilize whey, which contains lactose and galactose (Barrett et al. 2004); glycerol, the main by-product of biodiesel production (Rittmann et al. 2008); lignocellulose, which contains the pentoses xylose and arabinose (Kawaguchi et al. 2006, 2008); and rice straw hydrolysate (Meiswinkel et al. 2013), though there are still technical challenges related to upstream raw material processing and carbon use efficiency. In addition to these attempts to employ nonedible second-generation renewables, there is an increasing interest in the development of a lysine production process using mannitol, a major constituent of marine microalgae (seaweed), as a third-generation renewable resource that might be more efficiently and sustainably supplied from ocean farms (Hoffmann et al. 2018). The sugar alcohol mannitol can be metabolized into lysine by an engineered C. glutamicum strain that expressed the NADP-dependent glyceraldehyde 3-phosphate dehydrogenase (GapN) of S. mutans so as to couple the glycolysis to NADPH formation, resulting in a lysine yield of 0.24 mol/mol and a productivity of 1.3 mmol/g/h.
In addition to strain engineering, continuous improvement of the process and the development of a comprehensive methodology for assessing the process (Anaya-Reza and Lopez-Arenas 2017) have resulted not only in fermentation processes with increased product yields and reduced loads on downstream processing, but also in the commercialization of various product forms for novel intended uses (Hirao et al. 1989; Ikeda 2003, 2017; Kelle et al. 2005; Kobayashi et al. 2011). Meanwhile, anaerobic production of lysine through a C. glutamicum process remains a great challenge, though several attempts, including the operation of a nitrate respiration system (Nishimura et al. 2007; Takeno et al. 2007) and the use of an anode such as ferricyanide as the extracellular electron carrier (Xafenias et al. 2017; Vassilev et al. 2018), have indicated that it may be possible to turn an aerobic production process into an anaerobic process.
3.3 Arginine, Citrulline, and Ornithine
Arginine, a semi-essential amino acid, has lately attracted considerable attention for being a precursor to nitric oxide (NO), a key component of endothelial-derived relaxing factor (Appleton 2002). Citrulline and ornithine, precursors of arginine biosynthesis as well as intermediates in the urea cycle, are also important for human health since they are sources of endogenous arginine in the body (Hayashi et al. 2005, 2006c; Curis et al. 2007; Ochiai et al. 2012; Mori et al. 2015). As the economic values of these amino acids have increased, considerable attention has been given to improving our understanding of their metabolism in microbes (Utagawa 2004; Glansdorff and Xu 2007; Lee et al. 2010; Petri et al. 2013; Huang et al. 2015, 2016; Lubitz et al. 2016). In parallel, increasing efforts have been directed to the development of more efficient production strains by using recent technologies for strain development. For example, the genome breeding approach has been successfully applied to develop an arginine and citrulline producer from a C. glutamicum wild-type strain, as described earlier in this chapter (Ikeda et al. 2009). The reengineered strain was constructed by assembling just three mutations (argB26, ΔargR, and argB31) derived from three different lines of classical producers, resulting in a final titer of over 80 g/L (as a sum of arginine and citrulline) in 30 h of 5-L jar fermentor cultivation at a suboptimal temperature of 38 °C (Fig. 5b and 6).
Recently, a classically derived arginine-producing mutant of C. glutamicum was improved through stepwise metabolic engineering, including deregulation of arginine biosynthesis, increased NADPH availability, disruption of the glutamate exporter, and adequate modifications of the terminal pathway (Fig. 13). This systematic approach has ultimately led to the arginine hyper-producer AR6, which achieved a final titer of 92.5 g/L with a conversion yield of 40% on glucose plus sucrose after 72 h of 5-L jar fermentor cultivation (Park et al. 2014). It is notable that citrulline by-production was not observed in the fermentation.

Different approaches of metabolic engineering applied to the production of arginine
Likewise, a Chinese group has employed a similar metabolic engineering strategy for the development of the C. glutamicum arginine hyper-producer Cc6 (Fig. 13), which achieved a final titer of 87.3 g/L with a yield of 43.1% on glucose after 72 h of 5-L jar fermentor cultivation (Man et al. 2016b). They have also shown that modifications leading to decreased H2O2 synthesis and increased NADH and ATP levels contribute to improved arginine production (Man et al. 2016a).
Meanwhile, a German group took a different approach to developing producers of arginine, citrulline, and ornithine (Jensen et al. 2015). They first constructed the C. glutamicum ornithine-producing strain ORN2 (ΔargF, ΔargR, ΔargG) as a platform strain for subsequent development. Additional modifications, including deregulation of ArgB, overexpression of gdh, attenuation of pgi, and duplication of argCJB, resulted in the ornithine hyper-producer ORN6 with a yield of 52% on glucose. This ornithine hyper-producer was then converted into strains capable of producing citrulline (a yield of 41%) and arginine (a yield of 30%) by plasmid-mediated overexpression of argFB and argGFB, respectively. It is worth noting that the two specific mutations (A49V, M54 V) used for deregulation of ArgB are the same as those originally identified from the genomes of classical arginine producers during the genome breeding process mentioned earlier (Ikeda et al. 2009, 2010a).
Metabolic engineering of C. glutamicum has also led to industrially potent ornithine producers. For example, a Korean group has generated an ornithine producer by removal of competing pathways (ΔargF, ΔproB), deregulation of ornithine biosynthesis (ΔargR), overexpression of the terminal pathway with the use of plasmid pSY223 carrying argCJBD, and increased NADPH availability (pgi GTG, zwf ATG, Ptkt::Psod). The resulting strain YW06 (pSY223) produced 51.5 g/L of ornithine with a yield of 24% on glucose after 40 h of 6.6-L jar fermentor cultivation (Kim et al. 2015).
Chinese groups have also developed C. glutamicum ornithine producers through systematic metabolic engineering (Jiang et al. 2013a, b; Zhang et al. 2017a, 2018a, b, c). One of their strategies was to convert an industrial glutamate producer into an ornithine producer by adequately attenuating argF, followed by combining the common strategies including deregulation of ornithine biosynthesis, increased NADPH availability, disruption of the glutamate exporter, removal of competing pathways, and adequate modifications of the terminal pathway. The resulting strain SO16 produced 32.3 g/L of ornithine with a yield of 39.5% on glucose in shake flask cultivation (Zhang et al. 2018b). It is notable that overexpression of lysE contributed to increased ornithine production (Zhang et al. 2017a). Considering that the lysine and arginine exporter LysE was shown not to accept ornithine and citrulline as substrates (Bellmann et al. 2001), the positive effect of LysE on ornithine production remains elusive.
3.4 Tryptophan
Tryptophan is one of the essential amino acids required in the diet of humans and other mammals such as pigs and poultry. Since tryptophan is particularly scarce in cereal grains, this amino acid is of considerable value for animal nutrition. Furthermore, tryptophan is known to improve the sleep state and mood as it is a precursor of serotonin which acts as a neurotransmitter in the nervous system (Bender 1985). Due to these nutritional and medicinal benefits, the amino acid has various application fields including food additives, pharmaceuticals, and feed supplements. Accordingly, the scale of the tryptophan market has expanded from about 500 tons in the year 2000 to approximately 33,000 tons in 2016 (Ajinomoto 2016). As the scale of production has increased, tryptophan prices per kilogram have dropped to around US$8 (Ajinomoto 2016).
Biosynthesis of 1 mol of tryptophan from glucose requires 1 mol each of erythrose 4-phosphate (E4P) and phosphoenolpyruvate (PEP) as starting precursors and consumes an additional 1 mol each of PEP, glutamine, phosphoribosyl-5-pyrophosphate, and serine on its biosynthetic pathways (Umbarger 1978). Therefore, a balanced supply of the precursors is required for efficient production of the amino acid. Toward this goal, a sophisticated strategy has been applied to the pathway engineering of a classically derived tryptophan-producing C. glutamicum strain (Fig. 14) (Katsumata and Ikeda 1993; Ikeda et al. 1994). This strategy consisted of amplification of the first enzyme 3-deoxy-d-arabino-heptulosonate 7-phosphate synthase in the common pathway diverging from central metabolism to increase carbon flow down that pathway, followed by sequential removal of bottlenecks discerned by the accumulation of intermediates, resulting in a 61% increase in tryptophan production and a final yield of approximately 50 g/l. The remarkable improvement involves not only systematic genetic modifications to efficiently channel carbon towards tryptophan via plasmid-mediated amplification of all together eight genes of the pathways leading to tryptophan and serine, that is, aro II, trpEGDCBA, and serA (Fig. 14), but also construction of a plasmid stabilization system based on the presence of the serA gene on the plasmid and the gene’s absence from the chromosome. Further modifications in the central metabolism to increase the availability of PEP and E4P through decreased PPC activity and increased transketolase activity, respectively, have ultimately resulted in a final titer of 58 g/l with a conversion yield of 23.2% on sucrose after 80 h of 2-L jar fermentor cultivation with no need for antibiotics (Katsumata and Kino 1989; Ikeda and Katsumata 1999). It is notable that more than half of the product crystallized in the medium.

Schematic diagram of a genetically engineered tryptophan-producing C. glutamicum strain
In addition to pathway engineering, the impact of transport engineering on tryptophan production has been demonstrated in C. glutamicum (Ikeda and Katsumata 1994, 1995). A modification leading to a decreased rate of tryptophan uptake in a tryptophan-producing mutant resulted in increased production, while accelerated tryptophan uptake drastically decreased production. Considering this, the increased capacity of tryptophan efflux can be also a promising strategy for further increased production, although the excretion process of tryptophan in C. glutamicum remains to be elucidated.
Looking beyond C. glutamicum, the tryptophan exporter YddG has been reported in E. coli (Doroshenko et al. 2007; Airich et al. 2010). The YddG protein has been shown to mediate export of not only tryptophan but also phenylalanine and tyrosine and to enhance the production of the aromatic amino acids when overexpressed in E. coli. Following this, a Chinese group has improved a classically derived E. coli tryptophan producer by stepwise modifications including prevention of tryptophan uptake (Δmtr), overexpression of yddG, and decreased acetate by-production (Δpta), resulting in a final tryptophan titer of 48.68 g/l with a conversion yield of 21.87% on glucose after 38 h of 30-L fermentor cultivation (Wang et al. 2013).
Recently, starting from a wild-type background with respect to tryptophan biosynthesis, a genetically defined tryptophan-producing E. coli strain was developed through a rational metabolic engineering process that included interruption of tryptophan degradation (ΔtnaA), disruption of tryptophan importer (Δmtr, ΔtnaB), and stepwise modifications of the terminal pathways leading to both tryptophan and serine (Chen and Zeng 2017). The resulting strain S028 produced 40.3 g/L of tryptophan with a yield of 15% on glucose after 61 h of fed-batch fermentation. Intracellular metabolite analysis of this strain suggested that availability of glutamine and export of tryptophan were likely to limit tryptophan production .
More recently, the same group has reported that tryptophan biosynthesis in some microorganisms, including E. coli, Aspergillus niger, and Saccharomyces cerevisiae, is regulated through anthranilate-associated feed-forward regulation at the indole-3-glycerol phosphate synthase step (TrpC), in addition to already-known regulations such as repression, attenuation, and feedback inhibition (Chen et al. 2018). Based on the findings, the anthranilate-activated TrpC enzyme from A. niger was expressed in the tryptophan producer S028, leading to an increase in a conversion yield on glucose from 15 to 18% in fed-batch cultivation.
3.5 Branched-Chain Amino Acids
The branched-chain amino acids valine, leucine, and isoleucine are all essential for human and animal nutrition, and all have increasing uses in various fields including pharmaceuticals, cosmetics, agricultural chemicals, dietary supplements, and feed additives. Currently, their most popular use is as a supplement for athletes to promote strength; this use is based on the nutraceutical effect of these amino acids on skeletal muscles (Shimomura et al. 2006). The intermediates for these amino acids can also be used for the production of biofuels (Atsumi et al. 2008). In C. glutamicum, all three of these amino acids share common uptake and export systems (Ebbighausen et al. 1989; Kennerknecht et al. 2002) as well as common substrates and enzymes for their biosynthesis, and thus are closely related in their metabolic fate.
