
Abstract
Encountering a cognitive conflict not only slows current performance, but it can also affect subsequent performance, in particular when the conflict is induced with bivalent stimuli (i.e., stimuli with relevant features for two different tasks) or with incongruent trials (i.e., stimuli with relevant features for two response alternatives). The post-conflict slowing following bivalent stimuli, called “bivalency effect”, affects all subsequent stimuli, irrespective of whether the subsequent stimuli share relevant features with the conflict stimuli. To date, it is unknown whether the conflict induced by incongruent stimuli results in a similar post-conflict slowing. To investigate this, we performed six experiments in which participants switched between two tasks. In one task, incongruent stimuli appeared occasionally; in the other task, stimuli shared no feature with the incongruent trials. The results showed an initial performance slowing that affected all tasks after incongruent trials. On further trials, however, the slowing only affected the task sharing features with the conflict stimuli. Therefore, the post-conflict slowing following incongruent stimuli is first general and then becomes conflict-specific across trials. These findings are discussed within current task switching and cognitive control accounts.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Cognitive control is invoked when facing a conflict (Botvinick, Braver, Barch, Carter, & Cohen, 2001; Egner, 2007). It allows us to activate task-relevant features while inhibiting irrelevant features. In laboratory situations, conflict is typically induced either by incongruent stimuli or by bivalent stimuli. Incongruent stimuli are stimuli with relevant features for two different responses, such as the word “red” printed in blue. Bivalent stimuli are stimuli with relevant features for two different tasks, such as red or blue letters, when participants are asked to switch between a red/blue decision and an uppercase/lowercase decision. Responding to conflict stimuli not only slows performance on these stimuli, but also on subsequent (non-conflict) stimuli (Botvinick et al., 2001; Duthoo, Abrahamse, Braem, Boehler, & Notebaert, 2014; Loft, Kearney, & Remington, 2008; Verguts, Notebaert, Kunde, & Wühr, 2011; Woodward, Meier, Tipper, & Graf, 2003). Interestingly, the post-conflict slowing following bivalent stimuli occurs irrespective of whether the subsequent trials share relevant features with the conflict (see Meier & Rey-Mermet, 2012a, for an overview). Therefore, this post-conflict slowing is not specific to the conflict. To date, the conflict-specificity of the slowing following incongruent stimuli has not yet been investigated. The purpose of the present study was to fill this gap.
Post-conflict slowing following bivalent stimuli has been termed the bivalency effect (Woodward et al., 2003; see Meier & Rey-Mermet, 2012a, for a recent review). The paradigm typically used to investigate the bivalency effect involves three blocks with regular switches between three tasks, such as a parity decision (odd vs. even), a colour decision (red vs. blue), and a case decision (uppercase vs. lowercase). In the first and third blocks, all stimuli are univalent (i.e., black numerals for the parity decision, coloured symbols for the colour decision, and black letters for the case decision). In the second block, some letters for the case decisions are printed in red or blue colour, which turn them into bivalent stimuli. The bivalency effect is the performance slowing that occurs on all univalent trials following bivalent stimuli, including those sharing no features with bivalent stimuli (i.e., the parity-decision trials).
Previous research has shown that the bivalency effect occurs with different types of tasks, different types of bivalent stimuli, across different modalities, as well as with overlapping and non-overlapping response sets (Meier, Woodward, Rey-Mermet, & Graf, 2009; Metzak, Meier, Graf, & Woodward, 2013; Rey-Mermet & Meier, 2012, 2014). It is not affected by a manipulation of the interval between task triplets (i.e., 1000, 2000, 3000, or 5000 ms), and it is still significant after many subsequent univalent trials, up to more than 20 s after the conflict (Meier, Rey-Mermet, & Rothen, 2015; Meier et al., 2009; Rey-Mermet & Meier, 2013). The bivalency effect is also associated with activation in the dorsal anterior cingulate cortex, a brain area recruited for the adjustment of cognitive control, and with an ERP component reflecting interference in cognitive control (Grundy et al., 2013; Rey-Mermet, Koenig, & Meier, 2013; Woodward, Metzak, Meier, & Holroyd, 2008). Furthermore, it draws on memory resources because amnesic patients and older adults fail to show the typical pattern of a long-lasting performance slowing (Meier, Rey-Mermet, Woodward, Müri, & Gutbrod, 2013; Rey-Mermet & Meier, 2015).
To account for the bivalency effect, we have put forward an episodic context binding account (Meier et al., 2013, 2009; Rey-Mermet & Meier, 2015). In line with the adaptation-by-binding account in which the occurrence of a conflict strengthens all memory representations (Verguts & Notebaert, 2008, 2009), we have proposed that responding to a bivalent stimulus results in a memory representation that binds the conflict induced by the bivalent stimulus to its proximate context (e.g., the particular task triplet in the case of the paradigm used to investigate the bivalency effect). Thus, when a bivalent stimulus occurs within a task triplet, the whole context becomes conflict-loaded and thus on subsequent trials, the retrieval of this representation causes interference. As this memory representation includes the whole task triplet, performance is generally slowed for all subsequent univalent trials, irrespective of whether these share relevant features with the bivalent stimulus. Moreover, as the representation is updated each time a task is performed on univalent stimuli, the conflict-loaded memory representation may be attenuated gradually across the series of conflict-free trials. This can explain the reduction of the bivalency effect across univalent trials.
Post-conflict slowing has also been found in prospective memory research (Einstein & McDaniel, 2010; Smith, 2003). Prospective memory refers to the ability to remember to perform a particular task at some designated point in the future (McDaniel & Einstein, 2000). In a typical prospective memory experiment, participants are instructed to execute a particular action when a target event occurs in the course of an ongoing activity. Prospective memory targets induce a conflict because they have relevant features for both the ongoing task and the prospective memory task. In prospective memory research, the focus is on the performance cost when the prospective memory task is introduced compared to when no prospective memory task is introduced (Einstein & McDaniel, 2010; Smith, 2010, 2011). Interestingly, in addition to this monitoring cost, a performance slowing was found after prospective memory targets (Loft et al., 2008). To investigate more specifically the trajectory of this post-conflict slowing, we have included a prospective memory task in the typical bivalency effect paradigm by asking participants to press a different key (i.e., the key “h” instead of “b” or “n”) when encountering a bivalent stimulus (Meier & Rey-Mermet, 2012b). The results showed a performance slowing for the first three trials that immediately followed these targets. This slowing lasted up to 6 s, and it was interpreted as the result of an orienting response. As prospective memory targets occur infrequently, they capture attention. This results in an orienting response, which slows performance on subsequent trials (c.f., Notebaert et al., 2009; Notebaert & Verguts, 2011; Núñez Castellar, Kühn, Fias, & Notebaert, 2010). Ongoing task performance was also slowed on subsequent trials but only for those trials, which shared relevant features with the prospective memory targets. Moreover, this slowing was sporadic because it affected only some of these trials. Together, this indicates that the post-conflict slowing following prospective memory targets becomes more sporadic and conflict-specific across trials (see Meier & Rey-Mermet, 2012b).
In a different line of research, post-conflict slowing has been investigated with incongruent stimuli. Incongruent trials are presented predominantly within three tasks: the Stroop task (MacLeod, 1991; Stroop, 1935), the Simon task (Simon & Small, 1969) and the Flanker task (Eriksen & Eriksen, 1974). In the Stroop task, participants are asked to indicate the colour in which the colour word is displayed while ignoring the meaning of the word (e.g., the word “red” printed in blue, with “blue” as correct response). Stimuli are incongruent when the meaning of the word does not correspond to the print colour; in contrast, they are congruent when the meaning of the word corresponds to the print colour (e.g., the word “red” printed in red). In the Simon task, participants are asked to classify stimuli presented on the left and right side of the screen by pressing left and right keys. Stimuli are incongruent when the stimulus location is different from the response location (e.g., a stimulus presented on the right side requiring a left key press); in contrast, they are congruent when the stimulus location is the same as the response location (e.g., a stimulus presented on the left side, requiring a left key press). In the Flanker task, participants encounter a row of stimuli and they are asked to classify the central stimulus while ignoring the flanking stimuli. Trials are incongruent when the central stimulus is different from the flanking stimulus (e.g., HSH); in contrast, they are congruent when the central stimulus requires the same response as the flanking stimuli (e.g., HHH).
