
Abstract
The howler monkey clade includes species of Alouatta and four extinct genera, Stirtonia, Paralouatta, Protopithecus, and probably Solimoea as well. Contrary to expectations, this radiation may have originated as a largely frugivorous group; advanced, Alouatta-like leaf-eating is a novelty well-developed in the Alouatta-Stirtonia sublineage only. Revised body mass estimates place Stirtonia and Paralouatta within the size range exhibited by the living forms and confirm the place of Protopithecus in a larger, baboon-like size range. While their dentitions are more primitive than the Alouatta-Stirtonia pattern, the cranial anatomy of Protopithecus and Paralouatta is distinctly similar to living howler monkeys in highly derived features relating to enlargement of the subbasal space in the neck and in head carriage, suggesting that ancestral alouattines may have had an enlarged hyolaryngeal apparatus. All alouattines also have relatively small brains, including Protopithecus, a genus that was probably quite frugivorous. The successful origins of the alouattine clade may owe more to key adaptations involving communication and energetics than dental or locomotor breakthroughs. While the fossil record confirms aspects of previous character-analysis reconstructions based on the living forms, alouattines experienced a complexity of adaptive shifts whose history cannot be recoverable without a more complete fossil record.*
Resumen
El clado de los monos aulladores incluye las especies de Alouatta y cuatro géneros extintos, Stirtonia, Paralouatta, Protopithecus y probablemente Solimoea. Contrario a las expectativas, esta radiación pudo haberse originado a partir de hábitos frugívoros. La avanzada folivoría de Alouatta es una novedad desarrollada solamente en el sublinaje de Alouatta-Stirtonia. Las estimaciones de masa corporal ubican a Stirtonia y Paralouatta dentro del rango que exhiben las formas vivientes y confirman la posición de Protopithecus en un rango de tamaño mayor, similar al de los babuinos africanos. Considerando que la dentición es más primitiva que el patrón observado en Alouatta-Stirtonia, la anatomía craneana de Protopithecus y Paralouatta es similar a la de los aulladores vivientes debido a los rasgos altamente especializados relacionados al agrandamiento del espacio sub-basal en el cuello, así como en la posición de la cabeza, sugiriendo que los alouatinos ancestrales pudieron haber tenido un gran aparato hiolaríngeo. Todos los alouatinos también presentan un cerebro pequeño, incluyendo Protopithecus, género que probablemente haya sido frugívoro. El origen exitoso del clado de los alouatinos pudo deberse más a adaptaciones de comunicación y energéticas que a cambios dentarios o locomotores. Mientras que el registro fósil confirma ciertos aspectos de análisis de caracteres previos basados en formas vivientes, los alouatinos experimentaron una complejidad de adaptaciones cuya historia no podría reconstruirse sin el registro fósil.*
* Since this chapter was written, additional study by Halenar and Rosenberger (2013) of the material discussed here as Protopithecus led to the conclusion that the two samples actually represent two different genera. The essentially complete Bahian skeleton, which forms the basis of the present discussion, is being assigned to a new genus and species, Cartelles coimbrafilhoi, within subfamily Alouattinae. The original Lund material from Minas Gerais bears the original name Protopithecus, but its affinities are more likely to be found among atelines than alouattines.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
Fossils discovered in recent years have added important information to our knowledge of the diversity and evolution of platyrrhines closely related to one of the most anatomically divergent members of the radiation, the living howler monkeys, Alouatta. While the record is still scant, these additions mean the alouattine-plus-ateline clade, i.e., the fully prehensile-tailed New World monkeys (NWMs), is becoming one of the better-known lineages among the platyrrhines. Only pitheciines are better represented taxonomically among Tertiary and Quaternary remains (Rosenberger 2002).
The first historical narratives of the evolution of howler monkeys are of recent vintage, and they relied extensively on character analysis of the morphology and behavioral ecology of living atelids rather than paleontology (e.g., Rosenberger and Strier 1989; Strier 1992). Out of necessity, these studies focused on the contrasts between the living members of the two sister clades, alouattines (Alouatta) and atelines (Lagothrix, Ateles, Brachyteles). Few relevant, informative fossils were known prior to the 1980s. The one exception was Stirtonia tatacoensis from the middle Miocene La Venta beds of Colombia, 13.5–11.8 MA (Flynn et al. 1997). It was first found as dental remains in the late 1940s (Stirton 1951) and the species has been widely recognized as being both similar and related closely to Alouatta (e.g., Szalay and Delson 1979; Setoguchi et al. 1981; Delson and Rosenberger 1984; Rosenberger 1992; Hartwig and Meldrum 2002; but see Hershkovitz 1970). In the late 1980s, a second species, S. victoriae, was discovered at La Venta (Kay et al. 1987), and an isolated Stirtonia molar from the younger, late middle Miocene Solimões Formation in western Brazil, about 8 MA, also came to light recently (Kay and Frailey 1993). Stirtonia reinforced the notion that leaf-eating was an enduring and essential aspect of the howler monkey’s ecophylogenetic biology. The type specimen of another species related to Alouatta, Protopithecus brasiliensis from the Quaternary of Brazil, had been known since 1838 (Lund 1838), but the fossil was based on a partial humerus and femur and could not be properly interpreted for another 150 years (Hartwig and Cartelle 1996; see footnote above).
Finds in Brazil and Cuba add another dimension of complexity to the Stirtonia-Alouatta story, introducing an unexpected anatomical diversity. This panorama of diversity highlights the unusual nature of living Alouatta as a genus and suggests a need to reevaluate the Rosenberger and Strier (1989)/Strier (1992) model of alouattine evolution. Besides Protopithecus brasiliensis, the Alouatta clade also includes Paralouatta varonai from the Quaternary of Cuba (Rivero and Arredondo 1991) and perhaps Paralouatta marianae from the Miocene of Cuba (see MacPhee et al. 2003). If the latter species, known only from a single astragalus, is indeed an alouattine, these congeners represent a lengthy span of geological time. More problematic is Solimoea acrensis, described from a small set of isolated dental elements, two specimens including three teeth, discovered in Brazil’s Solimões Formation (Kay and Cozzuol 2006). The best evidence of its affinities consists of a single lower molar, which has distinctive crown morphology. The species was originally interpreted as a stem ateline, but we present reasons why it is probably an alouattine. Finally, also from a late Pleistocene cave of Bahia, Brazil, is a little known extinct species of howler monkey, Alouatta mauroi (Tejedor et al. 2008), which we mention only for the sake of completeness.
Our purpose here is to establish the taxonomic composition and interrelationships of living and extinct alouattines, present new information pertaining to their craniodental diversity, and explore several aspects of alouattine evolutionary history as an adaptive array. The phylogenetics and differentiation of this group has not been discussed previously. Part of the reason for this is that the composition of the subfamily Alouattinae is a matter of debate. In addition to the question of Solimoea, raised here for the first time, there are different views about the affinities of Paralouatta (e.g., Rivero and Arredondo 1991; MacPhee and Horovitz 2002; Rosenberger 2002), which MacPhee and colleagues (MacPhee et al. 1995; Horovitz and MacPhee 1999) maintain is monophyletically related to the other extinct Caribbean primates and, among the extant forms, to mainland Callicebus, a pitheciid. The present study emphasizes why, from a functional-morphological perspective, an affinity with alouattines is the more parsimonious hypothesis, as Rivero and Arredondo (1991) originally proposed.
2 Methods
Craniodental measurements of the modern samples used in this study are largely from collections in the American Museum of Natural History, the United States National Museum, the Field Museum of Natural History, the Natural History Museum (London), the Museu Nacional de Rio de Janeiro, and the Zoologisk Museum, Statens Naturhistoriske Museum (Copenhagen). Species identifications and sample sizes are given where appropriate. Standard linear craniodental measurements were taken to the 0.10 mm with digital calipers. Some teeth were measured using high-resolution laser scans of epoxy casts, using Landmark Editor (Wiley et al. 2005). Endocranial volumes were taken by pouring small plastic beads or other filler into the cavity then transferring the mass to a graduated glass beaker, except in the case of Paralouatta varonai. It was CT scanned in Havana, Cuba, using a medical scanner and a slice thickness of 0.8 mm. Using ImageJ (http://rsb.info.nih.gov/ij/), the endocranial cavity was then outlined as individual slices, composited, and measured. Some measurement error was unavoidable due to difficulty in separating bone from the matrix-filled cavity, but our figures here are consistent with other measurements used in the context of our assessment of relative brain size (see below).
Genealogical interrelationships were inferred using conventional, non-algorithmic procedures of character analysis and cladistic reconstruction. Our methodology is based on the functional-adaptational approach (see Szalay and Bock 1991). Reviews of the methodology as applied to atelids can be found in Rosenberger and Strier (1989), Rosenberger et al. (1990), and Rosenberger (1992), where additional references to the literature on cladistic phylogeny reconstruction can be found. Our intent has been to produce a character analysis that elucidates the homologies and polarities of functionally relevant anatomical features. We thus use functional-adaptive inference as well as taxonomic distributional information. The latter relies on commonality and out-group comparisons in order to develop hypotheses about the directionality of change in traits, but functional-adaptive information is necessary to hypothesize why such changes may have taken place. Although we do not specifically present distributional information on non-atelids, we draw on the morphology of the other platyrrhines, living and extinct, as a collective out-group in working out polarities.
We focus on large-scale morphological features that are demonstrably important in distinguishing Alouatta from other atelids at the genus level and are also relevant functionally to the evolution of howler monkey craniodental adaptations, since we are interested in establishing how unit characters evolved within functional complexes as a part of the phylogenetic history of alouattines. Our rationale presumes that the Alouatta cranium and dentition, which is radically different from most primates in many ways, is composed of an assortment of features that are derived relative to other atelids and platyrrhines. We hypothesize that the evolution of many craniodental features has been driven specifically by a novel adaptive complex relating to howling and folivory.
