
Abstract
Influenza represents a substantial global healthcare burden, with annual epidemics resulting in 3–5 million cases of severe illness with a significant associated mortality. In addition, the risk of a virulent and lethal influenza pandemic has generated widespread and warranted concern. Currently licensed influenza vaccines are limited in their ability to induce efficacious and long-lasting herd immunity. In addition, and as evidenced by the H1N1 pandemic in 2009, there can be a significant delay between the emergence of a pandemic influenza and an effective, antibody-inducing vaccine. There is, therefore, a continued need for new, efficacious vaccines conferring cross-clade protection—obviating the need for biannual reformulation of seasonal influenza vaccines. Development of such a vaccine would yield enormous health benefits to society and also greatly reduce the associated global healthcare burden. There are a number of alternative influenza vaccine technologies being assessed both preclinically and clinically. In this review we discuss viral vectored vaccines, either recombinant live-attenuated or replication-deficient viruses, which are current lead candidates for inducing efficacious and long-lasting immunity toward influenza viruses. These alternate influenza vaccines offer real promise to deliver viable alternatives to currently deployed vaccines and more importantly may confer long-lasting and universal protection against influenza viral infection.
Similar content being viewed by others


Introduction
Influenza epidemics are associated with a nontrivial morbidity and mortality; up to one billion infections occur annually with upwards of a half a million associated deaths (1,2). The mortality associated with the first influenza pandemic of the 21st century, caused by the swine-origin influenza A H1N1/09 virus, was not as high as first anticipated, nor when compared with the 1918 pandemic, which claimed an estimated 15–50 million lives worldwide (3). However, the continued risk of a lethal and transmissible influenza pandemic has generated widespread and warranted concern (4).
There are three genera of influenza virus that infect humans, influenza A, B and C viruses. Type A influenza viruses are the most virulent influenza viruses infecting humans and can be divided into different subtypes on the basis of the antigenic properties of the virion surface proteins, hemagglutinin (HA) and neuraminidase (NA) (5,6). Currently there are 17 HA subtypes and 9 NA subtypes identified, and most of the possible combinations have been found circulating in wild birds (5–7). However, the majority of influenza in humans is caused by only three subtypes of HA (H1, H2 and H3) and two subtypes of NA (N1 and N2) (8). Indeed, recent pandemics have been caused by three subtypes, namely: H1N1 (in 1918 and 2009, the Spanish and Swine flu pandemics, respectively), H2N2 (in 1957, known as the Asian flu) and H3N2 (in 1968, known as the Hong Kong flu) (8).
In contrast to influenza A viruses, B viruses mutate slowly (9). At present, there are two antigenically distinct type B influenza lineages (Victoria and Yamagata), which are concurrently circulating (10,11), and individuals exposed to one lineage have limited cross-protective immunity because of antigenic dissimilarity between strains (11). The burden of disease due to influenza B viral infection falls largely on children and young adults, and this influenza virus causes seasonal influenza epidemics every 2–4 years (11). By contrast, influenza C virus, although a common cause of mild upper respiratory illness, rarely causes severe illness or local epidemics because most people acquire protective antibodies early in life (12).
Influenza Vaccination of Population Cohorts
Licensed influenza vaccines principally mediate protection through the induction of antibodies against virion surface proteins. There are two primary types of commercially available seasonal influenza vaccines: live attenuated, cold-adapted influenza vaccines (LAIV; administered by nasal spray), and inactivated, principally trivalent, vaccine (parenterally administrated). Seasonal influenza vaccines are biannually formulated to include HA protein, or attenuated influenza virus, of the three influenza strains representative of the circulating strains in the northern or southern hemisphere. Routinely, formulations contain two human influenza A subtypes (H3N2 and H1N1) and one of the two influenza B lineages (Yamagata or Victoria) (2). Many countries have implemented influenza healthcare strategies in which the vaccination is administered to discrete population cohorts at risk of complications, the very young and the elderly.
The global burden of influenza disease mainly occurs in infants aged 5 years or under; accounting for 90 million infections annually (13,14). Children shed virus at a high rate and routinely congregate in high numbers (for example, schools, nurseries), thus facilitating easy dissemination of the virus to family members and in turn the wider public. The question of whether to mass vaccinate young children against influenza is a point of considerable debate among both the clinical immunology and public health communities (15), and at present there is no clear consensus. Supporters of vaccinating children against influenza argue that conferring protection to this segment of society will not only benefit children, but will also benefit the wider adult population (1,15,16). Opposition to this healthcare strategy hinges on the idea that individuals, including children, may accrue better heterosubtypic immunity through natural exposure to influenza virus as opposed to vaccination (17–23). Heterosubtypic, or cross-clade, memory can be broadly classified as protection induced to an influenza subtype or strain that confers protection against other strains and can be mediated by both humoral and cellular immunity (24–27).
Approximately 90% of influenza-related deaths, outside of pandemic settings, occur among people aged 65 years or older and this heightened susceptibility is increased as individuals age; persons aged 85 years or older are 16 times more likely to die of an influenza-related death compared with those aged 65 to 69 years (13,28). The progressive decline of the immune system is thought to be the principle cause of increased susceptibility to infection in the older adult. The age-related decline in immune responses (immunosenescence) affects both the innate and adaptive immune response (29).
Limitations of Currently Licensed Influenza Vaccines
Although influenza vaccines have undoubtedly saved numerous lives during both pandemic and epidemic outbreaks, there remains considerable variation in how influenza vaccines are used and the efficacy achieved in targeted population cohorts. As such, influenza and its associated complications still represent a considerable global healthcare burden. For instance, in healthy adults, the efficacy of inactivated vaccines is estimated at 50–75% (30,31), whereas in children under the age of 2 years, efficacy associated with trivalent influenza vaccine (TIV) vaccination is comparable to placebo (32). In the elderly, the validity of using inactivated influenza vaccines has been questioned (33), with a recent systematic review asserting that there is no conclusive evidence regarding efficacy or effectiveness of influenza vaccines for people aged 65 years or older (34).
