
Abstract
In the field of forensic entomology, Chrysomya rufifacies (Macquart 1843) (Diptera: Calliphoridae) has proven to be an important species for postmortem interval (PMI) assessment in criminal investigations. The developmental patterns of C. rufifacies exhibit temperature-dependent variations that are geographically specific, and these variations are influenced by seasonal changes. Hence, understanding the geographically specific development patterns of C. rufifacies in response to temperature and seasonal changes is crucial for improving the accuracy of PMI predictions. In the present study, we examined the developmental rates of C. rufifacies across a temperature gradient (20 °C, 25 °C, 30 °C, 35 °C, and 40 °C). Primary objective of present study was to gather precise developmental data essential for calculating the post-mortem interval (PMI) within varying temperature ranges. Furthermore, we investigated the influence of seasonal variations, encompassing the summer, rainy and winter seasons, on the growth and developmental patterns of C. rufifacies. The results of present studies show that temperature significantly affects the growth and development of C. rufifacies. The length of developmental stages and morphological parameters varied seasonally, with longer duration and colder temperatures in winter and shorter duration and warmer temperatures in summer. The finding that faster development was directly correlated with higher temperatures highlighted the significance of temperature in affecting an insect’s life cycle. Our research highlights the necessity of gathering information on evolution trends that are specific to a given area in order to accurately calculate PMI in forensic entomological investigations. Understanding the interactions between temperature, seasonal variation, and morphological traits is critical for establishing baseline data for PMI estimates. The accuracy of PMI estimates will ultimately increase thanks to this information, and forensic entomologists will have even more opportunities to contribute to forensic investigations.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Forensic entomologists use the invaluable potential of insects as evidence to establish exact timelines, determine the identity of suspects, confirm or disprove alibis and, most importantly, determine the postmortem interval (time since death). By closely examining the activities and life cycles of insects, these experts play a critical role in unravelling criminal mysteries and assisting law enforcement in solving crimes [1]. Forensic entomologists mostly rely on laboratory studies to calculate the postmortem interval (PMI) by analyzing the developmental data of arthropods. The colonization and developmental patterns of insects are affected by the surrounding temperature conditions. Insects are also dependent on accumulated heat energy (degree days) to move from one life stage to the next. They can only develop at certain threshold temperatures, as their growth is restricted outside these critical limits [2].
C. rufifacies have a wide distribution and is of particular interest as a blowfly species [3,4,5,6] for its particular importance to forensic science in many countries around the world. C. rufifacies plays a crucial role as an indicator species in forensic entomology and serves as a valuable tool for estimating the minimum postmortem interval (PMImin) in crime scene investigations. However, it is important to remember that development rates can vary greatly depending on local conditions. To ensure accurate estimates, it is therefore essential to base PMImin calculations on regionally generated development data.
By using specific developmental data from the local environment, forensic entomologists can improve the precision and reliability of their time-of-death determinations, contributing to more effective and scientifically sound crime scene investigations. Climate changes have a significant impact on decomposition rates, insect development, and succession patterns in different ecosystems at different times of the year and in different geographical locations [7]. These relationships underscore the critical role that temperature variation plays in shaping the intricate relationships between insects and their environment, making it an essential factor for forensic entomologists to consider when analyzing insect tracks at crime scenes.
The minimum postmortem interval (PMI), which is determined in forensic investigations, is based on the growth rate of the larvae, which is directly influenced by temperature [8]. Extreme temperatures can hinder growth by setting a lower developmental threshold beyond which any further progress comes to a halt [9, 10]. Temperature fluctuations significantly influence the rate of decomposition by slowing it down in winter and prolonging it in summer [11, 12].
The present study investigates how temperature fluctuations in India affect the life cycle of the forensic fly C. rufifacies. By analyzing its growth in response to seasonal and constant temperatures, the research aims to enhance forensic investigations and understand climate change’s impact on crucial insect species.
