
Abstract
Ammonia oxidation is the first step in nitrification and was thought to be performed solely by specialized bacteria. The discovery of ammonia-oxidizing archaea (AOA) changed this view. We examined the large scale and spatio-temporal occurrence, abundance and role of AOA throughout Chinese inland waters (n = 28). Molecular survey showed that AOA was ubiquitous in inland waters. The existence of AOA in extreme acidic, alkaline, hot, cold, eutrophic and oligotrophic environments expanded the tolerance limits of AOA, especially their known temperature tolerance to −25 °C and substrate load to 42.04 mM. There were spatio-temporal divergences of AOA community structure in inland waters and the diversity of AOA in inland water ecosystems was high with 34 observed species-level operational taxonomic units (OTUs; based on a 15% cutoff) distributed widely in group I.1b, I.1a and I.1a-associated. The abundance of AOA was quite high (8.5 × 104 to 8.5 × 109 copies g−1) and AOA outnumbered ammonia-oxidizing bacteria (AOB) in the inland waters where little human activities were involved. On the whole AOB predominate the ammonia oxidation rate over AOA in inland water ecosystems and AOA play an indispensable role in global nitrogen cycle considering that AOA occupy a broader habitat range than AOB, especially in extreme environments.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Introduction
The biogeochemical nitrogen cycle is primarily driven by microorganisms with nitrification as the key process. Ammonia oxidation is the rate-limiting step of nitrification1,2 and is catalyzed by ammonia-oxidizing bacteria (AOB) and ammonia-oxidizing archaea (AOA) through ammonia monooxygenase (AMO) as the key enzyme. Studies on AOB date back to over a century ago3 and the knowledge about their physiology, ecology and genomics is extensive4,5,6,7. AOA were discovered recently with the identification of genes predicted to encode AMO in the phylum Crenarchaeota8,9, which aroused global scientific interest and stimulated the reassessment of the role of different ammonia oxidizers in the nitrification process.
Archaea capable of oxidizing ammonia to nitrite have been assigned to a novel phylum named Thaumarchaeota10,11, which separates them from the phyla Euryarchaeota and Crenarchaeota. They are found widely distributed in various natural environments4 and have considerable diversity12. Their physiology, ecology and genomics in marine and soil ecosystems have been widely studied12,13,14. However, gaps in our understanding of AOA still exist2,15,16. The behaviors and biogeographical distribution of AOA in inland water ecosystems are not well characterized, especially not through large-scale studies. Although the significance of AOA in ecology and microbiology has been established17, the specific organisms responsible for ammonia oxidation remain to be identified.
The objectives of this study are to investigate the occurrence, biodiversity and distribution of AOA in inland water ecosystems through multiple samples, including those with extreme conditions, to broaden the known tolerance limits of AOA and to examine their ecological features. This study will also test the role and influence of AOA on ammonia oxidation and estimate their ecological potential in the aquatic nitrogen cycle. To achieve this, studies were performed in a wide range of inland water ecosystems throughout China. The abundance and community structure of AOA were analyzed using molecular methods and the nitrification rates were measured through incubation experiments.
Results
Ubiquity of AOA in inland waters
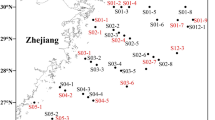
The presence of AOA in Chinese inland water ecosystems was investigated with a PCR survey of archaeal amoA genes resulting in the amplification of 635-bp fragments. PCR screening showed that AOA existed in all of the 100 sediment samples from the 28 sites, which verified the widespread occurrence of AOA in inland water ecosystems (Fig. 1). It is notable that AOA were still existed under extreme environmental conditions, including extremely high and low temperatures (75 °C and −25 °C), strong acidity and alkalinity (pH as low as 3.9 and up to 8.9) and oligotrophic and eutrophic conditions (Table 1).

Biogeographical distribution of sampling sites in Chinese inland water ecosystems.
Sites 1 to 14 were in order of longitude and sites 15 to 28 were in order of latitude as listed in Table 1. Different colors represent different types of inland waters as shown in the legend. The map were come from web of “Data Sharing Infrastructure of Earth System Science” http://www.geodata.cn. All of the maps used in the manuscript are free.
Community structure and diversity of AOA in inland waters
We examined the diversity of AOA and differences among AOA community structures in the inland water ecosystems. A database of 729 archaeal amoA gene sequences from this study was constructed and the community structures of AOA were investigated. After the diversity analysis using DOTUR software, a total of 216 unique OTUs (based on a 2% cutoff) were recovered. The observed Chao1 richness estimate and Shannon diversity index were as high as 324.90 and 4.88, indicating a high biodiversity of AOA.
Thirty-four species-level OTUs (each representing a specific AOA species), were obtained from our sequences, using 85% amoA gene sequence identity as a species threshold18. The evolutionary relationships between these species and published amoA gene sequences are shown in Fig. 2. The AOA species in Chinese inland water ecosystems were widely distributed in three AOA lineages (group I.1b, I.1a and I.1a-associated), which are the known lineages of AOA together with the ThAOA (Thermophilic AOA) group13. Most of the sequences clustered into the group I.1b (28 species representing 563 sequences), while a small quantity were affiliated with the group I.1a (4 species representing 135 sequences) and the remaining two species (representing 31 sequences) were assigned to the group I.1a-associated.

