
Abstract
Bone research in osteoporosis has quite rightly focused on the mineralised component of bone as this is the component that is ultimately responsible for bone strength. However, the non-mineralised component of bone, i.e. the bone marrow, is many times more metabolically active and responsive than the mineralised component of bone. Despite this, the bone marrow has been relatively overlooked with regard to the pathogenesis of osteoporosis and related conditions. This has changed with magnetic resonance imaging and positron emission tomography allowing non-invasive quantification of bone marrow physiology and pathology on a large scale. Aspects of the bone marrow that can be evaluated on imaging are marrow fat content, perfusion, molecular diffusion and metabolic activity. There are many ways in which bone marrow metabolism may potentially influence bone metabolism. For example, the bone marrow forms the microenvironment of biologically relevant endosteal and trabecular bone and this bone may be responding to changes in the bone marrow. Similarly, the bone marrow contains pluripotent mesenchymal stem cells with the ability to differentiate preferentially along either haematopoetic, adipocytic or osteoblastic cell lines. Preliminary research has shown how bone loss in senile osteoporosis mass is accompanied by scalar changes in marrow fat content, marrow perfusion and marrow diffusion. Similar to the bone loss of osteoporosis, the bone marrow changes in osteoporosis represent an exaggeration of physiological age-related change. Bone marrow changes occur in synchrony rather than pre- or post-date changes in the mineralised component of bone. Whether the bone marrow is an active contributor or a passive bystander to physiological and osteoporotic bone loss remains to be seen.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Background
Bone densitometry, high resolution imaging techniques to access bone architecture and advanced image analytic platforms have greatly improved our understanding of osteoporosis particularly with respect to bone structure and strength prediction over the past three decades (Link 2012). This osteoporotic research has quite rightly focused on the hard tissue component of bone as this is the component that ultimately gives bone its strength. The marrow cavity, nevertheless, also forms a major constituent of bone and has, until recently, received relatively less attention regarding osteoporotic research. Yet, bone marrow is a more metabolically active tissue than mineralised bone tissue and several plausible mechanisms existing through which the bone marrow may influence bone metabolism. Our knowledge of bone marrow metabolism has been greatly assisted by MR and PET-CT technology which allows, for the first time, a quantitative non-invasive study of the bone marrow. This study has focused on lifelong physiological changes in the bone marrow as well as how the bone marrow is affected in common musculoskeletal disorders such as osteoporosis, marrow infiltration, osteoarthritis and disc degeneration. The bone marrow is one of the most voluminous and metabolically active organs in the human body, that undergoes progressive change throughout life and is involved in perfusion or nutrition of adjacent structures. It is hoped that the bone marrow may provide some answers that exist regarding the pathogenesis of these common musculoskeletal diseases.
For example, with respect to osteoporosis, one could argue that current densitometry techniques and even high resolution imaging techniques are diagnosing osteoporosis too late (Griffith et al. 2010). By the time osteoporosis is recognised by densitometric techniques, bone strength is already significantly impaired. Pharmaceutical agents can stall or retard the osteoporotic process but will not return bone strength to normal. Thinned cortices and trabeculae may thicken with osteoporotic treatment, but those trabeculae that have absorbed will not return such that impaired bone strength persists even with a good treatment response. Also, for subjects with normal bone density or low bone mass (osteopenia), prediction of which subjects will progress to more severe degrees of bone loss and impairment of bone strength, is not sufficiently accurate to select those patients which will particularly benefit from osteoporotic treatment. In addition, osteoporosis is associated with several other conditions such as steroid use, atherosclerosis, vascular calcification, diabetes, dyslipidaemia and Alzheimer’s disease though the pathogenetic mechanisms linking these diseases to osteoporosis are not fully understood (Manolagas and Almeida 2007). One can appreciate that finding a contributory link between these diseases and osteoporosis would be a significant step towards the development of a common single therapy.
2 Bone Marrow
The bone marrow is supported by trabeculae and a fibrous tissue retinaculum and surrounded by a bone cortex of variable thickness ranging from approximately 1 to 5.5 mm. The actual composition of the bone marrow varies with anatomical location, physiological well-being and age (Hwang and Panicek 2007) though in general is made up of trabecular bone (approx 20 %), fatty marrow (approx 50 %), non-fatty functioning marrow (approx 25 %) and vascular channels (approx 5 %). Non-fatty functioning marrow comprises cells derived from the haematopoetic cell line (erythrocytes, granulocytes, lymphocytes, monocytes, platelets and osteoclasts) as well as stem cells.
At birth, the bone marrow is nearly entirely haematopoetic except for the epiphyses and apophyses which are mainly fat. With maturation, the haematopoetic appendicular marrow converts to a predominantly fatty marrow in a symmetrical centripetal fashion from the periphery to the central skeleton (Hwang and Panicek 2007). Superimposed on this centripedal conversion, haematopoetic marrow converts to fatty marrow in the tubular bones proceeding from the diaphysis to metaphysis (Hwang and Panicek 2007; Hartsock et al. 1965) (Fig. 1). At 10 years of age, marrow conversion of red to fatty marrow has begun in the diaphyses (Hwang and Panicek 2007). By 30 years of age, some red marrow remains only in the proximal metaphyses, and the axial skeletal (pelvis, spine, scapulae, clavicles, sternum and skull). In the event of an increased functional demand for haematopoesis such as smoking or malignancy, this sequence of events can reconvert with fatty marrow reconverting to red marrow in a reverse, symmetrical centrifugal manner (Poulton et al. 1993).

From birth, red marrow converts to fatty marrow from the periphery to the central skeleton. Superimposed on this centripedal pattern, red marrow converts to fatty marrow in the tubular bones proceeding from diaphysis to metaphysis until by the age of 20 years only the proximal metaphyseal area contains appreciable red marrow
Red and yellow marrow areas are not composed purely of either non-fatty cells or fat cells, respectively. ‘Red marrow’ typically contains about 60 % haematopoetic cells and about 40 % fat cells (Fig. 2a, b) while ‘fatty marrow’ contains about 5 % haematopoetic cells and about 95 % adipocytes (Hwang and Panicek 2007; Steiner et al. 1993). In other words, ‘fatty marrow’ tends to be more ‘pure’ than haematopoetic marrow. Fat cells (adipocytes) as expected contain more lipid than haematopoetic cells while haematopoetic cells contain slightly more water and protein than adipocytes. The approximate chemical composition of fatty marrow is about 80 % lipid, 15 % water and 5 % protein while that of red marrow is about 50 % lipid, 35 % water and 15 % protein (Hwang and Panicek 2007; Steiner et al. 1993) (Fig. 3). This is relevant since quantification techniques such as MR spectroscopy (MRS) use the fat: water ratio to determine the % marrow fat fraction [also referred to as marrow fat content (%)].

a Histology of predominantly red marrow. There is still quite an abundance of fat cells present. b Histology of predominantly fatty marrow. There are only a few red cells present. In other words fatty marrow is more fatty than red marrow is red

Chemical composition of red marrow and fatty marrow. Haematopoetic marrow contains more water than fatty marrow
The pH of the marrow cavity is not known though the pH of extravascular tissues is generally lower than that of arterial blood (pH 7.4) and venous blood (pH 7.36) (Arnett 2010). The oxygen tension of normal bone marrow is about 52 mmHg (6.6 %) which is lower than that of arterial blood (95 mmHg, ~12 %) and higher than that of veno-capillary blood (40 mmHg, ~5 %) (Arnett 2010). In normal tissues other than the bone marrow, median interstitial oxygen tension levels measure ~3–9 % (Arnett 2010).