Solimoea, which we limit to a single molar tooth as discussed below, is referenced only sparingly in the character analysis, which emphasizes cranial anatomy. The basis for our interpretation of the fossil’s affinities is presented in the body of the text following the same functional-adaptive lines employed to assess the cranium.
One feature we address but do not examine through a structured character analysis is body mass. While it has always been evident that body size would figure prominently in narrative explanations of platyrrhine evolution (see Hershkovitz 1972; Rosenberger 1980), its importance for atelids has become stunningly reaffirmed by discovering the large subfossils Protopithecus and Caipora. The initial body weight estimates for these genera (Cartelle and Hartwig 1996; Hartwig and Cartelle 1996) placed both well outside the range of modern forms. However, they were made using regression equations based on a catarrhine reference sample, a phylogenetically less desirable methodology (Hartwig 1995). Statistically robust equations based on platyrrhine postcranial elements which have been shown to be closely linked with body size (e.g., Ruff 2003) have recently been published and confirm the original estimates (Halenar 2011a, b). They have also been used to confirm an estimate of approximately 7–9.5 kg for Paralouatta (Cooke and Halenar 2012). We have taken a less formalistic approach in order to factor in this new information on size and integrate it with the broader analysis. The taxonomic terminology we use divides the monophyletic family Atelidae into alouattines (subfamily Alouattinae: extant Alouatta; extinct Stirtonia, Paralouatta, Protopithecus, and Solimoea) and atelines (subfamily Atelinae: extant Ateles, Brachyteles, and Lagothrix; extinct Caipora).
3 Results
3.1 Craniodental Morphology and Paleontological Synopsis
Two of the three fossil alouattine genera are represented by very good crania (Table 2.1). The third, Stirtonia, is known by excellent dental remains (e.g., Hershkovitz 1970; Szalay and Delson 1979; Setoguchi et al. 1981; Kay et al. 1987; Fleagle et al. 1997; Fleagle 1999; Hartwig and Meldrum 2002). The latter preserves both upper and lower cheek teeth that are unmistakably similar to Alouatta (see Figs. 2.1 and 2.2), so much so that Delson and Rosenberger (1984) suggested that generic separation obscures the possibility that Stirtonia and Alouatta may share an ancestor–descendant relationship and that classifying them as congeners ought to be considered. However, more work needs to be done to more accurately determine the relationships between Stirtonia and Alouatta.

Laser scan generated occlusal views of atelid left maxillary molars [digitized at 25 μm point intervals (here and below) from epoxy casts]. Teeth at left are first molars, in most cases brought to about the same mesiodistal lengths. (1) Ateles geoffroyi, (2) Caipora bambuiorum, (3) Lagothrix lagotricha, (4) Brachyteles arachnoides, (5) Alouatta seniculus, (6) Stirtonia tatacoensis, (7) Paralouatta varonai, (8) Protopithecus brasiliensis

Laser scan generated occlusal views of atelid left mandibular molars (protocols as above). (1) Ateles geoffroyi, (2) Caipora bambuiorum, (3) Lagothrix lagotricha, (4) Brachyteles arachnoides, (5) Alouatta seniculus, (6) Stirtonia tatacoensis, (7) Paralouatta varonai (m1, m3)
Like Alouatta, the upper molars of Stirtonia (Fig. 2.1) are relatively square, with an elevated, lobe-like hypocone; high-relief buccal cusps carrying a long ectoloph; deeply notched centrocrista; and a well-developed stylar shelf area. Lower molars have a small, elevated trigonid with protoconid and metaconid set at an oblique angle and a long talonid with a sharply angled, elongate cristid obliqua. This pattern of features, including elements that have been assessed quantitatively in Alouatta (e.g., Kay 1975; Rosenberger and Kinzey 1976; Kay and Hylander 1978; Kay et al. 1987), is universally interpreted as shearing, leaf-eating characteristics. The upper and lower premolars of Stirtonia are also consistent with an Alouatta-like morphology, as are the tooth proportions. Incisors are not known for Stirtonia, but the inter-canine span in the type mandible appears to be relatively narrow; Alouatta incisors are relatively small (see below). There is no information on the posterior part of the mandible of Stirtonia, the extreme expansion of which is diagnostic of Alouatta.
Protopithecus brasiliensis is now known from a nearly complete skeleton with a very well-preserved skull (Figs. 2.3, 2.4, and 2.5) that includes the anterior teeth, premolars, and a partial upper molar, as well as a mandible with anterior teeth and premolars. It presents an interesting mosaic of craniodental and postcranial traits not found in any other NWM (Hartwig and Cartelle 1996). It shares several cranial features exhibited only in Alouatta among the living platyrrhines, including a relatively extended basicranium and a compound temporo-nuchal crest, which led Hartwig and Cartelle to recognize its alouattine affinities. The teeth of Protopithecus are still incompletely analyzed. They are nonetheless highly informative for the present purpose (see below).

Crania of extant and extinct members of the alouattine and ateline radiations (lateral view). Left to right, top row: Brachyteles, Lagothrix, Alouatta. Bottom row: Caipora, Protopithecus, Paralouatta. Scale bars represent 1 cm. Note the similarities linking Alouatta, Protopithecus, and Paralouatta to the exclusion of the other three genera, especially size and shape of the neurocranium and the airorynchous facial skeleton. The latter trait is indicated by the more acute angle superimposed upon those three skulls between the nasal bridge and the tip of the incisors

Basal view of (left to right) Lagothrix, Alouatta, Protopithecus, and Paralouatta. Scale bars represent 1 cm. Note the anterior-posterior elongation of the alouattine cranial base, as well as the more marked postorbital constriction. The orientation of the foramen magnum and nuchal region of the fossils is intermediate between the ateline condition of Lagothrix and the alouattine condition of extant howler monkeys

Posterior view of (left to right) Caipora, Protopithecus, and Paralouatta, brought approximately to same cranial width. Contrast the relatively small, low braincase; cylindrical brain shape; and prominence of both the temporal (red arrow) and nuchal (blue arrow) crests of Protopithecus and Paralouatta with the rounded, globular braincase; lack of marked temporal lines; and a much less rugose nuchal plane of Caipora
Paralouatta has been classified as two species, P. varonai and P. marianae (Rivero and Arredondo 1991; MacPhee et al. 2003). The latter is known only by an astragalus. The former is represented by a fairly well-preserved but broken skull with worn teeth, a mandible, various isolated teeth (Figs. 2.1, 2.2, 2.3, 2.4, and 2.5), and postcranial material (Rivero and Arredondo 1991; Horovitz and MacPhee 1999; MacPhee and Meldrum 2006). The phylogenetic connection to Alouatta that we advocate is a matter of controversy. The first specimen, the skull, was found prior to the recovery of the new Brazilian Protopithecus material which, as we explain below, supports the case for the alouattine affinities of Paralouatta. When initially described, its overall morphology convinced Rivero and Arredondo (1991) that Paralouatta is closely related to its namesake Alouatta. However, MacPhee and colleagues argued that Paralouatta belongs to a newly recognized clade of Greater Antillean primates (MacPhee et al. 1995; Horovitz and MacPhee 1999; MacPhee and Horovitz 2004) most closely related as a group to Callicebus. This was based on the finding by Horovitz and MacPhee (1999) of three alleged unambiguous, observable craniodental characters that support the clade including Antillothrix bernensis, Xenothrix mcgregori, and Paralouatta varonai: nasal fossa wider than palate at level of M1, lower canine alveolus buccolingually smaller than p4, and m1 protoconid with bulging buccal surface. While this is an intriguing result given the isolation of these taxa from the mainland, it is far from definitive. Thus, Rosenberger (2002) held that Rivero and Arredondo (1991) were correct, as we further elaborate below. An added dimension to the paleobiology of Paralouatta was recently introduced by study of the postcranium. It led MacPhee and Meldrum (2006) to suggest Paralouatta may have been semiterrestrial.
The fourth fossil species we present as alouattine is Solimoea acrensis (Kay and Cozzuol 2006). The type specimen is an isolated lower molar with good crown morphology, identified as an m1. The general description given above for Alouatta and Stirtonia lower molars, which as we stated appears to be universally regarded as howler monkey-like and largely unique to NWMs, compares favorably with the pattern of Solimoea. All are relatively long teeth, with a compact, small elevated trigonid, obliquely oriented trigonid wall (postvallid), elongate talonid, and a long and deeply inflected cristid obliqua.
Caipora bambuiorum, from the same cavern that produced Protopithecus (Cartelle and Hartwig 1996), is in our view the only known extinct ateline (but see footnote above). It is included here for its comparative value in assessing the morphocline polarity of traits among the atelids.