In all three tasks, a congruency effect is observed because participants are slower and more prone to errors on incongruent trials than on congruent trials (Eriksen & Eriksen, 1974; MacLeod, 1991; Simon & Small, 1969; Stroop, 1935). Interestingly, this congruency effect is reduced when incongruent trials are presented more frequently than congruent trials, which results in a proportion congruency effect (Gratton, Coles, & Donchin, 1992; Hommel, 1994; Logan & Zbrodoff, 1979; Lowe & Mitterer, 1982). Moreover, the congruency effect is reduced after incongruent trials compared to after congruent trials (Duthoo, Abrahamse, Braem, Boehler, et al., 2014; Egner, 2007). More precisely, when trial N is incongruent (relative to when it is congruent), performance on trial N + 1 is faster if this trial is also incongruent; in contrast, performance is slower if trial N + 1 is congruent. This congruency sequence effect reflects a combination of two effects: a performance acceleration observed on an incongruent trial following an incongruent trial, and a performance slowing observed on a congruent trial following an incongruent trial.
In most studies (e.g., Akçay & Hazeltine, 2011; Egner, Delano, & Hirsch, 2007; Funes, Lupiáñez, & Humphreys, 2010a; Kunde, Augst, & Kleinsorge, 2012; Kunde & Stöcker, 2002; Mayr, Awh, & Laurey, 2003; Schlaghecken, Refaat, & Maylor, 2011; Spapé, Band, & Hommel, 2011; Verbruggen, Liefooghe, Notebaert, & Vandierendonck, 2005; Wendt, Kluwe, & Peters, 2006), the congruency sequence effect has been found to be conflict-specific (but see Fernandez-Duque & Knight, 2008; Freitas, Bahar, Yang, & Banai, 2007; Kan et al., 2013; Kleiman, Hassin, & Trope, 2014, for some exceptions). When Stroop and Simon tasks are for example combined, a congruency sequence effect occurs within a conflict type (either Stroop or Simon), but not across conflict types. This was interpreted as the result of conflict-specific cognitive control processes (e.g., Egner, 2008; see Braem, Abrahamse, Duthoo, & Notebaert, 2014, for a review of the literature).
Only a few studies have focussed on the post-conflict slowing following incongruent trials (Chang, Chen, Li, & Li, 2014; Verguts et al., 2011). In particular, Verguts et al. (2011) showed a post-conflict slowing after incongruent trials when the subsequent trials are neutral (i.e., involving only one relevant response feature), but not when they are congruent. This pattern was interpreted as the fact that different processes counteract on another in congruent trials. More precisely, the conflict induced by incongruent trials leads to an increased focusing on the relevant features for congruent trials. Thus, the task-focusing process masks the post-conflict slowing, and congruent trials seem not affected by the previously encountered conflict.
Together, these findings indicate that the post-conflict slowing can be conflict-specific (such as with prospective memory targets) or not (such as with bivalent stimuli). Moreover, although the congruency sequence effect was mainly found to be conflict-specific, it is still unknown whether the post-conflict slowing following incongruent stimuli is conflict-specific. The purpose of the present study was to investigate the conflict-specificity of the slowing following incongruent trials. To this end, we conducted six experiments with a paradigm similar to the one that was successfully used to investigate the conflict-specificity of the post-conflict slowing triggered by bivalent stimuli and prospective memory targets. That is, we explored the trajectory of the performance slowing following incongruent stimuli across two different tasks, namely a task whose stimuli share features with the conflict and a task whose stimuli share no features with the conflict.
More precisely, in Experiments 1a and 1b, participants were asked to switch between a digit identification (i.e., a task sharing no features with the conflict stimulus) and a Stroop task (i.e., a task sharing features with the conflict stimulus). For the digit identification, participants were asked to identify the digit, while for the Stroop task they were asked to indicate the ink colour of the colour words. All participants performed three blocks. In the first and third blocks, stimuli were univalent for the digit identification (i.e., black numerals) and congruent for the Stroop task (coloured words printed in their corresponding colours). In the second block, some colour words appeared in a non-corresponding colour (e.g., the word “red” printed in blue), which turn them into incongruent stimuli. In the subsequent experiments, we extended this paradigm by replacing the Stroop task with the Simon task (Experiments 2a and 2b) and the Flanker task (Experiment 3a and 3b).
In addition, we manipulated the interval between sequences of four trials so that the interval was 1000 ms in the Experiments 1a, 2a and 3a and 2000 ms in the Experiments 1b, 2b, and 3b. Previous research has revealed that the post-conflict slowing following bivalent stimuli is the same for both time intervals (see Meier et al., 2009; Rey-Mermet & Meier, 2013). However, the congruency sequence effect diminished across time (Duthoo, Abrahamse, Braem, & Notebaert, 2014; Egner, Ely, & Grinband, 2010). Therefore, it remains an open question whether or not the post-conflict slowing following incongruent trials persists across time.
In each experiment, we thus manipulated three variables within-subject (block, task, and trial sequence). The variable block takes into account the three blocks (block 1, block 2, block 3). The variable task takes into account the task sharing no features with the conflict stimulus (i.e., the digit identification) and the task sharing features with the conflict stimulus (i.e., the colour decision for the Stroop and Simon conflict types, and the letter decision for the Flanker conflict). The variable trial sequence takes into account the number of trials following an incongruent stimulus (i.e., N + 1, N + 2, N + 3, and N + 4, with N referring to the trials containing an incongruent stimulus). For the sake of parsimony, we present the analyses across all experiments by adding conflict type (Stroop, Simon, Flanker) and interval (1000, 2000 ms) as additional (between-subjects) variables.
We hypothesized that if the post-conflict slowing following incongruent trials is not conflict-specific and thus similar to the post-conflict slowing following bivalent stimuli (Woodward et al., 2003; see Meier & Rey-Mermet, 2012a, for a review), performance after incongruent trials would be slowed for both tasks. Thus, in this case, similar to the bivalency effect, the post-conflict slowing following incongruent trials would result from episodic context binding (Meier et al., 2013, 2009; Rey-Mermet & Meier, 2015). Alternatively, if the post-conflict slowing following incongruent stimuli is specific to the conflict (Braem et al., 2014; Meier & Rey-Mermet, 2012b), performance after incongruent trials would be slowed for the task sharing features with the conflict stimulus (i.e., colour/letter decisions), but not for the task sharing no features with the conflict stimulus (i.e., digit identification). In this case, it is also possible that no post-conflict slowing occurs because the post-conflict slowing and the task-focusing process may counteract on congruent trials (Verguts et al., 2011).
General method
Participants
Participants were 156 volunteers (26 in each experiment) from the University of Bern. We replaced three participants because their accuracy level for incongruent trials was less than 50 %. Demographic characteristics of the sample are described in the left part of Table 1. The study was approved by the local ethical committee of the University of Bern, and all participants gave written consent.
Materials
All stimuli were presented on a grey background in 60-point Times New Roman. In all experiments, the task sharing no features with the conflict stimulus was a digit identification on the numerals 1, 2, 3, and 4. These stimuli were displayed in black at the center of the screen. Participants used the response keys “v”, “b”, “n”, “m” in Experiments 1a, 1b, 3a and 3b, and the response keys “g”, “b”, “n”, “j” in Experiments 2a and 2b, with their left and right index and middle fingers. These keys were mapped to the digits 1, 2, 3, and 4, respectively. The task sharing features with the conflict stimulus differed across experiments and is described in the following sections. An overview of the differences between the experiments is presented in the right part of Table 1.
Experiments 1a and 1b (Stroop conflict)
The task sharing relevant features with the conflict stimulus was a colour decision on colour words. The stimuli were displayed at the center of the computer screen. Stimuli were congruent when the four German words blue, red, green, and yellow (i.e., “blau”, “rot”, “grün”, and “gelb”) were displayed in blue, red, green, and yellow, respectively. Stimuli were incongruent when the colour words were printed in a non-corresponding colour (e.g., the colour word “red” printed in blue). For each participant, six incongruent stimuli were determined randomly and without replacement. The response keys (i.e., “v”, “b”, “n”, “m”) were mapped to the colours blue, red, green, and yellow, respectively.
