
Abstract
The purpose of the present study was to determine how long-lasting the post-conflict slowing following incongruent stimuli is. In previous research, incongruent stimuli have been used to induce a conflict because they have relevant features for two different response alternatives. So far, the post-conflict slowing following incongruent stimuli has mainly been assessed up to one trial. In the first two experiments, we assessed the persistence of the post-conflict slowing across several trials. To this end, we presented a few incongruent stimuli among non-conflict stimuli. The results showed a consistent slowing for the first few trials immediately following the incongruent trials. In addition, a sporadic slowing was still found on later trials. In two subsequent experiments, we investigated to what extent the infrequency of incongruent trials — rather than their conflict — induced this slowing. To determine this, we used the same design as in the first two experiments, but we presented non-conflict stimuli as infrequent stimuli. The results showed a slowing on one subsequent trial, ruling out the possibility that the post-conflict slowing following incongruent trials was only caused by infrequency. Together, the findings of the present study indicate that the conflict induced by incongruent trials can have a longer lasting impact on subsequent trials than previously thought.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
A current issue in cognitive psychology concerns the cognitive control processes following a conflict. When facing a conflict, cognitive control allows us to select goal-relevant features and inhibiting irrelevant features. Thus, responding to a conflict slows performance. Interestingly, performance is also slowed on subsequent (non-conflict) stimuli (e.g., Botvinick, Braver, Barch, Carter, & Cohen, 2001; Duthoo, Abrahamse, Braem, Boehler, & Notebaert, 2014; Loft, Kearney, & Remington, 2008; Verguts, Notebaert, Kunde, & Wühr, 2011; Woodward, Meier, Tipper, & Graf, 2003). The purpose of the present study was to determine how long-lasting this post-conflict slowing is.
Different kinds of post-conflict slowing
So far, the post-conflict slowing has been investigated when the conflict occurs in the task-switching paradigm, the prospective memory paradigm as well as in the Stroop, Simon, and flanker tasks (see Table 1 for a description of these paradigms and their trials). In the task-switching paradigm, the post-conflict slowing has been called bivalency effect (Grundy et al., 2013; Meier, Rey-Mermet, & Rothen, 2015; Meier, Rey-Mermet, Woodward, Müri, & Gutbrod, 2013; Meier, Woodward, Rey-Mermet, & Graf, 2009; Rey-Mermet, Koenig, & Meier, 2013; Rey-Mermet & Meier, 2012a, 2012b, 2013, 2014, 2015; Woodward, Metzak, Meier, & Holroyd, 2008). In the paradigm typically used to investigate this effect, participants are asked to switch during three blocks between three tasks, such as a parity decision (odd vs. even), a color decision (red vs. blue), and a case decision (uppercase vs. lowercase). In the first and third blocks (the pure blocks), all stimuli are univalent (i.e., black numerals for the parity decision, colored symbols for the color decision, and black letters for the case decision). In the second block (the mixed block), occasionally the letters for the case decisions are printed in red or blue color, which turns them into bivalent stimuli. The bivalency effect is the performance slowing that occurs on all univalent trials following bivalent stimuli, including those sharing no relevant features with bivalent stimuli (i.e., the parity-decision trials). Critically, the bivalency effect has been found to persist across many subsequent trials (Meier et al., 2015; Meier et al., 2013; Meier et al., 2009; Rey-Mermet & Meier, 2013, 2015). Moreover, increasing the interval from 1,000 ms to 5,000 ms after each task triplet does not affect its magnitude (Meier et al., 2009). Thus, the bivalency effect affects subsequent trials for more than 20 seconds after the occurrence of the conflict (see Meier & Rey-Mermet, 2012a, for a review).
In the prospective memory paradigm (see Table 1), the conflict is triggered by prospective memory targets (Loft et al., 2008; Meier & Rey-Mermet, 2017, Meier & Rey-Mermet, 2012b). Loft et al. (2008) first revealed the presence of a slowing induced by prospective memory targets. That is, after participants were instructed to perform a prospective memory task, performance was slower on ongoing trials when prospective memory targets were presented compared to when they were not presented. In our study (Meier & Rey-Mermet, 2012b), we specifically investigated the persistence of this slowing. Thus, we used the typical bivalency effect paradigm but we asked participants to press another key (i.e., the key h instead of b or n) when they encountered the targets (i.e., the red or blue letters). The results showed a performance slowing for the first three trials that immediately followed these targets. This slowing lasted up to 6 seconds. Interestingly, we found that ongoing task performance was also slowed on subsequent trials but only for those trials sharing relevant features with the prospective memory targets (Meier & Rey-Mermet, 2017, Meier & Rey-Mermet, 2012b).
In the Stroop, Simon, and flanker tasks (see Table 1), the conflict is triggered by incongruent trials (Eriksen & Eriksen, 1974; MacLeod, 1991; Simon & Small, 1969; Stroop, 1935). In all three tasks, responding to incongruent trials results in slower and more error-prone performance than responding to congruent trials (Eriksen & Eriksen, 1974; MacLeod, 1991; Simon & Small, 1969; Stroop, 1935). Interestingly, this congruency effect is reduced when incongruent trials are presented more frequently than congruent trials, which results in a proportion congruency effect (Gratton, Coles, & Donchin, 1992; Hommel, 1994; Logan & Zbrodoff, 1979; Lowe & Mitterer, 1982). Moreover, the congruency effect is also reduced after incongruent trials compared to after congruent trials (Duthoo, Abrahamse, Braem, Boehler, & Notebaert, 2014; Egner, 2007). This congruency sequence effect refers to the combination of two effects: a performance acceleration observed on incongruent trials following incongruent trials and a performance slowing observed on congruent trials following incongruent trials. Thus, the congruency sequence effect demonstrates that the conflict induced by incongruent stimuli on Trial T has an impact on the immediate subsequent performance (i.e., on T+1). Only a few studies have explored the impact on subsequent trials, specifically, on T+2 trials (Akçay & Hazeltine, 2008; Horga et al., 2011; Mayr, Awh, & Laurey, 2003; Stürmer, Leuthold, Soetens, Schröter, & Sommer, 2002; Wendt, Kluwe, & Peters, 2006). The main goal of these studies was to determine whether an incongruent trial could reduce the congruency effect on trial T+2. The results were mixed. Some studies found no impact on T+2 (see Stürmer et al., 2002, Experiment 3; Wendt et al., 2006, Experiments 1 and 2a), while others did (Akçay & Hazeltine, 2008, Experiment 2; Mayr et al., 2003, Experiment 2; Wendt et al., 2006, Experiment 3). However, these studies focused on the performance acceleration on incongruent trials following incongruent trials.
Only two studies have focused on the post-conflict slowing following incongruent trials (Rey-Mermet & Meier, 2016; Verguts et al., 2011).Footnote 1 The first study focused on the immediate subsequent trial (Verguts et al., 2011) and showed a post-conflict slowing on this trial only if it had one common feature with the incongruent trials (i.e., in our terminology, if this trial was neutral). If this subsequent trial had several features in common with the incongruent trials (e.g., if it was congruent or incongruent), no post-conflict slowing was found. This was explained by assuming that a post-focusing process (i.e., an increased attention to the relevant response feature) masks the post-conflict slowing. In our study (Rey-Mermet & Meier, 2016), the post-conflict slowing was assessed across several trials. The goal of that study was to determine whether this slowing could generalize to trials sharing no relevant features with the conflict (i.e., univalent trials). To this end, we asked participants to switch between a task including occasionally the incongruent trials (e.g., the Stroop, Simon, or flanker task) and a task sharing no relevant features with the conflict (i.e., a digit classification with univalent trials). The results revealed an initial performance slowing that affected both tasks after incongruent trials. This slowing affected 12 trials. On further trials, however, the slowing mainly affected the task sharing features with the conflict stimuli.
Different explanations underlying the post-conflict slowing
To explain the different kinds of post-conflict slowing, different explanations have been put forward. For example, we accounted for the post-conflict slowing following bivalent and incongruent trials by proposing an episodic context binding explanation (Meier & Rey-Mermet, 2012a; Meier et al., 2013; Meier et al., 2009; Rey-Mermet & Meier, 2015, 2016). According to this account, responding to a particular trial results in a memory representation that is bound to the proximate context (e.g., the particular task sequence of parity, color, and case decisions in the case of bivalent stimuli). This context is retrieved and updated each time a task is performed. When a conflict stimulus occurs within a task sequence, the whole context becomes conflict-loaded, and thus, on subsequent trials, the retrieval of this representation causes interference. As the representation included the whole task sequence, performance is generally slowed for several subsequent trials, which results in a long-lasting and task-unspecific post-conflict slowing.