Since the year 2000, rational metabolic engineering has been applied to the production of the branched-chain amino acids by C. glutamicum many times, with a special emphasis on valine production (Pátek 2007; Park and Lee 2010; Wang et al. 2018a). The strategies used to improve production of valine include (1) eliminating bottlenecks in the terminal pathway, either by conferring isoleucine auxotrophy which allows the attenuation control of the ilvBNC operon to be circumvented (Radmacher et al. 2002), by deregulating the key regulatory enzyme acetohydroxyacid synthase (Elisáková et al. 2005), or by overexpressing the gene set responsible for valine biosynthesis (Radmacher et al. 2002; Blombach et al. 2007a; Bartek et al. 2010); (2) increasing the availability of precursor pyruvate, either by blocking pantothenate synthesis (Radmacher et al. 2002; Bartek et al. 2008), by inactivating pyruvate dehydrogenase, pyruvate carboxylase, and pyruvate:quinine oxidoreductase (Blombach et al. 2007a, 2008, 2009a), or by introducing an H+-ATPase defect which contributes to the enhanced glycolysis and thus to the increased supply of pyruvate (Li et al. 2007; Wada et al. 2008); and (3) increasing NADPH supply by inactivating phosphoglucose isomerase (Blombach et al. 2008; Bartek et al. 2010). These modifications have mostly been achieved through plasmid-mediated amplification and/or deletion of the targeted genes, possibly leading to perturbations of the natural homeostatic mechanisms of the cell. To alleviate such side-effects on cell physiology, the desired metabolic engineering has been achieved through purposeful mutagenesis of promoters of the chromosomal genes involved in the valine biosynthesis pathway and in competing pathways (Holátko et al. 2009). The resulting plasmid-free valine producer was auxotrophic to pantothenate and bradytrophic to isoleucine, carried a feedback-resistant acetohydroxy acid synthase, and expressed the genes ilvD and ilvE from strong mutant promoters. This new type strain with all mutations constructed within the chromosome has been shown to produce 136 mM (15.9 g/L) valine from 4% glucose after 48 h of flask cultivation.
In addition to such conventional aerobic processes, a different bioprocess has been reported for valine production (Hasegawa et al. 2012, 2013). In this alternative process, engineering of the redox balance in combination with the use of growth-arrested packed cells has allowed C. glutamicum to produce valine at high yields under anaerobic conditions. Theoretically, the biosynthesis of 1 mol of valine from 1 mol of glucose generates 2 mol of NADH during glycolysis but requires 2 mol of NADPH at the IlvC and IlvE steps, thus causing a redox imbalance during anaerobic valine production (Fig. 15). This problem was overcome by switching the cofactor requirement of IlvC from NADPH to NADH and introducing NAD-specific exogenous leucine dehydrogenase instead of NADPH-specific IlvE (Fig. 15). Further modifications intended to reconcile redox balance with high-yield valine production and low by-product formation have ultimately resulted in a final titer of 1280 mM (150 g/l) with a conversion yield of 88% (mol/mol) or 57.2% (w/w) on glucose after 24 h under growth-arrested anaerobic conditions.

Schematic diagram of a genetically engineered C. glutamicum strain for valine production under oxygen deprivation conditions
Recently, a Chinese group has developed valine producers starting from a different C. glutamicum wild-type strain, ATCC 13869, through systematic metabolic engineering involving deletion of the three genes aceE, alaT, and ilvA and overexpression of the six genes ilvB, ilvN, ilvC, lrp 1, brnF, and brnE, resulting in a final titer of 51.2 g/l with a conversion yield of 30.8% on glucose after 96 h of 5-L fermentor cultivation (Chen et al. 2015). The same group has also found, through transcriptomic and proteomic analysis of a classically derived C. glutamicum valine-producing mutant, that the up-regulation of the genes responsible for ribosome elongation factors and ribosomal proteins is involved in valine production (Zhang et al. 2018d).
On the other hand, evolutionary approaches such as biosensor-driven screening or applications of genetic suppression have revealed non-intuitive beneficial mutations for valine production . These include a knockout mutation in the ureD gene involved in the degradation of urea to carbon dioxide and ammonium and an icd knockdown mutation allowing a metabolic shift from the TCA cycle to the glyoxylate shunt (Mahr et al. 2015; Schwentner et al. 2018). Recent developments in valine production by metabolically engineered C. glutamicum strains have been summarized in a previous review (Wang et al. 2018a).
For leucine production , rationally designed C. glutamicum strains have also been reported by a German group (Vogt et al. 2014). Leucine biosynthesis has 2-ketoisovalerate as a common substrate for valine and additionally requires a supply of acetyl-CoA in the leucine-specific pathway. Therefore, researchers have sought to expand the pathways leading to the common substrate 2-ketoisovalerate, and the targets of engineering include citrate synthase for increased availability of the precursor acetyl-CoA and the leucine-specific enzymes. Systematic metabolic engineering starting from the wild-type strain ATCC 13032 has led to an industrially potent, genetically defined, and plasmid-free leucine producer capable of a final titer of 181 mM (23.7 g/L) with a yield of 21.8% (w/w) on glucose after 72 h of fed-batch fermentation.
Metabolic engineering of C. glutamicum has also led to isoleucine producers. Isoleucine is synthesized from threonine through five enzymatic steps. In the first step, catalysed by the key enzyme threonine dehydratase (IlvA), threonine is converted into 2-ketobutyrate, which is then converted into isoleucine by four enzymes common to valine biosynthesis. Therefore, metabolic engineering involves modifications that cause increased supply of threonine and deregulation and overexpression of IlvA, as well as modifications affecting the enzymes common to valine biosynthesis (Morbach et al. 1995, 1996; Yin et al. 2012; Vogt et al. 2015; Dong et al. 2016). Modifications leading to overexpression of the global regulator Lrp and the branched-chain amino acid exporter BrnFE (Xie et al. 2012; Yin et al. 2013) and increased availability of NADPH (Shi et al. 2013; Ma et al. 2016) are also useful for efficient production of isoleucine.
Recently, a Chinese group has reported that ribosome elongation factor G and recycling factor, both of which are relevant to protein synthesis, contribute to increased isoleucine production when overexpressed (Zhao et al. 2015). Plasmid-mediated amplification of the corresponding genes fusA and frr, together with the isoleucine-biosynthetic genes ilvA, ilvB, and ilvN and the NAD kinase gene ppnk, in a classical isoleucine producer of C. glutamicum has resulted in a final titer of 28.5 g/L with a yield of 13.9% on glucose after 72 h of fed-batch fermentation.
The branched-chain amino acids are mainly used for pharmaceutical purposes and are required to have the highest degree of purity. From this perspective, attempts have been made to minimize the by-production of other amino acids to a level at which supplementary purification of the desired amino acid is not necessary. For example, the by-production of alanine occurs during valine production but can be overcome by deletion of the alanine aminotransferase gene alaT in a C. glutamicum valine producer, thereby facilitating cost-effective downstream processing (Marienhagen and Eggeling 2008). On the other hand, production of leucine is often accompanied by accumulation of valine since C. glutamicum predominantly uses the single transaminase IlvE for the synthesis of the branched-chain amino acids from the respective keto acids, thus causing co-production of the amino acids. This problem has been overcome by using different types of aminotransferases (Feng et al. 2018). Overexpression of endogenous AspB or heterologous E. coli TyrB instead of native IlvE in an isoleucine-auxotrophic leucine producer of C. glutamicum has led to leucine production with almost no by-production of valine.
3.6 Methionine
Methionine, another essential amino acid for animals, has a great deal of commercial value as a feed additive. Moreover, methionine is important as a precursor of S-adenosylmethionine. For this purpose, methionine is produced exclusively by chemical synthesis in d, l-forms, as this amino acid is considered to have a similar effect on animal nutrition in both l- and d, l-forms. Nowadays, however, there is an increasing interest in the development of environmentally friendly fermentation methods using renewable feedstocks to produce methionine.
In the hope of discovering a method for the rational construction of a methionine producer, methionine biosynthesis and its regulation are being studied in C. glutamicum. This microbe possesses both transsulfuration and direct sulfhydrylation pathways, in contrast to E. coli and most other microorganisms, which utilize only one of these two pathways (Lee and Hwang 2003; Hwang et al. 2007). Two regulatory genes in C. glutamicum have been identified as relevant to methionine biosynthesis: mcbR (cg3253) and NCgl2640. Inactivation of either in wild-type C. glutamicum results in increased methionine production (Mampel et al. 2005; Rey et al. 2003, 2005). The common strategy underlying the metabolic engineering is to redirect carbon from the lysine pathway into the methionine pathway, and all the following achievements have in principle been performed using C. glutamicum lysine producers as platforms. For example, the introduction of feedback-resistant lysC and hom genes and deletion of the thrB gene resulted in 2.9 g/L of methionine together with 23.8 g/L of lysine (Park et al. 2007b). Overexpression of the homologous metX and metY genes in another lysine-producing C. glutamicum strain was reported in a patent by Möckel et al. (2002) as resulting in a final titer of 16 g/L of methionine. Deletion of the mcbR gene and overexpression of the brnFE genes encoding an exporter for methionine (Trötschel et al. 2005) led to 6.3 g/L of methionine production after 64 h in fed-batch fermentation (Qin et al. 2015).
Recently, Li et al. (2016) have developed a C. glutamicum methionine producer through the combination of rational metabolic engineering and random mutagenesis. The procedure and its effects on methionine production are summarized as follows (Fig. 16). The first step was to abolish the reuptake of methionine, followed by random mutagenesis. Deletion of the metD locus comprising genes metQNI for the methionine uptake system (Trötschel et al. 2008) and subsequent random mutagenesis conferred methionine production of 2.54 g/L on wild-type C. glutamicum ATCC 13032 after 72 h in fed-batch fermentation. Although the mutations obtained by the random mutagenesis remain undefined, they are associated with increased expression of the genes mainly involved in methionine biosynthesis, including hom, metX, metY, metB, aecD, metE, metH, and metK. The second step was to block or weaken competitive branch pathways. Deletion of the thrB gene and replacement of the start codon ATG by the rare GTG in the dapA gene (dapA A1G) resulted in methionine production of 2.99 g/L. The third step was to enhance the precursor supply for methionine biosynthesis. Introduction of the feedback-resistant lysC gene (lysC T311I) (Ohnishi et al. 2002) and a version of the pyc gene that had undergone a C-to-T exchange at nucleotide position 1327 and a GTG-to-ATG exchange at the start codon (pyc P458S G1A) resulted in methionine production of 5.89 g/L. The pyc P458S gene is responsible for an increased supply of oxaloacetate (Ohnishi et al. 2002). The fourth step was to improve the NADPH supply. Introduction of mutant alleles of genes zwf (zwf A243T) and gnd (gnd S361F), both of which are responsible for an increased supply of NADPH (Ohnishi et al. 2005; Becker et al. 2007), achieved methionine production of 6.85 g/L after 72 h in fed-batch fermentation, which corresponds to a conversion yield of 8% (mol/mol) on glucose.

Schematic diagram of a genetically engineered methionine-producing C. glutamicum strain
In E. coli, on the other hand, an attempt at systematic metabolic engineering resulted in a strain that produces methionine at an industrially useful level (Figge et al. 2009). The key to success here was achieving a balanced supply of three important precursors for methionine biosynthesis: O-succinylhomoserine, cysteine, and the C1 carbon methyl-tetrahydrofolate (CH3-THF). An imbalanced supply of these precursors causes the formation of undesired by-products such as homolanthionine and isoleucine through the involvement of certain methionine-biosynthetic enzymes themselves. The engineered E. coli strain has achieved a yield of 19.9% after 50 h in fed-batch fermentation without the formation of any detectable undesirable by-products. Based on this yield, the methionine titer is estimated at more than 35 g/L. The procedure and impact of this metabolic engineering project can be found in the first edition of this book. Very recently, in an engineered E. coli strain based on the similar concept of balancing the supply of the three precursors, deletion of the metI gene involved in methionine uptake (Merlin et al. 2002) and overexpression of the yjeH gene encoding an exporter of methionine (Liu et al. 2015) resulted in methionine production of 17 g/L after 48 h in fed-batch fermentation (Huang et al. 2018). The fact that the final titer in this report is lower than that reported by Figge et al. (2009) appears to be attributable to insufficient optimization of the expression of the manipulated genes or the culture conditions.