The marrow cavity is supplied by large nutrient arteries that pass through the cortex into the medullary canal (Travlos 2006). Ascending and descending nutrient branches give rise to small thin-walled arterioles that extend towards the periphery where they give rise to capillaries piercing the bone cortex and also merge with thin venous sinuses lined by flat endothelial cells. These endothelial cells lack a tight junction though may overlap or interdigitate facilitating two-way passage of haematopoetic cells (Travlos 2006; Lichtman 1981; Brookes 1974). The venous system drains via collecting venules back to the nutrient or emissary veins. This arrangement of vessel from central to peripheral, leads to a higher number of vascular channels and slower flow at the periphery of the marrow cavity. The thin (50–150 μm) trabeculae do not possess a Haversian system or capillary system though do, similar to cortical bone, posses a fine canalicular network linking embedded osteocytes to the bone surface. The cortex receives its blood flow from capillaries piercing its endosteal and periosteal surfaces and running within the Haversian system. As a rough guide, the outer one-third of the cortex is supplied by the periosteal arteries while the inner two-thirds of the cortex and the constituents of the marrow cavity are supplied by the nutrient arteries. Absolute bone blood flow in humans is not easy to measure though has been estimated to be approximately 11 % of cardiac output or 7 ml/min/100 g in humans (Brookes 1974; Van Dyke et al. 1971). More recently, in a study of ten young patients, lower vertebral body blood flow measured by PET and a 15O-labelled CO2 steady-state technique was deemed to be approximately 15 ml/min/100 g bone marrow (Kahn et al. 1994).
3 Links Between the Bone Marrow to Bone Metabolism
There are many tens of ways in which bone marrow properties may affect bone metabolism. From the imaging perspective, these following seem to be the most relevant.
-
(1)
There exists in the bone marrow, pluripotent mesenchymal stem cells that have the potential to differentiate along osteoblastic, adipocytic and haematological cell lines (Gimble and Nuttall 2004). Reduction in estrogen and oxidative stress may cause a drift in mesenchymal stem cell differentiation towards adipocytosis and away from osteoblastogenesis or haematopoesis (D’Ippolito et al. 2006; Fatokun et al. 2006; Shouhed et al. 2005; Kha et al. 2004; Duque 2008; Rosen and Klibanski 2009). Also adipocytes once formed are potentially self promotive whilst simultaneously actively suppressing osteoblastogenesis (Gimble and Nuttall 2004; Duque 2008; Lecka-Czernik et al. 2002).
-
(2)
Bone receives much of its signalling from the bone marrow and the most metabolically active bone areas are those in immediate contact with the bone marrow. The most metabolically active component of bone is the endosteal surface of the cortex with trabeculae bone being the next most metabolically active area (Parfitt 2002). The active unit of bone metabolism, i.e. the basic multicellular unit also lies in close contact with the marrow. Bone metabolism is possibly influenced by changes in the marrow microenvironment. For example, decreased Ph and deceased oxygenation will increase osteoclast formation and activity (Arnett 2010).
-
(3)
Mechano conduction and mechano sensation are terms which embody the principle of bone metabolism being influenced by interstitial fluid flow along osteocytes. Reduced bone blood flow will lead to reduced interstitial fluid flow and reduced shear stresses between osteocytes (Letechipia et al. 2010; Cowin 2002; McCarthy 2005, 2011). These shear stresses stimulate local release of bone remodelling mediators such as NO and PGI2 with the functionally important bone remodelling units and may be related to the rapid loss on bone (and muscle) mass seen microgravity. Osteocytes also produce VEGF which may stimulate bone perfusion.
-
(4)
Good perfusion is a pre-requisite for fracture healing and most bone perfusion comes from the marrow. In contrast, compromised perfusion may aggravate microfracture accumulation which is an integral part of insufficiency fracture development.
-
(5)
Long chain polyunsaturated fatty acids, or a change in the n − 6/n − 3 ratio, can affect bone metabolism. For example, long chain n − 6 fatty acids such as arachidonic acid and its metabolite prostaglandin PGE2 are pro-inflammatory with PGE2 being a potent stimulator of RANKL expression. This can reduce the OPG/RANKL ratio and may increase osteoclastogenesis (Coetzee et al. 2007). Alternatively, long chain n − 3 fatty acids such as eicosapentaenoic acid, docosahexaenoic acid and γ-linolenic have anti-inflammatory activity and may inhibit this PGE2-stimulated increase in RANKL expression (Poulsen et al. 2008). In other words, a change in the fatty acid milieu of bone may affect bone metabolism.
-
(6)
Bone and vasculature metabolism are so closely connected that, at a molecular level, there exists well over a hundred potential mechanisms whereby arteries can interact with bone and vice versa (Demer and Tintut 2009; Hamerman 2005). Broadly speaking, an arterial disorder may be affecting bone; a bone disorder may be affecting arteries or both tissues may be influenced by common extraneous factor or factors. For example, endothelial dysfunction itself has a potent downstream effect on bone metabolism by decreasing local production of nitric oxide and prostaglandin E2 (PGE2) (both of which stimulate osteoblasts and inhibit osteoclasts), decreasing production of PGI2 (which inhibits osteoclasts) and decreasing production of the bone matrix protein osteopontin (Bloomfield et al. 2002). In other words, endothelial dysfunction may, through local mediators, reduce osteoblastic and increase osteoclastic activity (Bloomfield et al. 2002; Wimalawansa 2010).
The remaining part of this chapter will address functional imaging techniques used to assess changes in the bone marrow, will look at what is known about lifelong changes in marrow fat, perfusion and diffusion and will look at how these processes are greatly affected in osteoporosis.
4 Bone Marrow Fat
4.1 Measurement of Marrow Fat
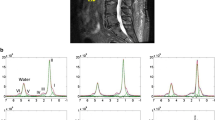
Proton MRS is the most widely used method to quantitatively assess marrow fat. MRS uses the fat: water ratio to determine the fat content (Figs. 4, 5). An obvious limitation is that a constant water content (%) is assumed. In other words, fat: water ratios may change due to a change in water content rather than fat content. MRS requires a minimal volume of approximately 1 cm2 to acquire a sufficient signal to noise ratio. Other non-spectroscopic yet precise methods of quantifying fat fraction are available such as the two-point Dixon method which involves sequential suppression or fat and water, the three-point Iterative Decomposition of water and fat with Echo Asymmetry and Least-squares estimation (IDEAL) (Gerdes et al. 2007), or the analogous Gradient-Echo Sampling of the Free Induction Decay and Echo method (Wehrli et al. 2000). The accuracy of MRI spectroscopic and non-spectroscopic methods in detecting the relative amounts of water and fat has been tested against 11 different emulsions of increasing fat content. This study confirmed a high correlation (r 2 > 0.92) between MR methods of fat quantification and the % fat volume fraction within test bottles (Bernard et al. 2008). Also, reproducibility of proton MRS in a clinical setting is high, ranging from 0.78 to 0.85, with the highest reproducibility being in those areas with the highest inherent fat content, i.e. the femoral head and lowest in the femoral neck (Griffith et al. 2009).

1H MR spectroscopy examination. a Sagittal T2-weighted MR image showing positioning of volume of interest (VOI) for proton spectroscopy of L3 vertebral body. b Coronal oblique T1-weighted image of proximal femur showing VOI’s used to measure fat content (%) in the femoral head, neck, and shaft

Typical 1H MR spectra in a normal subject with little marrow fat and b osteoporotic subject with a large amount of marrow fat
4.2 Physiological Changes in Bone Marrow Fat Content
An inverse relationship between increasing marrow fat and trabecular bone loss in senile osteoporosis has been recognised histologically for 40 years (Dunnill et al. 1967). However, it is only recently, though MRS and other MR-based techniques that marrow fat content can be quantified non-invasively on a large scale (De Bisschop et al. 1993; Schellinger et al. 2000; Kugel et al. 2001; Jung et al. 2000; Wehrli et al. 2000; Shih et al. 2004; Chen and Shih 2006; Liney et al. 2007) and at different anatomical sites (Duda et al. 1995) There is a gradual physiological increase in percentage marrow fat content with advancing years (Kugel et al. 2001; Griffith et al. 2012). An easy approximation to remember is that vertebral body marrow fat content is 25 % at 25 years and 65 % at 65 years of age (Kugel et al. 2001; Griffith et al. 2012).
There is also a distinct sex difference does exist in marrow fat content (Kugel et al. 2001; Griffith et al. 2012). Young males have about 10 % more fat in their marrow than females of equivalent age up to about 50 years of age (Kugel et al. 2001). Males show a gradual steady increase in marrow fat content of 7 % per decade throughout life from young to old (Kugel et al. 2001; Griffith et al. 2012) (Table 1, Fig. 6). Females, in contrast, show a less steep increase in marrow fat of about 2–7 % up to 55 years and then a dramatic increase between the ages of 55 and 65 years (Kugel et al. 2001; Griffith et al. 2012) (Table 1, Fig. 6). By 60 years of age, healthy females tend to possess about 10 % more marrow fat in their vertebrae than males (Griffith et al. 2012) (Table 1, Fig. 6).