Phylogenetic tree showing the spatial divergences of AOA community structure among various inland waters.
Phylogenetic trees were constructed with the neighbor-joining method using Maximum Composite Likelihood with 1000 bootstraps. The scale bar represents 5% of the sequence divergence. Pie charts for each species-level OTU show the composition of sequences from different origins with the colors corresponding to the legend.
Among the 34 species obtained in this study, three were found to be closely related to Candidatus Nitrososphaera gargensis (species-3), Nitrososphaera viennensis EN76 (species-7) and Ca. Nitrosotalea devanaterra (species-30) respectively and one (species-33) was similar to both Ca. Nitrosoarchaeum limnia SFB1 and Ca. Nitrosoarchaeum koreensis MY1 (with identities >85.0% on the nucleotide level). This indicates that the above five known species of AOA exist in Chinese inland water ecosystems. The other 30 AOA species found in inland water sediments had not been previously characterized, which reminds us that our knowledge of AOA in natural environments is incomplete and more unidentified AOA species probably exist13.
Most of our sampled inland waters harbored species-12 (24 inland waters) and species-3 (21 inland waters). 18 species existed in three or fewer of the sampled inland waters and 11 species were specific to a particular sampling site (Fig. 2). Although some AOA species were capable of living in a broad range of environments, most of them were selective and quite a few were sensitive to different environments. The distribution of AOA species in inland water ecosystems was nonrandom on a geographical scale. As a result, the community structure of AOA varied greatly among inland waters (Supplementary Table S1). This is also a reason for the high biodiversity of AOA in inland water ecosystems.
Besides the spatial distribution of AOA in Chinese inland water ecosystems, the temporal distribution of AOA was studied in the Pearl River. The AOA community structure in sediments of the Pearl River in winter and summer were analyzed (Fig. 3). Results showed that a population shift in AOA over different seasons occurred in the Pearl River, with most of the AOA in summer clustered into group I.1b, while winter AOA species distributed evenly in group I.1a and I.1b. In summary, this study documented the spatio-temporal divergences of AOA community structure.

Phylogenetic tree showing the temporal divergences of AOA community structure in the Pearl River.
Phylogenetic trees were constructed with the neighbor-joining method using Maximum Composite Likelihood with 1000 bootstraps. The scale bar represents 5% of the sequence divergence. Numbers in the parentheses after each species in red give the numbers of sequences obtained in summer and blue give those in winter.
AOA in extreme environments
pH
Species-20 and 30 were detected in the sediments of the Tieshanping River with a pH as low as 3.9 (Table 2), indicating the strong acidity tolerance of these two species. Species-30 was identified as Ca. Nitrosotalea devanaterra, which has been detected in acidic soil with a pH of 4.5 and incubated under a pH of 4.0-5.519. This study extended their tolerance limit to pH of 3.9. Ca. Nitrosotalea devanaterra (Species-30) was found to dominate the AOA population (75%) in the sediments of the Tieshanping River, with the remaining population belonging to species-20. In the soil of the Jiaxing paddy field with a pH of 6.55, Ca. Nitrosotalea devanaterra were also detected, accounting for a small part of the AOA population (20%). In other inland waters with a higher pH, no Ca. Nitrosotalea devanaterra were detected. The results provided evidence that Ca. Nitrosotalea devanaterra may exist only in acidic and neutral environments. The simple composition of the AOA population in the sediments of the Tieshanping River also reflected that only a few specialized AOA species could survive in environments with extreme acidity.
Eight AOA species were detected in alkaline sediments with a pH up to 8.9 (Table 2), including Ca. Nitrososphaera gargensis, N. viennensis EN76, Ca. Nitrosoarchaeum limnia SFB1 and Ca. Nitrosoarchaeum koreensis MY1, indicating their strong tolerance to alkalinity. N. viennensis EN76 and Ca. Nitrosoarchaeum koreensis MY1 were incubated under pH up to 9 and 820,21 and Ca. Nitrososphaera gargensis and Ca. Nitrosoarchaeum limnia SFB1 were detected in environments of pH of 7.9 and 7.0–7.213,22. This study expands their alkalinity tolerance to approximately 9. Unlike the AOA community structure in an acidic environment (Tieshanping River), that in the alkaline environment (Tarim River) was complex and spread widely between group I.1a (31.8%, 2 species) and I.1b (68.2%, 6 species).
Substrate
The sediment samples in this study had a wide gradient of ammonium concentrations from 0.10 mM (Tianchi Lake), to 42.04 mM (Baiyangdian Lake). Five uncharacterized AOA species belonging to group I.1b were detected in the oligotrophic Tianchi Lake (Table 2), indicating that these five species can live in environments with very low substrate concentration (0.10 mM).
Ca. Nitrososphaera gargensis, N. viennensis EN76 and four other species were observed in the sediments of Baiyangdian Lake (Table 2), indicating their strong tolerance to high ammonium concentration (up to 42.04 mM). Ca. Nitrososphaera gargensis was detected in environments with ammonium concentration of 5.6 mM22 and N. viennensis EN76 was incubated under ammonium concentration up to 15 mM20. This study expands their tolerance to high substrate content to 42.04 mM.