Material and method
Beginning of the experiment and maintenance of colony
Liver samples were purchased from a local butcher shop and allowed to putrefy, attracting flies that laid eggs on the decaying liver. Yes, bait was placed directly on the ground. Container was not used to protect the bait; instead, guards were implemented to prevent access by larger animals. Additionally, some baits were used for field collection. Adult flies were collected using an insect-gathering net and transferred to the laboratory at the Department of Zoology, Government College of Arts and Science, affiliated with Dr. Babasaheb Ambedkar Marathwada University, Aurangabad, Maharashtra, India. The adult animals were housed in cages, under non-controlled temperature and humidity. The flies were fed daily diets of raw beef liver and diluted honey in water in the lab. The fresh liver served as an ideal place for the female flies to lay their eggs. Up until the post-feeding stage, fresh beef liver was fed to the maggot cultures [13,14,15].
Maintenance of the pure culture
The egg laying of the females was continuously monitored, and as soon as the eggs were laid, the females were separated to produce a pure culture of C. rufifacies. Using previously published taxonomic keys [16, 5, 17,18,19,20,21], flies were recognized visually under a stereoscopic microscope (Magnus MS-224) to morphologically analyze these adults and maggots. To validate the morphological identification, molecular taxonomy was carried out. Adult flies were killed, and their thorax was used to perform DNA barcoding using mtCOI (Genbank accession number (MG816778)) [14]. Prepupae were housed in 500-ml beakers filled with the dry soil necessary for pupation. In a separate rearing box with dimensions of 22 inches in length, 12 inches in width, and 10 inches in depth, adult flies that emerged from pupae were raised, and normal culture was maintained [22]. By extracting the eggs or larvae of one female and cultivating them for identification and additional experimentation after the collection of the samples, pure cultures for each species were obtained [15, 22]. Oviposition observations were recorded hourly throughout the designated interval.
Evaluation of the effects of temperature and humidity
To examine the impact of different seasons on the life cycle of C. rufifacies, three replicates were conducted, and their averages were analyzed. For the seasonal study (the summer, rainy, and winter seasons, and each season lasts for six months), all cages were maintained in the laboratory under ambient room conditions, temperature and humidity were monitored using a thermo-hygrometer.
Five BOD incubators (by Narmada Instruments Corporation India, product code: ITB-9CF7F06A) with constant temperatures between 20 °C, 25 °C, 30 °C, 35 °C, and 40 °C were used for the constant temperature-dependent investigation. About 2 kg of beef liver samples were purchased from a nearby abattoir before the study was started. Each cage received 100 g of beef liver as its substrate. For the experiment, 150 larvae were collected, with 30 larvae allocated to each incubator, maintaining a constant temperature. This setup aims to investigate the impact of temperature on the growth and development of the larvae. Hourly development and morphological parameters were meticulously documented during the entire experiment, with three replicates recorded for each parameter. Feeding plates and honey water were changed every day, and cleaned plates were used every time. All the cages were cleaned at 5-day intervals. A pure culture of C. rufifacies colonies was consistently maintained throughout the year to conduct all the experiments. Fly samples from the pure culture were obtained for these trials to conduct research during the appropriate season.
Data collection
Upon placing jars containing fly eggs into an incubator, continuous day-to-day observations were performed. These observations spanned from the moment of hatching until the larvae reached the third instar, with hourly assessments conducted throughout the daylight hours. Once the eggs were hatched, two larvae from each jar were selected from the rearing jars, euthanized using hot water, and preserved in 70% alcohol for subsequent analysis of length, cross-width, and instar stage. Sampling occurred every 12 h after the larvae reached the third instar until the adult flies emerged. The length of each preserved larva was determined using a digital microscope (LB-236 Binocular Biological Digital Microscope), and the cross-width of the posterior segment was measured using a vernier calliper (Baker DC 20, with a precision reading of 0.02).
Statistical analysis
Statistical analyses were performed in R v. 4.2.2 [23]. Basic data manipulations were performed using tidyverse v. 1.3.2 [24], and data was checked for outliers and normality using rstatix v. 0.7.2 [25]. The distributions of most morphological measurements deviated from normality and were thus log-transformed so that a two-way repeated measures ANOVA (P = 0.05) could be performed on transformed and untransformed data, again using rstatix, to determine the effect of temperature on development time and larval morphometric measurements. The Mauchly test of the sphericity of data was performed, and the Greenhouse-Geisser correction was used when necessary. Figures were plotted using the ggplot2 version. 3.4.1 package [26].