Though some progress has been made toward creating improved methionine producers, methionine yields still remain low compared with those attained for other amino acids. Metabolic pathway analysis has been used to evaluate the theoretical maximum yields of methionine production on the substrates glucose, sulfate, and ammonia in C. glutamicum and E. coli (Krömer et al. 2006). The theoretical yield (mol-C methionine per mol-C glucose) of C. glutamicum was 0.49, while that of E. coli was somewhat higher at 0.52. This analysis also showed that introduction of the E. coli glycine cleavage system into C. glutamicum as an additional C1 source and the replacement of sulfate with thiosulfate or sulfide, thereby avoiding the need for reduction of oxidized sulfur, would increase the theoretical maximal methionine yields in C. glutamicum to 0.57 and 0.68, respectively. Furthermore, when methanethiol (also known as methylmercaptan) is used as a combined source for a C1 carbon and sulfur in C. glutamicum, the theoretical yield is estimated to reach its highest potential value at 0.91 (Krömer et al. 2006).
Recently, the potential utilization of methanethiol and its dimeric form dimethyldisulfide as both the C1 source and the sulfur source has been experimentally verified in C. glutamicum (Bolten et al. 2010). Isotope experiments have revealed that the S-CH3 group is entirely added to O-acetylhomoserine, directly yielding methionine (Fig. 17). This reaction has been shown to be catalyzed by MetY, creating a shortcut for methionine biosynthesis. The problem in this case would be the toxicity of these sulfur compounds to cells. A delivery system using a beaded macroporous polystyrene resin has been suggested as a potential way of alleviating the toxic effects (Bolten et al. 2010).

Proposed pathway for assimilation in C. glutamicum of methanethiol and dimethyldisulfide into methionine in addition to two known pathways of transsulfuration and direct sulfhydrylation
3.7 S-Adenosyl-Methionine
S-Adenosylmethionine (SAM) is a sulfonium compound recognized as a primary methyl donor for reactions catalyzed by methyltransferases. SAM is also required for a variety of reactions as a source of methylene groups, amino groups, ribosyl groups, aminoalkyl groups, and 5′-deoxyadenosyl radicals (Fontecave et al. 2004). In addition, SAM is also available worldwide as a drug and in the United States as a nutritional supplement (Lu and Mato 2012).
Recently, metabolic engineering for SAM production has been conducted using C. glutamicum. To achieve the following outcomes, SAM is accumulated in engineered cells; each SAM titer (g/L or mg/L) is thus provided as an intracellular concentration. The wild-type strain C. glutamicum ATCC 13032 has been reported to accumulate 0.32 g/L of SAM by the overexpression of the homologous metK gene encoding a methionine adenosyltransferase after 24 h of 72 h fed-batch culture with feeding methionine at a final concentration of 6 g/L (Han et al. 2016b). Since ATP as well as methionine is required for the MetK reaction, metabolic engineering aiming to enhance ATP supply by increasing oxygen availability for the respiratory chain has been performed using Vitreoscilla hemoglobin encoded by the vgb gene (Han et al. 2015). Overexpression of the metK and vgb genes in the isoleucine-producing strain C. glutamicum IWJ001 resulted in co-production of 0.67 g/L of SAM and 13.9 g/L of isoleucine after 72 h of fed-batch culture with feeding methionine (6 g/L), which quantities are 37-fold higher and 1.3-fold lower, respectively, than those produced by the control strain IWJ001.
Direct fermentation of SAM from glucose has also been attempted using an engineered C. glutamicum strain (Han et al. 2016a). This strategy is aimed at enhancing methionine biosynthesis and blocking competing branch pathways. The procedure and SAM production are summarized as follows (Fig. 18). First, the mcbR and thrB genes were deleted in wild type C. glutamicum ATCC 13032. This resulted in SAM accumulation of 33.2 mg/L (2.58 mg/g of dry cell weight) after 36 h on glucose in 50-mL batch culture. Second, in order to prevent homolanthionine accumulation and redirect the metabolic flux toward the direct sulfhydrylation pathway by O-acetylhomoserine sulfhydrylase encoded by the metY gene, the metB gene encoding cystathionine-γ-synthase was deleted. This resulted in SAM accumulation of 74.3 mg/L (6.12 mg/g of dry cell weight) after 36 h. Third, to improve assimilation of sulfur for methionine production using the sulfhydrylation pathway, NCgl2640, another regulatory gene for the metY gene, was deleted, which resulted in SAM accumulation of 95.4 mg/L (7.18 mg/g of dry cell weight) after 36 h. Finally, the metK and vgb genes were co-overexpressed, leading to SAM accumulation of 196.7 mg/L (12.15 mg/g of dry cell weight) after 48 h.

Schematic diagram of a genetically engineered S-adenosylmethionine-producing C. glutamicum strain. Information regarding feedback regulation is omitted here since it was already provided in Fig. 16
3.8 Cysteine
Cysteine, the other sulfur-containing amino acid, is nonessential but has a crucial function in metabolism as a precursor of sulfur-containing compounds such as methionine, thiamine, biotin, lipoic acid, and coenzyme A. In addition to its biological significance, cysteine is important commercially because of its various applications in the pharmaceutical, cosmetic, food, and livestock industries. As there is currently no efficient method of producing cysteine through fermentation, its production has depended on other methods including microbial conversion from DL-2-amino-Δ2thiazoline 4-carboxylic acid (Sano et al. 1977) and extraction from natural protein-rich resources such as hair and keratin.
As with methionine, it has been difficult to engineer strains that produce high yields of cysteine, though this amino acid is synthesized in C. glutamicum from serine via O-acetyl-serine in only two steps (Haitani et al. 2006). Typical strategies include deregulation of the key regulatory enzyme serine O-acetyltransferase, deletion of the cysteine desulfhydrase gene that catalyzes cysteine degradation to pyruvate, and overexpression of cysteine exporters; these methods have been shown to increase cysteine production in both E. coli and C. glutamicum, though the final titers were below 2 g/L (Wada et al. 2002; Wada and Takagi 2006). It has been suggested that the combination of these strategies and the improvement of other factors including an increased supply of the precursor serine and a decreased reuptake of the product would lead to further improvement (Wada and Takagi 2006).
In C. glutamicum, the transcriptional regulator CysR, whose gene is repressed by McbR, activates the expression of the genes involved in assimilatory sulfate reduction and of the other regulatory gene ssuR (Rückert et al. 2008). Subsequently, SsuR activates the expression of the genes involved in sulfonate utilization (Koch et al. 2005). Therefore, overexpression of the cysR gene enables the simultaneous expression of many genes associated with the assimilatory reduction of sulfur source. Overexpression of the cysE, cysK, and cysR genes leads to intracellular accumulation of approximately 60 mg/L of cysteine on glucose after 15 h of 50-mL batch culture (Joo et al. 2017). In a more advanced study using C. glutamicum, deletion of aecD and sdaA, two genes that are involved in cysteine and serine degradation, respectively, along with reduction in the expression level of the glyA gene and overexpression of the feedback-insensitive cysE gene, the cysK gene, the E. coli bcr gene involved in cysteine export, and the feedback-insensitive serA gene together with the serCB genes, allowed the wild-type strain to produce 950 mg/L of cysteine with a yield of 2.73% on glucose after 36 h batch cultivation in a medium containing 6 g/L of thiosulfate (Wei et al. 2018).
Recently, in E. coli, a novel CysM (O-acetylserine sulfhydrylase B)-independent thiosulfate assimilation pathway was identified. This novel route, specifically a thiosulfate sulfurtransferase (GlpE)-mediated bypass from thiosulfate to sulfite, was evaluated for cysteine production (Kawano et al. 2017). Whereas overexpression of the feedback-insensitive serA and cysE genes together with the native ydeD gene involved in efflux of cysteine allowed a wild-type E. coli strain to produce approximately 1.0 g/L of cysteine on glycerol after 72 h in an experiment where sulfate and thiosulfate were added as sulfur sources, the addition of glpE overexpression achieved cysteine production of 1.5 g/L under the same conditions (Kawano et al. 2017). Very recently, it has been reported in E. coli that overexpression of the feedback-insensitive cysE gene, the ydeD gene, and the feedback-insensitive serA gene along with the native serCB genes, as well as deletion of the sdaA and tnaA genes, which are responsible for degradation of serine and cysteine, respectively, resulted in cysteine production of 5.1 g/L after 32 h on glucose in fed-batch culture (Liu et al. 2018b).
In recent years, Pantoea ananatis and closely related species have received attention in the fermentation industry for their potential to overproduce a wide variety of useful chemicals (Takumi et al. 2017). Metabolic engineering for cysteine production has been performed using P. ananatis strain SC17 (Fig. 19). This was initiated by the following manipulations (Takumi and Nonaka 2014). First, to deregulate the cysteine biosynthetic pathway, a mutant allele of the E. coli cysE gene that encodes a feedback-resistant serine O-acetyltransferase was overexpressed. Simultaneously, to enhance the efflux of cysteine, the E. coli leuE encoding LeuE, which was originally identified as a leucine efflux pump (Kutukova et al. 2005) but which also exhibits cysteine efflux activity (Takumi and Nonaka 2014), was overexpressed. Second, to enhance thiosulfate uptake, the intrinsic cysPUWA genes encoding the thiosulfate transporter CysPUWA, a bottleneck in the intracellular sulfur supply pathway (Sirko et al. 1990), were overexpressed. Finally, to deregulate the biosynthetic pathway of serine that is the precursor of cysteine, a mutant allele of the homologous serA gene that encodes a feedback-resistant 3-phosphoglycerate dehydrogenase was overexpressed. Although the cysteine productivity of each intermediate strain derived during the above process has not been disclosed, the assembly of all these modifications to strain SC17 resulted in strain AG4854, which is capable of cysteine production of 1.3 g/L on glucose after 16 h cultivation with an estimated conversion yield of 3.1% on glucose (Takumi et al. 2017). While engineering of the transcriptional regulator CysB, a master regulator that induces most genes involved in sulfur assimilation and cysteine metabolism, is assumed to be beneficial to cysteine production as it simultaneously enhances the metabolism of sulfur and cysteine, it has a disadvantage in that it causes the expression of the ydjN gene encoding the cystine uptake transporter, resulting in reuptake of cysteine in the form of cystine, which is formed from cysteine in an oxidative environment (Nonaka 2018).

Schematic diagram of a genetically engineered cysteine-producing P. ananatis strain. Asterisked genes encode enzymes resistant to feedback inhibition. Deletion of the ccdA gene (1), overexpression of the cefA gene (2), or overexpression of the cefB gene (3) was conducted in strain AG4854 where CysM activity was enhanced as described in the text
Based on the finding that CysM is the bottleneck of cysteine production , continued research has demonstrated that conferring moderately and excessively enhanced CysM activity on strain AG4854 (which created strains AG6181 and AG6184, respectively) resulted in yields of 3.5% and 0.4%, respectively. The failure of strain AG6184 to produce cysteine was accompanied by increased activity of cysteine-inducible CcdA (cysteine desulfhydrase), the only major cysteine degradation enzyme in P. ananatis (Takumi and Nonaka 2016), which is presumably induced by the elevated intracellular cysteine levels resulting from the increased activity of CysM. Disruption of the ccdA gene improved the yields of strains AG6181 and AG6184 to 3.9% and 1.6%, respectively, but had negative effects on growth and glucose consumption, which were especially severe in strain AG6184, presumably due to the toxicity of intracellularly accumulated cysteine. An effort to elicit only positive effects from CysM overexpression on cysteine production was made using the cysteine efflux pumps CefA (Takumi and Nonaka 2016) and CefB (Takumi et al. 2017). Yet without the implementation of fine-tuned expression, overexpression of the cefA gene in AG6184 led to cysteine production of approximately 0.4 g/L with an estimated yield of approximately 1.0% after 28 h cultivation, whereas overexpression of the cefB gene achieved cysteine production of approximately 2.2 g/L with an estimated yield of approximately 5.5% after 22 h cultivation (Takumi et al. 2017). A combination of ccdA deletion and cefA and/or cefB overexpression was not conducted in this report. Although not all details of this experiment were given, metabolic engineering of P. ananatis has achieved cysteine production of close to 5.0 g/L (Nonaka 2018).
4 Conclusion and Future Prospects
As already mentioned, the global amino acid market per year has expanded to more than seven million tons and over US$13 billion in response to increased demand for amino acids. This market growth is expected to continue due to the ongoing increases in the nutritional values of amino acids and the growth of the numerous fields that use them. The giant market has been underpinned largely by recent advances in amino acid fermentation technologies, especially strain development technology.