Compared with inactivated vaccines, efficacy for live vaccines in children is considerably higher (approximately 82%); however, live vaccines are also associated with low effectiveness (approximately 33%) (32). Moreover, LAIV is not licensed in children under 2 years of age because it is associated with an increased risk of associated wheezing (32), and in adults over the age of 49 years, LAIV is not licensed owing to a lack of corroborated efficacy (35).
Thus, there is a clear need to develop effective and efficacious influenza vaccines for both infants and the elderly. Before being able to do this, however, we must understand what factors underpin the current limitations of licensed influenza vaccines.
Adaptive Immune Response following Vaccination
Humoral immune response. Currently licensed influenza vaccines induce a relatively narrow adaptive immune response with limited cross-clade immunity. Cross-clade protection refers to protective immunity against one influenza isolate, which subsequently confers protective efficacy upon exposure to different isolates.
The majority of humoral responses following influenza vaccination are directed toward HA, principally toward the globular head domain found in the HA1 segment. Due to the constantly evolving and mutating propensity of HA, these neutralizing antibodies offer little or no protection between subtypes or even between strains, and in some cases appear to be detrimental (18). This is a considerable disadvantage considering influenza A undergoes both antigenic drift and antigenic shift (36). Antigenic drift refers to small, gradual changes that occur through point mutations in the HA or NA coding genes. Antigenic shift refers to an abrupt, major alteration that generally produces a subtype change and can occur through either direct animal-to-human transmission or through a reassortment of human and animal influenza A (36). Most escape mutants contain mutations in the globular head domain, and this region of the protein contains widespread immunodominant decoy domains, which can readily mutate without significantly affecting virus fitness (37). Thus, yearly vaccinations with reformulated vaccines are required because of the antigenic differences that occur between, and even within, influenza strains.
Natural exposure to influenza is thought to induce a broad immune response, inducing cross-clade protection through T- and B-cell responses (25,38,39). Indeed, antibodies directed against the stalk region of HA have previously been shown to confer broad-spectrum protection toward different influenza isolates (40). Annual vaccination with TIV is thought to be a relatively poor inducer of these cross-clade immune responses (18,23).
Cellular immune response. Cellular immune responses are thought to play an important protective role during influenza infection, and human challenge studies have demonstrated a negative correlation between T-cell responses to viral antigens and influenza disease (38). T-cell responses, particularly against relatively well-conserved internal influenza antigens, can efficaciously provide cross-clade protection (38,41,42). Although human cytotoxic T-lymphocyte (CTL) immunity declines during periods of low influenza prevalence, these cells are still detectable after 5 years, with an estimated half-life of 2–3 years (43), making these cells an ideal target for boosting through vaccination.
Although cellular immunity does not prevent viral infection, it is important in clearing and curtailing infection. Cytotoxic CD8+ T cells play a major role and can eliminate virally infected cells (44,45). A direct cytolytic role for CD4+ T cells has been demonstrated (25,46). More importantly, CD4+ T cells can provide B-cell help, and prior CD4+ T-cell immunity can efficaciously augment HA-specific antibody responses (47,48).
Unfortunately, even though inactivated influenza vaccines routinely contain internal influenza antigens, specifically nucleoprotein and matrix protein 1 (49,50), these vaccines do not boost cellular immunity to an efficacious level (49,51).
The Challenge of Surveillance, Prediction and Deployment
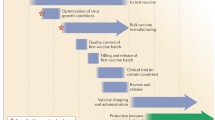
Prediction of the major circulating influenza subtypes on the basis of routine surveillance—required to update influenza vaccines each year—can fail. Indeed, mismatch between the vaccine components and circulating virus occurs approximately every 3–4 years (52,53). This mismatch has a significant effect on vaccine efficacy; in healthy adults, parenteral vaccine efficacy against seasonal influenza is approximately 75% when the circulating influenza and vaccine strain match; however, efficacy decreases to 44% when mismatch occurs (30).
Substantial costs are incurred because of the need for annual reformulation and given the manufacturing process, which for the recent 2009 H1N1 pandemic took over 6 months even with the implemented pandemic readiness plans (54). Each year, there is essentially a race between production, deployment and the spread of infection.
Limited Immunogenicity and Stability
One of the main challenges for inducing strong immune responses following vaccination with inactivated or subunit protein vaccines is the inherent poor immunogenicity of proteins, which can be partially overcome by the addition of adjuvants. Adjuvants are a common means to augment adaptive immune responses to vaccines and can be broadly classified into vehicles or immunostimulants (55). The former group largely enhance presentation to the immune system, whereas the latter enhance the adaptive immune response (55–57). Some adjuvants have been shown to advantageously increase and broaden the epitopes recognized by the immune repertoire (58,59). A number of strategies have been deployed to improve influenza vaccine efficacy for the elderly, and the inclusion of adjuvants as a means to augment humoral immunity in the elderly has demonstrated relatively encouraging results (60,61).
Another challenge is the long-term stability of recombinant proteins; the inclusion of some adjuvants can facilitate increased vaccine stability during storage or upon injection (62). However, even with this augmented stability there are often thermostability difficulties, especially when vaccines occupy a large amount of cold-chain storage space due to relatively complex administration or dosing regimens.
Alternative Vaccine Platforms
There are a number of platforms being explored for more efficacious influenza vaccine delivery and these can be broadly classified into: recombinant protein/peptide, DNA vaccines, viruslike particles, virosomes and viral vectors.
Recombinant Protein/Peptide
Recombinant technology has been used to generate influenza proteins in insect cells, bacteria and plants (63–65); and although much antigen can be produced, these proteins are generally poorly immunogenic and require complex immunization regimes and/or the inclusion of adjuvants to induce efficacious humoral immunity (66). For cellular immune responses, antigenic mapping of T-cell epitopes followed with antigen string delivery (in the form of peptides) is actively being investigated. Unfortunately, peptides are generally poorly immunogenic, and repeated administration to augment adaptive immune responses can induce tolerance rather that immunogencity (67).
DNA Vaccines
Immunization with DNA, encoding influenza proteins, has been studied for many years as an alternate vaccination strategy but clinical trials have met with limited success (68,69). DNA vaccines do not boost preexisting immunity (the scenario for the adult human population) and are generally only weakly immunogenic in humans (70).