Experiments 2a and 2b (Simon conflict)
The task sharing relevant features with the conflict stimulus was a colour decision on the symbols $, #, §, %, displayed either in blue, red, green, or yellow. These stimuli were presented in one of the four corners of a centred, non-displayed 4 cm × 4 cm square. The response keys (i.e., “g”, “b”, “n”, “j”) were mapped to the colours blue, red, green, and yellow, respectively. Stimuli were congruent when the position of the symbol on the computer screen corresponded to the location of the response key. In contrast, they were incongruent when the position of the symbol on the computer screen did not correspond to the location of the response key required by the colour decision. For each participant, six incongruent stimuli were determined randomly and without replacement.
Experiments 3a and 3b (Flanker conflict)
The task sharing relevant features with the conflict stimulus was a letter decision on letter triplicates. Stimuli were displayed in black at the center of the computer screen. The congruent stimuli were the four letters H, P, R, S, displayed as triplicates with the same letters (e.g., HHH). Incongruent stimuli were the same four letters, but the central letter was different from the flanking letters (e.g., SHS). For each participant, six incongruent stimuli were determined randomly and without replacement. The response keys (i.e., “v”, “b”, “n”, “m”) were mapped to the letters H, P, R, and S, respectively.
Procedure
In each experiment, participants were tested individually. They were informed to switch between two tasks: a digit identification task on numerals and either a colour decision on colour words for the Stroop conflict (Experiments 1a and 1b), a colour decision on coloured symbols for the Simon conflict (Experiments 2a and 2b), or a letter decision on letter triplicates for the Flanker conflict (Experiments 3a and 3b). Participants were instructed to press one of the four computer keys with their index and middle fingers of their left and right hands for each of the two tasks. The mapping information, printed on paper, was presented below the computer screen throughout the experiment. For the Stroop conflict, participants were informed that occasionally, the colour word would not correspond to the colour in which it was printed (e.g., the word “red” printed in blue), and that they had to proceed as usual by responding to the colour of the colour word. For the Simon conflict, they were informed that occasionally, the position of the stimulus would not correspond to the location of the response key (e.g., a red symbol requiring a lower left key press, but presented on the upper right corner), and that they had to proceed as usual by responding to the colour of the symbol. For the Flanker conflict, they were informed that occasionally, the central letter would not correspond to the flanking letters (e.g., HSH), and that they had to proceed as usual by responding to the central letter.
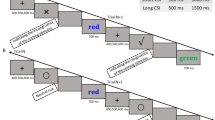
After the instructions, a block of 30 trial sequences was presented for practice. Each trial sequence included two digit identification tasks and two colour/letter decisions, as illustrated in Fig. 1. Within the trial sequence, the first task of each pair was a switch trial and the second task of each pair was a repetition trial. Within each pair, the particular stimulus was determined randomly and did not repeat. Task order followed an AABB design (i.e., two digit identification tasks were followed by two colour/letter decisions, which, in turn, were followed by two digit identification tasks, and so on). The stimulus for each trial was displayed until the participant responded. Then, the screen blanked for 500 ms before the next stimulus appeared. After each trial sequence, an additional blank interval was included so that the screen blanked either for 1000 ms (Experiments 1a, 2a, and 3a) or for 2000 ms (Experiments 1b, 2b, and 3b). After the practice block and a brief break, each participant completed three experimental blocks without break between blocks. The first block included 32 trial sequences, with the first two trial sequences serving as “warm-up” sequences which were discarded from the analyses. The second and third blocks had 30 trial sequences each.

Example of one trial sequence with 1000 ms as interval. a Stroop conflict. b Simon conflict. c Flanker conflict. On each trial sequence, participants carried out two digit identifications (task sharing no relevant features with the conflict stimulus), followed by two colour/letter decisions (task sharing relevant features with the conflict stimulus). On an incongruent trial sequence (not pictured here), the fourth trial involved a colour word printed in a non-corresponding colour for the Stroop conflict, a symbol whose position on the computer screen did not correspond to the location of the response key for the Simon conflict or a letter triplicate in which the central letter was different from the flanking letters for the Flanker conflict
In the first and third blocks, all stimuli were univalent for the digit identifications and congruent for the colour/letter decisions. In the second block, all stimuli were also univalent for the digit identifications. In contrast, for the colour/letter decisions, stimuli were congruent except for six trials in which they were incongruent. Incongruent stimuli were evenly interspersed among the 30 trial sequences of the block, thus occurring in every fifth trial sequence (specifically, the 3rd, 8th, 13th, 18th, 23rd, and 28th sequences contained an incongruent stimulus). Incongruent stimuli always appeared on the fourth trial of this trial sequence. The entire experiment lasted about 15 min.
Data preparation and analysis
For each participant in each experiment, the accuracy and the median reaction times (RTs) for correct responses were computed for each trial type (switch and repetition), each task (digit identification, colour/letter decision), and each trial sequence following an incongruent stimulus in block 2. The same was then applied for each corresponding trial sequence in the blocks 1 and 3. Specifically, the trial sequence containing an incongruent stimulus in block 2 was designated as trial sequence N. The successive trial sequences were then labelled as N + 1, N + 2, N + 3, and N + 4. Mean accuracies and median RTs were then averaged across switch and repetition trials for each task, each trial sequence, each block, and each participant.Footnote 1
We used these mean RTs for each participant to test for differences between the three blocks. To account for baseline RT differences between Stroop, Simon and Flanker conflict types, we also computed proportional scores. That is, for each trial sequence (i.e., N + 1, N + 2, N + 3, and N + 4), mean RT for block 2 was subtracted from the mean RT averaged across blocks 1 and 3; then, this difference was divided by the mean RT averaged across all blocks. Statistical analyses were performed on both dependent measures (i.e., mean RTs and proportional scores).
We first investigated the congruency effect by comparing performance on incongruent trials from block 2 with performance on the corresponding congruent trials from blocks 1 and 3 (i.e., those trials occurring on the same trial position as incongruent trials but in the blocks 1 and 3). To this end, we analysed the RTs with a three-way analysis of variance (ANOVA), with congruency (congruent-block 1, incongruent-block 2, congruent-block 3) as a within-subject factor, and conflict type (Stroop, Simon, Flanker) and interval (1000, 2000 ms) as between-subjects factors. For proportional scores, the equivalent analysis was a two-way ANOVA with the between-subjects factors conflict type (Stroop, Simon, Flanker) and interval (1000, 2000 ms).
The main objective of the present study was to determine the persistence of the impact of incongruent trials on tasks sharing relevant features with the conflict and on tasks sharing no features with the conflict. For RTs, we carried out a five-way ANOVA with block (block 1, block 2, block 3), task (digit identification, colour/letter decision) and trial sequence (N + 1, N + 2, N + 3, N + 4) as within-subject factors, and conflict type (Stroop, Simon, Flanker) and interval (1000 ms, 2000 ms) as between-subjects factors. On average, 9.8 valid trials per cell (range = 5–10) were available. We then disentangled the critical interactions by performing the follow-up ANOVAs. We also conducted follow-up one-way repeated-measures ANOVAs with the factor block (block 1, block 2, block 3) to assess more specifically performance difference between the different blocks. In these one-way ANOVAs, we focused on the quadratic component of the block effect because this is informative about a difference in block 2 compared to blocks 1 and 3. For the proportional scores, the equivalent analysis was a four-way ANOVA with task (digit identification, colour/letter decision) and trial sequence (N + 1, N + 2, N + 3, N + 4) as within-subject factors, and conflict type (Stroop, Simon, Flanker) and interval (1000, 2000 ms) as between-subjects factors. Again, the critical interactions were then disentangled by follow-up ANOVAs and one-sample t tests.
Means and standard errors for each experiment are presented in the “Appendix”.Footnote 2 As our focus was mainly on RTs, proportional scores results are referred to only when diverging from RTs results. An alpha level of 0.05 was used for all statistical tests. Greenhouse-Geisser corrections are reported where appropriate and effect sizes are expressed as partial η 2 values.