In the history of amino acid fermentation, the determination of the complete genome sequence of C. glutamicum was obviously an important milestone. The subsequent rapid progress in genomics, various “omics” technologies, and systems biology for this bacterium have dramatically transformed our approaches to strain development. For example, in-silico modeling and simulation approaches are now used routinely to help identify new targets for further engineering and strain improvement. The power of such systems-level approaches will surely increase as modeling is combined with the ever-accumulating “omics” data.
It should be noted, however, that not all purely rational approaches from scratch have necessarily resulted in commercially potent production strains, probably due to the existence of unknown mechanisms affecting industrially important properties, such as hyperproduction and high adaptability to large-scale processes. This means that there is a great deal more to learn from the genomes of classical strains. Since the dawn of the genomic era, new possibilities have emerged, including analysis of producer’s genomes, leading to the genome breeding approach, and systems metabolic engineering , leading to tailor-made cell factories with designed properties (Becker and Wittmann 2015; Ikeda 2017). The next-generation strains are expected to be created through the synergy of these approaches and through integration of the knowledge accumulated over decades of industrial strain development with emerging technologies such as biosensor-driven single cell screening , in-silico modeling, and carbon flux simulation.
At the same time, the amino acid industry is beginning to consider sustainable and environmentally-friendly manufacturing systems in response to the continuing crisis of global warming. From this standpoint, the industry is expected to develop strains enabling the use of feedstocks that are renewable and that do not compete with human food or energy sources. The development of innovative technologies enabling reduction in effluents and wastes generated during fermentation and purification processes is also expected. These remain important themes for future engineering.
References
Airich LG, Tsyrenzhapova IS, Vorontsova OV, Feofanov AV, Doroshenko VG, Mashko SV (2010) Membrane topology analysis of the Escherichia coli aromatic amino acid efflux protein YddG. J Mol Microbiol Biotechnol 19:189–197
Ajinomoto (2007) Fact sheets: amino acids business and feed-use amino acids business. http://www.ajinomoto.com/ar/i_r/pdf/fact/Aminoacids-Oct2007.pdf, http://www.ajinomoto.com/ar/i_r/pdf/fact/Feed-useAA-Oct2007.pdf
Ajinomoto (2016) FY2015 market and other information. https://www.ajinomoto.com/en/ir/event/presentation/main/09/teaserItems1/00/linkList/02/link/FY15_Data_E.pdf
Anaya-Reza O, Lopez-Arenas T (2017) Comprehensive assessment of the L-lysine production process from fermentation of sugarcane molasses. Bioprocess Biosyst Eng 40:1033–1048
Appleton J (2002) Arginine: clinical potential of a semi-essential amino acid. Altern Med Rev 7:512–522
Asakura Y, Kimura E, Usuda Y, Kawahara Y, Matsui K, Osumi T, Nakamatsu T (2007) Altered metabolic flux due to deletion of odhA causes L-glutamate overproduction in Corynebacterium glutamicum. Appl Environ Microbiol 73:1308–1319
Atsumi S, Hanai T, Liao JC (2008) Non-fermentative pathways for synthesis of branched-chain higher alcohols as biofuels. Nature 451:86–89
Barrett E, Stanton C, Zelder O, Fitzgerald G, Ross RP (2004) Heterologous expression of lactose- and galactose-utilizing pathways from lactic acid bacteria in Corynebacterium glutamicum for production of lysine in whey. Appl Environ Microbiol 70:2861–2866
Bartek T, Makus P, Klein B, Lang S, Oldiges M (2008) Influence of L-isoleucine and pantothenate auxotrophy for L-valine formation in Corynebacterium glutamicum revisited by metabolome analyses. Bioprocess Biosyst Eng 31:217–225
Bartek T, Blombach B, Zönnchen E, Makus P, Lang S, Eikmanns BJ, Oldiges M (2010) Importance of NADPH supply for improved L-valine formation in Corynebacterium glutamicum. Biotechnol Prog 26:361–371
Becker J, Wittmann C (2015) Advanced biotechnology: metabolically engineered cells for the bio-based production of chemicals and fuels, materials, and health-care products. Angew Chem Int Ed Engl 54:3328–3350
Becker J, Klopprogge C, Zelder O, Heinzle E, Wittmann C (2005) Amplified expression of fructose 1,6-bisphosphatase in Corynebacterium glutamicum increases in vivo flux through the pentose phosphate pathway and lysine production on different carbon sources. Appl Environ Microbiol 71:8587–8596
Becker J, Klopprogge C, Herold A, Zelder O, Bolten CJ, Wittmann C (2007) Metabolic flux engineering of L-lysine production in Corynebacterium glutamicum - over expression and modification of G6P dehydrogenase. J Biotechnol 132:99–109
Becker J, Klopprogge C, Schröder H, Wittmann C (2009) Metabolic engineering of the tricarboxylic acid cycle for improved lysine production by Corynebacterium glutamicum. Appl Environ Microbiol 75:7866–7869
Becker J, Zelder O, Häfner S, Schröder H, Wittmann C (2011) From zero to hero: design-based systems metabolic engineering of Corynebacterium glutamicum for L-lysine production. Metab Eng 13:159–168
Becker J, Gießelmann G, Hoffmann SL, Wittmann C (2018) Corynebacterium glutamicum for sustainable bioproduction: from metabolic physiology to systems metabolic engineering. Adv Biochem Eng Biotechnol 162:217–263
Bellmann A, Vrljić M, Pátek M, Sahm H, Krämer R, Eggeling L (2001) Expression control and specificity of the basic amino acid exporter LysE of Corynebacterium glutamicum. Microbiology 147:1765–1774
Bender DA (1985) Amino acid metabolism, 2nd edn. Wiley, New York
Binder S, Schendzielorz G, Stäbler N, Krumbach K, Hoffmann K, Bott M, Eggeling L (2012) A high-throughput approach to identify genomic variants of bacterial metabolite producers at the single-cell level. Genome Biol 13:R40
Blombach B, Schreiner ME, Holátko J, Bartek T, Oldiges M, Eikmanns BJ (2007a) L-valine production with pyruvate dehydrogenase complex-deficient Corynebacterium glutamicum. Appl Environ Microbiol 73:2079–2084
Blombach B, Schreiner ME, Moch M, Oldiges M, Eikmanns BJ (2007b) Effect of pyruvate dehydrogenase complex deficiency on L-lysine production with Corynebacterium glutamicum. Appl Microbiol Biotechnol 76:615–623
Blombach B, Schreiner ME, Bartek T, Oldiges M, Eikmanns BJ (2008) Corynebacterium glutamicum tailored for high-yield L-valine production. Appl Microbiol Biotechnol 79:471–479
Blombach B, Arndt A, Auchter M, Eikmanns BJ (2009a) L-valine production during growth of pyruvate dehydrogenase complex-deficient Corynebacterium glutamicum in the presence of ethanol or by inactivation of the transcriptional regulator SugR. Appl Environ Microbiol 75:1197–1200
Blombach B, Hans S, Bathe B, Eikmanns BJ (2009b) Acetohydroxyacid synthase, a novel target for improvement of L-lysine production by Corynebacterium glutamicum. Appl Environ Microbiol 75:419–427
Bolten CJ, Schröder H, Dickschat J, Wittmann C (2010) Towards methionine overproduction in Corynebacterium glutamicum: methanethiol and dimethyldisulfide as reduced sulfur sources. J Microbiol Biotechnol 20:1196–1203
Bommareddy RR, Chen Z, Rappert S, Zeng AP (2014) A de novo NADPH generation pathway for improving lysine production of Corynebacterium glutamicum by rational design of the coenzyme specificity of glyceraldehyde 3-phosphate dehydrogenase. Metab Eng 25:30–37
Börngen K, Battle AR, Möker N, Morbach S, Marin K, Martinac B, Krämer R (2010) The properties and contribution of the Corynebacterium glutamicum MscS variant to fine-tuning of osmotic adaptation. Biochim Biophys Acta 1798:2141–2149
Bott M, Niebisch A (2003) The respiratory chain of Corynebacterium glutamicum. J Biotechnol 104:129–153
Brockmann-Gretza O, Kalinowski J (2006) Global gene expression during stringent response in Corynebacterium glutamicum in presence and absence of the rel gene encoding (p)ppGpp synthase. BMC Genomics 7:230
Burkovski A (2008) Corynebacteria: genomics and molecular biology. Caister Academic, Norfolk
Burkovski A, Krämer R (2002) Bacterial amino acid transport proteins: occurrence, functions, and significance for biotechnological applications. Appl Microbiol Biotechnol 58:265–274
Chen L, Zeng AP (2017) Rational design and metabolic analysis of Escherichia coli for effective production of L-tryptophan at high concentration. Appl Microbiol Biotechnol 101:559–568
Chen Z, Bommareddy RR, Frank D, Rappert S, Zeng AP (2014) Deregulation of feedback inhibition of phosphoenolpyruvate carboxylase for improved lysine production in Corynebacterium glutamicum. Appl Environ Microbiol 80:1388–1393
Chen C, Li Y, Hu J, Dong X, Wang X (2015) Metabolic engineering of Corynebacterium glutamicum ATCC13869 for L-valine production. Metab Eng 29:66–75
Chen L, Chen M, Ma C, Zeng AP (2018) Discovery of feed-forward regulation in L-tryptophan biosynthesis and its use in metabolic engineering of E. coli for efficient tryptophan bioproduction. Metab Eng 47:434–444
Cheng J, Chen P, Song A, Wang D, Wang Q (2018) Expanding lysine industry: industrial biomanufacturing of lysine and its derivatives. J Ind Microbiol Biotechnol 45:719–734
Chinen A, Kozlov YI, Hara Y, Izui H, Yasueda H (2007) Innovative metabolic pathway design for efficient L-glutamate production by suppressing CO2 emission. J Biosci Bioeng 103:262–269
Curis E, Crenn P, Cynober L (2007) Citrulline and the gut. Curr Opin Clin Nutr Metab Care 10:620–626
Dai Z, Nielsen J (2015) Advancing metabolic engineering through systems biology of industrial microorganisms. Curr Opin Biotechnol 36:8–15
Dong X, Quinn PJ, Wang X (2011) Metabolic engineering of Escherichia coli and Corynebacterium glutamicum for the production of L-threonine. Biotechnol Adv 29:11–23
Dong X, Zhao Y, Zhao J, Wang X (2016) Characterization of aspartate kinase and homoserine dehydrogenase from Corynebacterium glutamicum IWJ001 and systematic investigation of L-isoleucine biosynthesis. J Ind Microbiol Biotechnol 43:873–885
Doroshenko V, Airich L, Vitushkina M, Kolokolova A, Livshits V, Mashko S (2007) YddG from Escherichia coli promotes export of aromatic amino acids. FEMS Microbiol Lett 275:312–318
Ebbighausen H, Weil B, Krämer R (1989) Transport of branched-chain amino acids in Corynebacterium glutamicum. Arch Microbiol 151:238–244
Eggeling L, Bott M (2005) Handbook of Corynebacterium glutamicum. CRC, Boca Raton
Eggeling L, Bott M (2015) A giant market and a powerful metabolism: L-lysine provided by Corynebacterium glutamicum. Appl Microbiol Biotechnol 99:3387–3394
Elisáková V, Pátek M, Holátko J, Nesvera J, Leyval D, Goergen JL, Delaunay S (2005) Feedback-resistant acetohydroxy acid synthase increases valine production in Corynebacterium glutamicum. Appl Environ Microbiol 71:207–213