In contrast, the post-conflict slowing following prospective memory targets have been explained with two further accounts. That is, the performance slowing occurring on the first three trials immediately following the targets was interpreted as an orienting response effect (Meier & Rey-Mermet, 2012b). According to this account (cf. Notebaert et al., 2009; Notebaert & Verguts, 2011; Núñez Castellar, Kühn, Fias, & Notebaert, 2010), infrequent stimuli capture attention, and it takes some time to redirect attention to the task to be performed, which slows performance on subsequent trials. As prospective memory targets are infrequent events, this means that they capture attention and redirect it away from the ongoing task. Thus, when the ongoing task has to be performed again, it takes some time to redirect attention to it, which results in performance slowing for the first few trials following the prospective memory targets.
The later post-conflict slowing occurring only on those trials sharing features with the prospective memory targets was interpreted as the result of the prospective memory response (Meier & Rey-Mermet, 2012b). More precisely, as prospective memory targets require a different response than the ongoing tasks (e.g., the key h), responding to the prospective memory task strengthens the association between the (unusual) prospective memory response and the ongoing tasks with overlapping features with the targets. On subsequent ongoing task trials, the reactivation of the prospective memory response may interfere with the activation of the ongoing task response, which results in a performance slowing for the tasks with overlapping features (see Metzker & Dreisbach, 2009). This later post-conflict slowing cannot, however, be accounted by an expectancy-based monitoring explanation. According to such an account (e.g., Meier, Zimmermann, & Perrig, 2006; Smith, 2003), monitoring for further prospective memory targets would steadily increase across trials, and this monitoring process would result in an increase of slowing across trials. However, the results showed a decline of the post-conflict slowing across trials (see also Meier & Rey-Mermet, 2017).
Different or same kind(s) of post-conflict slowing?
This overview reveals that the post-conflict slowing can persist across several trials, and its trajectory and the underlying processes differ only if the conflict is induced by prospective memory targets. At first sight, the post-conflict slowing following bivalent and incongruent trials does not seem to differ. However, this hypothesis might be questioned by at least two arguments. First, in the few studies in which the bivalency effect was compared after incongruent and congruent bivalent stimuli (as bivalent stimuli can be either incongruent or congruent; see Table 1), the results were inconclusive (see Grundy & Shedden, 2014a; Rey-Mermet & Meier, 2014, for behavioral studies; and see Grundy & Shedden, 2014b, for an EEG study). Grundy and colleagues found a larger post-conflict slowing after incongruent bivalent stimuli than after congruent bivalent stimuli, whereas we found no difference. This discrepancy might stem from the fact that Grundy and colleagues did not inform participants about the occurrence of bivalent stimuli (see Grundy & Shedden, 2014b, for an exception), while we did. Furthermore, Grundy and colleagues presented bivalent stimuli randomly, while we presented them regularly. Therefore, it is possible that the design used by Grundy and colleagues results in some uncertainty about which task to perform when incongruent bivalent trials were encountered (see Kray & Lindenberger, 2000; Metzak, Meier, Graf, & Woodward, 2013), and this uncertainty would persist across several trials, thus resulting in a larger post-conflict slowing after incongruent bivalent trials. In contrast, in our study, such task uncertainly was highly improbable because bivalent stimuli occurred regularly, and participants were instructed which task to perform on bivalent stimuli. In any case, it remains unclear whether the post-conflict slowing following bivalent (congruent) stimuli differs from the post-conflict slowing following (bivalent) incongruent trials.
A second reason to question the hypothesis of similar post-conflict slowing after bivalent and incongruent trials is that in all studies investigating the post-conflict slowing following incongruent trials (Grundy & Shedden, 2014a, 2014b; Rey-Mermet & Meier, 2016; Verguts et al., 2011), participants switched between at least two tasks. Although a task-switching paradigm is necessary to create bivalent stimuli, it is not the case for incongruent trials. Thus, using a task-switching design for both bivalent and incongruent trials might have promoted the similarities between both kinds of post-conflict slowing. Moreover, the results of Verguts et al. (2011) emphasize the importance of considering under which conditions the post-conflict slowing is investigated. The purpose of the present study was thus to investigate the trajectory of the post-conflict slowing following incongruent trials in a more “natural” paradigm for incongruent trials, that is, when no task switching is required. This is important because it might inform us about the unity or diversity of cognitive control processes. Thus, if no task-switching design is used but the post-conflict slowing following incongruent trials is as long-lasting as the post-conflict slowing following bivalent stimuli, this would suggest similar cognitive processes underlying both kinds of post-conflict slowing (e.g., episodic context binding). In this case, this would challenge research in which different sources of conflict were found to induce different kinds of cognitive control processes (see Braem, Abrahamse, Duthoo, & Notebaert, 2014; Egner, 2008, for reviews).
In the present study, we conducted four experiments (see Table 2, left part, for an overview of the manipulations). In Experiments 1 and 2, we investigated how long-lasting the post-conflict slowing following incongruent trials is. To this end, we occasionally presented incongruent trials among non-conflict stimuli (i.e., congruent trials in Experiment 1 and neutral trials in Experiment 2). In Experiments 3 and 4, we determined whether the slowing following incongruent trials results from their infrequency rather than their conflict. To this end, we examined to what extent infrequent (non-conflict) stimuli result in a slowing on subsequent trials. In Experiment 3, we thus reversed the ratio of incongruent and congruent trials used in Experiment 1 by occasionally presenting congruent trials among incongruent trials. In Experiment 4, we occasionally presented neutral trials among congruent trials. In each experiment, we investigated the persistence of the slowing following conflict or infrequent stimuli by determining the trajectory of the performance slowing across the subsequent trials that immediately followed.
Experiment 1
The goal of Experiment 1 was to determine how long-lasting the post-conflict slowing following incongruent trials is. To this end, we asked our participants to perform either a Stroop, a Simon, or a flanker task during three blocks. In the first and third blocks, only congruent stimuli were presented; in the second block, incongruent stimuli appeared occasionally. In addition, we manipulated the interval between sequences of four trials so that the interval was 1,000 ms for half of the participants and 2,000 ms for the other half. Previous research has revealed that the post-conflict slowing following bivalent stimuli persists across time (see Meier et al., 2009; Rey-Mermet & Meier, 2013). However, it might not be the case for the post-conflict slowing following incongruent trials as the congruency sequence effect diminished across time (Duthoo, Abrahamse, Braem, & Notebaert, 2014; Egner, Ely, & Grinband, 2010). Thus, increasing the interval from 1,000 ms to 2,000 ms after incongruent trials allowed us to examine whether the post-conflict slowing following incongruent trials persists across time in addition to across trials.
In the present experiment, we thus manipulated two variables within-subject (block and trial) and two variables (task and interval) between-subjects.Footnote 2 The variable block takes into account the three blocks (Block 1, Block 2, and Block 3). The variable trial takes into account the number of trials following an incongruent stimulus (i.e., T+1, T+2, etc., with T referring to the trial containing an incongruent stimulus). The variable task takes into account the three different tasks (i.e., Stroop, Simon, and flanker). The variable interval takes into account the two different intervals between the sequences of four trials (i.e., 1,000 ms or 2,000 ms).
We hypothesized that if the task-switching paradigm is not relevant for the persistence of the post-conflict slowing following incongruent trials (Rey-Mermet & Meier, 2016), then the post-conflict slowing would be, in the present study, as long-lasting as the bivalency effect (Meier et al., 2009; Rey-Mermet & Meier, 2013). In this case, performance after incongruent trials would be slowed across several trials. However, it is also possible that the post-conflict slowing would be masked because congruent trials were used as baseline (see Verguts et al., 2011). In this case, no slowing would be expected after incongruent trials.
Method
Participants
Participants were 156 volunteers (26 in each between-subjects condition) from the University of Bern. Overall, we replaced nine participants (five participants because of an accuracy level on incongruent trials less than 50%, two because of a technical error, and two because they did not follow task instructions). Demographic characteristics of the sample are described in Table 2 (right part). The study was approved by the local ethics committee of the University of Bern, and all participants gave written consent.
Materials
In the following section, the stimuli for each task are presented. An overview of the manipulation is also presented in Table 2 (left part).
Stroop task
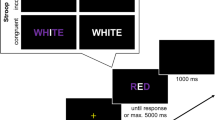
For the Stroop task, participants performed a color decision on color words. The congruent stimuli were the four German words for blue, red, green, and yellow (i.e., blau, rot, grün, and gelb), displayed in blue, red, green, and yellow, respectively. Incongruent stimuli were stimuli in which the color words were printed in a non-corresponding color (e.g., the color word red printed in blue). For each participant, six incongruent stimuli were determined randomly and without replacement. All stimuli were presented on a gray background at the center of the computer screen in 60-point Times New Roman. Participants used four response keys (v, b, n, m) with their left and right index and middle fingers. These response keys were mapped to the colors blue, red, green, and yellow, respectively.