Results
Congruency effect
For the congruency effect, the results of the ANOVAs are presented in Table 2, and the descriptive results are shown in Table 3. Critically, the RT analysis revealed a significant main effect of block, with a significant quadratic component, F(1, 150) = 334.09, p < 0.001, η 2 = 0.69. Thus, as expected, performance was slower on incongruent trials than on the corresponding congruent trials, which confirms the presence of the congruency effect (see Table 3). Moreover, the interaction between block and conflict type was significant. This results from a larger congruency effect in the Stroop conflict than in the Simon and Flanker conflict types (see Tables 2, 3). Importantly, however, the congruency effect was significant in all conflict types (Stroop: quadratic component of the block effect for RTs: F(1, 51) = 110, p < 0.001, η 2 = 0.68 and one sample t test for proportional scores: t(51) = 15.39, p < 0.001; Simon: quadratic component of the block effect for RTs: F(1, 51) = 132.08, p < 0.001, η 2 = 0.72 and one sample t test for proportional scores: t(51) = 13.83, p < 0.001; and Flanker: quadratic component of the block effect for RTs: F(1, 51) = 163.72, p < 0.001, η 2 = 0.76 and one sample t test for proportional scores: t(51) = 14.50, p < 0.001).
For accuracy, the three-way ANOVA revealed a significant main effect of congruency, with a significant quadratic component, F(1, 150) = 35.64, p < 0.001, η 2 = 0.19 (see Table 2). Thus, responses were less accurate on incongruent trials than on the corresponding congruent trials. This confirms the presence of the congruency effect in accuracy rates (see Table 3).
Impact of incongruent trials on subsequent univalent and congruent trials
Our main objective was to examine whether the performance slowing following incongruent trials persists across tasks (i.e., digit identification as well as colour/letter decisions). The most relevant results are thus the RTs from both tasks in blocks 1 and 3 compared to those in block 2. The descriptive results are depicted in Fig. 2.

Mean reaction times on the digit identifications and the colour/letter decisions from block 1 (empty squares), block 2 (filled circles), and block 3 (empty diamonds). Trial sequence N refers to the trial sequence containing an incongruent trial. Subsequent trial sequences (represented here) are labeled N + 1, N + 2, N + 3, and N + 4, respectively. Error bars represent standard errors. a Stroop conflict. b Simon conflict. c Flanker conflict
The results of the ANOVAs are shown in Table 4. The most relevant interactions are those involving block. Table 4 reveals three interactions of interest: the interaction between block and task, between block and trial sequence, and between block, task and trial sequence. To disentangle these interactions, we carried out follow-up two-way repeated-measures ANOVAs with the factors block (block 1, block 2, block 3) and task (digit identification, colour/letter decision) for each trial sequence. The statistical values of this analysis as well as those of the corresponding quadratic components of the block effect are summarized in Table 5. This reveals that the performance slowing was larger for the digit identifications than for the colour/letter decisions in the trial sequence N + 1, similar for both tasks in N + 2, and smaller for the digit identifications than for the colour/letter decisions in N + 3. In the last trial sequence (i.e., N + 4), the performance slowing was not significant for the digit identifications but still significant for the colour/letter decisions (see Fig. 2).
Furthermore, the interaction between block and interval was significant (see Table 4). Thus, although performance after incongruent trials was significantly slowed in both intervals [1000 ms: quadratic component of the block effect for RTs: F(1, 77) = 58.92, p < 0.001, η 2 = 0.43 and one sample t test for proportional scores: t(77) = 7.82, p < 0.001; as well as 2000 ms: quadratic component of the block effect for RTs F(1, 77) = 51.16, p < 0.001, η 2 = 0.40 and one sample t test for proportional scores: t(77) = 5.91, p < 0.001], performance was more slowed in the 1000 ms interval than in the 2000 ms interval. Thus, the post-conflict slowing was larger in the 1000 ms interval than in the 2000 interval, but its trajectory across trials and tasks did not differ for both intervals.
Finally, the interactions between block and conflict type as well as between block, task and conflict type were also significant (see Table 4). However, the equivalent effects—that is, the main effect of conflict type and the interaction between task and conflict type—were not significant when proportional scores were considered. This suggests that these significant interactions in RTs arose because of baseline RT differences between Stroop, Simon and Flanker conflict types. Together, this indicates that the trajectory of the post-conflict slowing was similar across conflict types.
General discussion
The purpose of the present study was to determine whether the post-conflict slowing following incongruent trials is conflict-specific so that it only affects the task sharing features with the conflict stimulus. To this end, we occasionally presented incongruent stimuli while participants switched between two tasks. One task was a digit identification and thus its stimuli shared no features with the conflict induced by incongruent stimuli. The other task included incongruent stimuli and thus its stimuli shared relevant features with the conflict. More specifically, this task was a colour decision on colour words in Experiments 1a and 1b (Stroop conflict), a colour decision on coloured symbols in Experiments 2a and 2b (Simon conflict), and a letter decision on letter triplicates in Experiments 3a and 3b (Flanker conflict). The goal was then to determine the trajectory of the performance slowing following incongruent trials across the task sharing relevant features with the conflict stimulus and the task sharing no features with the conflict stimulus.
The results showed a performance slowing on both tasks up to the third trial sequence after incongruent stimuli. In this trial sequence, however, the slowing was larger on colour/letter decisions than on digit identifications. This pattern was even more pronounced on the last trial sequence (i.e., N + 4) in which performance was still significantly slowed in the colour/letter decisions, but not anymore in the digit identifications. Together, this indicates that the conflict induced by incongruent trials results in a post-conflict slowing that is not conflict-specific initially. However, across subsequent trials, it becomes more conflict-specific.
First, it is interesting to note that we found a post-conflict slowing on congruent trials. Therefore, a task-focusing process does not always mask the post-conflict slowing following incongruent trials (Verguts et al., 2011). Another interesting point is that the post-conflict slowing occurs on several univalent trials, which indicates that it is not conflict-specific on several trials following incongruent trials. This contrasts with studies showing a conflict-specific congruency sequence effect (e.g., Akçay & Hazeltine, 2011; Egner, 2007; Kunde et al., 2012; Kunde & Stöcker, 2002; Mayr et al., 2003; Schlaghecken et al., 2011; Spapé et al., 2011; Verbruggen et al., 2005; Wendt et al., 2006; see also Egner, 2008). Rather, these results are in line with those studies in which the congruency sequence effect was found to generalize across conflict types (e.g., Fernandez-Duque & Knight, 2008; Freitas et al., 2007; Kan et al., 2013; Kleiman et al., 2014). Moreover, the long-lasting effect observed in the present study extends those studies because their focus was on the immediate subsequent trial only.
Can the post-conflict slowing following incongruent stimuli be explained with current task switching and cognitive control accounts?
Explanations from task switching and cognitive control research are based either on transient cognitive processes, which are assumed to be implemented trial by trial, or on sustained processes, which are assumed to affect performance across a series of several trials (see Braver, 2012; Braver, Gray, & Burgess, 2008). In the subsequent sections, we present these different explanations and we determine whether these explanations can account for the findings observed in the present study.
Transient cognitive processes
To account for the post-conflict slowing observed in the present study, one may put forward different transient processes, such as task-set inertia, task-set reconfiguration, conflict adaptation, or stimulus-task binding (Botvinick et al., 2001; Egner et al., 2007; Kiesel et al., 2010; Vandierendonck, Liefooghe, & Verbruggen, 2010; Waszak, Hommel, & Allport, 2003). For example, according to a task-set inertia account (Allport & Wylie, 1999, 2000; Wylie & Allport, 2000), when an incongruent stimulus occurs on a given trial, the task-set for the task-relevant feature (e.g., the colour of the colour word in the Stroop task) is activated while the task-set for the task-irrelevant feature is inhibited (i.e., the meaning of the word). This inhibition process is assumed to remain constant across the subsequent trials. Therefore, a task-set inertia process can explain the slowing on subsequent congruent trials because on these trials, the task-set for the task-irrelevant feature (i.e., the meaning of the word) can be further inhibited (even if it is not necessary anymore). However, this account cannot explain the slowing on univalent trials because these trials share no relevant or irrelevant features with the incongruent trials.