Feng LY, Xu JZ, Zhang WG (2018) Improved L-leucine production in Corynebacterium glutamicum by optimizing the aminotransferases. Molecules 23:2102
Figge R, Soucaille P, Barbier G, Bestel-Corre G, Boisart C, Chateau M (2009) Increasing methionine yield. International Patent Application WO 2009/043803 A2
Fontecave M, Atta M, Mulliez E (2004) S-adenosylmethionine: nothing goes to waste. Trends Biochem Sci 29:243–249
Glansdorff N, Xu Y (2007) Microbial arginine biosynthesis: pathway, regulation and industrial production. In: Wendisch VF (ed) Microbiology monographs, amino acid biosynthesis - pathways, regulation and metabolic engineering. Springer, Berlin, pp 219–257
Global Market Insights (2016) Glutamic acid and monosodium glutamate (MSG) market size, potential, industry outlook, regional analysis, application development, competitive landscape & forecast, 2016–2023. https://www.gminsights.com/industry-analysis/glutamic-acid-and-monosodium-glutamate-msg-market
Gutmann M, Hoischen C, Krämer R (1992) Carrier-mediated glutamate secretion by Corynebacterium glutamicum under biotin limitation. Biochim Biophys Acta 1112:115–123
Haitani Y, Awano N, Yamazaki M, Wada M, Nakamori S, Takagi H (2006) Functional analysis of L-serine O-acetyltransferase from Corynebacterium glutamicum. FEMS Microbiol Lett 255:156–163
Han G, Hu X, Wang X (2015) Co-production of S-adenosyl-L-methionine and L-isoleucine in Corynebacterium glutamicum. Enzym Microb Technol 78:27–33
Han G, Hu X, Qin T, Li Y, Wang X (2016a) Metabolic engineering of Corynebacterium glutamicum ATCC 13032 to produce S-adenosyl-L-methionine. Enzym Microb Technol 83:14–21
Han G, Hu X, Wang X (2016b) Overexpression of methionine adenosyltransferase in Corynebacterium glutamicum for production of S-adenosyl-L-methionine. Biotechnol Appl Biochem 63:679–689
Hara Y, Kadotani N, Izui H, Katashkina JI, Kuvaeva TM, Andreeva IG, Golubeva LI, Malko DB, Makeev VJ, Mashko SV, Kozlov YI (2012) The complete genome sequence of Pantoea ananatis AJ13355, an organism with great biotechnological potential. Appl Microbiol Biotechnol 93:331–341
Hasegawa S, Uematsu K, Natsuma Y, Suda M, Hiraga K, Jojima T, Inui M, Yukawa H (2012) Improvement of the redox balance increases L-valine production by Corynebacterium glutamicum under oxygen deprivation conditions. Appl Environ Microbiol 78:865–875
Hasegawa S, Suda M, Uematsu K, Natsuma Y, Hiraga K, Jojima T, Inui M, Yukawa H (2013) Engineering of Corynebacterium glutamicum for high-yield L-valine production under oxygen deprivation conditions. Appl Environ Microbiol 79:1250–1257
Hashimoto K, Nakamura K, Kuroda T, Yabe I, Nakamatsu T, Kawasaki H (2010) The protein encoded by NCgl1221 in Corynebacterium glutamicum functions as a mechanosensitive channel. Biosci Biotechnol Biochem 74:2546–2549
Hashimoto K, Murata J, Konishi T, Yabe I, Nakamatsu T, Kawasaki H (2012) Glutamate is excreted across the cytoplasmic membrane through the NCgl1221 channel of Corynebacterium glutamicum by passive diffusion. Biosci Biotechnol Biochem 76:1422–1424
Hayashi T, Juliet PA, Matsui-Hirai H, Miyazaki A, Fukatsu A, Funami J, Iguchi A, Ignarro LJ (2005) L-Citrulline and L-arginine supplementation retards the progression of high-cholesterol-diet-induced atherosclerosis in rabbits. Proc Natl Acad Sci U S A 102:13681–13686
Hayashi M, Mizoguchi H, Ohnishi J, Mitsuhashi S, Yonetani Y, Hashimoto S, Ikeda M (2006a) A leuC mutation leading to increased L-lysine production and rel-independent global expression changes in Corynebacterium glutamicum. Appl Microbiol Biotechnol 72:783–789
Hayashi M, Ohnishi J, Mitsuhashi S, Yonetani Y, Hashimoto S, Ikeda M (2006b) Transcriptome analysis reveals global expression changes in an industrial L-lysine producer of Corynebacterium glutamicum. Biosci Biotechnol Biochem 70:546–550
Hayashi T, Matsui-Hirai H, Miyazaki-Akita A, Fukatsu A, Funami J, Ding QF, Kamalanathan S, Hattori Y, Ignarro LJ, Iguchi A (2006c) Endothelial cellular senescence is inhibited by nitric oxide: implications in atherosclerosis associated with menopause and diabetes. Proc Natl Acad Sci U S A 103:17018–17023
Hermann T (2003) Industrial production of amino acids by coryneform bacteria. J Biotechnol 104:155–172
Hirao T, Nakano T, Azuma T, Sugimoto M, Nakanishi T (1989) L-lysine production in continuous culture of an L-lysine hyper- producing mutant of Corynebacterium glutamicum. Appl Microbiol Biotechnol 32:269–273
Hirasawa T, Shimizu H (2016) Recent advances in amino acid production by microbial cells. Curr Opin Biotechnol 42:133–146
Hoffmann SL, Jungmann L, Schiefelbein S, Peyriga L, Cahoreau E, Portais JC, Becker J, Wittmann C (2018) Lysine production from the sugar alcohol mannitol: design of the cell factory Corynebacterium glutamicum SEA-3 through integrated analysis and engineering of metabolic pathway fluxes. Metab Eng 47:475–487
Holátko J, Elisáková V, Prouza M, Sobotka M, Nesvera J, Pátek M (2009) Metabolic engineering of the L-valine biosynthesis pathway in Corynebacterium glutamicum using promoter activity modulation. J Biotechnol 139:203–210
Hua K (2013) Investigating the appropriate mode of expressing lysine requirement of fish through non-linear mixed model analysis and multilevel analysis. Br J Nutr 109:1013–1021
Huang Y, Zhang H, Tian H, Li C, Han S, Lin Y, Zheng S (2015) Mutational analysis to identify the residues essential for the inhibition of N-acetyl glutamate kinase of Corynebacterium glutamicum. Appl Microbiol Biotechnol 99:7527–7537
Huang Y, Li C, Zhang H, Liang S, Han S, Lin Y, Yang X, Zheng S (2016) Monomeric Corynebacterium glutamicum N-acetyl glutamate kinase maintains sensitivity to L-arginine but has a lower intrinsic catalytic activity. Appl Microbiol Biotechnol 100:1789–1798
Huang JF, Shen ZY, Mao QL, Zhang XM, Zhang B, Wu JS, Liu ZQ, Zheng YG (2018) Systematic analysis of bottlenecks in a multibranched and multilevel regulated pathway: the molecular fundamentals of L-methionine biosynthesis in Escherichia coli. ACS Synth Biol 7:2577–2589
Hwang BJ, Park SD, Kim Y, Kim P, Lee HS (2007) Biochemical analysis on the parallel pathways of methionine biosynthesis in Corynebacterium glutamicum. J Microbiol Biotechnol 17:1010–1017
Ikeda M (2003) Amino acid production processes. In: Faurie R, Thommel J (eds) Advances in biochemical engineering/biotechnology, Microbial production of L-amino acids, vol 79. Springer, Berlin, pp 1–35
Ikeda M (2012) Sugar transport systems in Corynebacterium glutamicum: features and applications to strain development. Appl Microbiol Biotechnol 96:1191–1200
Ikeda M (2017) Lysine fermentation: history and genome breeding. In: Yokota A, Ikeda M (eds) Advances in biochemical engineering/biotechnology, Amino acid fermentation, vol 159. Springer, Japan, pp 73–102
Ikeda M, Katsumata R (1994) Transport of aromatic amino acids and its influence on overproduction of the amino acids in Corynebacterium glutamicum. J Ferment Bioeng 78:420–425
Ikeda M, Katsumata R (1995) Tryptophan production by transport mutants of Corynebacterium glutamicum. Biosci Biotechnol Biochem 59:1600–1602
Ikeda M, Katsumata R (1999) Hyperproduction of tryptophan by Corynebacterium glutamicum with the modified pentose phosphate pathway. Appl Environ Microbiol 65:2497–2502
Ikeda M, Nakagawa S (2003) The Corynebacterium glutamicum genome: features and impacts on biotechnological process. Appl Microbiol Biotechnol 62:99–109
Ikeda M, Takeno S (2013) Amino acid production by Corynebacterium glutamicum. In: Yukawa H, Inui M (eds) Microbiology monographs, vol 23. Springer, Berlin, pp 107–147
Ikeda M, Nakanishi K, Kino K, Katsumata R (1994) Fermentative production of tryptophan by a stable recombinant strain of Corynebacterium glutamicum with a modified serine-biosynthetic pathway. Biosci Biotech Biochem 58:674–678
Ikeda M, Ohnishi J, Mitsuhashi S (2005) Genome breeding of an amino acid-producing Corynebacterium glutamicum mutant. In: Barredo JLS (ed) Microbial processes and products. Humana Press, Totowa, pp 179–189
Ikeda M, Ohnishi J, Hayashi M, Mitsuhashi S (2006) A genome-based approach to create a minimally mutated Corynebacterium glutamicum strain for efficient L-lysine production. J Ind Microbiol Biotechnol 33:610–615
Ikeda M, Mitsuhashi S, Tanaka K, Hayashi M (2009) Reengineering of a Corynebacterium glutamicum L-arginine and L-citrulline producer. Appl Environ Microbiol 75:1635–1641
Ikeda M, Nakano T, Mitsuhashi S, Hayashi M, Tanaka K (2010a) Process for producing L-arginine, L-ornithine or L-citrulline. US Patent 7741081B2
Ikeda M, Takeno S, Mizuno Y, Mitsuhashi S (2010b) Process for producing useful substance. International Patent Application WO 2010/024267 A1
Ikeda M, Mizuno Y, Awane S, Hayashi M, Mitsuhashi S, Takeno S (2011) Identification and application of a different glucose uptake system that functions as an alternative to the phosphotransferase system in Corynebacterium glutamicum. Appl Microbiol Biotechnol 90:1443–1451
Ikeda M, Noguchi N, Ohshita M, Senoo A, Mitsuhashi S, Takeno S (2015) A third glucose uptake bypass in Corynebacterium glutamicum ATCC 31833. Appl Microbiol Biotechnol 99:2741–2750
Izui H, Moriya M, Hirano S, Hara Y, Ito H, Matsui K (2006) Method for producing L-glutamic acid by fermentation accompanied by precipitation. US Patent 7015010B1
Jensen JV, Eberhardt D, Wendisch VF (2015) Modular pathway engineering of Corynebacterium glutamicum for production of the glutamate-derived compounds ornithine, proline, putrescine, citrulline, and arginine. J Biotechnol 214:85–94
Jiang LY, Chen SG, Zhang YY, Liu JZ (2013a) Metabolic evolution of Corynebacterium glutamicum for increased production of L-ornithine. BMC Biotechnol 13:47
Jiang LY, Zhang YY, Li Z, Liu JZ (2013b) Metabolic engineering of Corynebacterium glutamicum for increasing the production of L-ornithine by increasing NADPH availability. J Ind Microbiol Biotechnol 40:1143–1151
Joo YC, Hyeon JE, Han SO (2017) Metabolic design of Corynebacterium glutamicum for production of L-cysteine with consideration of sulfur-supplemented animal feed. J Agric Food Chem 65:4698–4707
Kabus A, Georgi T, Wendisch VF, Bott M (2007a) Expression of the Escherichia coli pntAB genes encoding a membrane-bound transhydrogenase in Corynebacterium glutamicum improves L-lysine formation. Appl Microbiol Biotechnol 75:47–53
Kabus A, Niebisch A, Bott M (2007b) Role of cytochrome bd oxidase from Corynebacterium glutamicum in growth and lysine production. Appl Environ Microbiol 73:861–868
Kalinowski J, Bathe B, Bartels D, Bischoff N, Bott M, Burkovski A, Dusch N, Eggeling L, Eikmanns BJ, Gaigalat L, Goesmann A, Hartmann M, Huthmacher K, Krämer R, Linke B, McHardy AC, Meyer F, Möckel B, Pfefferle W, Pühler A, Rey DA, Rückert C, Rupp O, Sahm H, Wendisch VF, Wiegräbe I, Tauch A (2003) The complete Corynebacterium glutamicum ATCC 13032 genome sequence and its impact on the production of L-aspartate-derived amino acids and vitamins. J Biotechnol 104:5–25