Results
In the present study, it was found that the duration of the life cycle stages of C. rufifacies differed in size in the different seasons. In summer, these stages were comparatively smaller than in the rainy season, but larger than their counterparts in the winter season (Table 1). The duration of this life cycle varied depending on the season and temperature. In summer, when the average maximum temperature was 31.6 °C and the average humidity was 26%, the life cycle was completed in about 218 h, which corresponds to 9 days and 2 h. In the rainy season, with an average temperature of 29.2 °C and an average humidity of 56%, the life cycle lasted 277 h, corresponding to 11 days and 13 h (Table 1). In contrast, in winter, with an average temperature of 24.1 °C and a humidity of 39%, the life cycle lasted approximately 319 h (13 days and 5 h) Figs. 1 and 2. Figure 3 shows the development time of C. rufifacies and PMI in hours in different seasons.

Morphological parameters of Chrysomya rufifacies in various seasons

Record of average temperature and humidity in various seasons

Developmental duration of Chrysomya rufifacies in various seasons
Notably, the various developmental phases of C. rufifacies exhibited differences in size across the seasons. In the summer, these stages were comparatively smaller than those in the rainy season but larger than their counterparts during the winter season. The warmer environmental conditions influence behaviour and physiology in the summer season. Flies become more active and move faster in warm weather, which provides ideal conditions for their life cycle, including faster development. Flies tend to be more prevalent during the summer due to increased breeding opportunities. Warmer temperatures lead to higher metabolic rates in flies. This means they require more energy to sustain their activities, such as flying, foraging, and mating. That’s why flies experience accelerated growth and development in the summer, as higher temperatures promote faster maturation from egg to adult, due to which flies may weigh less. In colder temperatures, flies tend to be less active and typically have lower metabolic rates in cold weather, such as winter or rainy season. Their metabolism slows down as temperatures drop, which is a common strategy for conserving energy in adverse environmental conditions.
Flies have lower energy requirements during the winter because they are less active and have fewer opportunities for reproduction. A slower metabolism helps them conserve energy and survive with limited resources. A decreased metabolic rate can extend the lifespan of flies during the winter. That’s why morphometric parameters may vary in the summer, winter, and rainy seasons. Knowledge of flies’ life cycles helps to calculate post-mortem intervals in respective seasons. The ANOVA results reveal that temperature significantly affects the developmental stage (p < 0.05), and morphological parameters (p < 0.05).
Table 1 shows the morphological parameters and development time of C. rufifacies in various seasons. Table 2 shows the morphological parameters and development time of C. rufifacies in various constant temperatures (20 °C, 25 °C, 30 °C, 35 °C, and 40 °C). Figure 1 presents morphological parameters of C. rufifacies across different seasons. Figure 2 illustrates the recorded average temperature and humidity variations across these seasons. Figure 3 displays the developmental duration of C. rufifacies under varying seasonal conditions. In contrast, Figs. 4, 5, 6 and 7 respectively, depict the mean length, mean weight, mean width, and mean developmental time of C. rufifacies under different constant temperature settings.

Mean length of Chrysomya rufifacies in various constant temperature

Mean weight of Chrysomya rufifacies in various constant temperature

Mean width of Chrysomya rufifacies in various constant temperature

Mean developmental time of Chrysomya rufifacies in various constant temperatures
The life cycle of C. rufifacies was investigated at various constant temperatures. The life cycle was completed in 406.14 h at the lowest temperature of 20 °C. The life cycle lasted for 395.85 h when the temperature reached 25 °C (Table 2). It was shown that the larvae gained more body weight, width, and length at lower temperatures. The life cycle then dropped at further temperature increases of 30 °C, 35 °C, and 40 °C to 373.30 h, 359.42 h, and 341.54 h, respectively. Table 2 shows the developmental period of C. rufifacies in all respective constant temperatures. The development of the flies was speeded up with the increase in temperature. The flies achieved their maximum weights, widths, and lengths at the lowest temperature, and they decreased as the temperature increased. The lowest weights, widths, and lengths were observed when larvae were reared at 40 °C (Table 2). The growth of C. rufifacies was completed faster at a temperature of 35 °C and lasted only 12 days, with a slight decrease in maggot weight. The modest mortality of C. rufifacies at 40 °C showed that the flies could not withstand the high temperatures. Remarkably, a consistent relationship between higher temperatures and reduced width was observed across different developmental stages. The comprehensive data presented in Table 2 provide detailed insights into the weight (in mg), length (in mm) and width (in mm) of the developmental stages of C. rufifacies under different constant temperature conditions. The results show a recognizable trend characterized by decreasing weights at increasing temperatures in the different developmental stages. This indicates a plausible correlation between temperature fluctuations and the progression of C. rufifacies development.