The sharp rise in marrow fat content with the menopause may be due to a reduced haematopoietic requirement with cessation of menstruation. This may not, however,be the only cause given that menstrual blood loss is generally quite low (median of about 43 ml per menstrual cycle) (Gao et al. 1987). The sharp increase in marrow fat content in early post-menopausal females may be a more direct effect of estrogen deficiency influencing fat deposition (or stem-cell differentiation) both inside and outside the skeleton. In this respect, the increase in marrow fat content does tally with changes in female extra-skeletal fat distribution recognised to occur at this time.
Androgen and estrogen levels both decline in later years, though estrogen levels fall more sharply in menopausal females leading to a higher circulating androgen: estrogen ratio. This, and other factors, promotes greater intra-abdominal or visceral fat, i.e. an ‘android’ pattern of fat deposition in post-menopausal females (Toth et al. 2000; Blouin et al. 2008). This is different to the gynoid-pattern of fat distribution seen in pre-menopausal women when fat accumulates in the gluteal and thigh areas (Toth et al. 2000; Blouin et al. 2008). Whilst there is no specific literature available on the relationship between estrogen and marrow fat content, it is known that visceral fat content (i.e. an android pattern of fat distribution) does correlate positively with marrow fat content (Bredella et al. 2011). It is feasible, therefore, that the increased marrow fat content seen in females in the post-menopausal era may be the bone-equivalent of android fat deposition. Android fat deposition is also associated with increased risk of cardiovascular disease and metabolic syndrome (Bredella et al. 2011).
Similar findings are found using multivoxel chemical shift registration MR imaging to measure variation in the water fraction of the lumbar vertebral bone marrow with age and sex (Ishijima et al. 1996). The water fraction for males was 75 % for young males, decreased to about 50 % for middle-aged males and remained almost constant for later years (Table 2, Fig. 7) (Ishijima et al. 1996). Conversely, in females, the water fraction for young females remained fairly constant at around 70 % but decreases quite rapidly around the time of menopause such that it is lower than in males during later years (Table 2, Fig. 7) (Ishijima et al. 1996). This tallies with the previously noted lifelong changes in % fat content since fatty marrow contains much less water (~5 %) than red marrow (~35 %) (Hwang and Panicek 2007).

Marrow water content (%) stratified for age and sex (Ishijima 1996)
Overall, there is at least a 40–50 % increase in fat cell content with increasing age. This increase in fat cell volume will occur at the expense of functioning marrow volume. Trabecular volume decreases by about one-third to one-half with increasing age, though the relative percentage of the marrow space occupied by trabecular bone is small. Since, the marrow cavity is a defined space and vascular sinusoids do not seem to expand with age, one can infer that an increase in marrow fat content is really a marker for a decrease in the amount of functioning marrow, i.e. a decrease in red marrow volume.
4.3 Changes in Marrow Fat Content in Osteoporosis
Over and above the physiological increase in marrow fat content with age, osteoporosis is associated with an even greater increase in marrow fat content. In the third lumbar vertebral body, for example, post-menopausal subjects with normal bone mineral density (BMD) have less marrow fat content than subjects with osteopenia. Similarly, subjects with osteopenia have less marrow fat content than this with osteoporosis (De Bisschop et al. 1993; Schellinger et al. 2000; Kugel et al. 2001; Jung et al. 2000; Wehrli et al. 2000; Shih et al. 2004; Chen and Shih 2006; Liney et al. 2007; Griffith et al. 2005, 2006; Shen et al. 2007; Tang et al. 2010; Liu et al. 2010) (Table 3). The proximal femur, which has a higher fat content than the vertebral body, also shows similar changes in increasing marrow fat content as the bone becomes more osteoporotic (Griffith et al. 2008) (Table 3). Even the femoral head, which has a very high intrinsic fat content, also shows an increase in marrow fat content with decreasing BMD though this increase is not as pronounced as in other areas.
4.4 Possible Erroneous Effect of Increasing Marrow Fat
It is possible that the aforementioned findings of increasing marrow fat content with decreasing BMD as measured by dual X-ray absorptiometry (DXA) may be spurious due to the effect of increasing marrow fat on BMD estimation by DXA. Increase in marrow fat content may cause an erroneous reduction in BMD measurements made by DXA (Sorenson 1990; Bolotin 1998; Bolotin et al. 2001; Bolotin 2007). This is because DXA evaluates BMD by measuring the transmission of X-rays at two different photon energies (Blake et al. 2009). The mathematical theory of DXA (basis set decomposition) holds that across a broad range of photon energies, the X-ray transmission factor through any physical object can be decomposed into the equivalent areal densities (g/cm2) of any two designated materials (Blake et al. 2009). For DXA scans, the two materials chosen are bone mineral (hydroxyapatite) and lean tissue. As a result, DXA measurements will only accurately reflect true BMD if the object being examined is composed entirely of hydroxyapatite and lean tissue. In practice, the human body is made up of not two but three main types of tissue, namely bone, lean tissue and fat. Neglecting the difference between lean and fat may lead to a spurious reduction in DXA–BMD measurement. When marrow fat content is known, DXA estimation of BMD needs to be corrected by 0.0014 g/cm2 in women and 0.0016 g/cm2 in men for every 1 % increase in marrow fat about zero (Blake et al. 2009). Applying this correction, the aforementioned results of increasing marrow fat with decreasing BMD still hold true.
4.5 Does Marrow Fat Composition Change with Reducing BMD?
Since changes in marrow fat composition can affect bone metabolism in vivo, and diets rich in polyunsaturated fats can affect BMD, it is conceivable that changes in marrow fat composition can affect bone metabolism (Yeung et al. 2005). To address, this question samples of marrow fat and subcutaneous fat from 126 subjects (98 females, 34 males, mean age 69.7 ± 10.5 years) undergoing orthopaedic surgery were analysed for fatty acid composition using gas chromatography and results correlated with BMD–DXA (Griffith et al. 2009; Yeung et al. 2008) (Fig. 8a, b). A total of 22 fatty acids were identified in marrow and subcutaneous fat. Significant differences existed between marrow and subcutaneous fat fatty acid composition as well as between marrow fat samples obtained from the relatively haematopoietic proximal femur and relatively fatty proximal tibia. Other than cis-7-hexadecenoic acid [C16:1 (n = 9)] and docosanoic acid [C22:0], no difference in marrow fatty acid composition was evident between subject groups of varying BMD (normal, low bone mass and osteoporosis). In particular, the overall polyunsaturated fatty acid content, the n − 6/n − 3 ratio and the percentage composition of those fatty acids most frequently implicated in bone remodelling, namely docosahexaenoic acid, arachidonic acid, γ-linolenic acid and eicosapentaenoic acid, were unchanged in subjects with normal BMD, low bone mass or osteoporosis (Griffith et al. 2009). Overall, it seems less likely that a change in marrow fat composition is directly affecting bone metabolism. The two associations found between fatty acid composition and BMD may be inconsequential given that they account for <1 % (for C16:1(n − 9)) and <0.1 % (for C22:0) of the total marrow fatty acid composition and they do not have any known effect on bone metabolism (Griffith et al. 2009).

a Aspiration of marrow fat from proximal tibia during knee replacement. b Gas chromatography spectrum of marrow fat. Each fatty acid methyl ester is quantified based on peak high relative to internal standard
5 Bone Marrow Perfusion
5.1 Measurement of Bone Marrow Perfusion
Dynamic contrast-enhanced magnetic resonance imaging (DCE-MRI), also known as MR perfusion imaging, measures bone marrow perfusion as opposed to bone marrow blood flow (Griffith and Genant 2011). DCE-MRI is a robust technique that yields empirical indices of perfusion such as maximal signal intensity enhancement (E max) and enhancement slope (E slope) (Figs. 9, 10). E slope and E max are derived from the first-pass phase of signal intensity enhancement and have been shown to be strongly predictive of tissue vascularity, microvessel density and tissue necrosis. In simple terms, E slope can be thought of as gadolinium delivery to the bone marrow and is a feature of blood supply, vascular sinusoidal size and permeability. E max is dependent on these factors though also on the perfusion requirements (i.e. metabolic activity) of the bone marrow. Reproducibility of bone marrow DCE-MRI is moderate to high ranging from 0.59 to 0.98 with best reproducibility in those areas with the highest inherent bone marrow perfusion (Griffith et al. 2009).