Temperature
Eight AOA species were observed in Aydingkol Lake with a high surface temperature (up to 75 °C) and six in the Songhua River with a low temperature (as low as −25 °C) (Table 2). Species-12 and 17, Ca. Nitrosoarchaeum limnia and Ca. Nitrosoarchaeum koreensis existed in both of the extreme-temperature environments, which expands their recognized temperature tolerance to between −25 °C and 75 °C. In addition, the moderately thermophilic Ca. Nitrososphaera gargensis, which had been detected in thermal spring microbial mats at 45 °C22, was also observed in Aydingkol Lake, which extends their known heat-tolerance to 75 °C. Previous studies have demonstrated the distribution of AOA in a broad range of temperature from 0.2 to 97 °C23,24,25. This study expands the lower limit of this range to −25 °C.
Abundance of archaeal and bacterial ammonia oxidizers
Results of qPCR showed that the abundances of AOA ranged from 8.5 × 104 to 8.5 × 109 copies g−1 dry sediment in Chinese inland waters. As the counterpart of AOA, AOB had the abundance of 2.9 × 103 to 4.3 × 109 copies g−1 (Fig. 4A). The relative abundance of AOA compared with AOB varied throughout the sampling sites (Fig. 4). and AOA outnumbered AOB in almost all the lakes and rivers, while the opposite was true in the other inland waters with strong human activities such as paddy fields, reservoirs, polluted groundwater, tidal land and constructed wetland. This result implies that AOA predominates the ammonia oxidizer population in inland waters with less human activities, while AOB dominates in inland waters with more human disturbance.

(A) The abundance of archaeal & bacterial amoA genes and potential nitrification rate (PNR) in various Chinese inland water ecosystems. (B) The abundance of archaeal & bacterial amoA genes in different types of inland waters. Boxes give the 25th and 75th percentiles; whiskers show the range from 1th to 99th percentiles; horizontal lines in and out the boxes represent the medians and maximum/minimum values respectively; little squares give the averages.
Spearman correlation analysis between archaeal & bacterial amoA abundance and environmental variables revealed that different parameters were related to the size of the AOA and AOB population, indicating a niche differentiation between these two groups. Archaeal amoA abundance had an obvious negative correlation with TC (r = −0.755, p < 0.01) and TS (r = −0.748, p < 0.01), while bacterial amoA abundance had an obvious positive correlation with NOx– content (r = 0.497, p < 0.01), TN (r = 0.529, p < 0.01) and TS (r = 0.484, p < 0.01) (Supplementary Table S2).
Contributions of AOA and AOB to microbial ammonia oxidation
Because AOA were found to be ubiquitous and abundant in Chinese inland water ecosystems, we expected that they would play a significant role in ammonia oxidation. To examine this assumption, potential nitrification rates (PNRs) were measured to estimate the combined activity of archaeal and bacterial ammonia oxidizers. The values ranged from 0 to 146.91 nmol N g−1 h−1 (n = 72, Fig. 4A). Spearmen correlation analysis between PNR, archaeal & bacterial amoA abundance and environmental variables showed bacterial amoA abundance and pH were significantly correlated with PNR (Supplementary Table S2). The archaeal amoA abundance showed no correlation with PNR. Multiple linear regression (stepwise regression) on PNR also showed that bacterial amoA abundance was the most determining variable for nitrification followed by pH (bacterial amoA abundance explained 29.5% of the variability of PNR, while pH explained 3.2%, n = 45, Supplementary Table S3). These results led to a possibility that AOB, rather than AOA, contribute more to nitrification in Chinese inland water ecosystems.
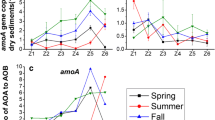
This hypothesis was further tested in the Tiaoxi River on a spatio-temporal scale. Sediment samples from different seasons were collected from four sites in the Tiaoxi River (Supplementary Fig. S1) under different ammonia loading levels (Supplementary Table S4). The community structure of AOA (Fig. 5A) and abundance of archaeal & bacterial amoA genes (Fig. 5B) were detected. AOA from different sites showed quite different community structures, although species-12 and 25 existed in all of the four sites. AOA outnumbered AOB in almost all of the sediment samples, while the correlation analysis showed that the abundance of AOB not AOA had a significant correlation with PNR (Supplementary Table S5), indicating that AOB might contribute more to ammonia oxidation than AOA on spatio-temporal scale.

The spatio-temporal variation of AOA & AOB abundance, PNR and AOA populations in four sites on the Tiaoxi River.
(A) Phylogenetic tree of the archaeal amoA gene sequences from sediments in the Tiaoxi River. The letters ABCD in the sequence names are used to distinguish sequences from different sites, marked by circles with different colors. The phylogenetic tree was constructed with the neighbor-joining method using Maximum Composite Likelihood with 1000 bootstraps. The scale bar represents 5% of the sequence divergence. (B)The seasonal variance in archaeal & bacterial amoA gene abundance and PNR in sediments from the Tiaoxi River. The ratios of archaeal amoA gene abundance to bacterial are listed above the diagrams. Error bars indicate standard deviation (n = 3).