Discussion
The influence of temperature on the growth and development of insects is considerable. The growth and performance of insects are greatly affected by sudden increases or decreases in temperature. In forensic investigations, it is important to consider how temperature affects the nutrition and development of maggots. Temperature and larval stages are the two main elements considered when estimating the timing of insect development in a forensic context [27]. According to Tomberlin et al. [28], critical temperatures can alter timings of forensic importance. Blowfly activity is affected by low temperatures [29] as well as heavy rain and excessive humidity [30], which can delay their arrival at a corpse. Greenberg [31] claimed that Calliphoridae cannot fly when it rains. Digby [32] also found that the ability of Calliphora vicina to fly is impaired by strong winds. Sarcophagid flies, on the other hand, are not affected by rain and may be the first insects to colonize a corpse during prolonged wet weather [33]. The activity of the blowfly is limited at temperatures above 30 °C and below 12 °C [34]. The study of insect growth rates and colonization and succession patterns at a site is necessary to estimate the postmortem interval (PMI). Temperature has a significant influence on insect activity and can change depending on the season and location [35]. Therefore, to provide a basis for forensic entomology, it is crucial to conduct carrion decomposition studies in different geographical areas and over several seasons in a given area. Similarly, precise data collection is crucial since it provides the basis for trustworthy and legitimate investigative results. To accurately estimate the postmortem interval (PMI), precise documentation of insect growth, ambient conditions, and related data is necessary. The results of criminal investigations may be impacted by minor mistakes or omissions in data collecting that cause large disparities in PMI computations.
In the present study, C. rufifacies was examined for seasonal variation, and its life cycle was recorded for the summer, rainy, and winter seasons. The results of our study shed light on the seasonal fluctuations in the life cycle of the blowfly species C. rufifacies, which is important for forensic entomology. One of the main objectives of the present study was to determine how seasonal variations, especially of temperature and humidity, influence the maturation schedule of this species. It became clear that these environmental conditions had a significant impact on how long C. rufifacies lived. In summer, with an average temperature of 31.6 °C and humidity of 31.6%, the life cycle is completed in an exceptionally short 218 h. Higher temperatures are known to speed up an insect’s metabolism and development. The accelerated development seen in our study is probably due to the higher metabolic activity in warmer environments. On the other hand, the life cycle of C. rufifacies lasted 317 h during the winter, which was characterised by an average temperature of 24.1 °C and 39% humidity. The slower development that occurs at lower temperatures can be attributed to the decrease in metabolic rate. For this reason, the developmental phases of the species were significantly prolonged during the winter. The life cycle lasted 277 h during the rainy season, when temperatures and humidity were in the medium range (average temperature of 29.2 °C and 67% humidity). These results indicate that although temperature remains the most important factor, the slowdown in development caused by lower temperatures can be somewhat offset by higher humidity. The relationship between humidity and temperature, which is most pronounced during the rainy season, favours an environment in which C. rufifacies can develop on a more balanced schedule.