Viruslike Particles
Self-assembled viral capsid proteins, viruslike particles (VLPs), have been produced from a number of viruses. A key restriction to certain VLP platforms is the small size of foreign epitopes that can be accommodated (for example, HA is a relatively big protein) (71). For the generation of VLPs appropriate protein glycosylation, correct assembly and ease of purification need to be considered. These concerns may dictate the use of mammalian cell culture systems, which is a less controllable and a more costly system (71).
Virosomes
Although similar to VLPs in many respects, virosomes, by contrast, are essentially assembled in vitro. There are a limited number of virosomal-adjuvanted-subunit influenza vaccines currently licensed; the generation of these vaccines requires good manufacturing practice (GMP)-grade influenza virus and, as such, these vaccines face similar obstacles as the production of TIV (72,73).
The Rise of the Viral Vector
Vectored vaccines, such as recombinant live-attenuated or replication-deficient viruses, are currently being developed for vaccine delivery and offer a number of advantages over other emerging technologies.
Induction of Both Arms of the Immune Response
Viral vectored vaccines are particularly good at expressing full-length protein and generating conformationally restricted epitopes, thus driving potent humoral responses following immunization (74). In addition, vectored vaccines can induce high levels of protective cellular immunity (75–77). Thus, viral vectors efficaciously induce both arms of the adaptive immune system.
High Levels and Longevity of Protein Expression
Following vaccination with viral vector vaccines, virus can infect local cells and produce high levels of protein (up to 100 µ g following vaccination with a modest amount of viral vector, 1010 viral particles (VP) of adenovirus or 107 plaque-forming units (PFU) of modified vaccinia virus Ankara) (78). Antigen has been detected at the immunization site up to 6 months after viral vectored vaccination (79,80), which is in contrast to the transient antigen depots following protein vaccination, even when coadministered with adjuvants (81). The increased presence of antigen may enhance the survival of certain memory T cells and augment high-affinity antibody selection (82,83).
Intrinsic Adjuvant Capacity
Viral infection results in both apoptosis and necrosis of localized tissue, facilitating the release of antigen, which is subsequently available to drive cellular and humoral immunity (84,85). The associated cell death following viral infection will also advantageously augment adaptive immune responses, circumventing the need for the inclusion of adjuvants.
Enhanced Stability
Live recombinant viral vectors must remain infectious to be effective, and conventional, continuous refrigeration is required to maintain stability and viability. However, recent methodologies have allowed complete recovery of viral titer and immunogenicity after storage at up to 45°C for 6 months (86).
In addition, viral vectored vaccines are relatively easy to manipulate and produce, and have well-established safety profiles. There are an increasing number of viral-based vectors currently employed as antigen delivery systems, including DNA virus vectors (for example, poxviruses, herpesvirus, adenovirus and baculovirus) and RNA virus vectors (for example, paramyxovirus, rhabdovirus, bunyavirus, alphavirus, coronavirus, retrovirus and flavivirus) (87). However, the majority of human clinical trials assessing viral vectored influenza vaccines use poxviral and adenoviral technologies, and these vectors are considered lead candidates for the immunogenic delivery of antigens (77).
Adenoviruses
Adenoviruses (Ads) are nonenveloped DNA viruses originally identified as infectious agents responsible for certain acute respiratory infections. Ad infection usually manifests as mild disease, which is promptly resolved; however, infection of immunocompromised individuals can result in more severe disease (88,89). Several properties of Ad viruses are attractive for vaccine use, including relative ease of manipulation, infection of both dividing and nondividing cells, high levels of transgene expression, ability to grow to high titers in vitro and lack of integration in the host genome and stability (physical and genetic) (87,89,90).
Ads are rendered replication defective by deletion of the E1 region genes, and although replication competent Ad vectors have been demonstrated to be quantitatively (reduction of required dose) and qualitatively (induction of site-specific immunity) more effective, the release of genetically modified Ad vectors into the general public, fuelling concerns of possible carriage, has limited their systematic use (89,90).
Nonreplicating human Ad (for example, Adhu5) vectors have been extensively used in gene therapy applications, but unfortunately, most adults have high titers of neutralizing antibodies against a number of Ad serotypes, including Adhu5, which has a negative impact on their effectiveness as vaccine platforms (91). To circumvent this limitation, alternative serotypes of adenovirus not found in human populations are being developed (92,93). These include Ads of rare serotypes (that is, Ad11, Ad26, Ad35, Ad28 and Ad49) and nonhuman Ads, principally of chimpanzee origin AdC (77,87,94). In addition, engineered chimeric vectors in which the Adhu5 capsid has been modified (thereby reducing the number of antigenic epitopes previously seen by man) are also being deployed (77,87,94).
Applicability of Adenoviral Vectored Vaccines for Influenza
A number of vaccine studies have demonstrated the feasibility of using recombinant Ad vectors expressing influenza viral proteins to protect model organisms from influenza challenge. A recombinant Ad vector expressing H5N1 HA induced both cellular and humoral immunity and was protective, following lethal challenge, in mice and chickens (95). Heterosubtypic immunity toward certain influenza viruses has been induced through in ovo vaccination with a nonreplicating Ad vector expressing HA from H5N9 (96), and indeed long-lasting immunity, 12 months postimmunization, has been demonstrated following Ad vaccination (97).
The application of Ad viral vectored vaccines has progressed to human studies, and a phase I clinical trial using a nonreplicating adenovirus vectored nasally administered influenza vaccine has demonstrated that the vaccine is well tolerated and efficaciously induces humoral immunity. A follow-on trial evaluating the dose range, safety and immunogenicity of an intranasally administered AdhVN1203/04.H5 (adenovirus vectored H5 HA) in healthy adults 19–49 years of age is expected to be completed imminently (ClinicalTrials.gov Identifier: NCT00755703).