3.2 Body Size
Body size deserves special mention here and we consider it separately from the rest of the character analysis for reasons given above. We provide a series of alternative weight estimates for the fossils based on regressions using different taxonomic samples of anthropoids and different independent variables, both dental and cranial (Conroy 1987; Kay et al. 1998; Sears et al. 2008) (Fig. 2.6). We caution, however, that difficulties remain and, as indicated above, the postcranial skeleton may be more suitable for estimating body size in Protopithecus and Caipora (Halenar 2011a, b). Some equations using skulls have relatively low R 2 values so they cannot be considered highly reliable for projections. While the equations for molars have R 2 values of 0.9 or greater, lower molars are missing from Protopithecus. Caipora, which is probably a frugivore, may also have relatively small teeth, which may bias a molar-based weight estimation. Nevertheless, in our analysis Stirtonia and Paralouatta fall within the range of modern howler monkeys in body mass, as does Solimoea. As noted, new body mass estimates for Protopithecus and Caipora were deemed necessary as the original estimates were calculated from regression equations based on a catarrhine reference sample (Hartwig 1995; Cartelle and Hartwig 1996; Hartwig and Cartelle 1996). Alternative regression equations to estimate size were calculated using a sample of primates encompassing a wide range of body sizes and locomotor patterns (for sample composition see Halenar 2011a, b). For this exercise, the centroid sizes of the epiphyses of various long bones were employed as the skeletal estimator and equations were generated based on the entire sample, the platyrrhines only, and the atelids only. Three aspects of “body size” were predicted for the fossil: body weight (kg), total length (TOTL; mm) which includes the length of the tail (TAILL), and trunk length, head, and body (TrL; mm) which includes the length of the skull and trunk (TOTL = TAILL + TrL; Ford and Corruccini 1985). A relatively wide range of body size estimates was thus recovered for Protopithecus: 12–35 kg, 1,479–1,887 mm TOTL, and 613–831 mm TrL. This range reflects the use of different skeletal elements, reference samples, and regression models. The equation with the combined highest R 2 (=0.98), lowest %SEE (=11.0), MPE (=14.7), and QMLE (=1.005) is that which uses the distal humerus with a platyrrhine-only reference sample; this gives an estimate of 28 kg for the more recently discovered specimen from Toca da Boa Vista and 24 kg for the original specimen discovered by Lund in Lagoa Santa. Condensing all of the estimates into an average, disregarding the obvious extreme outliers in estimate and confidence statistics, gives a body weight of approximately 23 kg, 1,675 mm TOTL, and 710 mm TrL. As an alternative to compiling an average value, a histogram of all body weight predictions shows 19, 21, and 25 kg as the most frequent estimates with a reasonable range from 17 to 29 kg. This puts Protopithecus in the size range of a large male baboon or proboscis monkey and confirms its place in a large-bodied category that no longer exists among extant platyrrhines.

Male and female body weights as reported in the literature for the living atelids (DiFiore and Campbell 2007) and their fossil relatives, the latter based on tooth and/or skull measurements. Weights for Stirtonia and Paralouatta are from Fleagle (1999) and MacPhee and Meldrum (2006), respectively. For other fossil species, including Paralouatta for which additional estimates are included, weights were calculated using the monkey, anthropoid, all primate, and female anthropoid regression equations of Conroy (1987) and the female platyrrhine equation of Kay et al. (1998). Body size estimates based on skull length and bizygomatic width were derived from Sears et al. (2008) equations. The highs and lows are shown instead of averages to demonstrate the wide and overlapping range of body sizes seen in the living atelids, making body mass a difficult character to code and interpret via character analysis. Estimates based on cranial measures are deemed less reliable because of low coefficients of determination (R 2) in the original regressions. Body mass estimates for Protopithecus and Caipora using craniodental measures are substantially below previous reports, but the original estimates of 20–25 kg are confirmed based on postcranial regression equations (Halenar 2011a)
For simplicity, and taking into account the considerations discussed above, within atelids we code the range of character states (Table 2.2) describing body mass at the generic level as medium and large, choosing these terms in part as a semantic device to distinguish them from other platyrrhines often regarded as being middling in size for the radiation, e.g., pitheciids and Cebus (e.g., Hershkovitz 1977). We recognize this grossly underrepresents intrageneric diversity (and likely selection for body mass at the species level) and especially the nature and complex distribution of sexual dimorphism among atelids. But it is a useful, operational approximation considering the foci of this study, fossils and genus-level systematics.
3.3 Character Analysis
Table 2.2 also summarizes the taxonomic distribution of the ten features we assess in detail. As mentioned, the major reasons for selecting these are that they tend to diagnose Alouatta as a genus, defining it morphologically, phylogenetically, and adaptively relative to other living NWM, and they are well represented in the cranial remains of three fossil genera. The fourth, Solimoea, is obviously an exception.
3.3.1 Facial Proportions
Rosenberger (1992) and Rosenberger and Strier (1989) suggested that the Lagothrix-like condition of the facial skeleton, here termed “moderately large”, is ancestral overall for atelids (Figs. 2.3 and 2.4). This was based, in part, on the interpretation that there are two other extremes in the atelid morphocline, exemplified by the Ateles and the Alouatta poles, each one highly likely to be derived since they are associated functionally with novel adaptations. In Ateles, ripe fruit frugivory is linked with reduction of the cheek teeth, well-developed anterior teeth (e.g., Rosenberger 1992; Anthony and Kay 1993), and a small face. This pattern occurs in Caipora as well. In Alouatta, massive changes in the placement and orientation of the large facial skeleton are associated with specializations of the cranial base related to extreme enlargement of the hyoid and the production of stentorian vocalizations (see Biegert 1963). Cheek teeth are also relatively large and anterior teeth are proportionately small (e.g., Rosenberger 1992; Anthony and Kay 1993). Regarding the fossils, we interpret the face of Protopithecus as moderately large, hence similar to the condition seen in Lagothrix, although more work needs to be done on the allometry of this region in the large-bodied fossil. Paralouatta, however, clearly does have a relatively large, long face resembling Alouatta in its proportions. Of the fossils under consideration, it is most comparable to Alouatta with a markedly prognathic snout, but similar prognathism is also evident in Protopithecus.
3.3.2 Craniofacial Haft
A feature correlated with facial size and prognathism is the orientation of the face relative to the braincase. Alouatta, is unusual and highly derived among platyrrhines in having an uptilted rostrum, a condition known as airorhynchy (Figs. 2.3 and 2.4). This design contributes to the expansion of space in the neck for the permanently inflated air sacs inside the hollowed-out hyoid bone and its associated cartilages. Airorhynchy is also linked functionally with elongation of the cranial base (see below). Paralouatta closely resembles Alouatta in this respect, although the dorsal tilt of the face seems to be less exaggerated. Even though the tip of the fossil’s snout is broken near the level of the canines, it is evident that the toothrow is nearly as arched in lateral view, forming an exaggerated curve of Spee. Protopithecus has a modestly uptilted face as well. The rostra of other platyrrhines are constructed differently and are generally non-airorhynchous, as in Caipora. The lateral profile of the Brachyteles dental arcade, with large postcanine teeth and a moderately deep but non-prognathic face, is slightly curved upward anteriorly.
3.3.3 Postorbital Constriction
The degree of postorbital constriction is influenced by braincase size and shape, craniofacial proportions, and the anteroposterior alignment of the face at the craniofacial junction (Fig. 2.3). The modern alouattines and atelines present contrasting character states. The constriction is moderate in atelines, including Caipora, but it is marked (i.e., narrow or waisted) in Alouatta. In atelines such as Ateles and Brachyteles, with retracted, subcerebral (below the horizontal axis of the brain) faces and large, relatively globular braincases, width at the craniofacial junction is not constricted. But even in Lagothrix, where the braincase is not globular, the constriction is unimpressive, as it tends to be in other platyrrhines, suggesting that this state is ancestral in atelids. In Alouatta, in contrast, the combination of a precerebral, uptilted face, massive width of the posterior face, and narrow braincase produces the markedly constricted effect. In ventral view (Fig. 2.4), Paralouatta resembles howler monkeys in these factors. The same is evident in Protopithecus, but it manifests differently because the braincase is quite wide posteriorly, owing largely to well-developed exocranial superstructures.
3.3.4 Cranial Crests
The development of exocranial temporal lines and nuchal crests may be strongly influenced by size, age, gender, and sexual dimorphism, indicating caution in making comparisons without population samples of fossil atelids (Figs. 2.3 and 2.5). Of the fossil specimens considered here, Caipora is a young adult; Protopithecus is an adult but with relatively unworn teeth; Paralouatta is an adult with advanced postcanine tooth wear. Judging by canine prominence, anterior premolar size, and the known level of sexual dimorphism in the living species, Caipora and Protopithecus appear to be males. The canine crowns of Paralouatta are broken away, but the expression of cranial crests suggests the skull may also be male.
Among modern atelids, moderate to prominent temporal lines, evenly developed anteriorly and posteriorly, are present in Lagothrix, Brachyteles, and Alouatta. Strong nuchal lines or crests tend to occur in the robust Alouatta males and are quite common interspecifically. Neither temporal lines nor nuchal crests are well-developed in Ateles, or in Caipora, which corresponds with their reduced cheek teeth and rounded, large braincases, among other factors. We surmise this is a correlate of the soft/ripe-frugivory feeding complex seen in Ateles. It is also related to what may be termed a semi-orthograde head carriage, i.e., the head is not strongly cantilevered off the vertebral column but tends to rest atop the cervical vertebrae in compliance with tail-assisted climbing and other semi-orthograde positional behaviors.
With a small braincase and large temporal and nuchal muscles, a compound temporo-nuchal crest is well-developed in Alouatta, although its distribution among the modern species has not been mapped out. Nevertheless, in the larger and more robust males, laterally away from the midline, the temporal enthuses fuse with the nuchal line to form a raised lateral margin of the nuchal region. By comparison with other atelids, these features are extremely well-developed in Protopithecus, probably as an elaboration of an Alouatta-like pattern exaggerated by the allometrics of a very large body size. The compound temporo-nuchal crest is present also in Paralouatta but exhibited less dramatically, comparing more favorably with the variations seen in Alouatta.