Simon task
For the Simon task, participants performed a color decision on symbols. The symbols were $, #, §, %, displayed either in blue, red, green, or yellow. All stimuli were presented on a gray background in 60-point Times New Roman in one of the four corners of a centered, non-displayed 4 cm × 4 cm square. Participants used four response keys (g, b, n, j) with their left and right index and middle fingers. These response keys were mapped to the colors blue, red, green, and yellow, respectively. Stimuli were congruent when the position of the symbol on the computer screen corresponds to the location of the response key required by the color decision. In contrast, they were incongruent when the position of the symbol on the computer screen does not correspond to the location of the relevant response key. For each participant, six incongruent stimuli were determined randomly and without replacement.
Flanker task
For the flanker task, participants performed a letter identification on letter triplicates. The congruent stimuli were the four letters H, P, R, and S, displayed as triplicates with the same letters (e.g., HHH). Incongruent stimuli were the same four letters, but the central letter was different from the flanking letters (e.g., SHS). For each participant, six incongruent stimuli were determined randomly and without replacement. All stimuli were presented on a gray background in 60-point Times New Roman at the center of computer screen. Participants used four response keys (v, b, n, m) with their left and right index and middle fingers. These response keys were mapped to the letters H, P, R, and S, respectively.
Procedure
In each condition, participants were tested individually. Participants were instructed to perform a color decision on color words for the Stroop task, a color decision on symbols for the Simon task, and a letter decision on letter triplicates for the flanker task. Participants were instructed to press one of the four computer keys with their index and middle fingers of their left and right hands for each task. The mapping information, printed on paper, was presented below the computer screen throughout the experiment. For the Stroop task, participants were informed that, occasionally, the color word would not correspond to the color in which it was printed (e.g., the word red printed in blue), and that they had to proceed as usual by responding to the color of the color word. For the Simon task, they were informed that, occasionally, the position of the stimulus would not correspond to the location of the response key (e.g., a red symbol requiring a lower left key press but presented on the upper right corner), and that they had to proceed as usual by responding to the color of the symbol. For the flanker task, they were informed that, occasionally, the central letter would not correspond to the flanking letters (e.g., HSH), and that they had to proceed as usual by responding to the central letter.
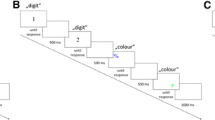
After the instructions, a block of 120 congruent trials was presented for practice. The stimulus for each trial was determined pseudorandomly so that neither the task-relevant stimulus feature nor the task-irrelevant stimulus feature repeated. The stimulus was displayed until the participant responded. Then, the screen blanked for 500 ms before the next stimulus appeared. After every four trials, an additional blank interval of 500 or 1,500 ms was included so that the screen blanked for 1,000 ms in half of participants, and for 2,000 ms in the other half. A sequence of four trials for each task (Stroop, Simon, and flanker) is illustrated in Fig. 1. After the practice block and a brief break, each participant completed three experimental blocks without break between blocks. The first block included 128 trials, with the first eight trials serving as warm-up trials that were discarded from the analyses. The second and third blocks had 120 trials each.

Example of one sequence of four congruent trials in Experiment 1. a Stroop task. On each trial, participants carried out a color decision on color words. On an incongruent trial (not pictured here), color words did not correspond to the color in which they were printed. b Simon task. On each trial, participants carried out a color decision on colored symbols. On an incongruent trial (not pictured here), the position of the symbol on the computer screen did not correspond to the location of the response key. c Flanker task. On each trial, participants carried out a letter decision on triplicates of letters. On an incongruent trial (not pictured here), the central letter was different from the flanking letters. (Color figure online)
In the first and third block, only congruent stimuli were presented. In the second block, stimuli were congruent, except on six trials in which incongruent stimuli appeared. Incongruent stimuli were always displayed on the fourth position of a four-trial sequence, and they were evenly interspersed among the 120 trials of the block. Thus, they occurred in every 20th trial, specifically in the 12th, 32nd, 52nd, 72nd, 92nd, and 112th trial. The entire experiment lasted about 15 minutes.
Data preparation and analysis
For each participant, the accuracy rates and the median reaction times (RTs) were computed for each trial following an incongruent stimulus in Block 2 and for each corresponding trial in the Blocks 1 and 3. Specifically, an incongruent stimulus was presented on every 20th trial in Block 2, and this trial was designated with the label T, with succeeding trials labeled T+1, T+2, and so on until T+19. Trials from Blocks 1 and 3 were labeled accordingly. To remove any confound with error and post-error slowing (e.g., Carter & van Veen, 2007; Kleiter & Schwarzenbacher, 1989; Notebaert et al., 2009), median RTs were computed on correct responses whose immediate preceding trial and incongruent trial also involved a correct response.
To ensure that participants were slower and less correct on incongruent trials than on congruent trials, we first compared performance on Trials T from Block 2 (i.e., the incongruent trials) with performance on Trials T from Blocks 1 and 3 (i.e., the corresponding congruent trials). To this end, we conducted a three-way analysis of variance (ANOVA) with block (Block 1, Block 2, Block 3) as a within-subject factor, and task (Stroop, Simon, flanker) and interval (1,000 ms, 2,000 ms) as between-subjects factors.
The main objective of the present experiment was to determine the trajectory of the post-conflict slowing following incongruent trials. Thus, we assessed whether performance in Block 2 following incongruent trials is slower than performance in Blocks 1 and 3. To this end, we carried out a four-way ANOVA with block (Block 1, Block 2, Block 3) and trial (T+1 until T+19) as within-subject factors, and task (Stroop, Simon, flanker) and interval (1,000 ms, 2,000 ms) as between-subjects factors. We then disentangled the critical interaction by performing follow-up two-way repeated-measures ANOVAs for each task, with the factors block (Block 1, Block 2, Block 3) and trial (T+1 until T+19). Finally, to assess the performance difference between the different blocks, we conducted follow-up one-way repeated-measures ANOVA with the factor block (Block 1, Block 2, Block 3). In these one-way ANOVAs, we focused on the quadratic component of the block effect because this is informative about a difference in Block 2 compared to Blocks 1 and 3.Footnote 3
Means and standard errors for each task and time interval are presented for RTs and accuracy in Table A1 of the supplementary material. As our focus was mainly on RT data, and accuracy was close to ceiling (98%), accuracy data is referred to only when diverging from RT data. We used an alpha level of 0.05, which was Bonferroni adjusted for multiple comparisons. Greenhouse–Geisser corrections are reported where appropriate and effect sizes are expressed as partial eta (ηp 2) values.
Results
Performance on Trial T
The three-way ANOVA across blocks, tasks, and intervals is shown in Table 3, separately for RT and accuracy. Performance on Trial T is presented in Table 4. Most importantly, the RT analysis revealed a significant main effect of block, with a significant quadratic component, F(1, 155) = 405.81, p < .001, ηp 2 = .72. Thus, as expected, performance was slower on incongruent trials from Block 2 than on the corresponding congruent trials from Blocks 1 and 3 (see Table 4). For accuracy, the three-way ANOVA revealed a significant interaction between block and task. The follow-up quadratic components revealed that responses were less accurate in incongruent trials from Block 2 than in the corresponding congruent trials from Blocks 1 and 3 for the Stroop and Simon tasks, F(1, 51) = 14.02, p < .001, ηp 2 = .22, and F(1, 51) = 88.40, p < .001, ηp 2 = .63, respectively, but not for the flanker task, F(1, 51) = 0, p = 1, ηp 2 = 0. Thus, for accuracy, the difference between incongruent and congruent trials was significant for the Stroop and Simon tasks only (see Table 4).
Impact of incongruent trials on subsequent congruent trials
The most relevant results are the RTs from the congruent trials in Block 2 compared to those in Blocks 1 and 3. These results are depicted in Fig. 2. The results of the four-way ANOVA are shown in Table 5. Critically, the RT analysis revealed a significant main effect of block and significant interactions between block and trial as well as between block, trial, and task. Thus, performance was slowed after incongruent trials in Block 2 compared to Blocks 1 and 3, and this slowing changed across subsequent congruent trials (see Fig. 2). Moreover, this change differed across tasks.