Discussion
To the best of our knowledge, this is the first report of the large-scale occurrence, ecological behaviors, biodiversity and potential function of AOA in inland waters. They were found to be ubiquitous, have high biodiversity and diverse community structure, have strong adaptability to extreme conditions, abundant and play a less important role in ammonia oxidation than AOB in inland water ecosystems.
The ubiquity of AOA in inland waters was demonstrated through sediment samples (n = 100) from 28 inland waters, including six sites with extreme pH, temperature or substrate conditions. The ubiquity of AOA may be explained by their high diversity and strong tolerance to extreme conditions. High diversity helps increase the capacity to adapt to environmental change and the strong tolerance to extreme conditions extends the occurrence to a largest scope1,26.
The similar phenomenon also appears in other ecosystems. AOA is also ubiquitous in soil ecosystems with quite high diversity27. There are few studies focused on the biodiversity of AOA in marine ecosystems on a large scale, but the observed unique archaeal amoA sequences specific to an individual sample location has indicated the high biodiversity of AOA in marine ecosystems28. For the whole natural environment, the considerable global AOA diversity was observed in a phylogenetic analysis on 12356 publicly available archaeal amoA sequences from different ecosystems18. These results demonstrate the high diversity of AOA in natural environments. As to the adaptability to extreme conditions, AOA were found in a wide pH range (2.5 to 9.024,29) and temperature range (0.2 to 97 °C23,24,25), indicating the great potential of AOA in adapting to extreme acidity and alkalinity and extreme low and high temperatures. In addition, AOA had a preference for low ammonium content30,31 and survived extreme low ammonium concentrations (≤10 nM20) with half-saturation constant (Km) of 133 nM total NH4+32. The detection of AOA in extreme conditions in this study broadened their known limits, especially those of identified species (Table 2). The cold-tolerance of AOA was expanded to −25 °C and the tolerance to high substrate content was extended to 42.04 mM. The results point to a function of AOA in extending ammonia oxidation to a much greater range of habitats. As a result, AOA are widely distributed in marine, soil and inland water environments1,13,33 and exist in extreme conditions such as hot springs34, Antarctic waters35, acid soils19 and oligotrophic environments32. High AOA diversity imply a strong capacity for ammonia oxidation in various ecosystems and an important role in global nitrification.
In this study, AOA were found to numerically outcompete their counterpart AOB in inland waters with less human disturbance, while AOB dominated in inland waters affected strongly by human activities. Evidence can also be found in previous studies. AOA appear to be numerically dominant in marine environments36,37,38 and soils17,39,40,41, while in environments involved with more human activities like fertilized agricultural soils42,43,44, polluted wetlands2, wastewater treatment plants45 and bioreactors46,47, AOB outnumbered AOA. The environmental changes caused by the human activities seem to create a position of advantage for AOB over AOA. This also demonstrated the niche differentiation between AOA and AOB. The factors influencing the relative abundance of AOA to AOB were various, including the ammonia concentration20,48,49, the dissolved oxygen concentration50 and the pH49. Considering the complex status of various ecosystems, the relative predominance of AOA and AOB may be not affected solely by a single parameter, but by a combination of influencing factors51.
With the increasing understanding of AOA, the function of AOA & AOB in ammonia oxidation was called into question. This study, based on a large number of samples, documented that AOB contributed more to nitrification than AOA in Chinese inland water ecosystems. Even so, the function of AOA can’t be ignored, especially in extreme environments. In other small-scale studies, different results have been obtained. For example, nitrification was suggested be driven by bacteria rather than archaea in six nitrogen-rich grassland soils in New Zealand30 and in sediments of four nitrogen-rich wetlands2. In six estuarine sediments, PNRs increased as the abundance of archaeal amoA increased, rather than bacterial amoA52 and in the Black Sea water column nitrification was mainly controlled by archaeal amoA expression in the lower oxic zone53. The relative importance of AOA vs. AOB in ammonia oxidation needs more researches. The in situ measurement of microbial ammonia oxidation rate and separation of the role of AOA & AOB are two key items. In this study we can make a conclusion that AOA may play an indispensable role in global nitrogen cycle considering that AOA occupy a broader habitat range than AOB, especially in extreme environments.
Methods
Study site background
A total of 100 sediment samples from 28 inland water ecosystems, including lakes, rivers, paddy fields, reservoirs, groundwater, swamp, tidal land and constructed wetland were investigated in Chinese territory (23 to 46° N and 86 to 130° E). Details for every sampling site including location, background and number of samples are listed in Table 1. Other information including the nitrogenous compounds content and some physicochemical characteristics of the sampling sites is listed in Supplementary Table S6.
Among the sampled inland waters, there were six with extreme conditions. Aydingkol Lake had the highest surface temperature (75 °C) and the Songhua River had the lowest temperature (−25 °C). Tianchi Lake was an oligotrophic lake (NH4+ as low as 0.10 mM) and Baiyangdian Lake was hyper-eutrophic (NH4+ up to 42.04 mM). The sediment in Tieshanping River had an acidic pH (as low as 3.9) and Tarim River had an alkaline pH (up to 8.9). These extreme conditions were all relatively stable. Furthermore, sediments from the Pearl River and the Tiaoxi River were sampled in different seasons to verify the results on a spatio-temporal scale.