Dynamic contrast-enhanced MR imaging a Sagittal T2-weighted MR image showing positioning of region of interest (ROI) for perfusion imaging of L3 vertebral body. b Coronal oblique T1-weighted image of proximal femur showing ROI’s used to measure marrow perfusion in the femoral head, neck and shaft

Time-intensity curve with E max and E slope. E max represents maximum enhancement while E slope represents the slope of the rapidly up-rising part of the curve
Perfusion data acquired from dynamic contrast-enhanced MR imaging is also amenable to two-compartment pharmacokinetic modelling using models such as the Tufts or Brix model (Fig. 11). The Tufts model uses a combination of arterial input function (AIF), and rate constants K trans, K ex and K el. AIF is assessed by analyzing the first pass intensity profile of the feeding artery. K trans refers to the transport constant and is influenced primarily by blood flow. K ex refers to capillary exchange and is influenced by capillary space, permeability, interstitial pressure and extracellular space. K el refers to elimination or wash-out and is influenced by venous return. The Brix model does not rely on AIF or K trans but still considers K ex and K el. It assumes a linear relationship between MR signal enhancement and tissue contrast concentration or, in other words, it assumes that tissue contrast concentration directly correlates with perfusion. No specific pharmacokinetic model to reflect the unique characteristics of marrow perfusion has been developed. Measurement of bone marrow perfusion can also be undertaken by PET-CT imaging undertaken using 18F-fluoride which has a half-life of 112 min. Since this tracer is metabolised in bone, 18F-fluoride imaging is a combined measure of both bone perfusion and bone metabolism as compared to MR perfusion imaging which only measures bone perfusion. Pure bone perfusion can be evaluated by PET using the freely diffusible tracer 15OH20. However, these studies are difficult to perform as 15OH20 has a half-life of only 122 s and thus requires an on-site cyclotron. Nevertheless, a highly significant correlation between blood perfusion measured using 18F-fluoride and true bone perfusion using 15OH20 has been reported (Piert et al. 2002).

Schematic diagram of a Tofts model and b Brix model
5.2 Physiological Age-Related Changes in Bone Marrow Perfusion
Bone marrow perfusion deceases with increasing age (Chen et al. 2001; Montazel et al. 2003; Baur et al. 1997). Subjects aged more than 50 years have a 62 % lower E max (21.88 ± 14.77) that those aged less than 50 years (58.21 ± 44.65, P < 0.005) (Chen et al. 2001). When this is further analysed according to sex, a greater discrepancy is observed. In women, E slope decreased by 80 % (from 87.17 ± 54.13 to 17.98 ± 13.80) in those older than age 50 years (P < 0.005). A similar trend is seen in men with E slope decreases by 33 % from 38.16 ± 21.69 to 25.38 ± 15.43 in subjects more than 50 years though this change did not reach statistical significance (P > 0.05) (Chen et al. 2001). Overall, vertebral bone marrow perfusion is higher in young females than young males (Chen et al. 2001). However, the rate of decrease of perfusion is less in males, which leads to vertebral bone marrow perfusion being higher in elderly males than elderly females (Chen et al. 2001). Similar findings were shown by Montazel JL et al. E max values being significantly higher in patients younger than 40 years than in those aged more than 40 years (P < 0.001). Perfusion parameters decreased with increasing age in a logarithmic relationship (r = 0.71) and correlated with increase in marrow fat content (Montazel et al. 2003). Savvopoulou et al. (2008) showed how the upper (L1, L2) lumbar vertebral bodies were better perfused than the lower (L3, L4, L5) vertebral bodies. In elderly subjects with normal BMD, E max was lower in females (32.3 ± 8.5 %) than males (34.5 ± 13 %) while E slope was higher in females (1.70 ± 5.2 %/s) than males (1.48 ± 0.7 %/s) (Griffith et al. 2005, 2006). To summarise, vertebral marrow perfusion is higher in young females than young males. However, perfusion decreases to a greater degree in females than males. Elderly females have reduced E max but not E slope compared to elderly males.
5.3 Changes in Bone Marrow Perfusion with Osteoporosis
Osteoporosis is associated with a decrease in bone perfusion over and above that accountable for by age alone. Normal BMD subjects have better bone marrow perfusion than osteopenic subjects, while osteopenic subjects have better bone marrow perfusion than osteoporotic subjects (Shih et al. 2004; Griffith et al. 2005, 2006, 2008) (Fig. 12) (Table 4). Similar changes occur in the proximal femur as in the vertebral body (Griffith et al. 2008; Wang et al. 2009). In the proximal femur, reduction in perfusion parameters is most pronounced in the biologically relevant femoral neck than the femoral head or sub-trochanteric regions (Wang et al. 2009).

Typical time-intensity curves for subjects with a normal BMD, b osteopenia and c osteoporosis
5.4 Causes of Marrow Perfusion Changes with Ageing and Osteoporosis
The reduction in marrow perfusion seen with advancing age and osteoporosis is most likely a feature of the marrow per se rather than due to a more generalised vascular or circulatory disturbance since any perfusion changes only occur in the marrow and are not seen in extra-osseous skeletal muscle with the same blood supply (Griffith et al. 2005, 2006, 2008) (Fig. 13).

Schematic representation of vertebral body arterial supply. Each lumbar artery divides to send a nutrient artery to the posterior aspect of the vertebral body and additional arteries to supply the paravertebral muscles. L lumbar artery, QL quadratus lumborum, S spinalis, L longissimus, IL iliocostalis
Overall, the most scenario is that a decrease in the more metabolically active functioning marrow content that is driving the marrow perfusion change seen in ageing and osteoporotic bone (Griffith et al. 2010). Using a combination of MRI and FDG-PET imaging data, the metabolic activity of red marrow as assessed by maximum SUV (standardised uptake value), was estimated to be seven times higher than that of fatty marrow (Basu et al. 2007). Observed changes in marrow fat and trabecular bone fraction with increasing age and osteoporosis essentially mirror a decrease in functioning marrow content. The percentage of the marrow cavity occupied by marrow fat increases from about 25 % in young females to about 70 % in elderly females while over the same period, the percentage occupied by trabecular bone decreases from about 20 to 15 % due to physiological age-related bone loss (Griffith et al. 2012; Müller et al. 1998) (Fig. 6). Within the size restraints of the marrow cavity, one can appreciate that, in line with these changes, the amount of functioning marrow should decrease from 55 to 20 % with ageing (Fig. 6). This reduction in functioning marrow content is manifested clinically as the ‘anaemia of old age’ and an impaired ability of older people to deal with biological stress. Similarly, because functioning marrow also comprises cells of the immune system, this decrease in functioning marrow may also contribute to the ‘immunosenescence’ of old age (Gameiro et al. 2010).
Atherosclerosis and endothelial function may also be related to the compromised bone perfusion seen with ageing and osteoporosis. Increasing age and osteoporosis are strongly associated with arthrosclerosis and, in particular, vascular calcification (Griffith et al. 2012; Nordström et al. 2010; Marcovitz et al. 2005; Collins et al. 2009). Low BMD independently predicts coronary artery disease in women undergoing coronary angiography better than traditional risk factors such as age, hypertension, diabetes, smoking, family history or dyslipidemia (Marcovitz et al. 2005). Histological studies have shown how progressive occlusion of intraosseous arteries, arterioles or arterial capillaries occurs with increasing age (Bridgeman and Brookes 1996) and in patients with proximal femoral osteoporosis (Laroche et al. 1995). Angiographic study has shown how arterioles within the centre of the vertebral body decrease in number, elongate and coil with advancing age (Ratcliffe 1986). A weak negative correlation (r = −0.33, P = 0.0018) was found between vertebral body E max and carotid artery intima-media thickness age after adjusting for the effect of sex, age, blood pressure, BMI, total cholesterol, high density lipoprotein and triglycerol level in a linear regression model (P = 0.008) (Chen et al. 2004). Vertebral body E max was significantly lower in those subjects with carotid intimal thickening group compared to those with normal intimal thickness (73 ± 23 vs. 90 ± 27, P = 0.0023) (Chen et al. 2004).