Replication competent Ad vectors, in general, induce stronger immune responses and a live, replication competent recombinant Ad vectored vaccine, expressing the HA from H5N1 (Ad4-H5-Vtn), will be administered either intranasally or tonsillarly in a phase 1 randomized, dose-escalation study expected to be completed in 2014 (Clinical-Trials.gov Identifier: NCT01443936).
Pox Virus
Poxviruses are among the most heavily exploited viral vectors, largely because of the overwhelming success of vaccinia virus in eradicating smallpox (87,98). As large DNA viruses, poxviruses can easily accommodate substantial amounts (over 25 kb) of extra DNA (99), thus facilitating a multivalent vaccine approach through the simultaneous expression of multiple transgenes (100,101). Poxviruses replicate within the cytoplasm of infected cells and do not integrate into the host genome, eliminating the potential for insertional mutagenesis. They have been developed as vaccines through two different approaches; the first relies on productive replication of an attenuated strain in a permissive host. The second uses replication-defective poxviruses, which may enter cells of nonpermissive hosts and express encoded proteins, but do not produce infectious virus (98,102).
Modified vaccinia virus Ankara (MVA), an attenuated strain of vaccinia virus, was originally developed during the smallpox eradication campaign, because wild-type strains can produce undesirable effects in humans (103,104). Attenuated vaccinia strains such as MVA undergo very limited, or nonproductive, replication in mammalian cells (102), although most of the virus proteins are produced, including inserted transgenes (98). MVA has an excellent safety profile in humans, having been used in many field studies (105,106)—safety has also been demonstrated in studies with immunocompromised macaques and humans (107,108).
One of the ongoing concerns with viral vector vaccines is their applicability for repeat vaccinations. Preclinical evaluation of MVA vectors has demonstrated their ability to repeatedly boost immune responses directed to recombinant antigens (109,110). Indeed, two phase I clinical trials using repeat vaccination with MVA vectored vaccines, at 6–8 months, demonstrate an ability to boost T-cell responses (111,112). These data suggest that vector-specific immunity does not greatly impede immune responses toward target antigens.
A major advantage of MVA over other vaccine preparations is that adjuvants are not required for high immunogenicity, because MVA has immunostimulatory properties (84,85) and can elicit potent humoral and antigen-specific T-cell responses (110,113). For the efficient activation of CD8+ T-cell responses with MVA vectors, delivery of full length antigen is generally found to be superior, suggesting that cross-priming may be particularly important in MVA-mediated CD8+ T-cell responses (110,114).
Applicability of Pox-Viral Vectored Vaccines for Influenza
There is now a body of evidence demonstrating that recombinant pox-viral vectors expressing influenza antigens are efficacious vaccines. Indeed, there is a licensed pox-viral influenza vaccine, TROVAC AIV H5, for the prevention of influenza in chickens. This vaccine uses the fowlpox virus encoding the HA gene from A/Turkey/Ireland/83 and has demonstrated protective efficacy against a wide range of highly pathogenic, sub-type-H5 viruses (115).
Preclinical work has demonstrated the feasibility of using MVA vectored influenza antigens as vaccines in mammals; mice vaccinated with MVA recombinants expressing the HA of H5N1 (A/Vietnam/1194/04; MVA-HA-VN/04) were fully protected against infection with the homologous strain and also against infection with H5 viruses from different clades (116–119). A pentavalent vaccina vaccine with HA, NA, nucleoprotein and the matrix proteins M1 and M2 was able to confer sterile cross-clade protection during stringent challenge studies (120).
Studies in higher primates have assessed the feasibility of using MVA-expressing hemagglutinin as an influenza vaccine. Immunization with A/Vietnam/1194/04 (MVA-HA-VN/04) induced cross-reactive antibodies and prevented virus replication in the respiratory tract of cynomolgus macaques (117). Indeed, a recent phase I clinical trial assessing an MVA vector encoding nucleoprotein and matrix protein 1 (MVA-NP + M1) has demonstrated that this vaccine was both safe and, more importantly, immunogenic—inducing high levels of interferon (IFN)-γ production from T-cells (75,121).
Future Directions
A promising avenue of influenza vaccine development is likely to come from the viral vectored vaccines currently progressing through phase I and II clinical trials. Designed to produce antiinfluenza responses, these vectored vaccines have demonstrated promising immunogenicity with good safety profiles and it is likely that these vaccines will be further used in prime-boost regimes aimed at augmenting protective immunity. These regimes involve priming the adaptive immune response toward an antigenic target through vaccination with a vector encoding the antigen of choice, followed by a boost, in which the same antigenic target is administered but in a different vector. Through this, the immune response is honed toward the antigen of choice, while avoiding boosting antivector immunity.
Prime-boost approaches, incorporating priming with either plasmid DNA vaccines or viral vector (typically Ad) followed by a boost with either an Ad-re-combinant or MVA expressing the same gene product, have been shown to enhance protective efficacy (122,123). Indeed, the use of heterologous regimens with Ad vectored vaccines boosted with MVA has been shown to augment antigen-specific T-cell responses and also to efficaciously induce humoral immunity (66,124–126).
As the supporting evidence evolves, a likely outcome is the use of viral vectored vaccines in prime-boost regimes to generate a strong and potentially heterosubtypic immune response. Indeed, heterosubtypic immunity toward certain influenza viruses has been induced through in ovo vaccination with a non-replicating Ad vector expressing HA from H5N9 (96). Furthermore, vaccination with MVA recombinants expressing the HA of H5N1 (A/Vietnam/1194/04; MVA-HA-VN/04) confers protection against infection with the homologous strain and also against H5 viruses from different clades (116–119). The induction of efficacious and long-lasting cross-clade immune responses will bypass the need for annual vaccine reformulation and immunization and, most importantly, will provide protection during future influenza pandemics.
Conclusions
Despite the long-standing service of currently licensed influenza vaccines, they carry a number of real limitations that need to be addressed before population-based influenza vaccination strategies are optimized. Undoubtedly, a vaccine that induced durable, sustainable and cross-clade protection against influenza would be of great medicinal and economic benefit worldwide.