Surface sediments (0–8 cm) were collected from each sampling site in 2012 and 2013. The samples were placed in sterile plastic bags, sealed and transported to the laboratory on ice. One part of each sample was used for the analysis of physicochemical characteristics immediately after arrival, one part was incubated to measure the potential nitrification rates (PNRs) and the rest was stored at −80 °C for later DNA extraction and molecular analysis.
DNA Extraction, PCR, Cloning and Sequences Analysis
DNA was extracted from about 0.3 g sediment using FastDNA® Spin Kit for Soil (MP Biomedicals, USA). Concentrations of the extracted DNA were determined by spectrophotometric analysis on a NanoDrop 2000 UV-Vis Spectrophotometer (Thermo Fisher Scientific, USA) and the quality was checked by electrophoresis on a 1% (weight/volume percent) agarose gel. The archaeal amoA (ammonia monooxygenase α-subunit) gene was amplified using primer pairs Archaea-amoAF/Archaea-amoAR according to Francis et al.28 with an annealing temperature of 53 °C. The sequences of primers and thermal profiles used in this study were shown in Supplementary Table S7. All PCR reactions were performed with the Ex TaqTM polymerase (Takara Dalian, China).
PCR amplified fragments were ligated directly using the pGEM-T® Easy Vector Systems (Promega, USA) according to the manufacturer’s instructions and then transformed to Escherichia coli JM109 competent cells for cloning. Selected clones were sequenced using T7 forward primers targeting vector sequences adjacent to the multiple cloning sites by an ABI PRISM 3730XL automated-sequencer (Applied Biosystems, USA). Sequences of archaeal amoA genes obtained in this study were deposited in the GenBank under the accession numbers (HM637849-HM637867, HQ538539-HQ538560, HQ538562, JF439021, JF439023-JF439028, JF439030-JF439044, JF439046-JF439066, KC108794-KC108815, KP167639-KP168260). All the sequences were aligned using the ClustalX 1.83 program. Phylogenetic analysis was performed using Mega 5.0 software54. Phylogenetic trees were constructed by neighbor-joining (NJ) with the Maximum Composite Likelihood and the robustness of tree topology was tested by bootstrap analysis with 1,000 replicates. The calculation of operational taxonomic unit (OTU) and diversity indices, including Chaol, richness estimate and Shannon diversity index, were generated by DOTUR by employing the furthest neighbor approach55.
Quantitative PCR analysis
Quantitative PCR (qPCR) was performed on an ABI 7300 real-time PCR instrument (Applied Biosystems, USA) with a SYBR Green qPCR kit (Takara Dalian, China). The qPCR thermal profiles of archaeal and bacterial amoA genes were performed with primers Archaea-amoAF/Archaea-amoAR and amoA-1F/amoA-2R7, with the annealing temperatures of 53 °C and 55 °C, respectively. The detailed information on sequences of primers and thermal profiles were listed in Supplementary Table S7. Positive clones of archaeal & bacterial amoA gene were selected to isolate plasmids using Wizard® Plus Minipreps DNA Purification System (Promega, USA) and standard curves were obtained with 10-fold serial dilutions of plasmid DNA containing the target genes. The results with amplification efficiency and correlation coefficient above 95% and 0.98 were employed.
Analytical Procedures of Environmental Variables
Ammonium (NH4+), nitrate plus nitrite (NOx-) were extracted from the fresh sediment samples with 2 M KCl solution and measured using a Continuous Flow Analyzer (SAN plus, Skalar Analytical, the Netherlands). The other physicochemical characteristics (total nitrogen (TN), total carbon (TC), total sulfur (TS), total phosphorus (TP)) of the sediment samples were also measured according to Bao56. The pH was determined using dry sediments after mixing with water at a ratio (dry sediment/water) of 1:5 and the organic matter was determined by K2Cr2O7 oxidation method. All analyses were performed on triplicate samples.
Potential nitrification rates (PNRs)
The potential nitrification rate was measured using a chlorate inhibition method with minor modifications57. Briefly, 3.0 g of fresh sediment was added to 50-mL centrifuge tube containing 20 mL phosphate buffer solution (NaCl 8.0 g L−1, KCl 0.2 g L−1, Na2HPO4 0.2 g L−1, NaH2PO4 0.2 g L−1, pH = 7.4). (NH4)2SO4 was added to the incubation system to a final concentration of ammonium similar to the in situ condition. Samples were run in triplicate with and without allylthiourea (ATU, an inhibitor of nitrification process) (100 μM final concentration) to identify the difference between NO2− accumulation by aerobic microbial nitrification and chemical ammonia oxidation58. Potassium chlorate (KClO3) with a final concentration of 10 mg L−1 was added to inhibit the nitrite oxidation. The suspension was incubated (150 rpm) in the dark at in situ temperature for 1 h, 4 h, 8 h, 12 h and 24 h, respectively. Three subsamples were taken at each time point. The produced nitrite was extracted with 5 mL of 2 M KCl solution and determined by a spectrophotometer at wave length of 540 nm with N-(1-naphthyl) ethylenediamine dihydrochloride. The nitrification rate was calculated from the linear increase of NO2– concentration in suspension.