Seasonal fluctuations also influence the morphological characteristics of the different developmental stages. The larvae gained more weight in winter than in the rainy season and in the summer. The highest weight was recorded in summer, when the temperature was high. The length and width of the larvae were also shorter in the summer season. However, the life cycle was longer in the winter than in the rainy and summer months. A similar study was conducted by Abd Al Galil and Zambare [36] who showed the effect of temperature on the life cycle of C. rufifacies which was influenced by temperature. Their results revealed that C. rufifacies took 11.46 ± 0.45 days during the rainy season, 10.04 ± 0.12 days during the summer, and 13.25 ± 0.25 days during the winter to complete its development. High temperatures both speeded up the development in the summer and slowed it down by roughly three days in the winter. In the dry season, the larvae are larger, in summer, they are smaller and in winter, they are even smaller. The length of the life cycle stages is significantly influenced by temperature. The time required to complete the life cycle increases at lower temperatures, while it decreases at higher temperatures [9, 37]. Higher temperatures usually accelerate the hatching of the eggs and the maturation of the Calliphoridae larvae, which can increase considerably in size within a few hours. The length of the larvae usually reaches its peak during the feeding phase and then gradually decreases again. The period before feeding and pupation usually takes up about 75% of the preadult cycle of Calliphoridae flies [37]. Even more than meteorological factors, temperatures, larval growth, and abundance determine how long the bloated stage, which is characterized by the accumulation of gases inside the body, lasts. When the insect larvae penetrate the host and release gas, the bloated stage is over [38]. According to Tantawi et al. [39], the bloated stage lasts longer in the summer than it does in the spring. The principal fly species that colonizes rabbit carcasses throughout these seasons has differences in its biology of reproduction, which is thought to be the cause of this variance.
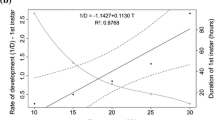
During the temperature-dependent study, the life cycle of the fly was completed in 406.14 h (20 °C), 395.85 h (25 °C), 373.30 h (30 °C), 359.42 h (35 °C) and 341.54 h (40 °C), respectively. At 20 and 25 °C, the fly took an extremely long time to complete its development. Due to the lowest temperature, development of the fly was delayed, but when the temperature was raised to 30 °C, 35 °C, and 40 °C, development was completed in a short time. Flores [40]. also mentioned that temperatures above 9 °C are necessary for C. rufifacies eggs to hatch, and larvae will only pupate at temperatures above 15 °C. For calculating the time of death based on insect developmental phases, precise temperature data is essential. Similar results were obtained by Rogers et al. [41], as per their study, 24 °C required more accumulated degree hours (ADH) than 30 °C for C. rufifacies growth. Interestingly, the species’ diet was more significantly impacted by the lower temperature of 24 °C, with notable differences in ADH requirements and survival rates. Compared to larvae raised on pork loin, those raised on pork mince showed higher development. Beef and pork liver growth was hindered at 24 °C, and specimens failed to reach the third instar. Among all rearing mediums, C. rufifacies did, however, successfully develop at 30 °C, which correlated with the finding of this study, which shows C. rufifacies grew well at 35 °C. At high temperatures (40 °C), the life cycle of the fly was disrupted, and 25% mortality was observed. Due to the high temperature, the emergence of the fly was affected, and flies died soon after the emergence. The length, weight, and width of the larvae were also minimal as compared to other constant temperature ranges under the study. This study supports the results obtained by Yanmanee et al. [7], who investigated the developmental rates of C. rufifacies in its native range in Thailand under nine constant temperature regimes: 15, 18, 21, 24, 27, 30, 33, 36, and 39 °C. The study revealed varying developmental times from egg to adult across these temperatures, with the longest duration observed at 15 °C (618 h) and the shortest at 33 °C (168 h). It is noteworthy that no pupae hatched at 39 °C, indicating the detrimental effect of extreme heat on the development of the insect. These results are of great importance for forensic entomology as they allow more accurate estimates of the post-mortem interval in the region and provide valuable insights into the influence of temperature on insect growth. Further research is needed to explore the underlying mechanisms and extend the study to other geographic regions.