At the very minimum, a revision of the current vaccine regimes for at-risk populations needs to be considered to optimize health outcomes. An improved regime is likely to include different vaccines for the different ages of man: for children, a four-pronged approach including the two currently circulating influenza A subtypes and both influenza B strains has been suggested (11). Whereas for the elderly, inclusion of the two currently circulating influenza A subtypes and both influenza B strains at a higher dose or the inclusion of adjuvants to augment the lower responses should be considered.
Undoubtedly, the continued and concerted effort invested in developing efficacious viral vectored influenza vaccines will deliver promising and real alternatives to those currently deployed and greatly impact upon public health in the 21st century.
Disclosure
The authors declare that they have no competing interests as defined by Molecular Medicine, or other interests that might be perceived to influence the results and discussion reported in this paper.
References
Molinari NA, et al. (2007). The annual impact of seasonal influenza in the US: measuring disease burden and costs. Vaccine. 25: 5086–96.
Lambert LC, Fauci AS. (2010). Influenza vaccines for the future. N. Engl. J. Med. 363:2036–44.
Johnson NP, Mueller J. (2002). Updating the accounts: global mortality of the 1918–1920 “Spanish” influenza pandemic. Bull. Hist. Med. 76:105–15.
Normile D. (2011). Avian influenza. Warning of H5N1 resurgence surprises community. Science. 333:1369.
Kovacova A, Ruttkay-Nedecky G, Haverlik IK, Janecek S. (2002). Sequence similarities and evolutionary relationships of influenza virus A hemagglutinins. Virus genes. 24:57–63.
Gamblin SJ, Skehel JJ. (2010). Influenza hemagglutinin and neuraminidase membrane glycoproteins. J. Biol. Chem. 285:28403–9.
Tong S, et al. (2012). A distinct lineage of influenza A virus from bats. Proc. Natl. Acad. Sci. U. S. A. 109:4269–74.
Elderfield R. Barclay W. (2011). Influenza pandemics. Adv. Exp. Med. Biol. 719:81–103.
Yamashita M, Krystal M, Fitch WM, Palese P. (1988). Influenza B virus evolution: co-circulating lineages and comparison of evolutionary pattern with those of influenza A and C viruses. Virology. 163:112–22.
Li WC, et al. (2008). Clinical and genetic characterization of severe influenza B-associated diseases during an outbreak in Taiwan. J. Clin. Virol. 42:45–51.
Belshe RB. (2010). The need for quadrivalent vaccine against seasonal influenza. Vaccine. 28(Suppl 4):D45–53.
Matsuzaki Y, et al. (2006). Clinical features of influenza C virus infection in children. J. Infect. Dis. 193:1229–35.
Thompson WW, et al. (2003). Mortality associated with influenza and respiratory syncytial virus in the United States. JAMA. 289:179–86.
Nair H, et al. (2011). Global burden of respiratory infections due to seasonal influenza in young children: a systematic review and meta-analysis. Lancet. 378:1917–30.
Heikkinen T, et al. (2006). Should healthy children be vaccinated against influenza? A consensus report of the Summits of Independent European Vaccination Experts. Eur. J. Pediatr. 165:223–8.
Reichert TA, et al. (2001). The Japanese experience with vaccinating schoolchildren against influenza. N. Engl. J. Med. 344:889–96.
Corbeel L. (2007). Should healthy children be vaccinated against influenza? Comments about this query. Eur. J. Pediatr. 166:629–31.
Skowronski DM, et al. (2010). Association between the 2008–09 seasonal influenza vaccine and pandemic H1N1 illness during Spring-Summer 2009: four observational studies from Canada. PLoS Med. 7:e1000258.
Choi YS, et al. (2011). Reduced antibody responses to the pandemic (H1N1) 2009 vaccine after recent seasonal influenza vaccination. Clin. Vaccine Immunol. 18:1519–23.
Huijskens E, et al. (2011). Immunogenicity, boost-ability, and sustainability of the immune response after vaccination against Influenza A virus (H1N1) 2009 in a healthy population. Clin. Vaccine Immunol. 18:1401–5.
Cowling BJ, et al. (2010). Protective efficacy of seasonal influenza vaccination against seasonal and pandemic influenza virus infection during 2009 in Hong Kong. Clin. Infect. Dis. 51:1370–9.
Bodewes R, et al. (2011). Annual vaccination against influenza virus hampers development of virus-specific CD8 T cell immunity in children. J. Virol. 85:11995–2000.
Glezen WP. (2010). How did the 2008–2009 seasonal influenza vaccine affect the pandemic? Clin. Infect. Dis. 51:1380–2.
Epstein SL, Price GE. (2010). Cross-protective immunity to influenza A viruses. Expert Rev. Vaccines. 9:1325–41.
Wilkinson TM, et al. (2012). Preexisting influenza-specific CD4(+) T cells correlate with disease protection against influenza challenge in humans. Nat. Med. 18:274–80.
Kaur K, Sullivan M, Wilson PC. (2011). Targeting B cell responses in universal influenza vaccine design. Trends Immunol. 32:524–31.
DiMenna LJ, Ertl HC. (2009). Pandemic influenza vaccines. Curr. Top. Microbiol. Immunol. 333:291–321.
Nordin J, et al. (2001). Influenza vaccine effectiveness in preventing hospitalizations and deaths in persons 65 years or older in Minnesota, New York, and Oregon: data from 3 health plans. J. Infect. Dis. 184:665–70.
Kovaiou RD, Herndler-Brandstetter D, Grubeck-Loebenstein B. (2007). Age-related changes in immunity: implications for vaccination in the elderly. Expert Rev. Mol. Med. 9:1–17.
Jefferson T, et al. (2010). Vaccines for preventing influenza in healthy adults. Cochrane Database Syst. Rev. (7):CD001269.
Jefferson TO, Rivetti D, Di Pietrantonj C, Rivetti A, Demicheli V. (2007). Vaccines for preventing influenza in healthy adults. Cochrane Database Syst. Rev. (2):CD001269.
Jefferson T, Rivetti A, Harnden A, Di Pietrantonj C, Demicheli V. (2008). Vaccines for preventing influenza in healthy children. Cochrane Database Syst. Rev. (2):CD004879.