Endothelial dysfunction is one of the earliest manifestations of atherosclerosis and may be aggravated by sex hormone depletion. Endothelial dysfunction leads to impaired vascular reactivity and seems to effect all arteries, including very likely the nutrient arteries of bones. Impaired endothelial function has been observed in young diabetics and overweight children (Khan et al. 2003). After adjusting for age and years since the menopause, women with low bone mass or osteoporosis had significantly impaired endothelial function that those of normal BMD (Sumino et al. 2007; Sanada et al. 2004; Samuels et al. 2001). Endothelium-dependent vasodilatation is about 20–25 % lower in the femoral nutrient arteries of aged experimental male rats (Prisby et al. 2007). Since blood flow is directly dependent on the vessel radius to the fourth power (Pouiseuille’s Law), a 25 % reduction of vessel diameter due to either arthrosclerosis and/or endothelial dysfunction could potentially reduce volumetric blood flow to the medullary canal by 33 % (Pfitzner 1976).
6 Bone Marrow Diffusion
The free movement of water molecules within the extracellular fluid of the bone marrow is affected by the cells that they encounter. The more closely packed the cells, the more restricted the water motion. Although cell packing is likely to be one of the main modulators, extracellular water motion is also dependent on other factors such as blood flow, capillary permeability, interstitial pressure, temperature and the viscosity of interstitial fluid. Diffusion-weighted MR imaging measures water diffusivity by applying ‘diffusion sensitising gradients’ to T2-weighted spin echo sequences using echoplanar readouts (Khoo et al. 2011). The strength and duration of diffusion sensitising gradients is indicated by their ‘b-value’ with a range of ‘b-values’. The ‘b-values’ applied to clinical diffusion-weighted imaging are such that extracellular and not the intracellular water diffusivity is being measured.
‘Apparent diffusion coefficient’ (ADC) provides a measure of water diffusivity. The ADC of water is 3 × 10−3 mm2/s (Mills 1973). The ADC of fat is close to zero with values of 0.011–0.012 × 10−3 for subcutaneous fat (Lehnert et al. 2004). ADC values of 0.2 × 10−3 and 0.1 × 10−3 have been reported for red and yellow marrow, respectively (Ward et al. 2000).
7 Age-Related Physiological Changes in Bone Marrow Diffusion
Age-related changes in bone marrow diffusion has only been addressed in a few diffusion-weighted studies (Hillengass et al. 2011; Yeung et al. 2004). Bone marrow ADC values in young (mean age 28 years) females were shown to be significantly higher (0.49 ± 0.08 × 10−3 mm2/s) than elderly (mean age 70 years) females (43 ± 0.08 × 10−3 mm2/s, P = 0.029) (Yeung et al. 2004). This is reflective of increased fat packing of bone marrow fat reducing water diffusivity (Nonomura et al. 2001). Against this, in another study of 36 healthy subjects (16 men, 14 women), mean age 56 years, no relationship was found between vertebral marrow ADC and age applying b values of 400 and 750 s/mm2 (Hillengass et al. 2011).
7.1 Diffusion Changes in Osteoporosis
Only a few studies have applied quantitative DWI to the study of bone marrow in patients with and without osteoporosis (Table 4) (Griffith et al. 2006; Liu et al. 2010; Yeung et al. 2004; Hatipoglu et al. 2007). In general, there is a reducing trend in molecular diffusion (as judged by ADC) with diminishing BMD most likely to fat packing of marrow reducing molecular diffusion. One can appreciate that the values obtained from different centres are quite comparable (Table 5).
More studies have applied quantitative DWI to distinguish between fractured osteoporotic and metastatic vertebral bodies (Zhou et al. 2002; Chan et al. 2002; Herneth et al. 2002; Maeda et al. 2003; Balliu et al. 2009; Tang et al. 2007; Biffar et al. 2010) occasionally using adjacent normal appearing vertebra as an internal control (Chan et al. 2002; Herneth et al. 2002; Maeda et al. 2003; Balliu et al. 2009; Tang et al. 2007; Biffar et al. 2010). One can appreciate that the values obtained for normal vertebrae in these studies are different from though reported in non-fractured spines (Table 6).
In general, osteoporotic vertebral fractures tend to have higher ADC values than metastatic vertebral fractures (Table 6). No particular ADC threshold to make this distinction has been adopted (Thawait et al. 2011). There is quite an overlap between reported ADC measurements for benign and metastatic fractures (Table 6). This may relate to selection criteria (such as fracture duration, fracture location, fracture severity, presence of intervertebral clefts, metastatic cell type, sclerotic or non-sclerotic type) or technical factors such as pulse sequencing and b values employed. With respect to differentiation from infectious lesions, reported mean ADC values from infectious vertebral lesions (0.963 ± 0.491 × 10−3 mm2/s) were not statistically different from those obtained from malignant lesions (0.917 ± 0.13 × 10−3 mm2/s) (Balliu et al. 2009).
Tang et al. have reported that the best differentiation of vertebral fracture type can be achieved at b values of around 300 s/mm2 (Tang et al. 2007) while Biffar et al. (2010) report that single shot TSE sequences proved more discriminatory than multi-shot echo planar imaging sequences.
Chemical shift imaging (or opposed phase imaging) has also been used to distinguish between benign and malignant vertebral fractures (Erly et al. 2006; Zampa et al. 2002). If a SIR threshold of 0.80 with >0.8 defined as malignant and <0.8 defined as a benign, in-phase/opposed-phase imaging had a sensitivity of 0.95 and specificity, of 0.89 in discriminating benign from malignant fractures (Erly et al. 2006). Another study has recommended an SIR threshold of 1.2, re-emphasising the variability in choosing the optimal threshold to distinguish benign from metastatic vertebral fracture.
7.2 Predictive Potential of Marrow Parameters
Currently, our ability to select subjects most prone to bone loss is limited, with clinical risk factors such as age, weight, weight loss over 2 years and baseline BMD being the best-recognised markers of future bone loss (Hannan et al. 2000; Lau et al. 2006; Dennison et al. 1999). To address the value of MR marrow parameters in predicting bone loss, a group of subjects with baseline MRS and perfusion imaging of the hip were followed up with hip densitometry at 2 and 4 years. Percentage reduction in femoral neck BMD at 4 years post-baseline was significantly greater in subjects with below median selected marrow or muscle perfusion parameters at baseline compared to those with above median perfusion parameters at baseline (Griffith and Genant 2011) (Fig. 14a–c). Similarly, subjects with more marrow fat at baseline had significantly greater BMD loss over the ensuring 2–4 years (Griffith and Genant 2011) (Fig. 14a–c). However, MR parameters where not sufficiently more predictive of bone loss than traditional risk factors to warrant using MRI to this effect in clinical practice. Nevertheless, the results were sufficiently encouraging to suggest that further refinement of marrow parameters may improve their ability to predict bone loss (Griffith and Genant 2011).

Bone loss over 4 years. Best MR predictors of bone loss in the femoral neck over 4 years when adjusted for covariates, were a muscle E max, b acetabulum E slope and c femoral neck fat content
7.3 Changes in the Extra-osseous Soft Tissues
Although beyond the scope of this chapter, osteoporosis and the menopause is also known to be associated with exaggerated disc degeneration (Wang and Griffith 2010), skeletal muscle loss (Crepaldi et al. 2007) and changes in fat distribution (Bredella et al. 2011; Bredella 2010).