Data analysis
The statistical analyses were conducted by PASW Statistics 18.0 (Predictive Analytics Software Statistics). Mann-Whitney U test and non-parameter paired test were used respectively for the comparison of two data groups and paired data. The correlations between different types of variables were computed by Spearman correlation analysis. Stepwise linear regression analysis was used to determine the most important factor for a dependent variable. Unless otherwise specified, the level of significance in this study was α = 0.05. Graphing was achieved using Origin 8.0 software.
Additional Information
How to cite this article: Zhou, L. et al. Species, Abundance and Function of Ammonia-oxidizing Archaea in Inland Waters across China. Sci. Rep. 5, 15969; doi: 10.1038/srep15969 (2015).
References
Stahl, D. A. & de la Torre, J. R. Physiology and diversity of ammonia-oxidizing archaea. Annu. Rev. Microbiol. 66, 83–101 (2012).
Wang, S., Wang, Y., Feng, X., Zhai, L. & Zhu, G. Quantitative analyses of ammonia-oxidizing Archaea and bacteria in the sediments of four nitrogen-rich wetlands in China. Appl. Microbiol. Biotechnol. 90, 779–787 (2011).
Winogradsky, S. Recherches sur les organismes de la nitrification. Ann. Inst. Pasteur 4, 11–12 (1890).
Monteiro, M., Séneca, J. & Magalhães, C. The history of aerobic ammonia oxidizers: from the first discoveries to today. J. Microbiol. 52, 537–547 (2014).
Kowalchuk, G. A. & Stephen, J. R. Ammonia-oxidizing bacteria: a model for molecular microbial ecology. Annu. Rev. Microbiol. 55, 485–529 (2001).
Arp, D. J., Chain, P. S. & Klotz, M. G. The impact of genome analyses on our understanding of ammonia-oxidizing bacteria. Annu. Rev. Microbiol. 61, 503–528 (2007).
Rotthauwe, J. H., Witzel, K. P. & Liesack, W. The ammonia monooxygenase structural gene amoA as a functional marker: molecular fine-scale analysis of natural ammonia-oxidizing populations. Appl. Environ. Microbiol. 63, 4704–4712 (1997).
Venter, J. C. et al. Environmental genome shotgun sequencing of the Sargasso Sea. Science 304, 66–74 (2004).
Treusch, A. H. et al. Novel genes for nitrite reductase and Amo-related proteins indicate a role of uncultivated mesophilic crenarchaeota in nitrogen cycling. Environ. Microbiol. 7, 1985–1995 (2005).
Brochier-Armanet, C., Boussau, B., Gribaldo, S. & Forterre, P. Mesophilic Crenarchaeota: proposal for a third archaeal phylum, the Thaumarchaeota. Nat. Rev. Microbiol. 6, 245–252 (2008).
Spang, A. et al. Distinct gene set in two different lineages of ammonia-oxidizing archaea supports the phylum Thaumarchaeota. Trends Microbiol. 18, 331–340 (2010).
Walker, C. et al. Nitrosopumilus maritimus genome reveals unique mechanisms for nitrification and autotrophy in globally distributed marine crenarchaea. Proc. Natl. Acad. Sci. USA. 107, 8818–8823 (2010).
Hatzenpichler, R. Diversity, physiology and niche differentiation of ammonia-oxidizing archaea. Appl. Environ. Microbiol. 78, 7501–7510 (2012).
James, I. P. & Graeme, W. N. Archaeal and bacterial ammonia oxidisers in soil: the quest for niche specialisation and differentiation. Trends Microbiol. 20, 523–531 (2012).
Wang, C., Zhu, G., Wang, W. & Yin, C. Preliminary study on the distribution of ammonia oxidizers and their contribution to potential ammonia oxidation in the plant-bed/ditch system of a constructed wetland. J. Soils Sed. 13, 1626–1635 (2013).
Wang, Y., Zhu, G., Song, L., Wang, S. & Yin, C. Manure fertilization alters the population of ammonia-oxidizing bacteria rather than ammonia-oxidizing archaea in a paddy soil. J. Basic Microbiol. 54, 190–197 (2014).
Prosser, J. I. & Nicol, G. W. Relative contributions of archaea and bacteria to aerobic ammonia oxidation in the environment. Environ. Microbiol. 10, 2931–2941 (2008).
Pester, M. et al. amoA-based consensus phylogeny of ammonia-oxidizing archaea and deep sequencing of amoA genes from soils of four different geographic regions. Environ. Microbiol. 14, 525–539 (2012).
Lehtovirta-Morley, L. E., Stoecker, K., Vilcinskas, A., Prosser, J. I. & Nicol, G. W. Cultivation of an obligate acidophilic ammonia oxidizer from a nitrifying acid soil. Proc. Natl. Acad. Sci. USA 108, 15892–15897 (2011).
Tourna, M. et al. Nitrososphaera viennensis, an ammonia oxidizing archaeon from soil. Proc. Natl. Acad. Sci. USA 108, 8420–8425 (2011).