3.3.5 Nuchal Plane
Alouatta is unusual among platyrrhines in having a nuchal plane that is flat, often rugose in texture, reduced in size, vertically oriented (Fig. 2.5), and exhibiting a semicircular dorsal perimeter when viewed from behind—all features corresponding with the cylindrical shape of the braincase and pronounced set of muscle attachments on the occiput. Sex differences exist, but this overall gestalt is fixed in howler monkeys. It relates to head carriage and craniofacial mass. The foramen magnum and occipital condyles are directed posteriorly rather than ventrally as in other NWM, meaning that the large, heavy head of Alouatta, which is eccentrically loaded up front due to its snouty prognathic design, tends to be extended dog- or lemur-like out from the shoulders and neck, in typical pronograde fashion. The flat, vertical nuchal plane presumably gives the trapezius and other neck muscles apt mechanical advantage in supporting the horizontally disposed skull. Following previous arguments, we regard the Lagothrix-like condition, a relatively flat, subvertical, and unreduced nuchal plane as ancestral in atelids. The contrasting rounded and unreduced morphology of Ateles and Caipora is considered derived for atelines. Paralouatta resembles Alouatta generally, but the plane of the nuchal region appears to be more primitive, slanted in a manner that compares with Lagothrix. Similarly, Protopithecus retains an inclined nuchal plane but it is also greatly expanded laterally, owing to the hypertrophic compound temporo-nuchal crests. We hypothesize that this is at least partly an allometric contingency but it may also reflect differences in the proportions of the jaw adductor muscles. The apparent lack of gonial expansion in comparison to Alouatta suggests that Protopithecus had a less elaborate masseter complex, while the enlarged temporo-nuchal crests suggest the posterior part of the temporalis muscle was exaggerated instead.
3.3.6 Foramen Magnum
Both the position (see Schultz 1955) and relative size of the foramen magnum differs among atelids. These features are related to head posture and endocranial volume. As indicated, it is extremely posteriorly positioned in Alouatta, Paralouatta, and Protopithecus, especially so in howler monkeys (Fig. 2.4), and the particulars conform to the degree of nuchal plane modifications in these genera. Alouatta exhibits the most derived pattern. The more anterior location of the foramen magnum in atelines is consistent with the more common location documented by Schultz, which is ancestral for NWMs and atelines. For convenience we code it as posterior, offsetting it from the condition in Saimiri and Cebus. They have foramina magna that are distinctly more “centrally” located within the long axis of the skull.
The foramen magnum also varies in proportions, with atelines and alouattines clearly having different scaling patterns (Fig. 2.7a). Relative to basicranial length, foramen magnum area (length × breadth) is small in Alouatta, Paralouatta, and Protopithecus, falling well below the scatter of points and the regression line representing modern atelines and Caipora. The size of the foramen is also closely correlated with endocranial volume across primates (e.g., Jerison 1973; Martin 1990). Brain size is relatively larger in atelines than alouattines (Fig. 2.7b), which helps explain why the foramen magnum is proportionately smaller in the latter. Again, the alouattine condition is very likely the derived pattern among atelids, given the rarity of de-encephalization, which is often associated in mammals with herbivory or folivory (see section below for an expanded explanation). But it is also possible that to some degree, relatively small brain size in this group reflects primitive platyrrhine proportions. The status of atelines also requires further examination. While Ateles and Brachyteles have been singled out as having derived, elevated relative brain sizes (Cole 1995, in Hartwig 2005), it appears from this assessment that all the atelines, including Lagothrix and Caipora, jointly share this pattern. Even Brachyteles, a genus that might be expected to have experienced selection for a reduced relative brain size as a correlate to its more leafy diet, follows the ateline pattern and is relatively larger-brained than any alouattine (Rosenberger et al. 2011).

Bivariate plots of (a) foramen magnum area and (b) endocranial volume relative to nasion-basion length in atelids. Note the separate distributions of the atelines, including Caipora, toward the top of the graph and the alouattines, including Protopithecus and Paralouatta, toward the bottom. Data points for the living genera are sex-pooled means from the following samples: Brachyteles arachnoides, 3; Ateles belzebuth, 16; Lagothrix lagotricha, 15; Alouatta belzebuth, 16. Alouattines have relatively smaller brains, even the frugivorous Protopithecus, while the leaf-eating ateline Brachyteles does not have a reduced brain size
3.3.7 Brain Size and Shape
As indicated, among modern platyrrhines, it is well established that howler monkeys have an unusually small brain size relative to body mass (e.g., Stephan and Andy 1964; Hershkovitz 1970; Stephan 1972; Clutton-Brock and Harvey 1980; Eisenberg 1981; Martin 1984, 1990; Hartwig 1996), and this likely represents, at least in part, an adaptation to folivory (e.g., Clutton-Brock and Harvey 1980; Eisenberg 1981; Martin 1984, 1990; Harvey and Clutton-Brock 1985; Rosenberger et al. 2011). Since folivory is clearly a derived habit among NWM, the correlative, relatively small Alouatta brain may have evolved via de-encephalization. This does not, however, mean there is no component of primitiveness in this character state, for early platyrrhines probably had smaller brain sizes than modern members (see Tejedor et al. 2006; Sears et al. 2008), parallel increases in relative brain size occurred, and basal lineages of the major clades may logically be expected to retain the primitive platyrrhine condition (see Hartwig et al. 2011).
The conjunction of a relatively small brain in howler monkeys with a posteriorly positioned foramen magnum, small nuchal plane, extended basicranial platform, and precerebral, airorhynchous face makes it likely that the cylindrical shape of the Alouatta braincase is a derived by-product of a spatial packaging phenomenon (i.e., Biegert 1963; Gould 1977; Ross and Ravosa 1993). The Protopithecus skull closely resembles Alouatta in this respect although its braincase differs in shape for it is wider posteriorly than anteriorly, a pattern not seen elsewhere among platyrrhines. However, some of this is an exocranial effect of the very wide nuchal plane, with well-developed lateral nuchal crests and a massive set of temporal roots supporting the zygomatic arches. The finding of Krupp et al. (2012) that the Protopithecus brain resembles Alouatta in overall shape helps explain why Protopithecus cannot share the globular braincase shape of Ateles, Brachyteles, and Caipora, all at the opposite end of the spectrum. Roughly speaking, the Protopithecus braincase may more closely resemble Lagothrix, whose morphology may be described as non-cylindrical for convenience. This would suggest it shares the ancestral condition for atelids.
3.3.8 Basicranial Shape
Alouatta is unusual among platyrrhines and other primates in having an elongate basicranium (Fig. 2.4), presumably as another derived correlate of subbasal spatial packaging, i.e., making room for the enlarged hyoid complex (Biegert 1963). However, it should be noted that within Alouatta, there is considerable interspecific variation in cranial base shape, with A. palliata showing a shorter, more rounded condition (Halenar 2008). A. seniculus males appear to be the most exaggerated, perhaps because the foramen magnum is shifted posteriorly to such an extreme degree. We designate the contrasting character states of Ateles and Caipora as short, but their modified, encephalized skulls suggest this may not be the ancestral atelid or ateline condition. We hypothesize that the deeper morphotype condition is more moderate and designate the primitive condition as “not elongate.” Hartwig and Cartelle (1996) pointed out that the Alouatta-like elongate pattern is evident in Protopithecus, and it is exhibited in Paralouatta as well (Rivero and Arredondo 1991; Halenar 2012). We consider the Protopithecus morphology less derived than in Alouatta and Paralouatta, largely because the nuchal plane continues to extend behind it. In agreement with many of the qualitative statements made above regarding facial proportions and airorynchy, 3D geometric morphometric analysis of the Protopithecus cranial base suggests that it exhibits an intermediate morphology between the extremely derived Alouatta and Ateles conditions; principal components analysis aligns the fossil with extant Lagothrix in terms of its degree of basicranial elongation and flexion (Halenar 2012).
3.3.9 Incisor Proportions
Morphologically, the incisors of atelids appear to show an acute sensitivity to selection reflecting critical dietary preferences (Fig. 2.8). Thus, Alouatta and Brachyteles, the most folivorous platyrrhines, have evolved relatively small-crowned incisors, probably independently (Eaglen 1984; Rosenberger 1992; Anthony and Kay 1993), whereas the other atelids have relatively larger incisors with the lower incisors being distinctly spatulate in shape. Reduced crowns like those of Alouatta and Brachyteles are not prevalent among other platyrrhines, making it likely that the unreduced condition is ancestral for atelids. The much enlarged incisors of Ateles and Lagothrix may be another specialization related to intensive fruit harvesting behaviors. This makes it difficult to specify the morphotype ateline condition. By default, we regard it as being intermediate. Importantly, the proportions of Paralouatta more closely resemble the condition seen in Alouatta and Brachyteles than Protopithecus or any modern atelines. Protopithecus incisors are quite large proportionately, although not to the extent seen in Ateles and Lagothrix. Stirtonia specimens lack incisors, but the well-preserved-type mandible of S. tatacoensis has canines positioned relatively close together, suggesting these teeth were not especially enlarged.

The index of lower incisor size was calculated by dividing the incisal crown cross-sectional area (length × breadth of i1) by first molar area (length × breadth). This exercise was repeated for maxillary and mandibular first molars so as to be able to include both Paralouatta and Protopithecus. Lower values, which indicate relatively small incisors, correspond with a more folivorous diet, as in Alouatta. The position of Protopithecus toward the higher end of the scale, with an index proportionately twice the size of Alouatta, suggests it was considerably frugivorous
3.3.10 Molar Relief and Crown Shape
Taking a very abstract approach in order to characterize the morphology of upper and lower molars simultaneously, we define two crown patterns as character states: “low relief”, with relatively low cusps and shallow, broad basins, and, “cristodont”, having more relief and an emphasis on relatively elevated cusps and lengthy crests, which thus restricts lower molar basins and lengthens the crown (Figs. 2.1 and 2.2). Alouatta is the archetypical example of the cristodont pattern with upper molars also exhibiting a set of strongly developed buccal ectoloph crests (especially the centrocrista between paracone and metacone) as well as a stylar region with a robust buccal cingulum, which is associated with localized crest development. Brachyteles (see Rosenberger 1992) shares several features of the cristodont pattern with Alouatta but appears to have evolved aspects of it by a different, convergent pathway emphasizing lingual, as opposed to buccal, shear. Hence, the massively developed metaconids and entoconids seen in Brachyteles molars (Fig. 2.2).