8 Summary
MRI and PET-CT have allowed, the for first time, a quantitative non-invasive assessment of the bone marrow providing us with more information on how the bone marrow changes in health and disease. We can now begin to more fully appreciate physiological age-related changes in the bone marrow that differ between sexes. One can also appreciate that osteoporosis is a disease associated with an exaggeration of physiological age-related changes not just with respect to bone loss but also marrow fat accumulation with a decrease in functioning marrow content and reduced bone perfusion. To fully comprehend the osteoporotic process, we should move away from simply thinking about osteoporosis as a disease of reduced bone density to a more encompassing paradigm which considers bone changes in conjunction with marrow changes, and changes in the extraossoeus soft tissues, particularly muscle. The MR imaging techniques we use to evaluate the bone marrow are still very much a work in progress. Radiologists are in an ideal position to move this bone marrow research forward and help explore in a wider sense connection between systemic diseases, the bone marrow and bone metabolism.
References
Arnett TR (2010) Acidosis, hypoxia and bone. Arch Biochem Biophys 1(503):103–119
Balliu E, Vilanova JC, Peláez I, Puig J, Remollo S, Barceló C, Barceló J, Pedraza S (2009) Diagnostic value of apparent diffusion coefficients to differentiate benign from malignant vertebral bone marrow lesions. Eur J Radiol 69:560–566
Basu S, Houseni M, Bural G, Chamroonat W, Udupa J, Mishra S, Alavi A (2007) Magnetic resonance imaging based bone marrow segmentation for quantitative calculation of pure red marrow metabolism using 2-deoxy-2-[F-18] fluoro-d-glucose-positron emission tomography: a novel application with significant implications for combined structure-function approach. Mol Imaging Biol 9:361–365
Baur A, Stabler A, Bartl R, Lamerz R, Scheidler J, Reiser M (1997) MRI gadolinium enhancement of bone marrow: age-related changes in normals and in diffuse neoplastic infiltration. Skeletal Radiol 26:414–418
Bernard C, Liney G, Manton D, Turnbull L, Langton C (2008) Comparison of fat quantification methods: a phantom study at 3.0 T. J Magn Reson Imaging 27:192–197
Biffar A, Baur-Melnyk A, Schmidt GP, Reiser MF, Dietrich O (2010) Multiparameter MRI assessment of normal-appearing and diseased vertebral bone marrow. Eur Radiol 20:2679–2689
Blake GM, Griffith JF, Yeung DK, Leung PC, Fogelman I (2009) Effect of increasing vertebral marrow fat content on BMD measurement, T-score status and fracture risk prediction by DXA. Bone 44:495–501
Bloomfield SA, Hogan HA, Delp MD (2002) Decreases in bone blood flow and bone material properties in aging Fischer-344 rats. Clin Orthop 396:248–257
Blouin K, Boivin A, Tchernof A (2008) Androgens and body fat distribution. J Steroid Biochem Mol Biol 108:272–280
Bolotin HH (1998) Analytic and quantitative exposition of patient-specific systematic inaccuracies inherent in planar DXA-derived in vivo BMD measurements. Med Phys 25:139–151
Bolotin HH (2007) DXA in vivo BMD methodology: an erroneous and misleading research and clinical gauge of bone mineral status, bone fragility and bone remodelling. Bone 41:138–154
Bolotin HH, Sievansen H, Grashuis JL, Kuiper JW, Jarvinen TL (2001) Inaccuracies inherent in patient-specific dual-energy x-ray absorptiometry bone mineral density measurements: comprehensive phantom-based evaluation. J Bone Miner Res 16:417–426
Bredella MA (2010) Perspective: the bone-fat connection. Skeletal Radiol 39:729–731
Bredella MA, Torriani M, Ghomi RH, Thomas BJ, Brick DJ, Gerweck AV, Rosen CJ, Klibanski A, Miller KK (2011) Vertebral bone marrow fat is positively associated with visceral fat and inversely associated with IGF-1 in obese women. Obesity (Silver Spring) 19:49–53
Bridgeman G, Brookes M (1996) Blood supply to the human femoral diaphysis in youth and senescence. J Anat 188:611–621
Brookes M (1974) Approaches to non-invasive blood flow measurement in bone. Biomed Eng 9:342–347
Chan JH, Peh WC, Tsui EY, Chau LF, Cheung KK, Chan KB, Yuen MK, Wong ET, Wong KP (2002) Acute vertebral body compression fractures: discrimination between benign and malignant causes using apparent diffusion coefficients. Br J Radiol 75:207–214
Chen WT, Shih TT (2006) Correlation between the bone marrow blood perfusion and lipid water content on the lumbar spine in female subjects. J Magn Reson Imaging 24:176–181
Chen WT, Shih TT, Chen RC, Lo SY, Chou CT, Lee JM, Tu HY (2001) Vertebral bone marrow perfusion evaluated with dynamic contrast-enhanced MR imaging: significance of aging and sex. Radiology 220:213–238
Chen WT, Ting-Fang Shih T, Hu CJ, Chen RC, Tu HY (2004) Relationship between vertebral bone marrow blood perfusion and common carotid intima-media thickness in aging adults. J Magn Reson Imaging 20:811–816
Coetzee M, Haag M, Kruger MC (2007) Effects of arachidonic acid, docosahexaenoic acid, prostaglandin E(2) and parathyroid hormone on osteoprotegerin and RANKL secretion by MC3T3-E1 osteoblast-like cells. J Nutr Biochem 18:54–63
Collins TC, Ewing SK, Diem SJ, Taylor BC, Orwoll ES, Cummings SR, Strotmeyer ES, Osteoporotic Fractures in Men (MrOS) Study Group (2009) Peripheral arterial disease is associated with higher rates of hip bone loss and increased fracture risk in older men. Circulation 119:2305–2312
Cowin SC (2002) Mechanosensation and fluid transport in living bone. J Musculoskelet Neuronal Interact 2:256–260
Crepaldi G, Romanato G, Tonin P, Maggi S (2007) Osteoporosis and body composition. J Endocrinol Invest 30:42–47
D’Ippolito G, Diabira S, Howard GA, Roos BA, Schiller PC (2006) Low oxygen tension inhibits osteogenic differentiation and enhances stemness of human MIAMI cells. Bone 39:513–522
De Bisschop E, Luypaert R, Louis O, Osteaux M (1993) Fat fraction of lumbar bone marrow using in vivo proton nuclear magnetic resonance spectroscopy. Bone 14:133–136
Demer LL, Tintut Y (2009) Mechanisms linking osteoporosis with cardiovascular calcification. Curr Osteoporos Rep 7:42–46
Dennison E, Eastell R, Fall CH, Kellingray S, Wood PJ, Cooper C (1999) Determinants of bone loss in elderly men and women: a prospective population-based study. Osteoporos Int 10:384–391
Duda SH, Laniado M, Schick F, Strayle M, Claussen CD (1995) Normal bone marrow in the sacrum of young adults: differences between the sexes seen on chemical-shift MR imaging. AJR Am J Roentgenol 164:935–940
Dunnill MS, Anderson JA, Whitehead R (1967) Quantitative histological studies on age changes in bone. J Pathol Bacteriol 94:275–291
Duque G (2008) Bone and fat connection in aging bone. Curr Opin Rheumatol 20:429–434
Erly WK, Oh ES, Outwater EK (2006) The utility of in-phase/opposed-phase imaging in differentiating malignancy from acute benign compression fractures of the spine. AJNR Am J Neuroradiol 27:1183–1188
Fatokun AA, Stone TW, Smith RA (2006) Hydrogen peroxide-induced oxidative stress in MC3T3-E1 cells: the effects of glutamate and protection by purines. Bone 39:542–551
Gameiro CM, Romão F, Castelo-Branco C (2010) Menopause and aging: changes in the immune system—a review. Maturitas 67:316–320
Gao J, Zeng S, Sun BL, Fan HM, Han LH (1987) Menstrual blood loss and hematologic indices in healthy Chinese women. J Reprod Med 32:822–826