Jung, M.-Y. et al. Enrichment and characterization of an autotrophic ammonia-oxidizing archaeon of mesophilic crenarchaeal group I. 1a from an agricultural soil. Appl. Environ. Microbiol. 77, 8635–8647 (2011).
Lebedeva, E. V. et al. Moderately thermophilic nitrifying bacteria from a hot spring of the Baikal rift zone. FEMS Microbiol. Ecol. 54, 297–306 (2005).
Nakagawa, T., Mori, K., Kato, C., Takahashi, R. & Tokuyama, T. Distribution of cold-adapted ammonia-oxidizing microorganisms in the deep-ocean of the northeastern Japan Sea. Microbes Environ. 22, 365–372 (2007).
Reigstad, L. J. et al. Nitrification in terrestrial hot springs of Iceland and Kamchatka. FEMS Microbiol. Ecol. 64, 167–174 (2008).
Erguder, T. H., Boon, N., Wittebolle, L., Marzorati, M. & Verstraete, W. Environmental factors shaping the ecological niches of ammonia-oxidizing archaea. FEMS Microbiol. Rev. 33, 855–869 (2009).
Hatzenpichler, R. Diversity, physiology and niche differentiation of ammonia-oxidizing archaea. Appl. Environ. Microbiol. 78, 7501–7510 (2012).
Gubry-Rangin, C. et al. Niche specialization of terrestrial archaeal ammonia oxidizers. Proc. Natl. Acad. Sci. USA 108, 21206–21211 (2011).
Francis, C. A., Roberts, K. J., Beman, J. M., Santoro, A. E. & Oakley, B. B. Ubiquity and diversity of ammonia-oxidizing archaea in water columns and sediments of the ocean. Proc. Natl. Acad. Sci. USA. 102, 14683–14688 (2005).
Zhang, C. L. et al. Global occurrence of archaeal amoA genes in terrestrial hot springs. Appl. Environ. Microbiol. 74, 6417–6426 (2008).
Di, H. et al. Nitrification driven by bacteria and not archaea in nitrogen-rich grassland soils. Nature Geoscience 2, 621–624 (2009).
Verhamme, D. T., Prosser, J. I. & Nicol, G. W. Ammonia concentration determines differential growth of ammonia-oxidising archaea and bacteria in soil microcosms. ISME J. 5, 1067–1071 (2011).
Martens-Habbena, W., Berube, P. M., Urakawa, H., de La Torre, J. R. & Stahl, D. A. Ammonia oxidation kinetics determine niche separation of nitrifying Archaea and Bacteria. Nature 461, 976–979 (2009).
Offre, P., Kerou, M., Spang, A. & Schleper, C. Variability of the transporter gene complement in ammonia-oxidizing archaea. Trends Microbiol. (2014).
De La Torre, J. R., Walker, C. B., Ingalls, A. E., Könneke, M. & Stahl, D. A. Cultivation of a thermophilic ammonia oxidizing archaeon synthesizing crenarchaeol. Environ. Microbiol. 10, 810–818 (2008).
Grzymski, J. J. et al. A metagenomic assessment of winter and summer bacterioplankton from Antarctica Peninsula coastal surface waters. ISME J. 6, 1901–1915 (2012).
Wuchter, C. et al. Archaeal nitrification in the ocean. Proc. Natl. Acad. Sci. USA 103, 12317–12322 (2006).
Santoro, A. E., Casciotti, K. L. & Francis, C. A. Activity, abundance and diversity of nitrifying archaea and bacteria in the central California Current. Environ. Microbiol. 12, 1989–2006 (2010).
Beman, J. M., Popp, B. N. & Francis, C. A. Molecular and biogeochemical evidence for ammonia oxidation by marine Crenarchaeota in the Gulf of California. ISME J 2, 453–453 (2008).
Zeglin, L. H., Taylor, A. E., Myrold, D. D. & Bottomley, P. J. Bacterial and archaeal amoA gene distribution covaries with soil nitrification properties across a range of land uses. Environ. Microbiol. Rep. 3, 717–726 (2011).
Leininger, S. et al. Archaea predominate among ammonia-oxidizing prokaryotes in soils. Nature 442, 806–809 (2006).
Adair, K. & Schwartz, E. Evidence that ammonia-oxidizing archaea are more abundant than ammonia-oxidizing bacteria in semiarid soils of Northern Arizona, USA. Microb. Ecol. 56, 420–426 (2008).
Zhu, G. et al. Anammox bacterial abundance, biodiversity and activity in a constructed wetland. Environ. Sci. Technol. 45, 9951–9958 (2011).
Jia, Z. & Conrad, R. Bacteria rather than Archaea dominate microbial ammonia oxidation in an agricultural soil. Environ. Microbiol. 11, 1658–1671 (2009).
Sun, R., Guo, X., Wang, D. & Chu, H. Effects of long-term application of chemical and organic fertilizers on the abundance of microbial communities involved in the nitrogen cycle. Appl. Soil Ecol. 95, 171–178 (2015).