Among platyrrhines, cristodont molars like those of Alouatta and Brachyteles do not occur outside of the atelids, so it is reasonable to regard this state as derived (in parallel). The low-relief pattern of Ateles and Lagothrix is also part of an unusual, large-basin occlusal morphology among NWMs, functionally related to masticating soft, ripe fruit (Kay 1975; Rosenberger 1992; Anthony and Kay 1993). Hence, we interpret both patterns as derived from a still hypothetical architecture we term “intermediate” for convenience. Among the fossils, Paralouatta upper molars (Fig. 2.1) clearly share with Alouatta well-developed buccal and stylar cristodont features, but the crown is more primitive lingually, retaining the well-differentiated hypocone, for example, that is broadly similar to many living NWM and middle Miocene fossils. The Paralouatta cusps and crests also tend to be more blunted than sharp. The morphology of Protopithecus is poorly known since the specimen lacks lower molars and the single M1 is broken; however, it evidently does not display the cristodont pattern. The occlusal surface of the upper molar appears to be relatively flat and the premolar cusps are bulbous. Both species of Stirtonia have very Alouatta-like, cristodont upper molars. Caipora exhibits an ateline-like, low-relief pattern.
The cross-sectional crown shape of lower molars also tends to distinguish most atelines from alouattines (Fig. 2.9). All alouattines have relatively long first lower molars. Length exceeds breadth by approximately 25 % or more. Here, again, Brachyteles converges on Alouatta, Stirtonia, Paralouatta, and Solimoea, while Caipora is an outlier among atelines. Other modern NWMs tend to have proportions similar to living atelines. First molars of species of Aotus, Callicebus, Pithecia, and Cebus, for example, have length/breadth ratios of 1.0–1.1 (Fig. 2.9). The overall functional continuity of this aspect with others that are part of the cristodont molar pattern indicates that elongation is a homologously derived element of crown design in alouattines, probably related to maximizing the linear length of shearing blades.
4 Discussion
4.1 Implications: Taxonomic Composition of the Fossil Alouattines and the Problem of Solimoea
The status of two of the three fossils at the focus of our character analysis has not been challenged. Stirtonia and Protopithecus present a robust, persuasive series of craniodental features tying them to Alouatta. There are also several postcranial features of the hip and thigh that may link Protopithecus and Alouatta (Halenar 2012). The affinities of Paralouatta have been debated (Rivero and Arredondo 1991; Horovitz and MacPhee 1999; Rosenberger 2002). As evident above, we have proceeded with the working hypothesis that the Cuban genus is an alouattine and refer readers elsewhere (Rosenberger 2002; Rosenberger et al. 2008) for arguments countering the notion that Paralouatta is a member of a monophyletic Caribbean group most closely affiliated with Callicebus. In nine of ten cranial features assessed here, Paralouatta shares the same derived state with Alouatta (Table 2.2). In two characters Paralouatta is “one step” less derived. In no cases are there any phenetic discrepancies to challenge the notion that these individual, intercorrelated elements are not homologous or functionally contrastive. We thus conclude that Paralouatta is a well-established alouattine.
The other species requiring attention is Solimoea acrensis. Kay and Cozzuol (2006) maintain that Solimoea acrensis is a stem ateline. The claim is based on a cladistic analysis using PAUP (Swofford 2002) of the two specimens they allocate to the taxon, an isolated lower molar inferred to be m1, the type specimen, and a referred maxillary fragment with P3–4, which is in poor condition. It is important to note that the Kay and Cozzuol (2006) analysis is not an independent assessment of morphological evolution among atelids because it is based on the “molecular scaffold” approach. In other words, the results of a molecular study were first used to arrange the topology of the tree. Then PAUP mapped characters onto the tree to produce the most economical distribution of states among the taxa.
We do not find the arguments compelling and suggest, alternatively, that Solimoea is an alouattine. There are major concerns that raise questions and warrant discussion: (1) the existence of distinctive phenetic similarities as well as a unique constellation of derived features shared by the type of Solimoea and alouattines, exclusively, and (2) Kay and Cozzuol’s reliance on characters from the maxillary specimen which may, in fact, not belong to the same taxon as the type.
The small-basin crown morphology of the type lower molar is far more similar to an alouattine than any of the wide-basined, extant atelines (Fig. 2.2). While Solimoea exhibits a crown pattern that appears to be less modified than the highly distinctive Stirtonia and Alouatta, it conforms to expectations of the alouattine morphotype. This configuration appears to be derived for atelids, based on character analysis and, especially, the expanded sense of alouattine diversity that is informed by taking Paralouatta into account. Solimoea shares with Alouatta and Stirtonia a morphological combination not seen elsewhere among NWM: (a) abbreviated, mesially narrowing, elevated trigonid and low, elongate, basined talonid; (b) obliquely oriented postvallid; (c) sharply angled cristid obliqua, forming a prominent ectoflexid; and (d) relatively long and narrow crown shape (Fig. 2.9). Buccally, the Solimoea lower molar also exhibits a resemblance to Paralouatta, whose maxillary molars demonstrate a primitive version of Alouatta-like ectoloph features as noted above. This is consistent with the Solimoea lower molar simply being more primitive, i.e., less of a “shearing folivore,” than the highly committed leaf-eaters Alouatta and Stirtonia. This functional and dietary inference is a conclusion also reached by Kay and Cozzuol (2006) based on quantification of shearing potential. Concerning resemblances between Solimoea and the ateline Brachyteles, some are evident in the angularity of the buccal aspect of the crown. However, this is probably partly due to primitiveness as well as a joint emphasis on shearing features.

Length breadth ratio (length/breadth) of m1 in selected platyrrhine species. Higher values indicate relatively longer and narrower teeth, a correlate of lengthened shearing blades, and are a derived feature associated with the alouattine clade and, independently, the semi-folivorous species Brachyteles. The crown morphology and length: breadth ratio of Solimoea aligns it with alouattines
The allocation of the maxillary specimen to the taxon is not convincing, for it is by no means evident that it is associated with the type lower molar. While there is a general conformity in the sizes of the lower molar and the upper premolars and they were recovered from the same locality, it would not be unusual for there to be several sympatric primate species and genera of similar size at an Amazonian locale (in this regard we note with interest that the gigantic Protopithecus and Caipora were found almost side by side in a cave, but their taphonomic histories remain a mystery). Kay and Cozzuol (2006) justify this allocation quantitatively, referencing the proportions of the crown areas (length × breadth) of the two specimens. They present a bivariate plot of m1 area vs. P4 area (Fig. 5, p. 677) based on a series of 13 Lagothrix lagotricha specimens and note that the plot point for the paired set of Acre fossils falls within the minimum convex polygon that bounds the distribution. We replicated and extended this exercise (Fig. 2.10) but arrive at a different conclusion. In our larger sample population of L. lagotricha, when jointly plotted the Acre specimens (termed Solimoea acrensis in the figure) do not lie within the polygon. It is also evident there is considerable overlap in the size relationships of m1 and P4 among species and genera of platyrrhines across a broad spectrum of body sizes at the 95 % confidence limits of populations, which undermines the taxonomic usefulness of this criterion (Fig. 2.10). The ellipses show that if the corresponding upper and lower teeth of most Brachyteles and Alouatta, or of most Ateles, Lagothrix, and Cebus, for example, were potted interchangeably by permutation, there would be no way of distinguishing or sorting confidently any individual tooth or tooth set to a species. Furthermore, our sample of howler monkeys uses A. seniculus only. If a smaller species was included in this case study, incidental sampling bias may even confound the metric associations of as many as six genera, Alouatta, Brachyteles, Ateles, Lagothrix, Cebus, and Solimoea.

Bivariate plot of m1 area (length × breadth) vs. P4 area (length × breadth) in selected platyrrhine species. A mixed species sample was used for Aotus, Cacajoa, and Chiropotes, including: A. vociferans, A. lemurinus, A. infulatus, A. nigriceps, A. trivirgatus, and A. brumbacki; Chiropotes albinasus and C. satans; and Cacajao calvus and C. melanocephalus. Mixed species samples were included to increase sample size and were only marginally more variable than single-species samples for this measure. Minimal convex polygons are shown in the main body of the figure and the inset shows 95 % confidence intervals. Solimoea is identified by an enlarged black dot in the inset. The multiple taxonomically overlapping proportions of these teeth across the range of sizes exhibited by platyrrhine species and genera, some of which can occur sympatrically, means that such size associations are not reliable as taxonomic identifiers in fossil assemblages like the Rio Acre sample involving Solimoea
In essence, the preserved morphology of the upper premolars is insufficient to properly test for an occlusal match with the lower molar, and compatibility in size is of little consequence. The premolars appear to be bunodont, rectangular, and of low relief, with large lingual occlusal surfaces, which is inconsistent with the non-bunodont, moderately high-relief morphology of the lower molar or with the latter’s abbreviated, oblique trigonid. It appears to us that these teeth may be mismatched taxonomically and, if so, this negates their utility in the generic diagnosis and cladistic analysis.
4.2 Interrelationships, Craniodental Morphology, and Adaptations of Fossil Alouattines
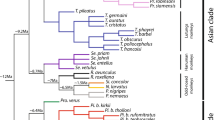
The cladistic interrelationships derived from our character analysis are summarized in Fig. 2.11. Our overall results continue to support prior arguments that Stirtonia is the fossil most closely related to Alouatta. For example, the upper molar morphology of Paralouatta tends to reinforce the Alouatta-Stirtonia linkage by default because the Cuban form’s crowns are blunter, but its upper molars present a W-shaped ectoloph and moderately well-developed stylar elements, structural features that eventually became trenchant shearing surfaces in Alouatta and Stirtonia. The lingual aspect of Paralouatta upper molars also had not yet developed the sharp, lobe-like hypocone, which is prominent in Alouatta and Stirtonia.