The development of flies is heavily influenced by temperature. Higher temperatures often hasten development, resulting in shorter life cycles, faster growth, and faster passage through several life stages [16]. Higher temperatures boost metabolic activity, while lower temperatures slow down fly development, extending life cycles. Lower temperatures align fly life cycles with favorable conditions, but extremes can be harmful. Each fly species has an optimal temperature range for efficient development. Understanding temperature effects is crucial in forensic calculations, especially for determining the post-mortem interval (PMI) [37]. Hence, scientists across the world are studying flies development with respect to time. Gabre et al. [42] examined the C. megacephala fly’s growth rate in Egypt at a temperature of 26 °C and a humidity level of 60–70%. Claver and Yaqub [43] raised the C. megacephala fly species on chicken meat in India under three distinct temperature and humidity conditions: 19–24 °C and 85–96% humidity, 24–28 °C and 82–94% humidity, and 28–32 °C and 78–84% humidity. This allowed them to study the development of this fly species. Bambaradeniya et al. [44] also studied C. megacephala at various temperature regimes in Sri Lanka. In India, Abd-Algalil and Zambare [36], Bansode et al. [13, 22, 27, 37, 45] have studied different types of Calliphoridae species to study the effect of temperature on flies growth and development. Such baseline region- and country-specific developmental data are crucial when applying insect shreds of evidence in criminal investigations to calculate PMI.
In the context of current global research initiatives focused on understanding the evolution of flies in relation to temperature, the importance of this study lies in the provision of regionally specific evolutionary data. However, to comprehensively address existing knowledge gaps, future research efforts should prioritize investigating additional variables that influence fly development. Expanding the study to a broader range of environmental conditions is of utmost importance. This ongoing research is essential for the continued advancement of forensic entomology and the creation of a solid foundation for the reliable use of insect evidence in criminal investigations.
Conclusions
The present study concludes by highlighting the crucial influence of temperature on the growth of C. rufifacies, an important aspect to be taken into account in forensic entomology. Gaining accurate postmortem evaluations necessitates a sophisticated comprehension of local climatic changes. C. rufifacies’s versatility emphasizes how important it is to understand temperature-dependent patterns in order to use them effectively in forensic applications. To have a better understanding, it is suggested that future studies explore the physiological and genetic mechanisms behind the biology and behavior of blowflies.
Data availability
The data and the results used to support the findings of this study are presented in the manuscript.
References
Keh B (1985) Scope and applications of forensic entomology. Annu Rev Entomol 30:137–154. https://doi.org/10.1146/annurev.en.30.010185.001033. PMID: 3882048
Greenberg B, Kunich JC (2002) Entomology and the law: flies as forensic indicators, 248 edn. Cambridge Univ. Press, Cambridge, pp 102–103
Sukontason KL, Sukontason K, Narongchai P, Lertthamnongtham S, Piangjai S, Olson JK (2001) Chrysomya rufifacies (Macquart) as a forensically-important fly species in 272 Thailand: a case report. J Vector Ecol J Soc Vector Ecol 26:162–164
Sukontason KL, Sukontason K, Lertthamnongtham S, Kuntalue B, Thijuk N, Vogtsberger RC et al (2003) Surface ultrastructure of Chrysomya rufifacies (Macquart) larvae 278 (Diptera: Calliphoridae). J Med Entomol 40:259–267
Sukontason K, Piangjai S, Siriwattanarungsee S, Sukontason KL (2008) Morphology and 274 developmental rate of blowflies Chrysomya megacephala and Chrysomya rufifacies in 275 Thailand: application in forensic entomology. Parasitol Res 102:1207–1216
Wells JD, Sperling FA (1999) Molecular phylogeny of Chrysomya albiceps and C. rufifacies (Diptera: Calliphoridae). J Med Entomol 36(3):222–226