Experiment 1: Impact of incongruent trials on subsequent congruent trials. Trial T refers to the incongruent trial in Block 2, and subsequent trials (represented here) are labeled T+1, T+2, and so on. Error bars represent within-subject confidence intervals (see Cousineau, 2005; Morey, 2008). a, b, c Mean reaction times on congruent trials from Block 1 (empty squares), Block 2 (filled circles), and Block 3 (empty diamonds). a Stroop task. b Simon task. c Flanker task. d Trajectory of the post-conflict slowing for each task (Stroop, Simon, and flanker). This slowing was computed as the difference between performance in Block 2 and performance averaged across Blocks 1 and 3. Filled symbols indicate a significant slowing (the alpha level of 0.05 was Bonferroni adjusted to 0.003 to account for the multiple comparisons)
To investigate this change more thoroughly, we performed additional two-way ANOVAs for each task separately, with block and trial as within-subject factors. These revealed a significant interaction between block and trial for all tasks – that is, the Stroop task: F(8.46, 431.42) = 2.91, p = .003, η p 2 = .05; the Simon task: F(16.08, 819.95) = 2.86, p < .001, ηp 2 = .05; and the flanker task: F(15.70, 800.81) = 2.77, p < .001, ηp 2 = .05. The follow-up relevant quadratic components are shown in Table 6 for each trial and each task. They revealed that, first, performance was slowed on the first two and three trials immediately following incongruent trials, and then performance slowing became more sporadic, affecting only some of the trials. This later and more sporadic slowing affected more subsequent trials in the Simon and flanker tasks than in the Stroop task. However, the post-conflict slowing was longer lasting for the Stroop task than for the Simon and flanker tasks because it came back at T+16 in the Stroop task but not in the Simon or flanker tasks (in these tasks, it only came back at T+12).
Table 5 also shows that the interaction between block and interval was significant. Follow-up one-way repeated-measures ANOVA with the factor block (Block 1, Block 2, Block 3) revealed a significant main effect of block with a significant quadratic component for both intervals — 1,000 ms interval: main effect, F(2, 154) = 35.55, p < .001, ηp 2 = .32, and quadratic component, F(1, 77) = 70.43, p < .001, ηp 2 = .48; 2,000 ms interval: main effect, F(1.69, 129.86) = 9.37, p < .001, ηp 2 = .11, and quadratic component, F(1, 77) = 20.34, p < .001, ηp 2 = .21. Thus, the performance slowing was found in both intervals, but it was larger in the 1,000 ms interval (Block 1: M = 616 ms, SE = 13; Block 2: M = 661 ms, SE = 12; Block 3: M = 627 ms, SE = 11) than in the 2,000 ms interval (Block 1: M = 634 ms, SE = 16; Block 2: M = 654 ms, SE = 13; Block 3: M = 620 ms, SE = 12).
Discussion
The results of Experiment 1 showed a performance slowing for the first few trials immediately following incongruent trials. This slowing occurred in all three tasks. It lasted circa 5 seconds (i.e., required for making up to the third trial, i.e., 3 decisions, each requiring approximately 650 ms, plus 2 blanks of 500 ms, plus 1 blank of 2,000 ms). The results also showed a more sporadic performance slowing on later trials. This later and more sporadic slowing affected more trials in the Simon and flanker tasks than in the Stroop task. However, it was longer lasting for the Stroop task than for the Simon and flanker tasks. Therefore, the present findings indicate that when participants are not required to switch between at least two tasks, the post-conflict slowing following incongruent trials persists across trials but is not as long-lasting as the post-conflict slowing following bivalent stimuli (Meier et al., 2009; Rey-Mermet & Meier, 2013, 2016).
In Experiment 1, the post-conflict slowing was investigated on congruent trials. However, this might not be optimal to find a long-lasting post-conflict slowing following incongruent trials because congruent trials could invoke a post-focusing process, which might mask the post-conflict slowing (Verguts et al., 2011). To test this possibility, we conducted a second experiment (i.e., Experiment 2) in which the post-conflict slowing was examined on neutral trials, that is, on trials on which no post-focusing process could occur. Therefore, we used the same design as Experiment 1, but we occasionally presented incongruent trials among neutral trials. As increasing the interval from 1,000 ms to 2,000 ms after incongruent did not affect the trajectory of the post-conflict slowing in Experiment 1, we removed this manipulation from the design of Experiment 2. Here, we expected to find a longer lasting post-conflict slowing following incongruent trials if the post-conflict slowing observed in Experiment 1 was masked on some congruent trials (Verguts et al., 2011).
Experiment 2
Method
Participants
Participants were 78 volunteers (26 in each task) from the University of Bern. We replaced two participants because of an accuracy level on incongruent trials less than 50%. Demographic characteristics of the sample are described in Table 2 (right part).
Materials
The material was the same as in Experiment 1, except that instead of congruent stimuli, neutral stimuli were presented. That is, for the Stroop task, the neutral stimuli were the symbols $ $ $, # # #, § § §, and % % %, displayed in blue, yellow, red, and green, respectively. For the Simon task, the neutral stimuli were the symbols $, #, §, and % displayed centrally. For the flanker task, the neutral stimuli were the four triplicates <H>, §P§, %R%, and +S+.
Procedure
The procedure was the same as in Experiment 1, except that there was only one interval (1,000 ms).
Data preparation and analysis
Data preparation and data analysis were the same as in Experiment 1, except for the following modifications. First, performance on Trial T was investigated with a two-way ANOVA with block (Block 1, Block 2, Block 3) as a within-subject factor, and task (Stroop, Simon, flanker) as a between-subjects factor. Second, the trajectory of the post-conflict slowing was assessed with a three-way ANOVA with block (Block 1, Block 2, Block 3) and trial (T+1 until T+19) as within-subject factors, and task (Stroop, Simon, flanker) as a between-subjects factor.
Results
Performance on Trial T
The two-way ANOVA across blocks and tasks is shown in Table 3. Performance on Trial T is presented in Table 4. Most importantly, the RT analysis revealed a significant interaction between block and task. The follow-up quadratic components revealed that responses were slower in incongruent trials from Block 2 than in the corresponding neutral trials from Blocks 1 and 3 in all three tasks — that is, the Stroop task: F(1, 25) = 91.50, p < .001, ηp 2 = .78; the Simon task: F(1, 25) = 65.03, p < .001, ηp 2 = .72; and the flanker task, F(1, 25) = 68.81, p < .001, ηp 2 = .73. Thus, for RTs, the difference between incongruent and neutral trials was larger for the Stroop and flanker tasks than for the Simon task (see Table 4). For accuracy, the three-way ANOVA revealed a significant main effect of block, with a significant quadratic component, F(1, 77) = 8.95, p = .004, ηp 2 = .10. Thus, as expected, performance was less correct on incongruent trials from Block 2 than on the corresponding neutral trials from Blocks 1 and 3 (see Table 4).
Impact of incongruent trials on subsequent neutral trials
The most relevant results are the RTs from the neutral trials in Block 2 compared to those in Blocks 1 and 3. These results are depicted in Fig. 3. For accuracy, the descriptive results are presented in Table A2 of the supplementary material. The results of the three-way ANOVA are shown in Table 7. Critically, RT analysis revealed a significant main effect of block and a significant interaction between block and trial. Thus, performance was slowed after incongruent trials in Block 2 compared to Blocks 1 and 3, and this slowing changed across subsequent neutral trials (see Fig. 3).

Experiment 2: Impact of incongruent trials on subsequent neutral trials. Trial T refers to the incongruent trial in Block 2, and subsequent trials (represented here) are labeled T+1, T+2, and so on. Error bars represent within-subject confidence intervals (see Cousineau, 2005; Morey, 2008). a, b, c. Mean reaction times on neutral trials from Block 1 (empty squares), Block 2 (filled circles), and Block 3 (empty diamonds). a Stroop task. b Simon task. c Flanker task. d Trajectory of the post-conflict slowing. This slowing was computed as the difference between performance in Block 2 and performance averaged across Blocks 1 and 3. Filled symbols indicate a significant slowing (the alpha level of 0.05 was Bonferroni adjusted to 0.003 to account for the multiple comparisons)
To investigate this change more thoroughly, we focused as in Experiment 1 on the follow-up relevant quadratic components. These are shown in Table 8 for each trial. They revealed that, first, performance was slowed on the first three trials immediately following incongruent trials, and then the performance slowing became more sporadic, affecting only some of the trials.
Discussion
The results of Experiment 2 replicate the findings of the first experiment by showing a performance slowing for the first few trials immediately following incongruent trials. It lasted circa 4 seconds (i.e., required for making up to the third trial, i.e., 3 decisions, each requiring approximately 660 ms, plus 2 blanks of 500 ms, plus 1 blank of 1,000 ms). The results also showed a performance slowing on later trials. In contrast to Experiment 1, however, the trajectory of the post-conflict slowing did not differ across the three tasks. Together, the results of Experiment 2 indicate that no post-focusing process masks the post-conflict slowing in Experiment 1 (cf. Verguts et al., 2011). More generally, these findings emphasize that even if the post-conflict slowing following incongruent trials was not as long-lasting as the post-conflict slowing following bivalent stimuli (e.g., Meier et al., 2009; Rey-Mermet & Meier, 2013), this post-conflict slowing clearly affected more than one trial.