Cladogram showing the proposed interrelationships of alouattine platyrrhines. Dotted lines indicate all possible positions for Solimoea. Images are for illustrative purposes and are not shown to scale
With a cylindrical braincase and constricted nuchal region, synapomorphies shared with Alouatta but combinatorially absent in Protopithecus, Paralouatta is not only more derived than Protopithecus in the direction of Alouatta. It also bears a distinctly closer phenetic resemblance to howler monkeys. This offsets Protopithecus as a basal member of the alouattine clade given what we know currently of their diversity. In the basicranium as well, Protopithecus appears to exhibit the ancestral end of the alouattine morphocline while Alouatta and Paralouatta occupy the opposite pole. Equally important, Protopithecus has a very different dental gestalt, lacking both the reduced incisors of Alouatta and Paralouatta and a cristodont molar pattern as seen in Alouatta. Among all the fossils, Protopithecus retains the largest combination of dental and cranial features consistent with the alouattine morphotype. In a general way, the morphological pattern is concordant with Rosenberger and Strier’s (1989) proposal of a Lagothrix-like craniodental morphology being ancestral for alouattines and atelines.
Based on one lower molar, the position of Solimoea is still difficult to fix. The shape of this tooth does not conform to the apparently open-basin crown morphology of the damaged Protopithecus upper molar (or with the bunodont pattern of existing lower premolars). Its small-basin design is not consistent with the advanced atelines, Lagothrix and Ateles. Resemblances to Brachyteles are confined to the more primitive buccal aspect of the crowns, and this generalized angularity of shape is also shared with alouattines. Solimoea most closely resembles Alouatta and Stirtonia overall. The buccally flaring, elevated protoconid; strongly angled pre- and post-hypoconid cristae; and strong trigonid-talonid height differential appear to be a derived combination that would facilitate folivory through selection for additional or more efficient shearing.
Cranially, both Protopithecus and Paralouatta present a mixture of features resembling the unique patterns exhibited by Alouatta in areas of the basicranium and nuchal region, overall braincase shape, and facial structure. The more detailed resemblances shared by Paralouatta and Alouatta imply an important functional overlap that appears to be related to a novel organization of the skull. These probably relate to fixation of an enlarged hyolaryngeal mechanism in the neck between the rami of the mandible and to a large, cantilevered head. The general organization of Protopithecus clearly indicates a shift from a more general, Lagothrix-like pattern in the Paralouatta-Alouatta direction.
An unexpected outcome of this study involves two related indicators of relative brain size, endocranial volume and foramen magnum area. The same reduced scaling conditions were observed among all alouattines known from cranial material but not in any of the atelines, not even the one genus with a pervasive tendency to eat leaves, Brachyteles. As noted previously, the relatively small brain size in Alouatta has been regarded as part of its folivorous feeding adaptation, so its occurrence in Protopithecus, with a decidedly frugivorous dentition, is thus counterintuitive. Noteworthy is a recent study (Krupp et al. 2012) that independently confirms the relatively small size of the Protopithecus brain and demonstrates, via an endocranial cast, its Alouatta-like shape and surface morphology. Given that the Protopithecus dentition is not howler monkey-like, these findings are consistent with the hypothesis that the shift toward folivory in Alouatta may have been predicated on an earlier reliance on seeds (Rosenberger et al. 2011). This initial dependence on seeds could have benefitted from similar energy-saving features like small brains (Rosenberger et al. 2011), if selection for large brains (possibly driven by features such as functional association with a large, complex social organization) was absent. If correct, the comparable small brains of Protopithecus, Paralouatta, and Alouatta more likely reflect a derived de-encephalization than a primitively atelid small-brain pattern, though parceling out these historical factors remains a difficult proposition as we noted above.
Of the three fossil genera, only Stirtonia, the howler monkey’s closest relative known thus far, can be considered a comparably committed leaf-eater by the detailed functional similarities shared with Alouatta in molar morphology and by its inferred body mass. The cheek teeth of Paralouatta are more bunodont and given to wearing more flatly, contrasting with Alouatta, which exposes lines of dentine along the crown perimeter and stems from a thin-enamel occlusal design that emphasizes and maintains shearing. Thus is it likely that Paralouatta molars are not designed optimally for shredding leaves.
Upper molars of Protopithecus lack crested shearing perimeter lines, and the large-basined crowns do not seem to resemble either the Alouatta-Stirtonia pattern or the morphology of Paralouatta. The lower premolars are also notably bunodont. Its incisor teeth are wide, spatulate, and relatively large in cross section, which is consistent with a generalized frugivory as opposed to pitheciine-like, sclerocarpic harvesting (Kinzey 1992; Rosenberger 1992). The summed cross-sectional area of Protopithecus incisors is 82 % of the area of the upper molar. Comparable ratios for leaf-eating folivores are 41 % in Alouatta seniculus and 46 % for Brachyteles arachnoides. Thus, one of the three alouattine fossils shows consistently strong indications of a non-leafy diet in aspects involving both anterior and posterior teeth. Paralouatta is also suggestive of the same.
5 Conclusion
With the addition of fossils, the subfamily Alouattinae now consists of four extinct genera in addition to Alouatta: Stirtonia, Paralouatta, Protopithecus, and Solimoea. It understates the case to say that the only extant alouattine is a platyrrhine outlier in its morphology, trophic adaptations, and clamorous mode of communicating. But as the diversity of this group is filled in by the discovery of related fossils, it becomes apparent that living howler monkeys are also something of an outlier among alouattines as well, for there is more than one “kind” of alouattine. Stirtonia is currently the howler monkey’s closest relative and its teeth are barely distinguishable from Alouatta, which suggests a fundamentally similar diet. The other fossils help strip back the feeding specializations of the Alouatta-Stirtonia group to disclose more primitive anatomies and different dietary adaptations and thus help close the trophic gap between alouattines and atelines. The other crucial aspect regarding evolutionary history revealed by the fossils comes from the cranium, which helps trace another signature adaptation of Alouatta, howling. We arrive at these conclusions regarding phylogeny, diversity, divergence, and evolutionary adaptation as outcomes reflecting the particular approach used in our analysis.
Confidence that Solimoea is an alouattine is elevated by the observation that its morphology falls within or along a morphocline that circumscribes the anatomical patterns of genera whose monophyletic affinities with Alouatta are corroborated independently by cranial morphology. It is also affirmed by the observation that what limited functional morphology can be drawn from the Solimoea tooth, i.e., its inferred mechanical shearing potential, is consistent with the notion that alouattines, living and extinct, exhibit a range of dental features relating to frugivory-folivory but that neither of the two most basal genera are projected to be as highly committed to folivory as are Alouatta and Stirtonia. In other words, the functional morphology of Solimoea is consistent with models of alouattine evolution, which predict what is self-evident in the broader context of NWM evolution—alouattines more primitive than Alouatta would have existed, and they would have exhibited a lesser emphasis on shearing features. On the other hand, there is nothing in any of the models that would predict primitive atelines would also resemble alouattines, only that they are not likely to be comparable to either Ateles or Brachyteles dietarily and morphologically. A Lagothrix-like dentition may still serve adequately as the default model of a morphotypic ateline dentition (Rosenberger and Strier 1989).
Of the two extinct genera known by cranial remains, the alouattine affinities of the Brazilian Protopithecus seems well established although there has been debate about the Cuban Paralouatta (e.g., MacPhee and Horovitz 2002; Rosenberger 2002). Here, too, functional morphology and a morphoclinal perspective weigh heavily in favor of Paralouatta being related cladistically to howler monkeys. With Protopithecus and Alouatta representing the range of alouattine extremes in terms of cranial character states and patterns, Paralouatta seems comfortably nested near the middle anatomically but decidedly closer to Alouatta at the more derived end of the spectrum. The functional explanation we propose as the underlying engine behind this transformation series relates more to howling adaptations than to mastication and diet. Protopithecus, with its relatively extended basicranium and uptilted face, exhibits the beginnings of a trend toward a greatly enlarged subbasal space, and this may represent a primitive version of the architecture supporting an enlarged hyolaryngeal apparatus. Paralouatta is clearly even more Alouatta-like in this regard (see also Halenar 2012).
The literature’s essentially unanimous endorsement of the hypothesis that Stirtonia is an alouattine quite closely related to modern howler monkeys is reinforced by finding that comparable aspects of the molars of Paralouatta (and Solimoea) are apparently more primitive, while the lesser known dentition of Protopithecus presents a different anatomical pattern, perhaps closer to atelines and thus possibly morphotype-like for alouattines. The evidence points strongly to the assessment that this most basal genus of the clade was decidedly frugivorous.
It is difficult to say how long the alouattine clade has been evolving. Recent molecular studies vary in their estimates. For example, Opazo et al. (2006) posit the origins of the clade at 16.75 million years. Schrago (2007) estimates a mean divergence date for Alouatta at 12.4 MA but this involves a broad range of ages, 9.1–18.6 MA. In a narrower study, Hodgson et al. (2009) estimate the divergence of Ateles relative to non-atelid platyrrhines, thus Alouatta by implication, at 18.0 MA, with a range of 15.7–21.6 MA. The fossil record offers an indication of a Miocene differentiation as well. There is one report (Tejedor 2002) of possible alouattines existing in Patagonia in the late-early Miocene, but it is based on meager evidence, isolated canine teeth showing certain resemblances to Alouatta. This would be a pre-La Venta occurrence, about four million years prior to Stirtonia victoriae and the younger S. tatacoensis. The isolated astragalus from Cuba allocated to Paralouatta marianae is dated stratigraphically to ~17 MA (MacPhee et al. 2003), also antedating La Venta, but the affinities of this bone must be considered tenuous.