Jackson ML, et al. (2008). Influenza vaccination and risk of community-acquired pneumonia in immunocompetent elderly people: a population-based, nested case-control study. Lancet. 372:398–405.
Jefferson T, et al. (2010). Vaccines for preventing influenza in the elderly. Cochrane Database Syst. Rev. (2):CD004876.
Ambrose CS, Luke C, Coelingh K. (2008). Current status of live attenuated influenza vaccine in the United States for seasonal and pandemic influenza. Influenza Other Respi. Viruses. 2:193–202.
Doherty PC, Turner SJ, Webby RG, Thomas PG. (2006). Influenza and the challenge for immunology. Nat. Immunol. 7:449–55.
Wilson IA, Cox NJ. (1990). Structural basis of immune recognition of influenza virus hemagglutinin. Annu. Rev. Immunol. 8:737–71.
McMichael AJ, Gotch FM, Noble GR, Beare PA. (1983). Cytotoxic T-cell immunity to influenza. N. Engl. J. Med. 309:13–7.
Skountzou I, et al. (2010). Immunity to pre-1950 H1N1 influenza viruses confers cross-protection against the pandemic swine-origin 2009 A (H1N1) influenza virus. J. Immunol. 185:1642–9.
Corti D, et al. (2011). A neutralizing antibody selected from plasma cells that binds to group 1 and group 2 influenza A hemagglutinins. Science. 333:850–6.
Epstein SL, et al. (2005). Protection against multiple influenza A subtypes by vaccination with highly conserved nucleoprotein. Vaccine. 23:5404–10.
Laddy DJ, et al. (2009). Electroporation of synthetic DNA antigens offers protection in nonhuman primates challenged with highly pathogenic avian influenza virus. J. Virol. 83:4624–30.
McMichael AJ, Gotch FM, Dongworth DW, Clark A, Potter CW. (1983). Declining T-cell immunity to influenza, 1977–82. Lancet. 2:762–4.
Topham DJ, Tripp RA, Doherty PC. (1997). CD8+ T cells clear influenza virus by perforin or Fas-dependent processes. J. Immunol. 159:5197–200.
Furuya Y, et al. (2010). Cytotoxic T cells are the predominant players providing cross-protective immunity induced by {gamma}-irradiated influenza A viruses. J. Virol. 84:4212–21.
Brown DM, Dilzer AM, Meents DL, Swain SL. (2006). CD4 T cell-mediated protection from lethal influenza: perforin and antibody-mediated mechanisms give a one-two punch. J. Immunol. 177:2888–98.
Russell SM, Liew FY, (1979). T cells primed by influenza virion internal components can cooperate in the antibody response to haemagglutinin. Nature. 280:147–8.
Scherle PA, Gerhard W. (1986). Functional analysis of influenza-specific helper T cell clones in vivo. T cells specific for internal viral proteins provide cognate help for B cell responses to hemagglutinin. J. Exp. Med. 164:1114–28.
Terajima M, et al. (2008). Influenza A virus matrix protein 1-specific human CD8+ T-cell response induced in trivalent inactivated vaccine recipients. J. Virol. 82:9283–7.
Renfrey S, Watts A. (1994). Morphological and biochemical characterization of influenza vaccines commercially available in the United Kingdom. Vaccine. 12:747–52.
He XS, et al. (2006). Cellular immune responses in children and adults receiving inactivated or live attenuated influenza vaccines. J. Virol. 80:11756–66.
Carrat F, Flahault A. (2007). Influenza vaccine: the challenge of antigenic drift. Vaccine. 25:6852–62.
Monto AS, et al. (2009). Influenza control in the 21st century: optimizing protection of older adults. Vaccine. 27:5043–53.
Osterhaus A, Fouchier R, Rimmelzwaan G. (2011). Towards universal influenza vaccines? Philos. Trans. R. Soc. Lond. B Biol. Sci. 366:2766–73.
Mastelic B, et al. (2010). Mode of action of adjuvants: implications for vaccine safety and design. Biologicals. 38:594–601.
Marrack P, McKee AS, Munks MW, (2009). Towards an understanding of the adjuvant action of aluminium. Nat. Rev. Immunol. 9:287–93.
Czerkinsky C, Holmgren J. (2010). Topical immunization strategies. Mucosal Immunol. 3:545–55.
Khurana S, et al. (2010). Vaccines with MF59 adjuvant expand the antibody repertoire to target protective sites of pandemic avian H5N1 influenza virus. Sci. Transl. Med. 2:15ra5.
Khurana S, et al. (2011). MF59 adjuvant enhances diversity and affinity of antibody-mediated immune response to pandemic influenza vaccines. Sci. Transl. Med. 3:85ra48.
Baldo V, et al. (2010). Immunogenicity of three different influenza vaccines against homologous and heterologous strains in nursing home elderly residents. Clin. Dev. Immunol. 2010:517198.
Schultze V, et al. (2008). Safety of MF59 adjuvant. Vaccine. 26:3209–22.
Schijns VE, Lavelle EC, (2011). Trends in vaccine adjuvants. Expert Rev. Vaccines. 10:539–50.
Turley CB, et al. (2011). Safety and immunogenicity of a recombinant M2e-flagellin influenza vaccine (STF2.4xM2e) in healthy adults. Vaccine. 29:5145–52.
Yusibov V, Streatfield SJ, Kushnir N. (2011). Clinical development of plant-produced recombinant pharmaceuticals: vaccines, antibodies and beyond. Hum. Vaccin. 7:313–21.
Krammer F, Grabherr R. (2010). Alternative influenza vaccines made by insect cells. Trends Mol. Med. 16:313–20.
de Cassan SC, et al. (2011). The requirement for potent adjuvants to enhance the immunogenicity and protective efficacy of protein vaccines can be overcome by prior immunization with a recombinant adenovirus. J. Immunol. 187:2602–16.
Aichele P, Brduscha-Riem K, Zinkernagel RM, Hengartner H, Pircher H. (1995). T cell priming versus T cell tolerance induced by synthetic peptides. J. Exp. Med. 182:261–6.