However, occasionally presenting incongruent trials among congruent trials (Experiment 1) or neutral trials (Experiment 2) has the disadvantage that incongruent trials are not only conflict stimuli but also infrequent events. Therefore, it is possible that the slowing following incongruent trials in Experiments 1 and 2 was not caused by the conflict induced by incongruent trials but rather by their infrequency (Notebaert et al., 2009; Notebaert & Verguts, 2011; Núñez Castellar et al., 2010; Rey-Mermet & Meier, 2013). To test this possibility, we conducted Experiment 3, in which we reversed the ratio of incongruent and congruent trials of Experiment 1. That is, we used the same design as Experiment 1, but we occasionally presented congruent trials among incongruent trials. We hypothesized that if the post-conflict slowing following incongruent trials was only caused by the infrequency of incongruent trials (Notebaert & Verguts, 2011; Rey-Mermet & Meier, 2013), infrequent congruent trials would result in a similar performance slowing as the one observed in Experiments 1 and 2.
Experiment 3
Method
Participants
Participants were 78 volunteers (26 in each task) from the University of Bern. Demographic characteristics of the sample are described in Table 2 (right part).
Materials
The material was the same as in Experiment 1.
Procedure
The procedure was the same as in Experiment 1, except for the following two modifications. First, only incongruent stimuli were presented in the first and third blocks. In the second block, stimuli were incongruent, except on six trials in which congruent stimuli appeared. Second, there was only one interval (1,000 ms).
Data preparation and analysis
The data preparation and data analysis were the same as in Experiment 2.
Results
Performance on Trial T
The two-way ANOVA across blocks and tasks is shown in Table 3. Performance on Trial T is presented in Table 4. Most importantly, the RT analysis revealed a significant interaction between block and task. The follow-up quadratic components revealed that responses were significantly slower in incongruent trials from Blocks 1 and 3 than in the corresponding congruent trials from Block 2 in the Simon task only (see Table 4), F(1, 25) = 26.88, p < .001, ηp 2 = .52. The difference between incongruent and congruent trials were not significant for the Stroop and flanker tasks — that is, the Stroop task: F(1, 25) = 6.29, p = .019, ηp 2 = .20; and the flanker task, F(1, 25) = 4.71, p = .040, ηp 2 = .16, with the alpha level of 0.05 being Bonferroni adjusted to 0.017. For accuracy, the three-way ANOVA revealed a significant main effect of block, with a significant quadratic component, F(1, 77) = 6.80, p = .011, ηp 2 = .08. Thus, in all three tasks, performance was less correct on incongruent trials from Blocks 1 and 3 than on the corresponding congruent trials from Block 2 (see Table 4).
Impact of congruent trials on subsequent incongruent trials
The most relevant results are the RTs from the incongruent trials in Block 2 compared to those in Blocks 1 and 3. These results are depicted in Fig. 4. For accuracy, the descriptive results are presented in the Table A3 of the supplementary material. The results of the three-way ANOVA are shown in Table 9. Critically, the RT analysis revealed a significant main effect of block and a significant interaction between block and trial. The follow-up quadratic components of the block effect are shown in Table 10. These revealed that performance was slowed only on the 12th trial following congruent trials.

Experiment 3: Impact of congruent trials on subsequent incongruent trials. Trial T refers to the congruent trial in Block 2, and subsequent trials (represented here) are labeled T+1, T+2, and so on. Error bars represent within-subject confidence intervals (see Cousineau, 2005; Morey, 2008). a, b, c Mean reaction times on incongruent trials from Block 1 (empty squares), Block 2 (filled circles), and Block 3 (empty diamonds). a Stroop task. b Simon task. c Flanker task. d Trajectory of the “post-infrequency” slowing. This slowing was computed as the difference between performance in Block 2 and performance averaged across Blocks 1 and 3. Filled symbols indicate a significant slowing (the alpha level of 0.05 was Bonferroni adjusted to 0.003 to account for the multiple comparisons)
Discussion
The results of Experiment 3 showed a performance slowing only on the 12th trial following congruent trials, probably induced by some kind of expectancy-based monitoring process. These findings are not compatible with a conservative version of an expectancy-based monitoring account (e.g., Meier et al., 2006; Smith, 2003) because according to such an account, participants would monitor for infrequent events so that the monitoring process would steadily increase across trials and thus result in an increase of the slowing across trials. Nevertheless, it seems plausible that participants anticipated the infrequent events on the 12th trial, thus directing their attention away from the trial processing and slowing down their performance. But why did they anticipate specifically on the 12th trial? As trials were presented in sequences of four, and infrequent events were presented on the fourth position of this sequence, this could have emphasized this position. Moreover, participants should have realized that the infrequent events did not occur every four trials but with a larger extent so that they expected infrequent events on the 12th trial. Thus, according to this explanation, participants did not steadily monitor for the occurrence of incongruent trials but rather anticipated them specifically due to the design of the experiment.
More generally, the findings of Experiment 3 showed that slowing following infrequent congruent trials is different from the post-conflict we observed in Experiments 1 and 2. This suggests that the conflict, but not the infrequency of incongruent trials, is responsible for the post-conflict slowing. However, in Experiment 3, it is possible that because incongruent trials were presented more frequently than congruent trials, more control processes were overall engaged. This could have reduced the slowing observed in this experiment, similar to a proportion congruency effect (Gratton et al., 1992; Hommel, 1994; Logan & Zbrodoff, 1979; Lowe & Mitterer, 1982). Thus, the design of Experiment 3 might be suboptimal to find the impact of infrequent events on subsequent trials. To ensure that the post-conflict slowing results from the conflict induced by incongruent trials, we conducted a fourth experiment. In this experiment, most trials were congruent, and infrequent (non-conflict) events were neutral trials. As for Experiment 3, we expected that if the post-conflict slowing following incongruent trials was only caused by the infrequency of incongruent trials (Notebaert & Verguts, 2011; Rey-Mermet & Meier, 2013), infrequent neutral trials would result in a similar performance slowing as the slowing observed in the first two experiments.
Experiment 4
Method
Participants
Participants were 78 volunteers (26 in each task) from the University of Bern. We replaced one participant because of an accuracy level on neutral trials less than 50%. Demographic characteristics of the sample are described in Table 2 (right part).
Materials
The material was the same as in Experiment 2.
Procedure
The procedure was the same as in Experiment 1, except for the following two modifications. First, only congruent stimuli were presented in the first and third blocks. In the second block, stimuli were congruent except on six trials in which neutral stimuli appeared. Second, there was only one interval (1,000 ms).
Data preparation and analysis
The data preparation and data analysis were the same as in Experiment 2.
Results
Performance on Trial T
The two-way ANOVA across blocks and tasks is shown in Table 3. Performance on Trial T is presented in Table 4. Most importantly, the RT analysis revealed a significant interaction between block and task. The follow-up quadratic components showed that responses were slower in neutral trials from Block 2 than in the corresponding congruent trials from Blocks 1 and 3 in all three tasks — that is, the Stroop task: F(1, 25) = 97.59, p < .001, ηp 2 = .80; the Simon task: F(1, 25) = 118.92, p < .001, ηp 2 = .83; and the flanker task, F(1, 25) = 71.27, p < .001, ηp 2 = .74. This difference was, however, larger for the Simon task than for the Stroop and flanker tasks (see Table 4).
For accuracy, the three-way ANOVA also revealed a significant interaction between block and task. The follow-up quadratic components revealed that responses were significantly less correct in neutral trials from Block 2 than in the corresponding congruent trials from Blocks 1 and 3 in the Simon task (see Table 4), F(1, 25) = 14.14, p = .001, ηp 2 = .36. The difference between neutral and congruent trials, was, however, not significant for the Stroop and flanker tasks — that is, the Stroop task: F(1, 25) = 2.00, p = .170, ηp 2 = .07; and the flanker task, F(1, 25) = 0, p = 1, ηp 2 < .001.
Impact of neutral trials on subsequent congruent trials
The most relevant results are the RTs from the congruent trials in Block 2 compared to those in Blocks 1 and 3. These results are depicted in Fig. 5. For accuracy, the descriptive results are presented in Table A4 of the supplementary material. The results of the three-way ANOVA are shown in Table 11. Critically, the RT analysis revealed a significant main effect of block and significant interactions between block and trial as well as between block, trial and task. Thus, performance was slowed after infrequent neutral trials in Block 2 compared to Blocks 1 and 3, and this slowing changed across subsequent congruent trials (see Fig. 5). Moreover, this change differed across tasks.