What can be said with some confidence is that by La Venta times, ~11–13 MA, and at the younger Acre site, ~ 8 MA, the modernized members of the Alouatta branch of the radiation existed, probably as committed howler monkey-like leaf-eaters living in the formative Amazon basin as this ecological community was being assembled (Rosenberger et al. 2009). The fossils outside this zone, Paralouatta varonai in Cuba and Protopithecus brasiliensis in eastern Brazil, shed light in a different direction, toward the remote origins of Alouatta. Despite the recent geological ages of these two species, they retain a variety of primitive morphologies and occupy positions on the alouattine cladogram basal to the differentiation of the Stirtonia-Alouatta lineage. This means that alouattines branching off before the La Venta horizon and outside of Amazonia may have been less committed adaptively to masticating leaves, and may thus come closer to approximating the original adaptive gestalt of the group.
Dentally, the fossils, all relatively large-bodied platyrrhines and all expected to have used, as atelids, fully prehensile tails, comprise an adaptive radiation of mixed feeders within the frugivore-folivore spectrum. At least two “stages” in the morphological evolution of the skull that relates to howling can be discerned. Paralouatta is sufficiently similar to living howler monkeys in the cranial base and occipital region to suggest the same set of novel specializations were present in terms of the biological roles of the hyoid complex and occipital region as they relate to vocalization and head carriage. Protopithecus is less advanced in that direction. But it signifies that at the basal branch of the radiation, the alouattine clade had already shifted toward some semblance of the loud-calling lifestyle of Alouatta before the clade produced the specialized capacity to harvest and masticate leaves. Long call adaptations also seem to have preceded the evolution of the howler monkey’s postcranial skeletal adaptations (see Hartwig and Cartelle 1996; Jones 2008; Halenar 2011a, b), also emerging in an alouattine that was more frugivorous, as exemplified by Protopithecus. As morphologists, we emphasize here that a shift in social behavior, possibly imprinted on the ancestral cranial morphology of the lineage, may have been instrumental in the successful differentiation of alouattines prior to the evolution of the modified dental and locomotor adaptations that one might have expected as essential niche characteristics of this radiation.
We have elsewhere suggested (Rosenberger et al. 2009) that the natural history and biogeography of living Alouatta species, potential pioneers due to their dietary flexibility, suggests the possibility that the genus arose not in the greater Amazon basin but elsewhere on the continent in less lush habitats. This idea appears to be consistent with the interpretations we present here, since two genera more basal to the Alouatta-Stirtonia clade occur outside Amazonia.
Another important insight is that relatively small brain sizes evolved among alouattines before their intense dental commitment to leaf-eating. Protopithecus appears to be a rare example of a small-brained, frugivorous anthropoid. This raises several interesting questions. Is there an evolutionary link between ostentatious howling, which may well have been part of the Protopithecus repertoire, and relatively modest brains, perhaps as a morphological constraint on cranial design? Have we overemphasized the physiological and adaptive connections between small brains and leaf-eating? Can facultative leaf-eating in a mixed feeder, perhaps enabled by large body size and concomitantly large guts—Protopithecus may be such an example—form a trophic substrate that would motivate selection for de-encephalization? Is relative brain size more sensitive to selection supporting folivorous or semi-folivorous diets (Rosenberger et al. 2011), or facultative leaf-eating, than dentition? Could de-encephalization have evolved as a seed-eating adaptation in the absence of selection for brain size increase? We can only speculate that howling, small brains, and leaf-eating are interconnected as low-energy balancing factors of potential adaptive value: long-distance advertisement that requires little movement or exposure, a brain that can be metabolically maintained relatively cheaply, and a food source that requires little exercise to acquire and produces energy slowly and at low dosages. These characteristics aptly describe facets central to the howler monkey lifestyle, but they offer little in the way of explaining how and why Alouatta came to be. The first batch of diverse alouattine fossils suggests some answers lay buried.
Abbreviations
- %:
-
Percent
- CT:
-
Computed Tomography
- e.g.:
-
For example
- Fig.:
-
Figure
- Figs.:
-
Figures
- i.e.:
-
In other words
- kg:
-
Kilograms
- m1:
-
First lower molar
- m3:
-
Third lower molar
- M1:
-
First upper molar
- MA:
-
Millions of years
- mm:
-
Millimeters
- NWM:
-
New World monkeys
- P3:
-
Third upper premolar
- p4:
-
Fourth lower premolar
- P4:
-
Fourth upper premolar
References
Anthony MRL, Kay RF (1993) Tooth form and diet in ateline and alouattine primates: reflections on the comparative method. Am J Sci 293:356–382
Biegert J (1963) The evaluation of characteristics of the skull, hands and feet for primate taxonomy. In: Washburn SL (ed) Classification and human evolution. Aldine, Chicago
Cartelle C, Hartwig WC (1996) A new extinct primate among the Pleistocene megafauna of Bahia, Brazil. Proc Natl Acad Sci U S A 93:6405–6409
Clutton-Brock TH, Harvey PH (1980) Primates, brains and ecology. J Zool 207:151–169
Cole TM (1995) Comparative craniometry of the Atelinae (Platyrrhini, Primates): function, development, and evolution. PhD Dissertation. State University of New York at Stony Brook, New York
Conroy GC (1987) Problems of body-weight estimation in fossil primates. Int J Primatol 8:115–137
Cooke SB, Halenar LB (2012) The evolution of body size in the Caribbean primates (Abstract). International Primatological Society meetings, 13 August 2012, Cancun
Delson E, Rosenberger AL (1984) Are there any anthropoid primate living fossils? In: Eldredge N, Stanley SM (eds) Living fossils. Springer, New York
DiFiore A, Campbell C (2007) The Atelines: Variation in ecology, behavior, and social organization. In: Campbell C, Fuentes A, MacKinnon K, Panger M, Bearder S (eds) Primates in Perspective. New York, Oxford University Press
Eaglen RH (1984) Incisor size and diet revisited: the view from a platyrrhine perspective. Am J Phys Anthropol 64:263–275
Eisenberg JF (1981) The mammalian radiations. University of Chicago, Chicago
Fleagle JG (1999) Primate adaptation and evolution. Academic, New York
Fleagle JG, Kay RF, Anthony MRL (1997) Fossil New World monkeys. In: Kay RF, Madden RH, Cifelli RL, Flynn JJ (eds) The history of a Neotropical Fauna: vertebrate paleobiology of the Miocene of tropical South America. Smithsonian Institution, Washington, DC
Flynn JJ, Guerrero J, Swisher CC (1997) Geochronology of the Honda group, Columbia. In: Kay RF, Madden RH, Cifelli RL, Flynn JJ (eds) The history of a Neotropical fauna: vertebrate paleobiology of the Miocene of tropical South America. Smithsonian Institution, Washington, DC
Ford SM, Corruccini RS (1985) Intraspecific, interspecific, metabolic and phylogenetic scaling in platyrrhine primates. In: Jungers WL (ed) Size and scaling in primate biology. Plenum, New York
Gould SJ (1977) Ontogeny and phylogeny. Harvard University, Cambridge
Halenar LB (2008) Agreement between interspecific variation in vocalization patterns and cranial base morphology in Alouatta: preliminary results and future directions (Abstract). Am J Phys Anthropol (Suppl. 46):111
Halenar LB (2011a) Reconstructing the locomotor repertoire of Protopithecus brasiliensis. I. Body size. Anat Rec 294:2024–2047
Halenar LB (2011b) Reconstructing the locomotor repertoire of Protopithecus brasiliensis. II. Forelimb morphology. Anat Rec 294:2048–2063
Halenar LB (2012) Paleobiology of Protopithecus brasiliensis, a plus-sized Pleistocene Platyrrhine from Brazil. PhD Dissertation, CUNY Graduate Center, New York
Halenar, LB, Rosenberger, AL (2013) A closer look at the “Protopithecus” fossil assemblages: new genus and species from Bahia, Brazil. J Hum Evol 65:374–390
Hartwig WC (1995) A giant New World monkey from the Pleistocene of Brazil. J Hum Evol 28:189–196
Hartwig WC (1996) Perinatal life history traits in New World monkeys. Am J Primatol 40:99–130
Hartwig WC (2005) Implications of molecular and morphological data for understanding atelid phylogeny. Int J Primatol 26:999–1015
Hartwig WC, Cartelle C (1996) A complete skeleton of the giant South American primate Protopithecus. Nature 381:307–311
Hartwig W, Meldrum DJ (2002) Miocene platyrrhines of the northern Neotropics. In: Hartwig WC (ed) The primate fossil record. Cambridge University, Cambridge
Hartwig WG, Rosenberger AL, Norconk M, Young Owl M (2011) Relative brain size, gut size and evolution in New World monkeys. Anat Rec 294:2207–2221
Harvey PH, Clutton-Brock TH (1985) Life history variation in primates. Evolution 39:559–581
Hershkovitz P (1970) Notes on Tertiary platyrrhine monkeys and description of a new genus from the late Miocene of Colombia. Folia Primatol 12:1–37
Hershkovitz P (1972) The recent mammals of the Neotropical Region: a zoogeographic and ecological review. In: Keast A, Erk FC, Glass B (eds) Evolution, mammals, and southern continents. State University of New York, Albany
Hershkovitz P (1977) Living New World monkeys (Platyrrhini), with an introduction to the primates, vol 1. University of Chicago, Chicago