Smith LR, et al. (2010). Phase 1 clinical trials of the safety and immunogenicity of adjuvanted plasmid DNA vaccines encoding influenza A virus H5 hemagglutinin. Vaccine. 28:2565–72.
Jones S, et al. (2009). DNA vaccination protects against an influenza challenge in a double-blind randomised placebo-controlled phase 1b clinical trial. Vaccine. 27:2506–12.
McConkey SJ, et al. (2003). Enhanced T-cell immunogenicity of plasmid DNA vaccines boosted by recombinant modified vaccinia virus Ankara in humans. Nat. Med. 9:729–35.
Grgacic EVL, Anderson DA. (2006). Virus-like particles: passport to immune recognition. Methods. 40:60–5.
Salvador A, Igartua M, Hernandez RM, Pedraz JL. (2011). An overview on the field of micro- and nanotechnologies for synthetic Peptide-based vaccines. J. Drug Deliv. 2011:181646.
Moser C, Amacker M, Zurbriggen R. (2011). Influenza virosomes as a vaccine adjuvant and carrier system. Expert Rev. Vaccines. 10:437–46.
Draper SJ, et al. (2008). Effective induction of high-titer antibodies by viral vector vaccines. Nat. Med. 14:819–21.
Berthoud TK, et al. (2011). Potent CD8+ T-cell immunogenicity in humans of a novel heterosubtypic influenza A vaccine, MVA-NP+M1. Clin. Infect. Dis. 52:1–7.
Gilbert SC. (2012). T-cell-inducing vaccines — what’s the future. Immunology. 135:19–26.
Rollier CS, Reyes-Sandoval A, Cottingham MG, Ewer K, Hill AVS, (2011). Viral vectors as vaccine platforms: deployment in sight. Curr. Opin. Immunol. 23:377–82.
Geiben-Lynn R, Greenland JR, Frimpong-Boateng K, Letvin NL. (2008). Kinetics of recombinant adenovirus type 5, vaccinia virus, modified vaccinia ankara virus, and DNA antigen expression in vivo and the induction of memory T-lymphocyte responses. Clin. Vaccine Immunol. 15:691–6.
Ronan EO, Lee LN, Beverley PC, Tchilian EZ. (2009). Immunization of mice with a recombinant adenovirus vaccine inhibits the early growth of Mycobacterium tuberculosis after infection. PLoS One. 4:e8235.
Tatsis N, et al. (2007). Adenoviral vectors persist in vivo and maintain activated CD8+ T cells: implications for their use as vaccines. Blood. 110:1916–23.
Hutchison, S, et al. (2011). Antigen depot is not required for alum adjuvanticity. FASEB J. 26:1272–9.
Gonzalez-Fernandez A, Milstein C. (1998). Low antigen dose favours selection of somatic mutants with hallmarks of antibody affinity maturation. Immunology. 93:149–53.
Gerlach C, van Heijst JW, Schumacher TN. (2011). The descent of memory T cells. Ann. N. Y. Acad. Sci. 1217:139–53.
Cubillos-Zapata C, et al. (2011). Differential effects of viral vectors on migratory afferent lymph dendritic cells in vitro predict enhanced immunogenicity in vivo. J. Virol. 85:9385–94.
Kastenmuller W, et al. (2006). Infection of human dendritic cells with recombinant vaccinia virus MVA reveals general persistence of viral early transcription but distinct maturation-dependent cytopathogenicity. Virology. 350:276–88.
Alcock R, et al. (2010). Long-term thermostabilization of live poxviral and adenoviral vaccine vectors at supraphysiological temperatures in carbohydrate glass. Sci. Transi. Med. 2:19ra12.
Robert-Guroff M. (2007). Replicating and non-replicating viral vectors for vaccine development. Curr. Opin. Biotechnol. 18:546–56.
Krilov LR. (2005). Adenovirus infections in the immunocompromised host. Pediatr. Infect. Dis. J. 24:555–6.
Kopecky-Bromberg SA, Palese P. (2009). Recombinant vectors as influenza vaccines. Curr. Top. Microbiol. Immunol. 333:243–67.
Souza APD, Haut L, Reyes-Sandoval A, Pinto AR. (2005). Recombinant viruses as vaccines against viral diseases. Braz. J. Med. Biol. Res. 38:509–22.
Lasaro MO, Ertl HJC. (2009). New insights on adenovirus as vaccine vectors. Mol. Ther. 17:1333–9.
Xiang ZQ, et al. (2006). Chimpanzee adenovirus antibodies in humans, sub-Saharan Africa. Emerg. Infect. Dis. 12:1596–9.
Dudareva M, et al. (2009). Prevalence of serum neutralizing antibodies against chimpanzee adenovirus 63 and human adenovirus 5 in Kenyan Children, in the context of vaccine vector efficacy. Vaccine. 27:3501–4.
Colloca S, et al. (2012). Vaccine vectors derived from a large collection of simian adenoviruses induce potent cellular immunity across multiple species. Sci. Transl. Med. 4:115ra2.
Gao WT, et al. (2006). Protection of mice and poultry from lethal H5N1 avian influenza virus through adenovirus-based immunization. J. Virol. 80:1959–64.
Toro H, et al. (2007). Protective avian influenza in ovo vaccination with non-replicating human adenovirus vector. Vaccine. 25:2886–91.
Hoelscher MA, et al. (2007). New pre-pandemic influenza vaccines: an egg- and adjuvant-independent human adenoviral vector strategy induces long-lasting protective immune responses in mice. Clin. Pharmacol. Ther. 82:665–71.
Brun A, et al. (2008). Antigen delivery systems for veterinary vaccine development: viral-vector based delivery systems. Vaccine. 26:6508–28.
Merchlinsky M, Moss B. (1992). Introduction of foreign DNA into the vaccinia virus genome by in vitro ligation: recombination-independent selectable cloning vectors. Virology. 190:522–6.
Boyle DB, Anderson MA, Amos R, Voysey R, Coupar BE. (2004). Construction of recombinant fowlpox viruses carrying multiple vaccine antigens and immunomodulatory molecules. Biotechniques. 37:104–6, 108–11.