According to a task-reconfiguration account (e.g., Goschke, 2000; Rogers & Monsell, 1995; Sohn & Carlson, 2000), when incongruent stimuli are processed, an additional decision is required to determine the relevant task-set. Therefore, in the present study, congruent trials can activate this additional task-decision process because they share features with incongruent trials. In contrast, for univalent trials sharing no relevant trials with incongruent trials, no additional, time-consuming task-decision process would be required. Thus, this account can explain the slowing on congruent trials, but not on univalent trials.
Moreover, task-set inertia and task-set reconfiguration are assumed to be involved in switch costs (i.e., the slowing occurring on switch trials compared to repetition trials) and restart costs (i.e., the slowing occurring in the first trial in a run compared to later trials of the run). However, even if these costs—in particular switch costs on either univalent or bivalent stimuli—were found to be affected by the presence of incongruent trials (Braem, Verguts, Roggeman, & Notebaert, 2012; Goschke, 2000), both costs have been found to last only for one subsequent trial (e.g., Allport & Wylie, 2000). It follows that if a task-set inertia or reconfiguration process occurred in the present study, it could explain the slowing on the first trial immediately following incongruent trials, but not the slowing on later (congruent and univalent) trials.
According to the conflict adaptation framework (Botvinick et al., 2001; Egner, 2007), processing task-relevant and -irrelevant features depends on whether the previous trial is congruent or incongruent. That is, when two congruent trials are presented consecutively, the processing of task-relevant and -irrelevant features (i.e., the colour and the meaning of the word in the Stroop task) are enhanced because both features are associated to the correct response. This results in fast performance. In contrast, when a congruent trial follows an incongruent trial, responding to the incongruent trial reduces the processing of task-irrelevant features and thus only the processing of task-relevant feature (i.e., the colour) is enhanced on the subsequent congruent trial. Therefore, performance on congruent trials is slower after incongruent trials than after congruent trials, which results in the post-conflict slowing. This account can explain the slowing on congruent trials because task-relevant and task-irrelevant features are similar across congruent and incongruent trials. However, it cannot explain the slowing on univalent trials in which task-relevant features (i.e., the numerical value of the digit) are different from the features of incongruent trials (namely, the colour and the meaning of the colour word for the Stroop conflict type, for example).
In a stimulus-task binding account (Waszak et al., 2003; Waszak, Hommel, & Allport, 2004), when a stimulus is presented within task, the stimulus features are bound to the task, and this stimulus-task binding can affect performance even after more than 100 intervening trials. Accordingly, when an incongruent stimulus is presented in the present study, the relevant and irrelevant features of the incongruent trials are bound to the task in which the incongruent stimulus is presented (i.e., the colour decision for the Stroop conflict type, for example). This stimulus-task binding can affect performance on this task for several trials, which may explain the performance slowing on the task sharing relevant features with the incongruent trials, as well as the long-lasting nature of this slowing. However, it cannot explain the slowing on the univalent trials because these stimuli share no features with the incongruent trials and thus are not bound to the task in which incongruent trials are presented.
Sustained cognitive processes
It is possible that the post-conflict slowing results from more sustained cognitive processes, such as a shift in response criterion (e.g., Woodward et al., 2003). According to such an explanation, participants would adopt a more cautious response style, which would result in a similar post-conflict slowing for univalent and congruent trials. However, the present results showed a larger slowing for congruent trials than for univalent trials (see Fig. 2), ruling out this account.
A further explanation can be derived from accounts explaining the proportion congruency effect (Lindsay & Jacoby, 1994; Logan & Zbrodoff, 1979; Lowe & Mitterer, 1982; West & Baylis, 1998). In these accounts, a high proportion of congruent trials (and thus a low proportion of incongruent trials) biases the cognitive system so that the processing of task-relevant features is enhanced. This leads to fast responses on congruent trials and slow responses to incongruent trials, which results in a large congruency effect. In contrast, a low proportion of congruent trials (and thus a high proportion of incongruent trials) biases the cognitive system so that the processing of task-irrelevant features is reduced. Therefore, in comparison with a high proportion of congruent trials, responses to congruent trials are slower while responses to incongruent trials are faster, and the congruency effect is thus smaller. In the present study, we also manipulated the proportion of congruent trials so that in the first and third blocks, 50 % of the trials were congruent and 50 % univalent, while in the second block, 40 % of the trials were congruent, 10 % incongruent and 50 % univalent. Therefore, it is possible that the processing of task-relevant features (i.e., the colour of colour word in the Stroop task) is particularly enhanced in the first and third blocks, which results in fast responses for congruent trials. In contrast, in the second block, the processing of task-relevant features is reduced because of the occasional occurrence of incongruent stimuli. Thus, performance on congruent trials would be slower after incongruent trials than after congruent trials, which explains the post-conflict slowing on congruent trials. However, this does not explain why performance is slowed on univalent trials because univalent trials have different task-relevant features (i.e., the numerical value of the digit) than congruent trials (i.e., the colour of the colour word).
It is noteworthy that similar to the congruency sequence effect, the proportion congruency effect was found to be conflict-specific in most studies (e.g., Crump, Gong, & Milliken, 2006; Crump, Vaquero, & Milliken, 2008; Fernandez-Duque & Knight, 2008; Jacoby, Lindsay, & Hessels, 2003). Nevertheless, when it was found to generalize across conflict types (see Funes, Lupiáñez, & Humphreys, 2010b), this was explained by assuming that the task-relevant features were so similar across conflict types that when the processing of task-relevant features was enhanced in one conflict type, it was also enhanced in the other conflict type. However, this cannot account for the post-conflict slowing observed in the present study, because task-relevant features for univalent and incongruent/congruent trials are dissimilar (i.e., the numerical value of the digit, and the colour of the colour word for the Stroop task, the colour of the symbol for the Simon task, and the central letter of the letter row for the Flanker task, respectively).
Finally, according to a contingency learning account (e.g., Schmidt, 2013a; Schmidt & De Houwer, 2011), when a stimulus and a response co-occur in time, then stimulus and response features are integrated into a common episodic memory representation, and this episodic memory representation is assumed to be strengthened by each new stimulus presentation. Therefore, as the congruent stimuli appeared in the present study less frequently in block 2 than in blocks 1 and 3 (50 vs. 40 %, respectively), their episodic memory representations could have been less strengthened in block 2 than in blocks 1 and 3, which could explain the slowing on congruent trials in block 2. Thus, contingency learning can account for the post-conflict slowing on congruent trials. However, it cannot account for the post-conflict slowing observed on univalent trials as these trials were presented in 50 % of the trials in all three blocks.
Together, this emphasizes that the post-conflict slowing following incongruent trials can only be explained to some extent by the current task switching and cognitive control accounts (e.g., Allport & Wylie, 2000; Botvinick et al., 2001; Egner, 2007; Funes et al., 2010b; Goschke, 2000; Logan & Zbrodoff, 1979; Schmidt, 2013b; Woodward et al., 2003). However, these accounts were put forward to explain the impact of incongruent trials within switch costs, restart cots, congruency sequence effects or proportion congruency effects, but not to explain the post-conflict slowing following incongruent trials. Therefore, it is possible that the accounts used to explain the post-conflict slowing following bivalent stimuli or prospective memory targets can be useful to explain the post-conflict slowing following incongruent trials.
Similar or dissimilar to the post-conflict slowing following other conflict stimuli?
Post-conflict slowing following prospective memory targets
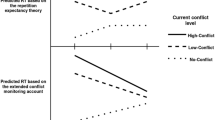
At first sight, the post-conflict slowing following incongruent trials may seem similar to the post-conflict slowing following prospective memory targets (Meier & Rey-Mermet, 2012b). After both incongruent trials and prospective memory targets, the slowing is first general and then becomes more conflict-specific across trials. Therefore, in both cases, the slowing observed on the first few trials could be explained with an orienting response account (Notebaert et al., 2009; Notebaert & Verguts, 2011; Núñez Castellar et al., 2010; Rey-Mermet & Meier, 2013). In the case of incongruent trials, this suggests that these trials capture attention because they occur infrequently, and the orienting response persists across a few subsequent trials, irrespective of whether the trials share relevant or irrelevant features with the conflict. As the impact of the orienting response is rather short-lived (about six trials; see Rey-Mermet & Meier, 2013), this process may explain the slowing on incongruent trialsFootnote 3 and on the first few trials following incongruent trials, but it may not explain the slowing on later trials.