Kallio PT, Kim DJ, Tsai PS, Bailey JE (1994) Intracellular expression of Vitreoscilla hemoglobin alters Escherichia coli energy metabolism under oxygen-limited conditions. Eur J Biochem 219:201–208
Kanehisa M, Goto S, Sato Y, Kawashima M, Furumichi M, Tanabe M (2014) Data, information, knowledge and principle: back to metabolism in KEGG. Nucleic Acids Res 42:D199–D205
Kase H, Nakayama K (1974) Mechanism of L-threonine and L-lysine production by analog-resistant mutants of Corynebacterium glutamicum. Agr Biol Chem 38:993–1000
Katsumata R, Ikeda M (1993) Hyperproduction of tryptophan in Corynebacterium glutamicum by pathway engineering. Bio/Technology 11:921–925
Katsumata R, Kino K (1989) Process for producing amino acids by fermentation. Japan Patent 01,317,395 A (P2,578,488)
Kawaguchi H, Vertès AA, Okino S, Inui M, Yukawa H (2006) Engineering of a xylose metabolic pathway in Corynebacterium glutamicum. Appl Environ Microbiol 72:3418–3428
Kawaguchi H, Sasaki M, Vertès AA, Inui M, Yukawa H (2008) Engineering of an L-arabinose metabolic pathway in Corynebacterium glutamicum. Appl Microbiol Biotechnol 77:1053–1062
Kawahara Y, Takahashi-Fuke K, Shimizu E, Nakamatsu T, Nakamori S (1997) Relationship between the glutamate production and the activity of 2-oxoglutarate dehydrogenase in Brevibacterium lactofermentum. Biosci Biotechnol Biochem 61:1109–1112
Kawano Y, Onishi F, Shiroyama M, Miura M, Tanaka N, Oshiro S, Nonaka G, Nakanishi T, Ohtsu I (2017) Improved fermentative L-cysteine overproduction by enhancing a newly identified thiosulfate assimilation pathway in Escherichia coli. Appl Microbiol Biotechnol 101:6879–6889
Kelle R, Hermann T, Bathe B (2005) L-Lysine production. In: Eggeling L, Bott M (eds) Handbook of Corynebacterium glutamicum. CRC, Boca Raton, pp 465–488
Kennerknecht N, Sahm H, Yen MR, Patek M, Saier MH Jr, Eggeling L (2002) Export of L-isoleucine from Corynebacterium glutamicum: a two-gene-encoded member of a new translocator family. J Bacteriol 184:3947–3956
Kiefer P, Heinzle E, Zelder O, Wittmann C (2004) Comparative metabolic flux analysis of lysine-producing Corynebacterium glutamicum cultured on glucose or fructose. Appl Environ Microbiol 70:229–239
Kim J, Fukuda H, Hirasawa T, Nagahisa K, Nagai K, Wachi M, Shimizu H (2009a) Requirement of de novo synthesis of the OdhI protein in penicillin-induced glutamate production by Corynebacterium glutamicum. Appl Microbiol Biotechnol 86(3):911–920. https://doi.org/10.1007/s00253-009-2360-6
Kim J, Hirasawa T, Sato Y, Nagahisa K, Furusawa C, Shimizu H (2009b) Effect of odhA overexpression and odhA antisense RNA expression on Tween-40-triggered glutamate production by Corynebacterium glutamicum. Appl Microbiol Biotechnol 81:1097–1106
Kim HI, Nam JY, Cho JY, Lee CS, Park YJ (2013) Next-generation sequencing-based transcriptome analysis of L-lysine-producing Corynebacterium glutamicum ATCC 21300 strain. J Microbiol 51:877–880
Kim SY, Lee J, Lee SY (2015) Metabolic engineering of Corynebacterium glutamicum for the production of L-ornithine. Biotechnol Bioeng 112:416–421
Kimura E (2003) Metabolic engineering of glutamate production. In: Faurie R, Thommel J (eds) Advances in biochemical engineering/biotechnology, Microbial production of L-amino acids, vol 79. Springer, Berlin, pp 37–58
Kjeldsen KR, Nielsen J (2009) In silico genome-scale reconstruction and validation of the Corynebacterium glutamicum metabolic network. Biotechnol Bioeng 102:583–597
Kobayashi M, Itoyama T, Mitani Y, Usui N (2011) Method for producing basic amino acid. European Patent 1182261 B1
Koch DJ, Rückert C, Albersmeier A, Hüser AT, Tauch A, Pühler A, Kalinowski J (2005) The transcriptional regulator SsuR activates expression of the Corynebacterium glutamicum sulphonate utilization genes in the absence of sulphate. Mol Microbiol 58:480–494
Komati Reddy G, Lindner SN, Wendisch VF (2015) Metabolic engineering of an ATP-neutral Embden-Meyerhof-Parnas pathway in Corynebacterium glutamicum: growth restoration by an adaptive point mutation in NADH dehydrogenase. Appl Environ Microbiol 81:1996–2005
Krömer JO, Sorgenfrei O, Klopprogge K, Heinzle E, Wittmann C (2004) In-depth profiling of lysine-producing Corynebacterium glutamicum by combined analysis of the transcriptome, metabolome, and fluxome. J Bacteriol 186:1769–1784
Krömer JO, Wittmann C, Schröder H, Heinzle E (2006) Metabolic pathway analysis for rational design of L-methionine production by Escherichia coli and Corynebacterium glutamicum. Metab Eng 8:353–369
Krömer JO, Bolten CJ, Heinzle E, Schröder H, Wittmann C (2008) Physiological response of Corynebacterium glutamicum to oxidative stress induced by deletion of the transcriptional repressor McbR. Microbiology 154:3917–3930
Kubota T, Tanaka Y, Takemoto N, Hiraga K, Yukawa H, Inui M (2015) Identification and expression analysis of a gene encoding a shikimate transporter of Corynebacterium glutamicum. Microbiology 161:254–263
Kutukova EA, Livshits VA, Altman IP, Ptisyn LR, Zyiatdinov MH, Tokmakova IL, Zakataeva NP (2005) The yeaS (leuE) gene of Escherichia coli encodes an exporter of leucine, and the Lrp protein regulates its expression. FEBS Lett 579:4629–4634
Lange C, Mustafi N, Frunzke J, Kennerknecht N, Wessel M, Bott M, Wendisch VF (2012) Lrp of Corynebacterium glutamicum controls expression of the brnFE operon encoding the export system for L-methionine and branched-chain amino acids. J Biotechnol 158:231–241
Lee HS, Hwang BJ (2003) Methionine biosynthesis and its regulation in Corynebacterium glutamicum: parallel pathways of transsulfuration and direct sulfhydrylation. Appl Microbiol Biotechnol 62:459–467
Lee JH, Wendisch VF (2017) Production of amino acids - genetic and metabolic engineering approaches. Bioresour Technol 245:1575–1587
Lee KH, Park JH, Kim TY, Kim KU, Lee SY (2007) Systems metabolic engineering of Escherichia coli for L-threonine production. Mol Syst Biol 3:149
Lee SY, Shin HS, Park JS, Kim YH, Min J (2010) Proline reduces the binding of transcriptional regulator ArgR to upstream of argB in Corynebacterium glutamicum. Appl Microbiol Biotechnol 86:235–242
Lee CS, Nam JY, Son ES, Kwon OC, Han W, Cho JY, Park YJ (2012) Next-generation sequencing-based genome-wide mutation analysis of L-lysine-producing Corynebacterium glutamicum ATCC 21300 strain. J Microbiol 50:860–863
Li L, Wada M, Yokota A (2007) A comparative proteomic approach to understand the adaptations of an H+-ATPase-defective mutant of Corynebacterium glutamicum ATCC14067 to energy deficiencies. Proteomics 7:3348–3357
Li Y, Cong H, Liu B, Song J, Sun X, Zhang J, Yang Q (2016) Metabolic engineering of Corynebacterium glutamicum for methionine production by removing feedback inhibition and increasing NAPDH level. Antonie Van Leeuwenhoek 109:1185–1197
Lindner SN, Seibold GM, Henrich A, Krämer R, Wendisch VF (2011a) Phosphotransferase system-independent glucose utilization in Corynebacterium glutamicum by inositol permeases and glucokinases. Appl Environ Microbiol 77:3571–3581
Lindner SN, Seibold GM, Krämer R, Wendisch VF (2011b) Impact of a new glucose utilization pathway in amino acid-producing Corynebacterium glutamicum. Bioeng Bugs 2:291–295
Liu Q, Zhang J, Wei XX, Ouyang SP, Wu Q, Chen GQ (2008) Microbial production of L-glutamate and L -glutamine by recombinant Corynebacterium glutamicum harboring Vitreoscilla hemoglobin gene vgb. Appl Microbiol Biotechnol 77:1297–1304
Liu Q, Liang Y, Zhang Y, Shang X, Liu S, Wen J, Wen T (2015) YjeH is a novel exporter of L-methionine and branched-chain amino acids in Escherichia coli. Appl Environ Microbiol 81:7753–7766
Liu C, Zhang B, Liu YM, Yang KQ, Liu SJ (2018a) New intracellular shikimic acid biosensor for monitoring shikimate synthesis in Corynebacterium glutamicum. ACS Synth Biol 7:591–601
Liu H, Fang G, Wu H, Li Z, Ye Q (2018b) L-cysteine production in Escherichia coli based on rational metabolic engineering and modular strategy. Biotechnol J 13:e1700695
Lu SC, Mato JM (2012) S-adenosylmethionine in liver health, injury, and cancer. Physiol Rev 92:1515–1542
Lubitz D, Jorge JM, Pérez-García F, Taniguchi H, Wendisch VF (2016) Roles of export genes cgmA and lysE for the production of L-arginine and L-citrulline by Corynebacterium glutamicum. Appl Microbiol Biotechnol 100:8465–8474
Ma W, Wang J, Li Y, Hu X, Shi F, Wang X (2016) Enhancing pentose phosphate pathway in Corynebacterium glutamicum to improve L-isoleucine production. Biotechnol Appl Biochem 63:877–885
Ma Q, Zhang Q, Xu Q, Zhang C, Li Y, Fan X, Xie X, Chen N (2017) Systems metabolic engineering strategies for the production of amino acids. Synth Syst Biotechnol 2:87–96
Ma Y, Chen Q, Cui Y, Du L, Shi T, Xu Q, Ma Q, Xie X, Chen N (2018) Comparative genomic and genetic functional analysis of industrial L-leucine- and L-valine-producing Corynebacterium glutamicum strains. J Microbiol Biotechnol 28:1916–1927
Mahr R, Gätgens C, Gätgens J, Polen T, Kalinowski J, Frunzke J (2015) Biosensor-driven adaptive laboratory evolution of l-valine production in Corynebacterium glutamicum. Metab Eng 32:184–194
Mampel J, Schröder H, Haefner S, Sauer U (2005) Single-gene knockout of a novel regulatory element confers ethionine resistance and elevates methionine production in Corynebacterium glutamicum. Appl Microbiol Biotechnol 68:228–236
Man Z, Rao Z, Xu M, Guo J, Yang T, Zhang X, Xu Z (2016a) Improvement of the intracellular environment for enhancing l-arginine production of Corynebacterium glutamicum by inactivation of H2O2-forming flavin reductases and optimization of ATP supply. Metab Eng 38:310–321
Man Z, Xu M, Rao Z, Guo J, Yang T, Zhang X, Xu Z (2016b) Systems pathway engineering of Corynebacterium crenatum for improved L-arginine production. Sci Rep 6:28629
Marienhagen J, Eggeling L (2008) Metabolic function of Corynebacterium glutamicum aminotransferases AlaT and AvtA and impact on L-valine production. Appl Environ Microbiol 74:7457–7462
Marx A, Hans S, Mockel B, Bathe B, de Graaf AA (2003) Metabolic phenotype of phosphoglucose isomerase mutants of Corynebacterium glutamicum. J Biotechnol 104:185–197
Meiswinkel TM, Gopinath V, Lindner SN, Nampoothiri KM, Wendisch VF (2013) Accelerated pentose utilization by Corynebacterium glutamicum for accelerated production of lysine, glutamate, ornithine and putrescine. Microb Biotechnol 6:131–140
Merlin C, Gardiner G, Durand S, Masters M (2002) The Escherichia coli metD locus encodes an ABC transporter which includes Abc (MetN), YaeE (MetI), and YaeC (MetQ). J Bacteriol 184:5513–5517
Mitsuhashi S (2014) Current topics in the biotechnological production of essential amino acids, functional amino acids, and dipeptides. Curr Opin Biotechnol 26:38–44
Mitsuhashi S, Hayashi M, Ohnishi J, Ikeda M (2006) Disruption of malate:quinone oxidoreductase increases L-lysine production by Corynebacterium glutamicum. Biosci Biotechnol Biochem 70:2803–2806