Perkus ME, Piccini A, Lipinskas BR, Paoletti E. (1985). Recombinant vaccinia virus: immunization against multiple pathogens. Science. 229:981–4.
Sutter G, Moss B. (1992). Nonreplicating vaccinia vector efficiently expresses recombinant genes. Proc. Natl. Acad. Sci. U. S. A. 89:10847–51.
Mayr A, Munz E. (1964). [Changes in the vaccinia virus through continuing passages in chick embryo fibroblast cultures]. Zentralbl Bakteriol Orig. 195:24–35.
Breman JG, Henderson DA. (2002). Diagnosis and management of smallpox. N. Engl. J. Med. 346:1300–8.
Webster DP, et al. (2005). Enhanced T cell-mediated protection against malaria in human challenges by using the recombinant poxviruses FP9 and modified vaccinia virus Ankara. Proc. Natl. Acad. Sci. U. S. A. 102:4836–41.
Bejon P, et al. (2006). A phase 2b randomised trial of the candidate malaria vaccines FP9 ME-TRAP and MVA ME-TRAP among children in Kenya. PLoS Clin. Trials. 1:e29.
Minassian AM, etal. (2011). A Phase I study evaluating the safety and immunogenicity of MVA85A, a candidate TB vaccine, in HIV-infected adults. BMJ Open. 1:e000223.
Hanke T, et al. (2005). Biodistribution and persistence of an MVA-vectored candidate HIV vaccine in SIV-infected rhesus macaques and SCID mice. Vaccine. 23:1507–14.
Ramirez JC, Gherardi MM, Rodriguez D, Esteban M. (2000). Attenuated modified vaccinia virus Ankara can be used as an immunizing agent under conditions of preexisting immunity to the vector. J. Virol. 74:7651–5.
Rimmelzwaan GF, Sutter G. (2009). Candidate influenza vaccines based on recombinant modified vaccinia virus Ankara. Expert Rev. Vaccines. 8:447–54.
O’Hara GA, et al. (2012). Clinical assessment of a recombinant simian adenovirus ChAd63: a potent new vaccine vector. J. Infect. Dis. 205:772–81.
Moorthy VS, et al. (2004). Phase 1 evaluation of 3 highly immunogenic prime-boost regimens, including a 12-month reboosting vaccination, for malaria vaccination in Gambian men. J. Infect. Dis. 189:2213–9.
Mooij P, et al. (2008). Differential CD4+ versus CD8+ T-cell responses elicited by different poxvirus-based human immunodeficiency virus type 1 vaccine candidates provide comparable efficacies in primates. J. Virol. 82:2975–88.
Gasteiger G, Kastenmuller W, Ljapoci R, Sutter G, Drexler I. (2007). Cross-priming of cytotoxic T cells dictates antigen requisites for modified vaccinia virus Ankara vector vaccines. J. Virol. 81:11925–36.
Bublot M, et al. (2006). Development and use of fowlpox vectored vaccines for avian influenza. Ann. N. Y. Acad. Sci. 1081:193–201.
Kreijtz JH, et al. (2009). Recombinant modified vaccinia virus Ankara expressing the hemagglutinin gene confers protection against homologous and heterologous H5N1 influenza virus infections in macaques. J. Infect. Dis. 199:405–13.
Kreijtz JH, et al. (2009). Preclinical evaluation of a modified vaccinia virus Ankara (MVA)-based vaccine against influenza A/H5N1 viruses. Vaccine. 27:6296–9.
Kreijtz JH, et al. (2009). MVA-based H5N1 vaccine affords cross-clade protection in mice against influenza A/H5N1 viruses at low doses and after single immunization. PLoS One. 4:e7790.
Hessel A, et al. (2011). Vectors based on modified vaccinia Ankara expressing influenza H5N1 hemagglutinin induce substantial cross-clade protective immunity. PLoS One. 6:e16247.
Poon LL, et al. (2009). Vaccinia virus-based multivalent H5N1 avian influenza vaccines adjuvanted with IL-15 confer sterile cross-clade protection in mice. J. Immunol. 182:3063–71.
Lillie PJ, et al. (2012). A preliminary assessment of the efficacy of a T cell-based influenza vaccine, MVA-NP+M1, in humans. Clin. Infect. Dis. 55:19–25.
Casimiro DR, et al. (2005). Attenuation of simian immunodeficiency virus SIVmac239 infection by prophylactic immunization with DNA and recombinant adenoviral vaccine vectors expressing Gag. J. Virol. 79:15547–55.
Mattapallil JJ, et al. (2006). Vaccination preserves CD4 memory T cells during acute simian immunodeficiency virus challenge. J. Exp. Med. 203:1533–41.
Capone S, et al. (2010). Immune responses against a liver-stage malaria antigen induced by simian adenoviral vector AdCh63 and MVA prime-boost immunization in non-human primates. Vaccine. 29:256–65.
Reyes-Sandoval A, et al. (2010). Prime-boost immunization with adenoviral and modified vaccinia virus Ankara vectors enhances the durability and polyfunctionality of protective malaria CD8(+) T-cell responses. Infect. Immun. 78:145–53.
Douglas AD, et al. (2010). Tailoring subunit vaccine immunogenicity: maximizing antibody and T cell responses by using combinations of adenovirus, poxvirus and protein-adjuvant vaccines against Plasmodium falciparum MSP1. Vaccine. 28:7167–78.
Acknowledgments
T Lambe is an Oxford Martin Fellow and receives financial support from the Medical Research Council, UK, and the Oxford Biomedical Research Centre. The author is grateful to S Gilbert and R O’Donnell for their helpful comments on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, and provide a link to the Creative Commons license. You do not have permission under this license to share adapted material derived from this article or parts of it.
The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
To view a copy of this license, visit (https://doi.org/creativecommons.org/licenses/by-nc-nd/4.0/)
About this article
Cite this article
Lambe, T. Novel Viral Vectored Vaccines for the Prevention of Influenza. Mol Med 18, 1153–1160 (2012). https://doi.org/10.2119/molmed.2012.00147
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.2119/molmed.2012.00147