However, there is a crucial difference between the post-conflict slowing following incongruent trials and prospective memory targets. That is, on later trials, the post-conflict slowing following incongruent trials occurs on most trials, whereas the post-conflict slowing following prospective memory targets is sporadic, affecting only some of the trials. Thus, in contrast to prospective memory targets, the post-conflict slowing following incongruent trials cannot be explained by an expectancy-based monitoring process (Meier & Rey-Mermet, 2012b; Meier, Zimmermann, & Perrig, 2006).
The post-conflict slowing following bivalent stimuli (i.e., the bivalency effect)
Apparently, the trajectory of the post-conflict slowing following incongruent stimuli and the trajectory following bivalent stimuli seem different. While the bivalency effect is long-lasting and general (e.g., Meier et al., 2009), the post-conflict slowing following incongruent trials is also long-lasting, but it becomes more conflict-specific across trials. However, the trajectory of the bivalency effect was assessed up to 12 trials (Meier et al., 2013, 2009; Rey-Mermet & Meier, 2013, 2015), whereas the trajectory of the post-conflict slowing following incongruent trials was assessed in the present study up to 16 trials. When only the first 12 trials are considered, the post-conflict slowing following incongruent trials is as general as the bivalency effect. That is, performance was slowed for both tasks, the task sharing relevant features with incongruent trials as well as the task sharing no relevant features with incongruent trials.
Similar to the bivalency effect, the post-conflict slowing may therefore be explained with an episodic context binding account (Meier et al., 2013, 2009; Rey-Mermet & Meier, 2015). It is possible that responding to the conflict induced by an incongruent stimulus results in a memory representation which binds the conflict with its proximate context (i.e., the task in which the conflict occurs and the task with univalent stimuli). Thus, a combined representation of the task sharing relevant features with the conflict stimuli and the task sharing no relevant features with the conflict stimuli is formed when a conflict is processed. On subsequent trials, the retrieval of this conflict-loaded representation causes interference and slows down performance. Critically, as the memory representation includes both tasks, performance is slowed for all trials, which results in the general post-conflict slowing observed after incongruent trials. However, as a new, conflict-unloaded memory representation is retrieved and updated each time a task is performed on a congruent or a univalent trial, these new memory representations may gradually overwrite the conflict-loaded memory representation across the series of conflict-free trials. This could explain the reduction of the post-conflict slowing across univalent and congruent trials. The present findings suggest that the conflict-loaded memory representation is no more effective after 12 trials. Moreover, to explain the specific slowing on the last trial sequence (N + 4) for the task whose stimuli shared the relevant features with the incongruent trials, one may assume that the stimulus-task binding occurring on the task in which incongruent trials are presented is longer-lasting than the episodic context binding. Thus, this stimulus-task binding still interferes with processing of the latest trials, which could explain this late specific slowing.
Together, the present results showed that the post-conflict slowing following incongruent trials was more general and longer-lasting than previously thought. This can be explained with the episodic context binding account, although this explanation is so far somewhat speculative. Therefore, further research would be necessary to determine exactly which cognitive processes are responsible for the pattern of results found in the present study. To this end, it would be interesting, for example, to test the post-conflict slowing following all three conflict stimuli—that is, incongruent stimuli, bivalent stimuli, and prospective memory targets—in the same experiment under the same exact experimental conditions. It would be also interesting to determine the impact of the instructions on the post-conflict slowing following bivalent and incongruent stimuli as the instructions seem critical for the post-conflict slowing following prospective memory targets. More generally, the present findings also suggest that to determine the conflict-specificity of cognitive processes, it would be interesting to disentangle the congruency sequence effect into its two effects (i.e., the performance acceleration on incongruent trials following incongruent trials and the slowing on congruent trials following incongruent trials) and to consider the proportion congruency effect in a trial-by-trial analysis.
Conclusion
To summarize, the findings of the present study show that the conflict induced by incongruent trials results in a post-conflict slowing which can persist across trials sharing features with the conflict as well as sharing no relevant features with the conflict. More specifically, the post-conflict slowing changes across trials so that it is first general and then becomes more conflict-specific. This effect occurred for the Stroop, the Simon as well as the Flanker task, demonstrating that the post-conflict slowing is a robust phenomenon. More generally, the present results highlight that different types of conflict (incongruent trials, bivalent stimuli or prospective memory targets) result in different trajectories of post-conflict slowing.
Notes
In the present study, participants were instructed to perform each task twice in succession (see Fig. 1). This results in switch and repetition trials which allows to examine whether responding to incongruent stimuli affects switch and repetition trials differentially, and thus contributes to switch costs (i.e., the slower performance on switch compared to repetition trials). We carried out all the analyses including switch vs. repetition as an additional independent variable. Overall, this resulted in the same pattern of findings.
In the analyses of the impact of incongruent trials on subsequent congruent trials, we focussed on reaction times data because accuracy on univalent and congruent trials was close to ceiling (mean accuracy 98 %) and even at ceiling in some conditions (see “Appendix”).
This may explain why the congruency effects were so large in the present study. It is possible that participants were slower on incongruent trials than on congruent trials not only because incongruent trials induced a conflict between two response alternatives but also because they induced an orienting response due to their infrequent occurrence.
References
Akçay, Ç., & Hazeltine, E. (2011). Domain-specific conflict adaptation without feature repetitions. Psychonomic Bulletin & Review, 18(3), 505–511. doi:10.3758/s13423-011-0084-y.
Allport, A., & Wylie, G. (1999). Task-switching: Positive and negative priming of task-set. In G. W. Humphrey, J. Duncan, & A. Treisman (Eds.), Attention, space and action: Studies in cognitive neuroscience (pp. 273–296). New York: Oxford University Press.
Allport, A., & Wylie, G. (2000). Task-switching, stimulus-response bindings, and negative priming. In S. Monsell & J. S. Driver (Eds.), Control of Cognitive Processes: Attention and Performance XVIII (pp. 35–70). Cambridge: MIT Press.
Botvinick, M. M., Braver, T. S., Barch, D. M., Carter, C. S., & Cohen, J. D. (2001). Conflict monitoring and cognitive control. Psychological Review, 108(3), 624–652. doi:10.1037//0033-295X.108.3.624.
Braem, S., Abrahamse, E. L., Duthoo, W., & Notebaert, W. (2014). What determines the specificity of conflict adaptation? A review, critical analysis, and proposed synthesis. Frontiers in Psychology,. doi:10.3389/fpsyg.2014.01134.
Braem, S., Verguts, T., Roggeman, C., & Notebaert, W. (2012). Reward modulates adaptations to conflict. Cognition, 125(2), 324–332. doi:10.1016/j.cognition.2012.07.015.
Braver, T. S. (2012). The variable nature of cognitive control: A dual mechanisms framework. Trends in Cognitive Sciences, 16(2), 106–113. doi:10.1016/j.tics.2011.12.010.
Braver, T. S., Gray, J. R., & Burgess, G. C. (2008). Explaining the many varieties of working memory variation: Dual mechanisms of cognitive control. In A. Conway, C. Jarrold, M. Kane, A. Miyake, & J. Towse (Eds.), Variation in Working Memory (pp. 76–106). Oxford University Press.
Chang, A., Chen, C.-C., Li, H.-H., & Li, C.-S. R. (2014). Event-related potentials for post-error and post-conflict slowing. PLoS One, 9(6), e99909. doi:10.1371/journal.pone.0099909.
Crump, M. J. C., Gong, Z., & Milliken, B. (2006). The context-specific proportion congruent Stroop effect: Location as a contextual cue. Psychonomic Bulletin & Review, 13(2), 316–321.
Crump, M. J. C., Vaquero, J. M. M., & Milliken, B. (2008). Context-specific learning and control: The roles of awareness, task relevance, and relative salience. Consciousness and Cognition, 17(1), 22–36. doi:10.1016/j.concog.2007.01.004.
Duthoo, W., Abrahamse, E. L., Braem, S., Boehler, C. N., & Notebaert, W. (2014a). The heterogeneous world of congruency sequence effects: An update. Frontiers in Psychology,. doi:10.3389/fpsyg.2014.01001.