Yanmanee S, Husemann M, Benbow ME, Suwannapong G (2016) Larval development rates of Chrysomya rufifacies Macquart, 1842 (Diptera: Calliphoridae) within its native range in South-East Asia. Forensic Sci Int 266:63–67
Johnson AP, Wighton SJ, Wallman JF (2014) Tracking movement and temperature selection of larvae of two forensically important blow fly species within a maggot mass. J Forensic Sci 59(6):1586–1591
Campobasso CP, Vella GD, Introna F (2001) Factors affecting decomposition and Diptera colonization. Forensic Sci Int 120:18–27
Gennard D (2012) Forensic entomology: an introduction. Wiley, Hoboken, NJ
Tarone AM, Foran DR (2006) Components of developmental plasticity in a Michigan population of Lucilia Sericata (Diptera: Calliphoridae). J Med Entomol 43(5):1023–1033
Boatright SA, Tomberlin JK (2010) Effects of temperature and tissue type on the development of Cochliomyia macellaria (Diptera: Calliphoridae). J Med Entomol 47(5):917–923
Bansode SA, More VR, Zambare SP (2017) Effect of seasonal variations on the life cycle of Lucilia Cuprina (Wiedemann, 1830)(Diptera: Calliphoridae). J Entomol Zool stud 5:1518–1522
Sarika B, Vinod R (2023) Molecular toxonomy of forensically important carrion flies using mt (DNA COI). Indian J Entomol 643–646
Annasaheb Bansode S, Ramrao More V (2023) Effect of lorazepam on the development of the hairy maggot blow fly, Chrysomya rufifacies (Macquart): implication for forensic entomology. J Toxicol 2023
Nandi BC (1994) Studies on calliphorid flies (Diptera: Calliphoridae) from Calcutta and adjoining areas. J Bengal Nat Hist Soc 13(2):37–47
James MT (1971) Genus Chrysomya in New Guinea. Pac Insects 13(2):361–369
Kamal RMZR, Mohamed AM, John J, Ahmad FMS, Wan OA, Baharudin O (2008) Descriptions of the larval instars of Chrysomya rufifacies (Macquart) (Diptera: Calliphoridae), a species of forensic importance in Malaysia. Jurnal Sains Kesihatan Malaysia 2:35–41
Sukontason KL, Narongchai P, Kanchai C, Vichairat K, Piangjai S, Boonsriwong W, Bunchu N, Sripakdee D, Chaiwong T, Kuntalue B, Siriwattanarungsee S, Sukontason K (2006) Morphological comparison between Chrysomya rufifacies (Macquart) and Chrysomya Villeneuvi Patton (Diptera: Calliphoridae) puparia, forensically important blow flies. Forensic Sci Int 164:230–234
Sukontason KL, Ngern-Klun R, Sripakdee D, Sukontason K (2007) Identifying fly puparia by clearing technique: application to forensic entomology. Parasitol Res 101:1407–1416
Sukontason K, Methanitikorn R, Sukontason KL, Piangjai S, Olson JK (2004) clearing technique to examine the cephalopharygeal skeletons of blow fly larvae. J Vector Ecol 29: 192–195
Bansode SA, More VR, Zambare SP, Fahd M (2016) Effect of constant temperature (20 0 C, 25 0 C, 30 0 C, 35 0 C, 40 0 C) on the development of the Calliphorid fly of forensic importance, Chrysomya Megacephala (Fabricus, 1794). J Entomol Zool Stud 4(3):193–197
Team RC (2020) RA language and environment for statistical computing, R Foundation for Statistical. Computing
Wickham H, Averick M, Bryan J, Chang W, McGowan LD, François R, Grolemund G, Hayes A, Henry L, Hester J, Kuhn M, Pedersen TL, Miller E, Bache SM, Müller K, Ooms J, Robinson D, Seidel DP, Spinu V, Takahashi K, Vaughan D, Wilke C, Woo K, Yutani H (2019) Welcome to the tidyverse. J Open Source Softw 4(43):1686. https://doi.org/10.21105/joss.01686
Kassambara A (2023) rstatix: Pipe-Friendly Framework for Basic Statistical Tests. R package version 0.7.2. https://rpkgs.datanovia.com/rstatix/
Wickham H (2016) ggplot2: Elegant graphics for data analysis. Springer-Verlag New York. ISBN 978-3-319-24277-4. https://ggplot2.tidyverse.org
Bansode SA, More VR, Zambare SP (2016) Effect of different constant temperatures on the life cycle of a fly of forensic importance Lucilia Cuprina. Entomol Ornithol Herpetol 5(3):2161–0983
Tomberlin JK, Mohr R, Benbow ME, Tarone AM, Vanlaerhoven S (2011) A roadmap for bridging basic and applied research in forensic entomology. Ann Rev Entomol 56(1):401–421
Shah ZA, Sakhawat TA, Eela NZ (2004) The effect of flesh age, trap colour, decomposition stage, temperature and relative humidity on the visitation pattern of blow and flesh flies. Int J Agric Biology 6(2):370–374