Gao, J. F., Luo, X., Wu, G. X., Li, T. & Peng, Y. Z. Quantitative analyses of the composition and abundance of ammonia-oxidizing archaea and ammonia-oxidizing bacteria in eight full-scale biological wastewater treatment plants. Bioresour. Technol. 138, 285–296 (2013).
Jin, T., Zhang, T. & Yan, Q. Characterization and quantification of ammonia-oxidizing archaea (AOA) and bacteria (AOB) in a nitrogen-removing reactor using T-RFLP and qPCR. Appl. Microbiol. Biotechnol. 87, 1167–1176 (2010).
Wells, G. F. et al. Ammonia-oxidizing communities in a highly aerated full-scale activated sludge bioreactor: betaproteobacterial dynamics and low relative abundance of Crenarchaea. Environ. Microbiol. 11, 2310–2328 (2009).
Herrmann, M., Scheibe, A., Avrahami, S. & Küsel, K. Ammonium availability affects the ratio of ammonia-oxidizing bacteria to ammonia-oxidizing archaea in simulated creek ecosystems. Appl. Environ. Microbiol. 77, 1896–1899 (2011).
Trias, R. et al. Emergent macrophytes act selectively on ammonia-oxidizing bacteria and archaea. Appl. Environ. Microbiol. 78, 6352–6356 (2012).
Abell, G. C. et al. Effects of estuarine sediment hypoxia on nitrogen fluxes and ammonia oxidizer gene transcription. FEMS Microbiol. Ecol. 75, 111–122 (2011).
Yao, H. et al. Multi-factorial drivers of ammonia oxidizer communities: evidence from a national soil survey. Environ. Microbiol. 15, 2545–2556 (2013).
Caffrey, J. M., Bano, N., Kalanetra, K. & Hollibaugh, J. T. Ammonia oxidation and ammonia-oxidizing bacteria and archaea from estuaries with differing histories of hypoxia. ISME J. 1, 660–662 (2007).
Lam, P. et al. Linking crenarchaeal and bacterial nitrification to anammox in the Black Sea. Proc. Natl. Acad. Sci. USA 104, 7104–7109 (2007).
Tamura, K. et al. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance and maximum parsimony methods. Mol. Biol. Evol. 28, 2731–2739 (2011).
Schloss, P. D. & Handelsman, J. Introducing DOTUR, a computer program for defining operational taxonomic units and estimating species richness. Appl. Environ. Microbiol. 71, 1501–1506 (2005).
Bao, S. Chemical Analysis for Agricultural Soil (China Agriculture Press, 2000).
Kurola, J., Salkinoja-Salonen, M., Aarnio, T., Hultman, J. & Romantschuk, M. Activity, diversity and population size of ammonia-oxidising bacteria in oil-contaminated landfarming soil. FEMS Microbiol. Lett. 250, 33–38 (2005).
Lam, P. et al. Linking crenarchaeal an d bacterial nitrification to anammox in the Black Sea. Proc. Natl. Acad. Sci. USA 104, 7104–7109 (2007).
Hatzenpichler, R. et al. A moderately thermophilic ammonia-oxidizing crenarchaeote from a hot spring. Proc. Natl. Acad. Sci. USA 105, 2134–2139 (2008).
Mosier, A. C., Lund, M. B. & Francis, C. A. Ecophysiology of an ammonia-oxidizing archaeon adapted to low-salinity habitats. Microb. Ecol. 64, 955–963 (2012).
Acknowledgements
This research is financially supported by the National Natural Science Foundation of China (No. 41322012 and 21277156), Strategic Priority Research Program of the Chinese Academy of Sciences (XDB15020303), Special Fund of State Key Joint Laboratory of Environment Simulation and Pollution Control (14L01ESPC) and Open Research Fund of Key Laboratory of Drinking Water Science and Technology, Chinese Academy of Sciences (15Z007KLDWST). Moreover, the author Guibing Zhu gratefully acknowledges the support of Humboldt Research Fellowship (1152633).
Author information
Authors and Affiliations
Contributions
L.Z., S.W., Y.Z. and C.X. performed the experiments. L.Z., S.W. and G.Z. analyzed and interpreted the data. L.Z. and G.Z. drafted the manuscript. All authors reviewed the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Zhou, L., Wang, S., Zou, Y. et al. Species, Abundance and Function of Ammonia-oxidizing Archaea in Inland Waters across China. Sci Rep 5, 15969 (2015). https://doi.org/10.1038/srep15969
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep15969
- Springer Nature Limited
This article is cited by
-
The effect of calcium addition on the nitrogen removal performance of an activated sludge system from a microbiological perspective
International Journal of Environmental Science and Technology (2024)
-
Nitrification mainly driven by ammonia-oxidizing bacteria and nitrite-oxidizing bacteria in an anammox-inoculated wastewater treatment system
AMB Express (2021)
-
Monthly distribution of ammonia-oxidizing microbes in a tropical bay
Journal of Microbiology (2021)
-
Resuscitation of anammox bacteria after >10,000 years of dormancy
The ISME Journal (2019)
-
Ammonia-oxidizing bacterial and archaeal communities in tropical bioaugmented zero water exchange shrimp production systems
Journal of Soils and Sediments (2019)