Duthoo, W., Abrahamse, E. L., Braem, S., & Notebaert, W. (2014b). Going, going, gone? Proactive control prevents the congruency sequence effect from rapid decay. Psychological Research, 78(4), 483–493. doi:10.1007/s00426-013-0498-4.
Egner, T. (2007). Congruency sequence effects and cognitive control. Cognitive, Affective, & Behavioral Neuroscience, 7(4), 380–390. doi:10.3758/CABN.7.4.380.
Egner, T. (2008). Multiple conflict-driven control mechanisms in the human brain. Trends in Cognitive Sciences, 12(10), 374–380. doi:10.1016/j.tics.2008.07.001.
Egner, T., Delano, M., & Hirsch, J. (2007). Separate conflict-specific cognitive control mechanisms in the human brain. NeuroImage, 35(2), 940–948. doi:10.1016/j.neuroimage.2006.11.061.
Egner, T., Ely, S., & Grinband, J. (2010). Going, going, gone: characterizing the time-course of congruency sequence effects. Cognition, 1, 154. doi:10.3389/fpsyg.2010.00154.
Einstein, G. O., & McDaniel, M. A. (2010). Prospective memory and what costs do not reveal about retrieval processes: A commentary on Smith, Hunt, McVay, and McConnell (2007). Journal of Experimental Psychology: Learning, Memory, and Cognition, 36(4), 1082–1088. doi:10.1037/a0019184.
Eriksen, B. A., & Eriksen, C. W. (1974). Effects of noise letters upon the identification of a target letter in a nonsearch task. Perception & Psychophysics, 16(1), 143–149. doi:10.3758/BF03203267.
Fernandez-Duque, D., & Knight, M. (2008). Cognitive control: Dynamic, sustained, and voluntary influences. Journal of Experimental Psychology: Human Perception and Performance, 34(2), 340–355. doi:10.1037/0096-1523.34.2.340.
Freitas, A. L., Bahar, M., Yang, S., & Banai, R. (2007). Contextual adjustments in cognitive control across tasks. Psychological Science, 18(12), 1040–1043.
Funes, M. J., Lupiáñez, J., & Humphreys, G. (2010a). Analyzing the generality of conflict adaptation effects. Journal of Experimental Psychology: Human Perception and Performance, 36(1), 147–161. doi:10.1037/a0017598.
Funes, M. J., Lupiáñez, J., & Humphreys, G. (2010b). Sustained vs. transient cognitive control: Evidence of a behavioral dissociation. Cognition, 114(3), 338–347. doi:10.1016/j.cognition.2009.10.007.
Goschke, T. (2000). Intentional reconfiguration and involuntary persistence in task set switching. In S. Monsell & J. Driver (Eds.), Control of Cognitive Processes: Attention and Performance XVIII (pp. 331–355). Cambridge: MIT Press.
Gratton, G., Coles, M. G., & Donchin, E. (1992). Optimizing the use of information: Strategic control of activation of responses. Journal of Experimental Psychology: General, 121(4), 480–506.
Grundy, J. G., Benarroch, M. F. F., Woodward, T. S., Metzak, P. D., Whitman, J. C., & Shedden, J. M. (2013). The bivalency effect in task switching: Event-related potentials. Human Brain Mapping, 34(5), 999–1012. doi:10.1002/hbm.21488.
Hommel, B. (1994). Spontaneous decay of response-code activation. Psychological Research, 56(4), 261–268.
Jacoby, L. L., Lindsay, D. S., & Hessels, S. (2003). Item-specific control of automatic processes: Stroop process dissociations. Psychonomic Bulletin & Review, 10(3), 638–644.
Kan, I. P., Teubner-Rhodes, S., Drummey, A. B., Nutile, L., Krupa, L., & Novick, J. M. (2013). To adapt or not to adapt: The question of domain-general cognitive control. Cognition, 129(3), 637–651. doi:10.1016/j.cognition.2013.09.001.
Kiesel, A., Steinhauser, M., Wendt, M., Falkenstein, M., Jost, K., Philipp, A. M., & Koch, I. (2010). Control and interference in task switching—A review. Psychological Bulletin, 136(5), 849–874. doi:10.1037/a0019842.
Kleiman, T., Hassin, R. R., & Trope, Y. (2014). The control-freak mind: Stereotypical biases are eliminated following conflict-activated cognitive control. Journal of Experimental Psychology: General, 143(2), 498–503. doi:10.1037/a0033047.
Kunde, W., Augst, S., & Kleinsorge, T. (2012). Adaptation to (non)valent task disturbance. Cognitive, Affective, & Behavioral Neuroscience, 12(4), 644–660. doi:10.3758/s13415-012-0116-8.
Kunde, W., & Stöcker, C. (2002). A Simon effect for stimulus-response duration. The Quarterly Journal of Experimental Psychology Section A, 55(2), 581–592. doi:10.1080/02724980143000433.
Lindsay, D. S., & Jacoby, L. L. (1994). Stroop process dissociations: The relationship between facilitation and interference. Journal of Experimental Psychology: Human Perception and Performance, 20(2), 219–234.
Loft, S., Kearney, R., & Remington, R. (2008). Is task interference in event-based prospective memory dependent on cue presentation? Memory & Cognition, 36(1), 139–148. doi:10.3758/MC.36.1.139.
Logan, G. D., & Zbrodoff, N. J. (1979). When it helps to be misled: Facilitative effects of increasing the frequency of conflicting stimuli in a Stroop-like task. Memory & Cognition, 7(3), 166–174. doi:10.3758/BF03197535.
Lowe, D. G., & Mitterer, J. O. (1982). Selective and divided attention in a Stroop task. Canadian Journal of Psychology, 36(4), 684–700. doi:10.1037/h0080661.
MacLeod, C. M. (1991). Half a century of research on the Stroop effect: An integrative review. Psychological Bulletin, 109(2), 163–203. doi:10.1037/0033-2909.109.2.163.
Mayr, U., Awh, E., & Laurey, P. (2003). Conflict adaptation effects in the absence of executive control. Nature Neuroscience, 6(5), 450–452. doi:10.1038/nn1051.
McDaniel, M. A., & Einstein, G. O. (2000). Strategic and automatic processes in prospective memory retrieval: A multiprocess framework. Applied Cognitive Psychology, 14(7), S127–S144.
Meier, B., & Rey-Mermet, A. (2012a). Beyond feature binding: Interference from episodic context binding creates the bivalency effect in task-switching. Frontiers in Psychology, 3, 386–394. doi:10.3389/fpsyg.2012.00386.
Meier, B., & Rey-Mermet, A. (2012b). Beyond monitoring: After-effects of responding to prospective memory targets. Consciousness and Cognition, 21(4), 1644–1653. doi:10.1016/j.concog.2012.09.003.
Meier, B., Rey-Mermet, A., & Rothen, N. (2015). Turning univalent stimuli bivalent: Synesthesia can cause cognitive conflict in task switching. Cognitive Neuroscience,. doi:10.1080/17588928.2015.1017449.
Meier, B., Rey-Mermet, A., Woodward, T. S., Müri, R., & Gutbrod, K. (2013). Episodic context binding in task switching: Evidence from amnesia. Neuropsychologia, 51(5), 886–892. doi:10.1016/j.neuropsychologia.2013.01.025.
Meier, B., Woodward, T. S., Rey-Mermet, A., & Graf, P. (2009). The bivalency effect in task switching: General and enduring. Canadian Journal of Experimental Psychology, 63(3), 201–210. doi:10.1037/a0014311.
Meier, B., Zimmermann, T. D., & Perrig, W. J. (2006). Retrieval experience in prospective memory: Strategic monitoring and spontaneous retrieval. Memory, 14(7), 872–889. doi:10.1080/09658210600783774.
Metzak, P. D., Meier, B., Graf, P., & Woodward, T. S. (2013). More than a surprise: The bivalency effect in task switching. Journal of Cognitive Psychology, 25(7), 833–842. doi:10.1080/20445911.2013.832196.
Notebaert, W., Houtman, F., Opstal, F. V., Gevers, W., Fias, W., & Verguts, T. (2009). Post-error slowing: An orienting account. Cognition, 111(2), 275–279. doi:10.1016/j.cognition.2009.02.002.