Smith KG (1986) A manual of forensic entomology
Greenberg B (1990) Nocturnal oviposition behaviour of blowflies (Diptera: Calliphoridae). J Med Entomol 27:807–810
Digby PSB (1958) Flight activity in the blowfly Calliphora Erythrocephala, in relation to wind speed, with special reference to adaptation. J Exp Biol 35:776–795
Erzinçlioglu Z (2000) Maggots, murder and men: Memories and reflections of a forensic entomologist. Harley Books, Colchester
Gennard D (2007) Forensic entomology: an introduction. Wiley, Chichester
Kenneth G, Hall RD. Testing reliability of animal models in research and training programs in forensic entomology, Part 11
Abd Al Galil FM, Zambare S (2015) Effect of temperature on the development of a calliphorid fly of forensic importance, Chrysomya rufifacies (Macquart, 1842). Int J 3:1099–1103
Bansode SA, More VR, Zambare SP (2016) Effect of constant temperature (20 ºC, 25 ºC, 30 ºC, 35 ºC, 40 ºC) on the development of Sarcophagidae: Diptera (FAB)(Sarcophagidae: Diptera). J Pet Environ Biotechnol 7(275):2
Anderson GS, Vanlaerhoven SL (1996) The initial studies on insect succession on carrion in South Western British Columbia. Forensic Sci 41:617–625
Tantawi TI, El-Kady EM, Greenberg B, El-Ghaffar HA (1996) Arthropod succession on exposed rabbit carrion in Alexandria, Egypt. J Med Entomol 33(4):566–580
Flores M (2013) Life-history traits of Chrysomya rufifacies (Macquart)(Diptera: Calliphoridae) and its associated non-consumptive effects on Cochliomyia macellaria (Fabricius)(Diptera: Calliphoridae) behavior and development (Doctoral dissertation)
Rogers EK, Franklin D, Voss SC (2021) Dietary effects on the development of Calliphora dubia and Chrysomya rufifacies (Diptera: Calliphoridae): implications for postmortem interval. J Med Entomol 58(1):79–87
Gabre RM, Adham FK, Chi H (2005) Life table of Chrysomya megacephala (Fabricius)(Diptera: Calliphoridae). Acta Oecol 27(3):179–183
Claver MA, Yaqub A (2015) Studies on the biology of the blowfly Chrysomya megacephala (Fabricius)(Diptera: Calliphoridae) under fluctuating temperatures in three different seasons. Int J Res Studies Biosci 3:107–112
Bambaradeniya YTB, Karunaratne WIP, Tomberlin JK, Goonerathne I, Kotakadeniya RB, Magni PA (2019) Effect of temperature and tissue type on the development of the forensic fly Chrysomya megacephala (Diptera: Calliphoridae). J Med Entomol 56(6):1571–1581
Ames C, Turner B (2003) Low-temperature episodes in the development of blowflies: implications for postmortem interval estimation. Med Vet Entomol 17(2):178–186
Acknowledgements
The authors are thankful to the Principal and H. O. D. Dept. of Zoology, Govt. College of Arts and Science, MH. India, for providing all the required facilities to carry out the experiments. The authors are thankful to Dr. Tyler A. Elliott, Research Associate, Adamowicz Lab, Department of Integrative Biology, University of Guelph, Canada, for helping with data analysis and proofreading.
Funding
There is no funding for this work.
Author information
Authors and Affiliations
Contributions
Both authors contributed equally.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they don’t have any conflicts of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Highlights
• Chrysomya rufifacies is pivotal for precise postmortem interval (PMI) determination in forensic entomology.
• Temperature significantly influences growth rates of Chrysomya rufifacies, with higher temperatures leading to faster development.
• Seasonal variations impact developmental phases, highlighting the need for region-specific data in PMI calculations.
• Understanding temperature and morphological features is crucial for establishing accurate baseline PMI estimates.
• This research enhances PMI accuracy, empowering forensic entomologists in criminal investigations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Bansode, S., More, V. Effects of temperature variation on the life cycle of the forensically important Calliphoridae fly Chrysomya rufifacies (Macquart 1843) (Diptera). Int J Trop Insect Sci 44, 747–757 (2024). https://doi.org/10.1007/s42690-024-01189-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42690-024-01189-6