Gerdes CM, Kijowski R, Reeder SB (2007) IDEAL imaging of the musculoskeletal system: robust water fat separation for uniform fat suppression, marrow evaluation, and cartilage imaging. AJR Am J Roentgenol 189:W284–W291
Gimble JM, Nuttall ME (2004) Bone and fat: old questions, new insights. Endocrine 23:183–188
Griffith JF, Genant HK (2011) New imaging modalities in bone. Curr Rheumatol Rep 13:241–250
Griffith JF, Yeung DK, Antonio GE, Lee FK, Hong AW, Wong SY, Lau EM, Leung PC (2005) Vertebral bone mineral density, marrow perfusion, and fat content in healthy men and men with osteoporosis: dynamic contrast-enhanced MR imaging and MR spectroscopy. Radiology 236:945–951
Griffith JF, Yeung DK, Antonio GE, Wong SY, Kwok TC, Woo J, Leung PC (2006) Vertebral marrow fat content and diffusion and perfusion indexes in women with varying bone density: MR evaluation. Radiology 241:831–838
Griffith JF, Yeung DK, Tsang PH, Choi KC, Kwok TC, Ahuja AT, Leung KS, Leung PC (2008) Compromised bone marrow perfusion in osteoporosis. J Bone Miner Res 23:1068–1075
Griffith JF, Yeung DK, Chow SK, Leung JC, Leung PC (2009) Reproducibility of MR perfusion and (1)H spectroscopy of bone marrow. J Magn Reson Imaging 29:1438–1442
Griffith JF, Engelke K, Genant HK (2010) Looking beyond bone mineral density: Imaging assessment of bone quality. Ann N Y Acad Sci 1192:45–56
Griffith JF, Yeung DKW, Ma HT, Leung JSC, Kwok TCY, Leung PC (2012) Bone marrow fat content in the elderly: a reversal of trend seen in younger subjects J Magn Reson Imaging (In press)
Hamerman D (2005) Osteoporosis and atherosclerosis: biological linkages and the emergence of dual-purpose therapies. QJM 98:467–484
Hannan MT, Felson DT, Dawson-Hughes B, Tucker KL, Cupples LA, Wilson PW, Kiel DP (2000) Risk factors for longitudinal bone loss in elderly men and women: the Framingham Osteoporosis Study. J Bone Miner Res 15:710–720
Hartsock RJ, Smith EB, Petty CS (1965) Normal variations with aging of the amount of hematopoetic tissue in bone marrow from the anterior iliac crest. A study made from 177 cases of sudden death examined by necropsy. Am J Clin Pathol 43:326–331
Hatipoglu HG, Selvi A, Ciliz D, Yuksel E (2007) Quantitative and diffusion MR imaging as a new method to assess osteoporosis. Am J Neuroradiol 28:1934–1937
Herneth AM, Philipp MO, Naude J, Funovics M, Beichel RR, Bammer R, Imhof H (2002) Vertebral metastases: assessment with apparent diffusion coefficient. Radiology 225:889–894
Hillengass J, Stieltjes B, Bäuerle T, McClanahan F, Heiss C, Hielscher T, Wagner-Gund B, Habetler V, Goldschmidt H, Schlemmer HP, Delorme S, Zechmann CM (2011) Dynamic contrast-enhanced magnetic resonance imaging (DCE-MRI) and diffusion-weighted imaging of bone marrow in healthy individuals. Acta Radiol 1(52):324–330
Hwang S, Panicek DM (2007) Magnetic resonance imaging of bone marrow in oncology, Part 1. Skeletal Radiol 36:913–920
Ishijima H, Ishizaka H, Horikoshi H, Sakurai M (1996) Water fraction of lumbar vertebral bone marrow estimated from chemical shift misregistration on MR imaging: normal variations with age and sex. AJR Am J Roentgenol 167:355–358
Jung CM, Kugel H, Schulte O, Heindel W (2000) Proton-MR spectroscopy of the spinal bone marrow. An analysis of physiological signal behavior. Radiologe 40:694–699
Kahn D, Weiner GJ, Ben-Haim S, Ponto LL, Madsen MT, Bushnell DL, Watkins GL, Argenyi EA, Hichwa RD (1994) Positron emission tomographic measurement of bone marrow blood flow to the pelvis and lumbar vertebrae in young normal adults. Blood 15(83):958–963
Kha HT, Basseri B, Shouhed D, Richardson J, Tetradis S, Hahn TJ, Parhami F (2004) Oxysterols regulate differentiation of mesenchymal stem cells: pro-bone and anti-fat. J Bone Miner Res 19:830–840
Khan F, Green FC, Forsyth JS, Greene SA, Morris AD, Belch JJ (2003) Impaired microvascular function in normal children: effects of adiposity and poor glucose handling. J Physiol 551:705–711
Khoo MM, Tyler PA, Saifuddin A, Padhani AR (2011) Diffusion-weighted imaging (DWI) in musculoskeletal MRI: a critical review. Skeletal Radiol 40:665–681
Kugel H, Jung C, Schulte O, Heindel W (2001) Age- and sex-specific differences in the 1H-spectrum of vertebral bone marrow. J Magn Reson Imaging 13:263–268
Laroche M, Ludot I, Thiechart M, Arlet J, Pieraggi M, Chiron P, Moulinier L, Cantagrel A, Puget J, Utheza G et al (1995) Study of the intraosseous vessels of the femoral head in patients with fractures of the femoral neck or osteoarthritis of the hip. Osteoporos Int 5:213–217
Lau EM, Leung PC, Kwok T, Woo J, Lynn H, Orwoll E, Cummings S, Cauley J (2006) The determinants of bone mineral density in Chinese men—results from Mr. Os (Hong Kong), the first cohort study on osteoporosis in Asian men. Osteoporos Int 17:297–303
Lecka-Czernik B, Moerman EJ, Grant DF, Lehmann JM, Manolagas SC, Jilka RL (2002) Divergent effects of selective peroxisome proliferator-activated receptor-gamma 2 ligands on adipocyte versus osteoblast differentiation. Endocrinology 143:2376–2384
Lehnert A, Machann J, Helms G, Claussen CD, Schick F (2004) Diffusion characteristics of large molecules assessed by proton MRS on a whole-body MR system. Magn Reson Imaging 22:39–46
Letechipia JE, Alessi A, Rodriguez G, Asbun J (2010) Would increased interstitial fluid flow through in situ mechanical stimulation enhance bone remodeling? Med Hypotheses 75:196–198
Lichtman MA (1981) The ultrastructure of the hemopoietic environment of the marrow: a review. Exp Hematol 9:391–410
Liney GP, Bernard CP, Manton DJ, Turnbull LW, Langton CM (2007) Age, gender, and skeletal variation in bone marrow composition: a preliminary study at 3.0 Tesla. J Magn Reson Imaging 26:787–793
Link TM (2012) Osteoporosis imaging: state of the art and advanced imaging. Radiology 263:3–17
Liu Y, Tang GY, Tang RB, Peng YF, Li W (2010) Assessment of bone marrow changes in postmenopausal women with varying bone densities: magnetic resonance spectroscopy and diffusion magnetic resonance imaging. Chin Med J (Engl) 123:1524–1527
Maeda M, Sakuma H, Maier SE, Takeda K (2003) Quantitative assessment of diffusion abnormalities in benign and malignant vertebral compression fractures by line scan diffusion-weighted imaging. AJR Am J Roentgenol 181:1203–1209
Manolagas SC, Almeida M (2007) Gone with the Wnts: beta-catenin, T-cell factor, forkhead box O, and oxidative stress in age-dependent diseases of bone, lipid, and glucose metabolism. Mol Endocrinol 21:2605–2614
Marcovitz PA, Tran HH, Franklin BA, O’Neill WW, Yerkey M, Boura J, Kleerekoper M, Dickinson CZ (2005) Usefulness of bone mineral density to predict significant coronary artery disease. Am J Cardiol 96:1059–1063
McCarthy ID (2005) Fluid shifts due to microgravity and their effects on bone: a review of current knowledge. Ann Biomed Eng 33:95–103
McCarthy EF (2011) Perspective: skeletal complications of space flight. Skeletal Radiol 40:661–663
Mills R (1973) Self-diffusion in normal and heavy water in the range 1–45 deg. J Phy Chem 77:685–688
Montazel JL, Divine M, Lepage E, Kobeiter H, Breil S, Rahmouni A (2003) Normal spinal bone marrow in adults: dynamic gadolinium-enhanced MR imaging. Radiology 229:703–709