Experiment 4: Impact of neutral trials on subsequent congruent trials. Trial T refers to the neutral trial in Block 2, and subsequent trials (represented here) are labeled T+1, T+2, and so on. Error bars represent within-subject confidence intervals (see Cousineau, 2005; Morey, 2008). a, b, c Mean reaction times on congruent trials from Block 1 (empty squares), Block 2 (filled circles), and Block 3 (empty diamonds). a Stroop task. b Simon task. c Flanker task. d Trajectory of the “postx-infrequency” slowing for each task (Stroop, Simon, and flanker). This slowing was computed as the difference between performance in Block 2 and performance averaged across Blocks 1 and 3. Filled symbols indicate a significant slowing (the alpha level of 0.05 was Bonferroni adjusted to 0.003 to account for the multiple comparisons)
To investigate this change more thoroughly, we performed additional two-way ANOVAs for each task separately, with block and trial as within-subject factors. These ANOVAs revealed a significant interaction between block and trial for the Stroop task, F(36, 900) = 1.63, p = .012, ηp 2 = .06, and the flanker task, F(36, 900) = 2.01, p < .001, ηp 2 = .07. For the Simon task, however, the interaction did not approach the level of significance, F(36, 900) = 1.41, p = .059, ηp 2 = .06. For all three tasks, the follow-up relevant quadratic components of the block effect are shown in Table 12. These revealed a significant performance slowing on the first trial following infrequent neutral trials for the flanker task, but no significant performance slowing for the Stroop task (see Fig. 5). For the sake of comparison, we also computed the quadratic components of the block effect for each trial of the Simon task (see Table 12). These revealed a significant performance slowing on the first trial following infrequent neutral trials (see Fig. 5).
Discussion
The results of Experiment 4 showed at best a performance slowing on the first trial following infrequent neutral trials, probably indicating some orienting response (Notebaert et al., 2009; Notebaert & Verguts, 2011; Núñez Castellar et al., 2010; Rey-Mermet & Meier, 2013). However, no slowing was observed on the later trials. These results are in contrast with those of Experiment 3, in which a slowing was only observed on a later trial. Together, the results of Experiments 3 and 4 reveal that when incongruent trials were presented frequently and thus induced more cognitive control processes, there was no slowing caused by an orienting response but a slowing induced by some anticipation process. In contrast, when no incongruent trials were presented and thus cognitive control processes were less necessary, only a slowing due to an orienting response (if any) occurred. More generally, the results of both Experiments 3 and 4 indicate that the slowing following infrequent events is different from the slowing following incongruent trials. This suggests that the conflict, but not the infrequency of incongruent trials, is responsible for the post-conflict slowing.
Analyses across experiments
To strengthen the finding that the post-conflict slowing is different from the slowing following an infrequent event, we conducted a follow-up analysis in which experiment was added as a between-subjects variable. That is, we carried a four-way ANOVA with block (Block 1, Block 2, Block 3) and trial (T+1 until T+19) as within-subject factors, and task (Stroop, Simon, flanker) and experiment (Experiment 1, Experiment 2, Experiment 3, Experiment 4) as between-subjects factors. As shown in Table 13 (left part), the results revealed a significant four-way interaction, ensuring that the slowing we observed in each experiment differed across experiments.
Moreover, combining the data from Experiment 1 (only the 1,000 ms interval condition) and Experiment 3 allowed us to analyze a fully counterbalanced design with one condition in which incongruent trials were presented infrequently across congruent trials (i.e., Experiment 1) and another condition in which congruent trials were presented infrequently across incongruent trials (i.e., Experiment 3). With these data, we conducted a four-way ANOVA with block (Block 1, Block 2, Block 3) and trial (T+1 until T+19) as within-subject factors, and task (Stroop, Simon, flanker) and experiment (Experiment 1 with the 1,000 ms interval condition, Experiment 3) as between-subjects factors. As shown in Table 13 (right part), the results also revealed a significant four-way interaction, ensuring statistically that the post-conflict slowing observed after incongruent trials in Experiment 1 was different from the slowing observed after infrequent congruent trials in Experiment 3.
General discussion
The purpose of the present study was to determine how long-lasting the post-conflict slowing following incongruent trials is. To this end, we performed two experiments in which we occasionally presented incongruent trials among non-conflict stimuli (i.e., congruent trials in Experiment 1 and neutral trials in Experiment 2), and we assessed the persistence of slowing following incongruent trials by determining the trajectory of performance slowing across the subsequent trials that immediately followed. In both experiments, the results showed a performance slowing for the first few trials immediately following incongruent trials. On some later trials, performance was still slowed. To ensure that this performance slowing was not caused by the infrequency of incongruent trials among non-conflict stimuli, we conducted two further experiments. In Experiment 3, congruent trials were occasionally presented among incongruent trials; in Experiment 4, neutral trials were occasionally presented among congruent trials. In both experiments, the results revealed that performance was only affected at best on one trial after infrequent events were presented. This rules out the explanation that the post-conflict slowing we observed in Experiments 1 and 2 was only caused by the infrequency of incongruent trials. This rather demonstrates that the conflict induced by incongruent trials is responsible for the post-conflict slowing.
The question is now: How can we explain the post-conflict slowing following incongruent trials? As the post-conflict slowing following incongruent trials was not in the present study as long-lasting as the post-conflict slowing following bivalent stimuli (Meier et al., 2009; Rey-Mermet & Meier, 2013), this calls into question the episodic context binding explanation, at least when no task switching is required (but see Rey-Mermet & Meier, 2016). Moreover, as stated above, because the post-conflict slowing was not caused by the infrequency of incongruent trials, an orienting response account is not sufficient to explain the slowing on the first few trials following incongruent trials (Notebaert et al., 2009; Notebaert & Verguts, 2011; Núñez Castellar et al., 2010; Rey-Mermet & Meier, 2013). Finally, according to a conservative version of the expectancy-based monitoring account (e.g., Meier et al., 2006; Smith, 2003), the performance slowing would steadily increase across the later trials. However, although the present findings show a post-conflict slowing following incongruent trials on some later trials, the magnitude of this slowing did not follow a pattern, such as a linear increase (see Figs. 2 and 3).
Most probably, the post-conflict slowing we observed in the present study results from a focusing process (Ullsperger, Bylsma, & Botvinick, 2005; cf. Verguts et al., 2011). That is, encountering the conflict induced by incongruent trials widens attention or more precisely directs attention to irrelevant features. Then, a focusing process is necessary to redirect attention to the relevant response feature. As the post-conflict slowing occurred on congruent and neutral trials (see Experiments 1 and 2), attention seems not only directed to irrelevant response features (as it would be the case in congruent trials) but also to other irrelevant features (as it must be the case for neutral trials as they have no irrelevant response feature). Thus, this additional focusing process slows performance, resulting in the post-conflict slowing. Critically, to explain the irregularity of the post-conflict slowing across later trials, this focusing process cannot be an all-or-none process that is triggered as soon as a stimulus is presented after an incongruent stimulus. The present results rather suggest that this process is variable. Accordingly, when attention is sufficiently widened (e.g., because the current trial is the fourth of the sequence of four trials and participants anticipated incongruent trials on this trial as they realized that incongruent trials were sometimes presented on this position), a focusing process is necessary, and thus a post-conflict slowing occurs on later trials. In contrast, when attention is not widened, no focusing process is required, and therefore no slowing occurs. These variations might be explained by the specific designs of the experiments (see Verguts et al., 2011) as well as by fluctuations in current attentional demands and/or in current motivation states (see De Jong, 2000).
One may wonder why in the present study, and in particular in Experiment 1, a post-conflict slowing was found on congruent trials and even might be caused by a focusing process, whereas Verguts et al. (2011) argue that the post-conflict slowing was masked by a focusing process on congruent trials. The reason for this difference might be in the conditions under which the post-conflict slowing was investigated. In Verguts et al. (2011), the post-conflict slowing was computed on congruent or neutral trials whose immediate preceding trial was either neutral, congruent, or incongruent. Thus, as all trial types were intermixed, it is possible that control processes carried over the trials. This would slow the trials used as baseline, which might mask some post-conflict slowing. In the present study, the post-conflict slowing emerged from the comparison between a pure block of congruent or neutral trials and a mixed block including some incongruent trials. Thus, the carry-over of cognitive processes was avoided because baseline trials (i.e., congruent and neutral trials) were presented in pure blocks. Moreover, as the focusing process is an additional process in the block with incongruent trials compared to the pure block, this results in a performance slowing.
Together with our previous findings (Rey-Mermet & Meier, 2016) in which the post-conflict slowing was observed on tasks sharing features with the incongruent trials and on tasks sharing no relevant features with the incongruent trials (i.e., tasks including univalent stimuli), the present results suggest that with our design, the trials used as baseline (i.e., congruent, neutral, or univalent) do not seem to be critical for the post-conflict slowing following incongruent trials. Rather, the conditions under which the post-conflict slowing was investigated (e.g., whether participants were asked to switch between tasks or whether all trial types were intermixed) seem central. At a global level, this is in line with Verguts et al. (2011) who emphasize the importance of considering under which conditions the post-conflict slowing is investigated. Furthermore, this underscores the necessity of further experiments to determine why using a task-switching design results in a long-lasting post-conflict slowing or to investigate the real impact of this post-conflict slowing in studies investigating the congruency sequence effect or proportion congruency effect, for example.