Notebaert, W., & Verguts, T. (2011). Conflict and error adaptation in the Simon task. Acta Psychologica, 136(2), 212–216. doi:10.1016/j.actpsy.2010.05.006.
Núñez Castellar, E., Kühn, S., Fias, W., & Notebaert, W. (2010). Outcome expectancy and not accuracy determines posterror slowing: ERP support. Cognitive, Affective & Behavioral Neuroscience, 10(2), 270–278. doi:10.3758/CABN.10.2.270.
Rey-Mermet, A., Koenig, T., & Meier, B. (2013). The bivalency effect represents an interference-triggered adjustment of cognitive control: An ERP study. Cognitive, Affective, & Behavioral Neuroscience, 13(3), 575–583. doi:10.3758/s13415-013-0160-z.
Rey-Mermet, A., & Meier, B. (2012). The bivalency effect: Adjustment of cognitive control without response set priming. Psychological Research, 76(1), 50–59. doi:10.1007/s00426-011-0322-y.
Rey-Mermet, A., & Meier, B. (2013). An orienting response is not enough: Bivalency not infrequency causes the bivalency effect. Advances in Cognitive Psychology, 9(3), 146–155. doi:10.2478/v10053-008-0142-9.
Rey-Mermet, A., & Meier, B. (2014). More conflict does not trigger more adjustment of cognitive control for subsequent events: A study of the bivalency effect. Acta Psychologica, 145, 111–117. doi:10.1016/j.actpsy.2013.11.005.
Rey-Mermet, A., & Meier, B. (2015). Age affects the adjustment of cognitive control after a conflict: Evidence from the bivalency effect. Aging, Neuropsychology, and Cognition, 22(1), 72–94. doi:10.1080/13825585.2014.889070.
Rogers, R. D., & Monsell, S. (1995). Costs of a predictable switch between simple cognitive tasks. Journal of Experimental Psychology: General, 124(2), 207–231. doi:10.1037/0096-3445.124.2.207.
Schlaghecken, F., Refaat, M., & Maylor, E. A. (2011). Multiple systems for cognitive control: Evidence from a hybrid prime-Simon task. Journal of Experimental Psychology: Human Perception and Performance, 37(5), 1542–1553. doi:10.1037/a0024327.
Schmidt, J. R. (2013a). Questioning conflict adaptation: Proportion congruent and Gratton effects reconsidered. Psychonomic Bulletin & Review, 20(4), 615–630. doi:10.3758/s13423-012-0373-0.
Schmidt, J. R. (2013b). The Parallel Episodic Processing (PEP) model: Dissociating contingency and conflict adaptation in the item-specific proportion congruent paradigm. Acta Psychologica, 142(1), 119–126. doi:10.1016/j.actpsy.2012.11.004.
Schmidt, J. R., & De Houwer, J. (2011). Now you see it, now you don’t: Controlling for contingencies and stimulus repetitions eliminates the Gratton effect. Acta Psychologica, 138(1), 176–186. doi:10.1016/j.actpsy.2011.06.002.
Simon, J. R., & Small, A. M. (1969). Processing auditory information: Interference from an irrelevant cue. Journal of Applied Psychology, 53(5), 433–435. doi:10.1037/h0028034.
Smith, R. E. (2003). The cost of remembering to remember in event-based prospective memory: Investigating the capacity demands of delayed intention performance. Journal of Experimental Psychology: Learning, Memory, and Cognition, 29(3), 347–361. doi:10.1037/0278-7393.29.3.347.
Smith, R. E. (2010). What costs do reveal and moving beyond the cost debate: Reply to Einstein and McDaniel (2010). Journal of Experimental Psychology: Learning, Memory, and Cognition, 36(4), 1089–1095. doi:10.1037/a0019183.
Smith, R. E. (2011). Prospective memory: Beyond the cost debate. Zeitschrift Für Psychologie, 219(2), 75–76. doi:10.1027/2151-2604/a000050.
Sohn, M.-H., & Carlson, R. A. (2000). Effects of repetition and foreknowledge in task-set reconfiguration. Journal of Experimental Psychology: Learning, Memory, and Cognition, 26(6), 1445–1460. doi:10.1037//0278-7393.26.6.1445.
Spapé, M. M., Band, G. P. H., & Hommel, B. (2011). Compatibility-sequence effects in the Simon task reflect episodic retrieval but not conflict adaptation: Evidence from LRP and N2. Biological Psychology, 88(1), 116–123. doi:10.1016/j.biopsycho.2011.07.001.
Stroop, J. R. (1935). Studies of interference in serial verbal reactions. Journal of Experimental Psychology, 18(6), 643–662. doi:10.1037/h0054651.
Vandierendonck, A., Liefooghe, B., & Verbruggen, F. (2010). Task switching: Interplay of reconfiguration and interference control. Psychological Bulletin, 136(4), 601–626. doi:10.1037/a0019791.
Verbruggen, F., Liefooghe, B., Notebaert, W., & Vandierendonck, A. (2005). Effects of stimulus–stimulus compatibility and stimulus–response compatibility on response inhibition. Acta Psychologica, 120(3), 307–326. doi:10.1016/j.actpsy.2005.05.003.
Verguts, T., & Notebaert, W. (2008). Hebbian learning of cognitive control: Dealing with specific and nonspecific adaptation. Psychological Review, 115(2), 518–525. doi:10.1037/0033-295X.115.2.518.
Verguts, T., & Notebaert, W. (2009). Adaptation by binding: a learning account of cognitive control. Trends in Cognitive Sciences, 13(6), 252–257. doi:10.1016/j.tics.2009.02.007.
Verguts, T., Notebaert, W., Kunde, W., & Wühr, P. (2011). Post-conflict slowing: Cognitive adaptation after conflict processing. Psychonomic Bulletin & Review, 18(1), 76–82. doi:10.3758/s13423-010-0016-2.
Waszak, F., Hommel, B., & Allport, A. (2003). Task-switching and long-term priming: Role of episodic stimulus-task bindings in task-shift costs. Cognitive Psychology, 46(4), 361–413. doi:10.1016/S0010-0285(02)00520-0.
Waszak, F., Hommel, B., & Allport, A. (2004). Semantic generalization of stimulus-task bindings. Psychonomic Bulletin & Review, 11(6), 1027–1033. doi:10.3758/BF03196732.
Wendt, M., Kluwe, R. H., & Peters, A. (2006). Sequential modulations of interference evoked by processing task-irrelevant stimulus features. Journal of Experimental Psychology: Human Perception and Performance, 32(3), 644–667. doi:10.1037/0096-1523.32.3.644.
West, R., & Baylis, G. C. (1998). Effects of increased response dominance and contextual disintegration on the Stroop interference effect in older adults. Psychology and Aging, 13(2), 206–217.
Woodward, T. S., Meier, B., Tipper, C., & Graf, P. (2003). Bivalency is costly: Bivalent stimuli elicit cautious responding. Experimental Psychology, 50(4), 233–238. doi:10.1026//1618-3169.50.4.233.
Woodward, T. S., Metzak, P. D., Meier, B., & Holroyd, C. B. (2008). Anterior cingulate cortex signals the requirement to break inertia when switching tasks: a study of the bivalency effect. NeuroImage, 40(3), 1311–1318. doi:10.1016/j.neuroimage.2007.12.049.
Wylie, G., & Allport, A. (2000). Task switching and the measurement of “switch costs”. Psychological Research, 63(3–4), 212–233. doi:10.1007/s004269900003.
Acknowledgments
This work was supported by the Center for Cognition, Learning, and Memory, University of Bern, Switzerland. We thank Anne-Catherine Amstutz, Rahel Gfeller, Eva Gut, Tiffany Jacob, Melanie Künzli, Isabelle Marti, Seline Messmer, Sabrina Schmied, and Michèle Spichtig for testing the participants. We also thank Michel Druey for helpful comments on an earlier version.
Author information
Authors and Affiliations
Corresponding author
Appendix
Appendix
See Table 6.
Rights and permissions
About this article

Cite this article
Rey-Mermet, A., Meier, B. Post-conflict slowing after incongruent stimuli: from general to conflict-specific. Psychological Research 81, 611–628 (2017). https://doi.org/10.1007/s00426-016-0767-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00426-016-0767-0