Mizuno Y, Nagano-Shoji M, Kubo S, Kawamura Y, Yoshida A, Kawasaki H, Nishiyama M, Yoshida M, Kosono S (2016) Altered acetylation and succinylation profiles in Corynebacterium glutamicum in response to conditions inducing glutamate overproduction. Microbiology 5:152–173
Möckel B, Pfefferle W, Huthmacher K, Rückert C, Kalinowski J, Pühler A, Binder M, Greissinger D, Thierbach G (2002) Nucleotide sequences which code for the metY gene. International Patent Application WO 02/18613
Moon MW, Kim HJ, Oh TK, Shin CS, Lee JS, Kim SJ, Lee JK (2005) Analyses of enzyme II gene mutants for sugar transport and heterologous expression of fructokinase gene in Corynebacterium glutamicum ATCC 13032. FEMS Microbiol Lett 244:259–266
Morbach S, Sahm H, Eggeling L (1995) Use of feedback-resistant threonine dehydratases of Corynebacterium glutamicum to increase carbon flux towards L-isoleucine. Appl Environ Microbiol 61:4315–4320
Morbach S, Sahm H, Eggeling L (1996) L-isoleucine production with Corynebacterium glutamicum: further flux increase and limitation of export. App Environ Microbiol 62:4345–4351
Mori A, Morita M, Morishita K, Sakamoto K, Nakahara T, Ishii K (2015) L-Citrulline dilates rat retinal arterioles via nitric oxide- and prostaglandin-dependent pathways in vivo. J Pharmacol Sci 127:419–423
Mustafi N, Grünberger A, Kohlheyer D, Bott M, Frunzke J (2012) The development and application of a single-cell biosensor for the detection of L-methionine and branched-chain amino acids. Metab Eng 14:449–457
Nagano-Shoji M, Hamamoto Y, Mizuno Y, Yamada A, Kikuchi M, Shirouzu M, Umehara T, Yoshida M, Nishiyama M, Kosono S (2017) Characterization of lysine acetylation of a phosphoenolpyruvate carboxylase involved in glutamate overproduction in Corynebacterium glutamicum. Mol Microbiol 104:677–689
Nakamura J, Hirano S, Ito H, Wachi M (2007) Mutations of the Corynebacterium glutamicum NCgl1221 gene, encoding a mechanosensitive channel homolog, induce L-glutamic acid production. Appl Environ Microbiol 73:4491–4498
Niebisch A, Kabus A, Schultz C, Weil B, Bott M (2006) Corynebacterial protein kinase G controls 2-oxoglutarate dehydrogenase activity via the phosphorylation status of the OdhI protein. J Biol Chem 281:12300–12307
Nishimura T, Vertès AA, Shinoda Y, Inui M, Yukawa H (2007) Anaerobic growth of Corynebacterium glutamicum using nitrate as a terminal electron acceptor. Appl Microbiol Biotechnol 75:889–897
Nonaka G (2018) Bacterial metabolism and fermentative production of cysteine. Bioscience and Industry 76:110–104
Nottebrock D, Meyer U, Krämer R, Morbach S (2003) Molecular and biochemical characterization of mechanosensitive channels in Corynebacterium glutamicum. FEMS Microbiol Lett 218:305–309
Ochiai M, Hayashi T, Morita M, Ina K, Maeda M, Watanabe F, Morishita K (2012) Short-term effects of L-citrulline supplementation on arterial stiffness in middle-aged men. Int J Cardiol 155:257–261
Ohnishi J, Ikeda M (2006) Comparisons of potentials for L-lysine production among different Corynebacterium glutamicum strains. Biosci Biotechnol Biochem 70:1017–1020
Ohnishi J, Mitsuhashi S, Hayashi M, Ando S, Yokoi H, Ochiai K, Ikeda M (2002) A novel methodology employing Corynebacterium glutamicum genome information to generate a new L-lysine-producing mutant. Appl Microbiol Biotechnol 58:217–223
Ohnishi J, Hayashi M, Mitsuhashi S, Ikeda M (2003) Efficient 40°C fermentation of L-lysine by a new Corynebacterium glutamicum mutant developed by genome breeding. Appl Microbiol Biotechnol 62:69–75
Ohnishi J, Katahira R, Mitsuhashi S, Kakita S, Ikeda M (2005) A novel gnd mutation leading to increased L-lysine production in Corynebacterium glutamicum. FEMS Microbiol Lett 242:265–274
Park JH, Lee SY (2010) Fermentative production of branched chain amino acids: a focus on metabolic engineering. Appl Microbiol Biotechnol 85:491–506
Park JH, Lee KH, Kim TY, Lee SY (2007a) Metabolic engineering of Escherichia coli for the production of L-valine based on transcriptome analysis and in silico gene knockout simulation. Proc Natl Acad Sci U S A 104:7797–7802
Park SD, Lee JY, Sim SY, Kim Y, Lee HS (2007b) Characteristics of methionine production by an engineered Corynebacterium glutamicum strain. Metab Eng 9:327–336
Park SH, Kim HU, Kim TY, Park JS, Kim SS, Lee SY (2014) Metabolic engineering of Corynebacterium glutamicum for L-arginine production. Nat Commun 5:4618
Pátek M (2007) Branched-chain amino acids. In: Wendisch VF (ed) Microbiology monographs, amino acid biosynthesis - pathways, regulation and metabolic engineering. Springer, Berlin, pp 129–162
Petersen S, Mack C, de Graaf AA, Riedel C, Eikmanns BJ, Sahm H (2001) Metabolic consequences of altered phosphoenolpyruvate carboxykinase activity in Corynebacterium glutamicum reveal anaplerotic mechanisms in vivo. Metab Eng 3:344–361
Peters-Wendisch PG, Schiel B, Wendisch VF, Katsoulidis E, Möckel B, Sahm H, Eikmanns BJ (2001) Pyruvate carboxylase is a major bottleneck for glutamate and lysine production by Corynebacterium glutamicum. J Mol Microbiol Biotechnol 3:295–300
Petri K, Walter F, Persicke M, Rückert C, Kalinowski J (2013) A novel type of N-acetylglutamate synthase is involved in the first step of arginine biosynthesis in Corynebacterium glutamicum. BMC Genomics 14:713
Qin T, Hu X, Hu J, Wang X (2015) Metabolic engineering of Corynebacterium glutamicum strain ATCC 13032 to produce L-methionine. Biotechnol Appl Biochem 62:563–573
Radmacher E, Eggeling L (2007) The three tricarboxylate synthase activities of Corynebacterium glutamicum and increase of L-lysine synthesis. Appl Microbiol Biotechnol 76:587–589
Radmacher E, Vaitsikova A, Burger U, Krumbach K, Sahm H, Eggeling L (2002) Linking central metabolism with increased pathway flux: L-valine accumulation by Corynebacterium glutamicum. Appl Environ Microbiol 68:2246–2250
Research and Markets (2018) Amino acids market: global industry trends, share, size, growth, opportunity and forecast 2018–2023. https://www.researchandmarkets.com/reports/4514538/amino-acids-market-global-industry-trends
Rey DA, Pühler A, Kalinowski J (2003) The putative transcriptional regulator McbR, member of the TetR-family, is involved in the regulation of the metabolic network directing the synthesis of sulfur containing amino acids in Corynebacterium glutamicum. J Biotechnol 103:51–65
Rey DA, Nentwich SS, Koch DJ, Rückert C, Pühler A, Tauch A, Kalinowski J (2005) The McbR repressor modulated by the effector substance S-adenosylmethionine controls directly the transcription of a regulon involved in Sulphur metabolism of Corynebacterium glutamicum ATCC 13032. Mol Microbiol 56:871–887
Riedel C, Rittmann D, Dangel P, Möckel B, Sahm H, Eikmanns BJ (2001) Characterization, expression, and inactivation of the phosphoenolpyruvate carboxykinase gene from Corynebacterium glutamicum and significance of the enzyme for growth and amino acid production. J Mol Microbiol Biotechnol 3:573–583
Rieping M, Hermann T (2007) L-threonine. In: Wendisch VF (ed) Microbiology monographs, amino acid biosynthesis - pathways, regulation and metabolic engineering. Springer, Berlin, pp 71–92
Rittmann D, Lindner SN, Wendisch VF (2008) Engineering of a glycerol utilization pathway for amino acid production by Corynebacterium glutamicum. Appl Environ Microbiol 74:6216–6222
Rückert C, Milse J, Albersmeier A, Koch DJ, Pühler A, Kalinowski J (2008) The dual transcriptional regulator CysR in Corynebacterium glutamicum ATCC 13032 controls a subset of genes of the McbR regulon in response to the availability of sulphide acceptor molecules. BMC Genomics 9:483
Sanchez S, Rodríguez-Sanoja R, Ramos A, Demain AL (2018) Our microbes not only produce antibiotics, they also overproduce amino acids. J Antibiot 71:26–36. 2018
Sano K, Shiio I (1971) Microbial production of L-lysine. IV. Selection of lysine-producing mutants from Brevibacterium flavum by detecting threonine sensitivity or halo-forming method. J Gen Appl Microbiol 17:97–113
Sano K, Yokozeki K, Tamura F, Yasuda N, Noda I, Mitsugi K (1977) Microbial conversion of DL-2-amino-Δ2thiazoline-4-carboxylic acid to L-cysteine and L-cystine: screening of microorganisms and identification of products. Appl Environ Microbiol 34:806–810
Sato H, Orishimo K, Shirai T, Hirasawa T, Nagahisa K, Shimizu H, Wachi M (2008) Distinct roles of two anaplerotic pathways in glutamate production induced by biotin limitation in Corynebacterium glutamicum. J Biosci Bioeng 106:51–58
Sawada K, Zen-In S, Wada M, Yokota A (2010) Metabolic changes in a pyruvate kinase gene deletion mutant of Corynebacterium glutamicum ATCC 13032. Metab Eng 12:401–407
Schendzielorz G, Dippong M, Grünberger A, Kohlheyer D, Yoshida A, Binder S, Nishiyama C, Nishiyama M, Bott M, Eggeling L (2014) Taking control over control: use of product sensing in single cells to remove flux control at key enzymes in biosynthesis pathways. ACS Synth Biol 3:21–29
Schulte J, Baumgart M, Bott M (2017) Development of a single-cell GlxR-based cAMP biosensor for Corynebacterium glutamicum. J Biotechnol 258:33–40
Schultz C, Niebisch A, Gebel L, Bott M (2007) Glutamate production by Corynebacterium glutamicum: dependence on the oxoglutarate dehydrogenase inhibitor protein OdhI and protein kinase PknG. Appl Microbiol Biotechnol 76:691–700
Schwentner A, Feith A, Münch E, Busche T, Rückert C, Kalinowski J, Takors R, Blombach B (2018) Metabolic engineering to guide evolution - creating a novel mode for L-valine production with Corynebacterium glutamicum. Metab Eng 47:31–41
Shi F, Li K, Huan X, Wang X (2013) Expression of NAD(H) kinase and glucose-6-phosphate dehydrogenase improve NADPH supply and L-isoleucine biosynthesis in Corynebacterium glutamicum ssp. lactofermentum. Appl Biochem Biotechnol 171:504–521
Shiio I, Miyajima R (1969) Concerted inhibition and its reversal by end products of aspartate kinase in Brevibacterium flavum. J Biochem 65:849–859
Shiio I, Toride Y, Sugimoto S (1984) Production of lysine by pyruvate dehydrogenase mutants of Brevibacterium flavum. Agric Biol Chem 48:3091–3098
Shimomura Y, Yamamoto Y, Bajotto G, Sato J, Murakami T, Shimomura N, Kobayashi H, Mawatari K (2006) Nutraceutical effects of branched-chain amino acids on skeletal muscle. J Nutr 136:529S–532S
Shingu H, Terui G (1971) Studies on process of glutamic acid fermentation at the enzyme level. Part I. on the change of α-ketoglutaric acid dehydrogenase in the course of culture. J Ferment Technol 49:400–405
Sirko A, Hryniewicz M, Hulanicka D, Böck A (1990) Sulfate and thiosulfate transport in Escherichia coli K-12: nucleotide sequence and expression of the cysTWAM gene cluster. J Bacteriol 172:3351–3357
Sprenger GA (2007) Aromatic amino acids. In: Wendisch VF (ed) Microbiology monographs, Amino acid biosynthesis - pathways, regulation and metabolic engineering. Springer, Berlin, pp 93–127