More generally, the results of the present study reveal that the post-conflict slowing following incongruent trials can be different from the post-conflict slowing following bivalent stimuli or prospective memory targets (see, e.g., Meier & Rey-Mermet, 2012a, 2012b). Therefore, the source of conflict (bivalent stimulus, prospective memory target, or incongruent trial) is important in determining the kind of adjustment of cognitive control underlying the post-conflict slowing (see Braem et al., 2014; Egner, 2008). However, these sources of conflict are interconnected (e.g., Meier & Rey-Mermet, 2012b; Steinhauser & Hübner, 2009). That is, bivalent stimuli and prospective memory targets are incongruent because they may involve competing responses (see Table 1). Conversely, incongruent stimuli might be considered as bivalent. For example, in a task-switching paradigm, participants can encounter Stroop stimuli and thus can be asked to switch between naming the color of the word and reading the word. Therefore, at a conceptual level, the different sources of conflict share critical features. Further research is thus necessary to understand what is critical in these sources of conflict to induce different kinds of post-conflict slowing.
To summarize, the results of the present study show that the conflict induced by incongruent trials results in a consistent slowing for the first few subsequent trials. In addition, a slowing was also found on later trials. Moreover, this post-conflict slowing was not caused by an orienting response due to the infrequency of incongruent trials. Together, the present findings demonstrate that the post-conflict slowing induced by incongruent stimuli is longer lasting than previously thought. Interestingly, this post-conflict slowing differs from the post-conflict slowing induced by bivalent stimuli or prospective memory targets.
Notes
There are also some studies focusing on the impact of several successive incongruent trials. These revealed a performance slowing on the immediate subsequent congruent trial even when the two or three previous trials were incongruent (Duthoo, Abrahamse, Braem, & Notebaert, 2014; Jiménez & Méndez, 2013, 2014).
In order to be as concise as possible and to assess statistically the differences between the post-conflict slowing induced by the Stroop, Simon and flanker tasks, we added task as a between-subjects variable. However, the different tasks were thought as separate experiments. Therefore, although recruitment and testing conditions were similar across all tasks, the data collection for the three tasks was not started simultaneously. However, as soon as half of participants were tested, all three tasks were tested concurrently.
To account for baseline RT differences between Stroop, Simon, and flanker tasks, we computed proportional scores for each participant in each experiment. That is, for each trial (i.e., T+1 until T+19), mean RT for Block 2 was subtracted from the mean RT averaged across Blocks 1 and 3; then, this difference was divided by the mean RT averaged across all blocks. In each experiment, the analyses on the proportional scores revealed the same findings as those with the RT, ruling out the possibility that our findings are caused by baseline RT difference between the tasks.
References
Akçay, Ç., & Hazeltine, E. (2008). Conflict adaptation depends on task structure. Journal of Experimental Psychology: Human Perception and Performance, 34(4), 958–973. doi:10.1037/0096-1523.34.4.958
Botvinick, M. M., Braver, T. S., Barch, D. M., Carter, C. S., & Cohen, J. D. (2001). Conflict monitoring and cognitive control. Psychological Review, 108(3), 624–652. doi:10.1037//0033-295X.108.3.624
Braem, S., Abrahamse, E. L., Duthoo, W., & Notebaert, W. (2014). What determines the specificity of conflict adaptation? A review, critical analysis, and proposed synthesis. Frontiers in Psychology, 5. doi:10.3389/fpsyg.2014.01134
Carter, C. S., & van Veen, V. (2007). Anterior cingulate cortex and conflict detection: An update of theory and data. Cognitive, Affective, & Behavioral Neuroscience, 7(4), 367–379. doi:10.3758/CABN.7.4.367
Cousineau, D. (2005). Confidence intervals in within-subject designs: A simpler solution to Loftus and Masson’s method. Tutorials in Quantitative Methods for Psychology, 1(1), 42–45.
De Jong, R. (2000). An intention-activation account of residual switch costs. In S. Monsell & J. S. Driver (Eds.), Control of cognitive processes: Attention and performance XVIII (pp. 356–376). Cambridge: MIT Press.
Duthoo, W., Abrahamse, E. L., Braem, S., Boehler, C. N., & Notebaert, W. (2014). The heterogeneous world of congruency sequence effects: An update. Frontiers in Psychology, 5. doi:10.3389/fpsyg.2014.01001
Duthoo, W., Abrahamse, E. L., Braem, S., & Notebaert, W. (2014). Going, going, gone? Proactive control prevents the congruency sequence effect from rapid decay. Psychological Research, 78(4), 483–493. doi:10.1007/s00426-013-0498-4
Egner, T. (2007). Congruency sequence effects and cognitive control. Cognitive, Affective, & Behavioral Neuroscience, 7(4), 380–390. doi:10.3758/CABN.7.4.380
Egner, T. (2008). Multiple conflict-driven control mechanisms in the human brain. Trends in Cognitive Sciences, 12(10), 374–380. doi:10.1016/j.tics.2008.07.001
Egner, T., Ely, S., & Grinband, J. (2010). Going, going, gone: Characterizing the time-course of congruency sequence effects. Cognition, 1, 154. doi:10.3389/fpsyg.2010.00154
Eriksen, B. A., & Eriksen, C. W. (1974). Effects of noise letters upon the identification of a target letter in a nonsearch task. Perception & Psychophysics, 16(1), 143–149. doi:10.3758/BF03203267
Gratton, G., Coles, M. G., & Donchin, E. (1992). Optimizing the use of information: Strategic control of activation of responses. Journal of Experimental Psychology: General, 121(4), 480–506.
Grundy, J. G., Benarroch, M. F. F., Woodward, T. S., Metzak, P. D., Whitman, J. C., & Shedden, J. M. (2013). The Bivalency effect in task switching: Event-related potentials. Human Brain Mapping, 34(5), 999–1012. doi:10.1002/hbm.21488
Grundy, J. G., & Shedden, J. M. (2014a). A role for recency of response conflict in producing the bivalency effect. Psychological Research, 78(5), 679–691. doi:10.1007/s00426-013-0520-x
Grundy, J. G., & Shedden, J. M. (2014b). Support for a history-dependent predictive model of dACC activity in producing the bivalency effect: An event-related potential study. Neuropsychologia, 57, 166–178. doi:10.1016/j.neuropsychologia.2014.03.008
Hommel, B. (1994). Spontaneous decay of response-code activation. Psychological Research, 56(4), 261–268.
Horga, G., Maia, T. V., Wang, P., Wang, Z., Marsh, R., & Peterson, B. S. (2011). Adaptation to conflict via context-driven anticipatory signals in the dorsomedial prefrontal cortex. Journal of Neuroscience, 31(45), 16208–16216. doi:10.1523/JNEUROSCI.2783-11.2011
Jiménez, L., & Méndez, A. (2013). It is not what you expect: Dissociating conflict adaptation from expectancies in a Stroop task. Journal of Experimental Psychology: Human Perception and Performance, 39(1), 271–284. doi:10.1037/a0027734
Jiménez, L., & Méndez, A. (2014). Even with time, conflict adaptation is not made of expectancies. Frontiers in Psychology, 5. doi:10.3389/fpsyg.2014.01042
Kleiter, G. D., & Schwarzenbacher, K. (1989). Beyond the answer: Post-error processes. Cognition, 32(3), 255–277. doi:10.1016/0010-0277(89)90037-1
Kray, J., & Lindenberger, U. (2000). Adult age differences in task switching. Psychology and Aging, 15(1), 126–147. doi:10.1037/0882-7974.15.1.126
Loft, S., Kearney, R., & Remington, R. (2008). Is task interference in event-based prospective memory dependent on cue presentation? Memory & Cognition, 36(1), 139–148. doi:10.3758/MC.36.1.139
Logan, G. D., & Zbrodoff, N. J. (1979). When it helps to be misled: Facilitative effects of increasing the frequency of conflicting stimuli in a Stroop-like task. Memory & Cognition, 7(3), 166–174. doi:10.3758/BF03197535
Lowe, D. G., & Mitterer, J. O. (1982). Selective and divided attention in a Stroop task. Canadian Journal of Psychology, 36(4), 684–700. doi:10.1037/h0080661
MacLeod, C. M. (1991). Half a century of research on the Stroop effect: An integrative review. Psychological Bulletin, 109(2), 163–203. doi:10.1037/0033-2909.109.2.163
Mayr, U., Awh, E., & Laurey, P. (2003). Conflict adaptation effects in the absence of executive control. Nature Neuroscience, 6(5), 450–452. doi:10.1038/nn1051
McDaniel, M. A., & Einstein, G. O. (2000). Strategic and automatic processes in prospective memory retrieval: A multiprocess framework. Applied Cognitive Psychology, 14(7), S127–S144.