Hodgson JA, Sterner KN, Matthews LJ, Burrell AS, Jania RA, Raaum RL, Stewart C-B, Disotell TR (2009) Successive radiations, not stasis, in the South American primate fauna. Proc Natl Acad Sci U S A 106:5534–5539
Horovitz I, MacPhee RDE (1999) The quaternary Cuban platyrrhine Paralouatta varonai and the origin of the Antillean monkeys. J Hum Evol 36:33–68
Jerison HJ (1973) Evolution of the brain and intelligence. Academic, New York
Jones AL (2008) The evolution of brachiation in ateline primates, ancestral character states and history. Am J Phys Anthropol 137:123–144
Kay RF (1975) The functional adaptations of primate molar teeth. Am J Phys Anthropol 43:195–216
Kay RF, Cozzuol MA (2006) New platyrrhine monkeys from the Solimoes Formation (late Miocene, Acre State, Brazil). J Hum Evol 50:673–686
Kay RF, Frailey CD (1993) Large fossil platyrrhines from the Rio Acre local fauna, late Miocene, western Amazonia. J Hum Evol 25:319–327
Kay RF, Hylander WI (1978) The dental structure of mammalian folivores with special reference to Primates and Phalangeroidea (Marsupialia). In: Montgomery GG (ed) The ecology of arboreal folivores. Smithsonian Institution, Washington, DC
Kay RF, Madden R, Plavcan JM, Cifelli RL, Diaz JG (1987) Stirtonia victoriae, a new species of Miocene Colombian primate. J Hum Evol 16:173–196
Kay RF, Johnson D, Meldrum DJ (1998) A new pitheciin primate from the middle Miocene of Argentina. Am J Primatol 45:317–336
Kinzey WG (1992) Dietary adaptations in the Pitheciinae. Am J Phys Anthropol 88:499–514
Krupp A, Cartelle C, Fleagle JG (2012). Size and external morphology of the brains of the large fossil platyrrhines Protopithecus and Caipora. (Abstract) Am J Phys Anthropol (Suppl. 54):186
Lund PW (1838) Blik paa Brasiliens dyreverden for sidste jordomvaeltning. Det Kongelige Danske Videnskabernes Selskabs Naturvidenskabelige og Mathematiske Afhandlinger 8:61–144
MacPhee RDE, Horovitz I (2002) Extinct quaternary platyrrhines of the Greater Antilles and Brazil. In: Hartwig WC (ed) The primate fossil record. Cambridge University, Cambridge
MacPhee RDE, Horovitz I (2004) New craniodental remains of the Quaternary Jamaican monkey Xenothrix mcgregori (Xenotrichini, Callicebinae, Pitheciidae), with a reconsideration of the Aotus hypothesis. Am Mus Novitates 3434:1–51
MacPhee RDE, Iturralde-Vinent MA (1995) Origin of the Greater Antillean land mammal fauna, 1: new Tertiary fossils from Cuba and Puerto Rico. Am Mus Novitates 3141:1–32
MacPhee RDE, Meldrum J (2006) Postcranial remains of the extinct monkeys of the Greater Antilles (Platyrrhini, Callicebinae, Xenotrichini), with a consideration of semiterrestriality in Paralouatta. Am Mus Novitates 3516:1–65
MacPhee RDE, Horovitz I, Arredondo O, Vasquez OJ (1995) A new genus for the extinct Hispaniolan monkey Saimiri bernensis Rimoli, 1977, with notes on its systematic position. Am Mus Novitates 3134:1–21
MacPhee RDE, Iturralde-Vinent M, Gaffney ES (2003) Domo de Zaza, an early Miocene vertebrate lin South-Central Cuba, with notes on the tectonic evolution of Puerto Rico and the Mona Passage. Am Mus Novitates 3394:1–42
Martin RD (1984) Body size, brain size and feeding strategies. In: Chivers D, Wood B, Bilsborough A (eds) Food acquisition and processing in primates. Plenum, New York
Martin RD (1990) Primate origins and evolution: a phylogenetic reconstruction. Chapman and Hall, London
Opazo JC, Wildman DE, Prychitko T, Johnson RM, Goodman M (2006) Phylogenetic relationships and divergence times among New World monkeys (Platyrrhini, Primates). Mol Phylo Evol 40:274–280
Rivero M, Arredondo O (1991) Paralouatta varonai, a new quaternary platyrrhine from Cuba. J Hum Evol 21:1–11
Rosenberger AL (1980) Gradistic views and adaptive radiation of platyrrhine primates. Z Morphol Anthropol 71:157–163
Rosenberger AL (1992) Evolution of feeding niches in new world monkeys. Am J Phys Anthropol 88:525–562
Rosenberger AL (2002) Platyrrhine paleontology and systematics: the paradigm shifts. In: Hartwig WC (ed) The primate fossil record. Cambridge University, Cambridge
Rosenberger AL, Kinzey WG (1976) Functional patterns of molar occlusion in platyrrhine primates. Am J Phys Anthropol 45:281–298
Rosenberger AL, Strier KB (1989) Adaptive radiation of the ateline primates. J Hum Evol 18:717–750
Rosenberger AL, Setoguchi T, Shigehara N (1990) The fossil record of callitrichine primates. J Hum Evol 19:209–236
Rosenberger AL, Halenar LB, Cooke SB, Hartwig WC (2008) Morphology and evolution of the spider monkey, genus Ateles. In: Campbell C (ed) Spider monkeys: behavior, ecology and evolution of the genus Ateles. Cambridge University, New York
Rosenberger AL, Tejedor MF, Cooke SB, Pekkar S (2009) Platyrrhine ecophylogenetics, past and present. In: Garber P, Estrada A, Bicca-Marques JC, Heymann EW, Strier KB (eds) South American primates: comparative perspectives in the study of behavior, ecology and conservation. Springer, New York
Rosenberger AL, Halenar LB, Cooke SB (2011) The making of platyrrhine semi-folivores: models for the evolution of folivory in primates. Anat Rec 294:2112–2130
Ross C, Ravosa M (1993) Basicranial flexion, relative brain size, and facial kyphosis in nonhuman primates. Am J Phys Anthropol 91:305–324
Ruff CB (2003) Long bone articular and diaphyseal structure in Old World monkeys and apes II: estimation of body mass. Am J Phys Anthropol 120:16–37
Schrago CG (2007) On the time scale of the New World primate diversification. Am J Phys Anthropol 132:344–354
Schultz AH (1955) The position of the occipital condyles and of the face relative to the skull base in primates. Am J Phys Anthropol 13:97–120
Sears KE, Finarelli JA, Flynn JJ, Wyss AR (2008) Estimating body mass in New World “monkeys” (Platyrrhini, Primates) from craniodental measurements, with a consideration of the Miocene platyrrhine, Chilecebus carrascoensis. Am Mus Novitates 3617:1–29
Setoguchi T (1980) Discovery of a fossil primate from the Miocene of Colombia. Monkey 24:64–69
Setoguchi T, Rosenberger AL (1985) Miocene marmosets: first evidence. Intl J Primatol 6:615–625
Setoguchi T, Watanabe T, Mouri T (1981) The upper dentition of Stirtonia (Ceboidea, Primates) from the Miocene of Colombia, South America and the origin of the postero-internal cusp of upper molars of howler monkeys (Alouatta). Kyoto University Overseas Research Reports of New World Monkeys 3:51–60
Stephan H (1972) Evolution of primate brains: a comparative anatomical investigation. In: Tuttle RH (ed) The functional and evolutionary biology of primates. Aldine, Chicago
Stephan H, Andy OJ (1964) Quantitative comparison of brain structures from insectivores to primates. Am Zool 4:59–74
Stirton RA (1951) Ceboid monkeys from the Miocene of Colombia. Univ Calif Pub Geol Sci 28:315–356
Strier KB (1992) Ateline adaptations: behavioral strategies and ecological constraints. Am J Phys Anthropol 88:515–524
Swofford DL (2002) PAUP. Phylogenetic analysis using parsimony. Version 4.0b10 (Altivec). Sinauer, Sunderland
Szalay FS, Bock WJ (1991) Evolutionary theory and systematics: relationships between process and patterns. J Zool Syst Evol Res 29:1–39
Szalay FS, Delson E (1979) Evolutionary history of the primates. Academic, New York
Tejedor MF (2002) Primate canines from the early Miocene Pinturas Formation, southern Argentina. J Hum Evol 43:127–141
Tejedor MF, Tauber AA, Rosenberger AL, Swisher CC, Palacios ME (2006) New primate genus from the Miocene of Argentina. Proc Natl Acad Sci U S A 103:5437–5441
Tejedor MF, Rosenberger AL, Cartelle C (2008) Nueva especie de Alouatta (Primates, atelineae) del Pleistoceno tardío de Bahía, Brasil. Ameghin 45:247–251
Wiley DF, Amenta N, Alcantara DA, Ghosh D, Kil YK, Delson E, Harcourt-Smith W, Rohlf FJ, St. John K, Hamann B (2005) Evolutionary morphing (extended abstract and associated video presentation). Proceedings of IEEE visualization conference 2005, pp. 1–8
Acknowledgements
We owe much to many: to the editors of this volume for inviting us to contribute; to the museums mentioned above that make our research possible, especially our home institution, the American Museum of Natural History; to Leandro Salles and Castor Cartelle and their museums, MNRJ and PUC MINAS, for making this project possible; to Marilyn Norconk for sharing her insights on platyrrhines; to Andi Jones and Mike Rose for discussions on the Brazilian fossils and use of their photographs; to the agencies at Brooklyn College (Tow) and the City University of New York (PSC CUNY) for financial assistance to ALR; to the Wenner-Gren Foundation and CUNY NYCEP for a postdoctoral fellowship awarded to MFT which helped support our collaboration; and to NSF DDIG awards to LBH (0925704) and SBC (0726134) and an Alumnae Association of Barnard College Graduate Fellowship to help support SBC in her research on Caribbean primates. We thank the reviewers and editors for many helpful suggestions. The software package PAST was employed for several computations and charts (http://folk.uio.no/ohammer/past/).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2015 Springer Science+Business Media New York
About this chapter
Cite this chapter
Rosenberger, A.L. et al. (2015). Fossil Alouattines and the Origins of Alouatta: Craniodental Diversity and Interrelationships. In: Kowalewski, M., Garber, P., Cortés-Ortiz, L., Urbani, B., Youlatos, D. (eds) Howler Monkeys. Developments in Primatology: Progress and Prospects. Springer, New York, NY. https://doi.org/10.1007/978-1-4939-1957-4_2
Download citation
DOI: https://doi.org/10.1007/978-1-4939-1957-4_2
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4939-1956-7
Online ISBN: 978-1-4939-1957-4
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)