Müller R, Van Campenhout H, Van Damme B, Van Der Perre G, Dequeker J, Hildebrand T, Rüegsegger P (1998) Morphometric analysis of human bone biopsies: a quantitative structural comparison of histological sections and micro-computed tomography. Bone 23:59–66
Nonomura Y, Yasumoto M, Yoshimura R, Haraguchi K, Ito S, Akashi T, Ohashi I (2001) Relationship between bone marrow cellularity and apparent diffusion coefficient. J Magn Reson Imaging 13:757–760
Nordström A, Eriksson M, Stegmayr B, Gustafson Y, Nordström P (2010) Low bone mineral density is an independent risk factor for stroke and death. Cerebrovasc Dis. 29:130–136
Parfitt AM (2002) Misconceptions (2): turnover is always higher in cancellous than in cortical bone. Bone 30:807–809
Pfitzner J (1976) Poiseuille and his law. Anaesthesia 31:273–275
Piert M, Machulla HJ, Jahn M, Stahlschmidt A, Becker GA, Zittel TT (2002) Coupling of porcine bone blood flow and metabolism in high-turnover bone disease measured by [(15)O]H(2)O and [(18)F]fluoride ion positron emission tomography. Eur J Nucl Med Mol Imaging 29:907–914
Poulsen RC, Wolber FM, Moughan PJ, Kruger MC (2008) Long chain polyunsaturated fatty acids alter membrane-bound RANK-L expression and osteoprotegerin secretion by MC3T3-E1 osteoblast-like cells. Prostaglandins Other Lipid Mediat 85:42–48
Poulton TB, Murphy WD, Duerk JL, Chapek CC, Feiglin DH (1993) Bone marrow reconversion in adults who are smokers: MR Imaging findings. Am J Roentgenol 161:1217–1221
Prisby RD, Ramsey MW, Behnke BJ, Dominguez JM 2nd, Donato AJ, Allen MR, Delp MD (2007) Aging reduces skeletal blood flow, endothelium-dependent vasodilation, and NO bioavailability in rats. J Bone Miner Res 22:1280–1288
Ratcliffe JF (1986) Arterial changes in the human vertebral body associated with aging. The ratios of peripheral to central arteries and arterial coiling. Spine 11:235–240
Rosen CJ, Klibanski A (2009) Bone, fat, and body composition: evolving concepts in the pathogenesis of osteoporosis. Am J Med 122:409–414
Samuels A, Perry MJ, Gibson RL, Colley S, Tobias JH (2001) Role of endothelial nitric oxide synthase in estrogen-induced osteogenesis. Bone 29:24–29
Sanada M, Taguchi A, Higashi Y, Tsuda M, Kodama I, Yoshizumi M, Ohama K (2004) Forearm endothelial function and bone mineral loss in postmenopausal women. Atherosclerosis 176:387–392
Savvopoulou V, Maris TG, Vlahos L, Moulopoulos LA (2008) Differences in perfusion parameters between upper and lower lumbar vertebral segments with dynamic contrast-enhanced MRI (DCE MRI). Eur Radiol 18:1876–1883
Schellinger D, Lin SC, Fertikh D et al (2000) Normal lumbar vertebrae: anatomic, age, and sex variance in subjects at proton MR spectroscopy-initial experience. Radiology 215:910–916
Shen W, Chen J, Punyanitya M, Shapses S, Heshka S, Heymsfield SB (2007) MRI-measured bone marrow adipose tissue is inversely related to DXA-measured bone mineral in Caucasian women. Osteoporos Int 18:641–647
Shih TT, Chang CJ, Hsu CY, Wei SY, Su KC, Chung HW (2004) Correlation of bone marrow lipid water content with bone mineral density on the lumbar spine. Spine (Phila Pa 1976) 15(29):2844–2850
Shouhed D, Kha HT, Richardson JA, Amantea CM, Hahn TJ, Parhami F (2005) Osteogenic oxysterols inhibit the adverse effects of oxidative stress on osteogenic differentiation of marrow stromal cells. J Cell Biochem 95:1276–1283
Sorenson JA (1990) Effects of nonmineral tissues on measurement of bone mineral content by dual-photon absorptiometry. Med Phys 17:905–912
Steiner RM, Mitchell DG, Rao VM, Schweitzer ME (1993) Magnetic resonance imaging of diffuse bone marrow disease. Radiol Clin North Am 31:383–409
Sumino H, Ichikawa S, Kasama S, Takahashi T, Sakamoto H, Kumakura H, Takayama Y, Kanda T, Murakami M, Kurabayashi M (2007) Relationship between brachial arterial endothelial function and lumbar spine bone mineral density in postmenopausal women. Circ J 71:1555–1559
Tang G, Liu Y, Li W, Yao J, Li B, Li P (2007) Optimization of b value in diffusion-weighted MRI for the differential diagnosis of benign and malignant vertebral fractures. Skeletal Radiol 36:1035–1041
Tang GY, Lv ZW, Tang RB, Liu Y, Peng YF, Li W, Cheng YS (2010) Evaluation of MR spectroscopy and diffusion-weighted MRI in detecting bone marrow changes in postmenopausal women with osteoporosis. Clin Radiol 65:377–381
Thawait SK, Marcus MA, Morrison WB, Klufas RA, Eng J, Carrino J (2011) Research synthesis: what is the diagnostic performance of MRI to discriminate benign from malignant vertebral compression fractures? Systematic review and meta-analysis. Spine (Phila Pa 1976) [Epub ahead of print]
Toth MJ, Tchernof A, Sites CK, Poehlman ET (2000) Menopause-related changes in body fat distribution. Ann N Y Acad Sci 904:502–506
Travlos GS (2006) Normal structure, function, and histology of the bone marrow. Toxicol Pathol 34:548–565
Van Dyke D, Parker H, Anger HO, McRae J, Dobson EL, Yano Y, Naets JP, Linfoot J (1971) Markedly increased bone blood flow in myelofibrosis. J Nucl Med 12:506–512
Wang YX, Griffith JF (2010) Effect of menopause on lumbar disk degeneration: potential etiology. Radiology 257:318–320
Wang YX, Griffith JF, Kwok AW, Leung JC, Yeung DK, Ahuja AT, Leung PC (2009) Reduced bone perfusion in proximal femur of subjects with decreased bone mineral density preferentially affects the femoral neck. Bone 45:711–715
Ward R, Caruthers S, Yablon C, Blake M, DiMasi M, Eustace S (2000) Analysis of diffusion changes in posttraumatic bone marrow using navigator-corrected diffusion gradients. Am J Roentgenol 174:731–734
Wehrli FW, Hopkins JA, Hwang SN, Song HK, Snyder PJ, Haddad JG (2000) Cross-sectional study of osteopenia with quantitative MR imaging and bone densitometry. Radiology 217:527–538
Wimalawansa SJ (2010) Nitric oxide and bone. Ann N Y Acad Sci 1192:391–403
Yeung DK, Wong SY, Griffith JF, Lau EM (2004) Bone marrow diffusion in osteoporosis: evaluation with quantitative MR diffusion imaging. J Magn Reson Imaging 19:222
Yeung DK, Griffith JF, Antonio GE, Lee FK, Woo J, Leung PC (2005) Osteoporosis is associated with increased marrow fat content and decreased marrow fat unsaturation: a proton MR spectroscopy study. J Magn Reson Imaging 22:279–285
Yeung DK, Lam SL, Griffith JF, Chan AB, Chen Z, Tsang PH, Leung PC (2008) Analysis of bone marrow fatty acid composition using high-resolution proton NMR spectroscopy. Chem Phys Lipids 151:103–109
Zampa V, Cosottini M, Michelassi C, Ortori S, Bruschini L, Bartolozzi C (2002) Value of opposed-phase gradient-echo technique in distinguishing between benign and malignant vertebral lesions. Eur Radiol 12:1811–1818
Zhou XJ, Leeds NE, McKinnon GC, Kumar AJ (2002) Characterization of benign and metastatic vertebral compression fractures with quantitative diffusion MR imaging. Am J Neuroradiol 23:165–170
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2013 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Griffith, J.F. (2013). Bone Marrow Changes in Osteoporosis. In: Guglielmi, G. (eds) Osteoporosis and Bone Densitometry Measurements. Medical Radiology(). Springer, Berlin, Heidelberg. https://doi.org/10.1007/174_2012_614
Download citation
DOI: https://doi.org/10.1007/174_2012_614
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-27883-9
Online ISBN: 978-3-642-27884-6
eBook Packages: MedicineMedicine (R0)