Meier, B., & Rey-Mermet, A. (2012a). Beyond feature binding: Interference from episodic context binding creates the bivalency effect in task-switching. Frontiers in Psychology, 3, 386–394. doi:10.3389/fpsyg.2012.00386
Meier, B., & Rey-Mermet, A. (2012b). Beyond monitoring: After-effects of responding to prospective memory targets. Consciousness and Cognition, 21(4), 1644–1653. doi:10.1016/j.concog.2012.09.003
Meier, B., & Rey-Mermet, A. (2017). After-effects without monitoring costs: The impact of prospective memory instructions on task switching performance. Acta Psychologica. Advance online publication. doi:10.1016/j.actpsy.2017.04.010
Meier, B., Rey-Mermet, A., Woodward, T. S., Müri, R., & Gutbrod, K. (2013). Episodic context binding in task switching: Evidence from amnesia. Neuropsychologia, 51(5), 886–892. doi:10.1016/j.neuropsychologia.2013.01.025
Meier, B., Rey-Mermet, A., & Rothen, N. (2015). Turning univalent stimuli bivalent: Synesthesia can cause cognitive conflict in task switching. Cognitive Neuroscience, 1–8. doi:10.1080/17588928.2015.1017449
Meier, B., Woodward, T. S., Rey-Mermet, A., & Graf, P. (2009). The bivalency effect in task switching: General and enduring. Canadian Journal of Experimental Psychology, 63(3), 201–210. doi:10.1037/a0014311
Meier, B., Zimmermann, T. D., & Perrig, W. J. (2006). Retrieval experience in prospective memory: Strategic monitoring and spontaneous retrieval. Memory, 14(7), 872–889. doi:10.1080/09658210600783774
Metzak, P. D., Meier, B., Graf, P., & Woodward, T. S. (2013). More than a surprise: The bivalency effect in task switching. Journal of Cognitive Psychology, 25(7), 833–842. doi:10.1080/20445911.2013.832196
Metzker, M., & Dreisbach, G. (2009). Bidirectional priming processes in the Simon task. Journal of Experimental Psychology: Human Perception and Performance, 35(6), 1770–1783. doi:10.1037/a0015787
Morey, R. D. (2008). Confidence intervals from normalized data: A correction to Cousineau (2005). Tutorials in Quantitative Methods for Psychology, 4(2), 61–64.
Notebaert, W., Houtman, F., Opstal, F. V., Gevers, W., Fias, W., & Verguts, T. (2009). Post-error slowing: An orienting account. Cognition, 111(2), 275–279. doi:10.1016/j.cognition.2009.02.002
Notebaert, W., & Verguts, T. (2011). Conflict and error adaptation in the Simon task. Acta Psychologica, 136(2), 212–216. doi:10.1016/j.actpsy.2010.05.006
Núñez Castellar, E., Kühn, S., Fias, W., & Notebaert, W. (2010). Outcome expectancy and not accuracy determines posterror slowing: ERP support. Cognitive, Affective & Behavioral Neuroscience, 10(2), 270–278. doi:10.3758/CABN.10.2.270
Rey-Mermet, A., Koenig, T., & Meier, B. (2013). The bivalency effect represents an interference-triggered adjustment of cognitive control: An ERP study. Cognitive, Affective, & Behavioral Neuroscience, 13(3), 575–583. doi:10.3758/s13415-013-0160-z
Rey-Mermet, A., & Meier, B. (2012a). The bivalency effect: Adjustment of cognitive control without response set priming. Psychological Research, 76(1), 50–59. doi:10.1007/s00426-011-0322-y
Rey-Mermet, A., & Meier, B. (2012b). The bivalency effect: Evidence for flexible adjustment of cognitive control. Journal of Experimental Psychology: Human Perception and Performance, 38(1), 213–221. doi:10.1037/a0026024
Rey-Mermet, A., & Meier, B. (2013). An orienting response is not enough: Bivalency not infrequency causes the bivalency effect. Advances in Cognitive Psychology, 9(3), 146–155. doi:10.2478/v10053-008-0142-9
Rey-Mermet, A., & Meier, B. (2014). More conflict does not trigger more adjustment of cognitive control for subsequent events: A study of the bivalency effect. Acta Psychologica, 145, 111–117. doi:10.1016/j.actpsy.2013.11.005
Rey-Mermet, A., & Meier, B. (2015). Age affects the adjustment of cognitive control after a conflict: Evidence from the bivalency effect. Aging, Neuropsychology, and Cognition, 22(1), 72–94. doi:10.1080/13825585.2014.889070
Rey-Mermet, A., & Meier, B. (2016). Post-conflict slowing after incongruent stimuli: From general to conflict-specific. Psychological Research. doi:10.1007/s00426-016-0767-0
Simon, J. R., & Small, A. M. (1969). Processing auditory information: Interference from an irrelevant cue. Journal of Applied Psychology, 53(5), 433–435. doi:10.1037/h0028034
Smith, R. E. (2003). The cost of remembering to remember in event-based prospective memory: Investigating the capacity demands of delayed intention performance. Journal of Experimental Psychology: Learning, Memory, and Cognition, 29(3), 347–361. doi:10.1037/0278-7393.29.3.347
Steinhauser, M., & Hübner, R. (2009). Distinguishing response conflict and task conflict in the Stroop task: Evidence from ex-Gaussian distribution analysis. Journal of Experimental Psychology: Human Perception and Performance, 35(5), 1398–1412. doi:10.1037/a0016467
Stroop, J. R. (1935). Studies of interference in serial verbal reactions. Journal of Experimental Psychology, 18(6), 643–662. doi:10.1037/h0054651
Stürmer, B., Leuthold, H., Soetens, E., Schröter, H., & Sommer, W. (2002). Control over location-based response activation in the Simon task: Behavioral and electrophysiological evidence. Journal of Experimental Psychology: Human Perception and Performance, 28(6), 1345–1363. doi:10.1037//0096-1523.28.6.1345
Ullsperger, M., Bylsma, L. M., & Botvinick, M. M. (2005). The conflict adaptation effect: It’s not just priming. Cognitive, Affective, & Behavioral Neuroscience, 5(4), 467–472.
Verguts, T., Notebaert, W., Kunde, W., & Wühr, P. (2011). Post-conflict slowing: Cognitive adaptation after conflict processing. Psychonomic Bulletin & Review, 18(1), 76–82. doi:10.3758/s13423-010-0016-2
Wendt, M., Kluwe, R. H., & Peters, A. (2006). Sequential modulations of interference evoked by processing task-irrelevant stimulus features. Journal of Experimental Psychology: Human Perception and Performance, 32(3), 644–667. doi:10.1037/0096-1523.32.3.644
Woodward, T. S., Meier, B., Tipper, C., & Graf, P. (2003). Bivalency is costly: Bivalent stimuli elicit cautious responding. Experimental Psychology, 50(4), 233.
Woodward, T. S., Metzak, P. D., Meier, B., & Holroyd, C. B. (2008). Anterior cingulate cortex signals the requirement to break inertia when switching tasks: A study of the bivalency effect. NeuroImage, 40(3), 1311–1318. doi:10.1016/j.neuroimage.2007.12.049
Acknowledgements
This work was supported by the Center for Cognition, Learning, and Memory, University of Bern, Switzerland. We thank Lia Äschlimann, Shaila Baumann, Jessica Borer, Salome Bötschi, Clelia Castiglione, Valerie Clausen, Tiziano D’Autilia, Samuel Egli, Danja Greder, Valerie Härry, Maya Holdener, Sadrije Imeri, Olivia Köchli, Cécile Maurer-Kiener, Katja Näf, Kathrin Oegerli, Pascale Raemy, Astrid Rösch, Vanessa Salvatore, Alenka Schmid, Livia Schnider, Lisa Sidler, Lisa-Maria Summermatter, Matthias Smith, Stefanie Suter and Noemi Wimmer for testing the participants. We also thank Michel Druey for helpful comments on an earlier version.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(DOCX 85 kb)
Rights and permissions
About this article

Cite this article
Rey-Mermet, A., Meier, B. How long-lasting is the post-conflict slowing after incongruent trials? Evidence from the Stroop, Simon, and flanker tasks. Atten Percept Psychophys 79, 1945–1967 (2017). https://doi.org/10.3758/s13414-017-1348-z
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-017-1348-z