Strelkov S, von Elstermann M, Schomburg D (2004) Comprehensive analysis of metabolites in Corynebacterium glutamicum by gas chromatography/mass spectrometry. Biol Chem 385:853–861
Takeno S, Ohnishi J, Komatsu T, Masaki T, Sen K, Ikeda M (2007) Anaerobic growth and potential for amino acid production by nitrate respiration in Corynebacterium glutamicum. Appl Microbiol Biotechnol 75:1173–1182
Takeno S, Murata R, Kobayashi R, Mitsuhashi S, Ikeda M (2010) Engineering of Corynebacterium glutamicum with an NADPH-generating glycolytic pathway for L-lysine production. Appl Environ Microbiol 76:7154–7160
Takeno S, Hori K, Ohtani S, Mimura A, Mitsuhashi S, Ikeda M (2016) L-lysine production independent of the oxidative pentose phosphate pathway by Corynebacterium glutamicum with the Streptococcus mutans gapN gene. Metab Eng 37:1–10
Takumi K, Nonaka G (2014) An L-amino acid-producing bacterium and a method for producing an L-amino acid. European Patent EP 2218729 B1
Takumi K, Nonaka G (2016) Bacterial cysteine-inducible cysteine resistance systems. J Bacteriol 198:1384–1392
Takumi K, Ziyatdinov MK, Samsonov V, Nonaka G (2017) Fermentative production of cysteine by Pantoea ananatis. Appl Environ Microbiol 83:e02502–e02516
Trötschel C, Deutenberg D, Bathe B, Burkovski A, Krämer R (2005) Characterization of methionine export in Corynebacterium glutamicum. J Bacteriol 187:3786–3794
Trötschel C, Follmann M, Nettekoven JA, Mohrbach T, Forrest LR, Burkovski A, Marin K, Krämer R (2008) Methionine uptake in Corynebacterium glutamicum by MetQNI and by MetPS, a novel methionine and alanine importer of the NSS neurotransmitter transporter family. Biochemistry 47:12698–12709
Umbarger HE (1978) Amino acid biosynthesis and its regulation. Annu Rev Biochem 47:533–606
Usuda Y, Hara Y, Kojima H (2017) Toward sustainable amino acid production. In: Yokota A, Ikeda M (eds) Advances in biochemical engineering/biotechnology, Amino acid fermentation, vol 159. Springer, Tokyo, pp 289–304
Utagawa T (2004) Arginine metabolism: enzymology, nutrition, and clinical significance. J Nutr 134:2854S–2857S
van Ooyen J, Noack S, Bott M, Reth A, Eggeling L (2012) Improved L-lysine production with Corynebacterium glutamicum and systemic insight into citrate synthase flux and activity. Biotechnol Bioeng 109:2070–2081
Vassilev I, Gießelmann G, Schwechheimer SK, Wittmann C, Virdis B, Krömer JO (2018) Anodic electro-fermentation: anaerobic production of L-lysine by recombinant Corynebacterium glutamicum. Biotechnol Bioeng 115:1499–1508
Vogt M, Haas S, Klaffl S, Polen T, Eggeling L, van Ooyen J, Bott M (2014) Pushing product formation to its limit: metabolic engineering of Corynebacterium glutamicum for L-leucine overproduction. Metab Eng 22:40–52
Vogt M, Krumbach K, Bang WG, van Ooyen J, Noack S, Klein B, Bott M, Eggeling L (2015) The contest for precursors: channelling L-isoleucine synthesis in Corynebacterium glutamicum without byproduct formation. Appl Microbiol Biotechnol 99:791–800
Vrljić M, Sahm H, Eggeling L (1996) A new type of transporter with a new type of cellular function: L-lysine export from Corynebacterium glutamicum. Mol Microbiol 22:815–826
Wada M, Takagi H (2006) Metabolic pathways and biotechnological production of L-cysteine. Appl Microbiol Biotechnol 73:48–54
Wada M, Awano N, Haisa K, Takagi H, Nakamori S (2002) Purification, characterization and identification of cysteine desulfhydrase of Corynebacterium glutamicum, and its relationship to cysteine production. FEMS Microbiol Lett 217:103–107
Wada M, Hijikata N, Aoki R, Takesue N, Yokota A (2008) Enhanced valine production in Corynebacterium glutamicum with defective H+-ATPase and C-terminal truncated acetohydroxyacid synthase. Biosci Biotechnol Biochem 72:2959–2965
Wang J, Cheng LK, Wang J, Liu Q, Shen T, Chen N (2013) Genetic engineering of Escherichia coli to enhance production of L-tryptophan. Appl Microbiol Biotechnol 97:7587–7596
Wang X, Zhang H, Quinn PJ (2018a) Production of L-valine from metabolically engineered Corynebacterium glutamicum. Appl Microbiol Biotechnol 102:4319–4330
Wang Y, Cao G, Xu D, Fan L, Wu X, Ni X, Zhao S, Zheng P, Sun J, Ma Y (2018b) A novel Corynebacterium glutamicum L-glutamate exporter. Appl Environ Microbiol 84(6):e02691–e02617
Webster DA (1987) Structure and function of bacterial hemoglobin and related proteins. In: Eichhorn GC, Marzilli LG (eds) Advances in inorganic chemistry. Elsevier, New York, pp 245–265
Wei L, Wang H, Xu N, Zhou W, Ju J, Liu J, Ma Y (2018) Metabolic engineering of Corynebacterium glutamicum for L-cysteine production. Appl Microbiol Biotechnol 103(3):1325–1338. https://doi.org/10.1007/s00253-018-9547-7
Wen J, Bao J (2019) Engineering Corynebacterium glutamicum triggers glutamic acid accumulation in biotin-rich corn Stover hydrolysate. Biotechnol Biofuels 12:86
Wendisch VF (2007) Microbiology monographs, amino acid biosynthesis - pathways, regulation and metabolic engineering. Springer, Berlin
Wendisch VF, Bott M, Kalinowski J, Oldiges M, Wiechert W (2006) Emerging Corynebacterium glutamicum systems biology. J Biotechnol 124:74–92
Willis LB, Lessard PA, Sinskey AJ (2005) Synthesis of L-threonine and branched-chain amino acids. In: Eggeling L, Bott M (eds) Handbook of Corynebacterium glutamicum. CRC, Boca Raton, pp 511–531
Wittmann C, Heinzle E (2002) Genealogy profiling through strain improvement by using metabolic network analysis: metabolic flux genealogy of several generations of lysine-producing Corynebacteria. Appl Environ Microbiol 68:5843–5859
Wu Y, Li P, Zheng P, Zhou W, Chen N, Sun J (2015) Complete genome sequence of Corynebacterium glutamicum B253, a Chinese lysine-producing strain. J Biotechnol 207:10–11
Xafenias N, Kmezik C, Mapelli V (2017) Enhancement of anaerobic lysine production in Corynebacterium glutamicum electrofermentations. Bioelectrochemistry 117:40–47
Xie X, Xu L, Shi J, Xu Q, Chen N (2012) Effect of transport proteins on L-isoleucine production with the L-isoleucine-producing strain Corynebacterium glutamicum YILW. J Ind Microbiol Biotechnol 39:1549–1556
Xu J, Zhang J, Guo Y, Zai Y, Zhang W (2013) Improvement of cell growth and L-lysine production by genetically modified Corynebacterium glutamicum during growth on molasses. J Ind Microbiol Biotechnol 40:1423–1432
Xu J, Han M, Zhang J, Guo Y, Qian H, Zhang W (2014a) Improvement of L-lysine production combines with minimization of by-products synthesis in Corynebacterium glutamicum. J Chem Technol Biotechnol 89:1924–1933
Xu J, Han M, Zhang J, Guo Y, Zhang W (2014b) Metabolic engineering Corynebacterium glutamicum for the L-lysine production by increasing the flux into L-lysine biosynthetic pathway. Amino Acids 46:2165–2175
Xu JZ, Zhang JL, Guo YF, Jia QD, Zhang WG (2014c) Heterologous expression of Escherichia coli fructose-1,6-bisphosphatase in Corynebacterium glutamicum and evaluating the effect on cell growth and L-lysine production. Prep Biochem Biotechnol 44:493–509
Xu JZ, Wu ZH, Gao SJ, Zhang W (2018a) Rational modification of tricarboxylic acid cycle for improving L-lysine production in Corynebacterium glutamicum. Microb Cell Factories 17:105
Xu JZ, Yang HK, Liu LM, Wang YY, Zhang WG (2018b) Rational modification of Corynebacterium glutamicum dihydrodipicolinate reductase to switch the nucleotide-cofactor specificity for increasing L-lysine production. Biotechnol Bioeng 115:1764–1777
Xu JZ, Yang HK, Zhang WG (2018c) NADPH metabolism: a survey of its theoretical characteristics and manipulation strategies in amino acid biosynthesis. Crit Rev Biotechnol 38:1061–1076
Xu JZ, Yu HB, Han M, Liu LM, Zhang WG (2019) Metabolic engineering of glucose uptake systems in Corynebacterium glutamicum for improving the efficiency of L-lysine production. J Ind Microbiol Biotechnol 46(7):937–949. https://doi.org/10.1007/s10295-019-02170-w
Yin L, Hu X, Xu D, Ning J, Chen J, Wang X (2012) Co-expression of feedback-resistant threonine dehydratase and acetohydroxy acid synthase increase L-isoleucine production in Corynebacterium glutamicum. Metab Eng 14:542–550
Yin L, Shi F, Hu X, Chen C, Wang X (2013) Increasing L-isoleucine production in Corynebacterium glutamicum by overexpressing global regulator Lrp and two-component export system BrnFE. J Appl Microbiol 114:1369–1377
Yokota A, Ikeda M (2017) Amino acid fermentation. In: Advances in biochemical engineering/biotechnology, vol 159. Springer, Berlin
Yukawa H, Omumasaba CA, Nonaka H, Kós P, Okai N, Suzuki N, Suda M, Tsuge Y, Watanabe J, Ikeda Y, Vertès AA, Inui M (2007) Comparative analysis of the Corynebacterium glutamicum group and complete genome sequence of strain R. Microbiology 153:1042–1058
Zhang L, Li Y, Wang Z, Xia Y, Chen W, Tang K (2007) Recent developments and future prospects of Vitreoscilla hemoglobin application in metabolic engineering. Biotechnol Adv 25:123–136
Zhang B, Yu M, Zhou Y, Li Y, Ye BC (2017a) Systematic pathway engineering of Corynebacterium glutamicum S9114 for L-ornithine production. Microb Cell Factories 16:158
Zhang Y, Cai J, Shang X, Wang B, Liu S, Chai X, Tan T, Zhang Y, Wen T (2017b) A new genome-scale metabolic model of Corynebacterium glutamicum and its application. Biotechnol Biofuels 10:169
Zhang B, Ren LQ, Yu M, Zhou Y, Ye BC (2018a) Enhanced L-ornithine production by systematic manipulation of L-ornithine metabolism in engineered Corynebacterium glutamicum S9114. Bioresour Technol 250:60–68
Zhang B, Yu M, Wei WP, Ye BC (2018b) Optimization of l-ornithine production in recombinant Corynebacterium glutamicum S9114 by cg3035 overexpression and manipulating the central metabolic pathway. Microb Cell Factories 17:91
Zhang B, Yu M, Zhou Y, Ye BC (2018c) Improvement of L-ornithine production by attenuation of argF in engineered Corynebacterium glutamicum S9114. AMB Express 8:26
Zhang H, Li Y, Wang C, Wang X (2018d) Understanding the high L-valine production in Corynebacterium glutamicum VWB-1 using transcriptomics and proteomics. Sci Rep 8:3632
Zhang X, Zhang X, Xu G, Zhang X, Shi J, Xu Z (2018e) Integration of ARTP mutagenesis with biosensor-mediated high-throughput screening to improve L-serine yield in Corynebacterium glutamicum. Appl Microbiol Biotechnol 102:5939–5951
Zhao J, Hu X, Li Y, Wang X (2015) Overexpression of ribosome elongation factor G and recycling factor increases L-isoleucine production in Corynebacterium glutamicum. Appl Microbiol Biotechnol 99:4795–4805
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2020 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Ikeda, M., Takeno, S. (2020). Recent Advances in Amino Acid Production. In: Inui, M., Toyoda, K. (eds) Corynebacterium glutamicum. Microbiology Monographs, vol 23. Springer, Cham. https://doi.org/10.1007/978-3-030-39267-3_7
Download citation
DOI: https://doi.org/10.1007/978-3-030-39267-3_7
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-39266-6
Online ISBN: 978-3-030-39267-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)