
Abstract
Purpose
The blowfly Sarconesia chlorogaster (Diptera: Calliphoridae) is of limited forensic use in South America, due to the poorly known relationship between development time and temperature. The purpose of this study was to determine development time of S. chlorogaster at different constant temperatures, thereby enabling the forensic use of this fly.
Methods
Development time of this species was examined by observing larval development at six temperatures (10, 15, 20, 25, 30, 35 °C). The thermal constant (K), the minimum development threshold (t 0), and development rate were calculated using linear regressions of the development time interval at five temperatures (10–30 °C).
Results
Development interval from egg to adult varied from 14.2 to 95.2 days, depending on temperature. The t0 calculated for total immature development is 6.33 °C and the overall thermal constant is 355.51 degree-days (DD). Temperature affected the viability of pupae, at 35 °C 100 % mortality was observed.
Conclusion
Understanding development rate across these temperatures now makes development of S. chlorogaster a forensically useful tool for estimating postmortem interval.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Forensic entomology helps determine postmortem interval by providing tools that estimate the time elapsed between the time at which an insect lays eggs on a body and its discovery (minimum postmortem interval, PMImin) [1]. Among the most useful methods for estimating PMImin is aging the insect larvae collected from the remains. To this end, fly larvae are commonly used because the stages of larval development and the time it takes for them to go through those stages are known for several species over a range of temperatures (using a species-specific model of accumulated degree-hours or degree-days [2]).
Flies in the family Calliphoridae (Diptera) are the most frequently used group of insects for PMI estimation [3], using models of accumulated degree-hours. Sarconesia chlorogaster (Wiedemann, 1830), along with other Calliphoridae, has become a key species in forensics in South America because larvae are often found feeding on carcasses [4–8] and on human remains [9]. This fly is endemic to South America, in Argentina, Uruguay, Bolivia, Peru, Chile, and southern Brazil [10–12].
One of the key components of using insects to estimate PMI is that their developmental rate is temperature dependent [13]. Thus, precise estimates of PMI will require knowledge of the developmental rates of the fly species over the range of temperatures encountered in the field. The life cycle of S. chlorogaster has been described at specific controlled [14, 15] and fluctuating temperatures [16–18]. While providing information showing that development rate is temperature dependent, previous studies did not examine the range of temperatures often encountered in field conditions and so the forensic potential of this species is limited.
Because S. chlorogaster is found on bodies and thus offers great potential for estimating PMI [9], and considering the limited information about its biology and development rates, we aimed to develop a model of accumulated degree-hours based on the growth rate of S. chlorogaster at constant temperatures. We also examined the effect of these different temperatures on immature stage development rate and estimated the minimum development threshold (t0) and thermal constant (K). With this information, the forensic use of, and general understanding of the biology of this species, will be greatly improved.
Materials and methods
Development of immature stages at controlled temperatures
To establish colonies in the laboratory, S. chlorogaster were collected in Curitiba (25°25′S, 49°14′W), in the state of Paraná, Brazil, using a trap baited with rotting sardines. This colony was maintained under a natural photoperiod with varying room temperatures. Adults were fed with sugar, milk powder, raw ground beef, and water ad libitum. Eggs for this study were obtained from raw ground beef that was placed in the colony cages for 3 h to allow egg deposition. Eggs were then transferred to containers and placed in incubators (122FC; Eletrolab®, São Paulo, Brazil).
Each incubator was adjusted to maintain constant temperature (either 10, 15, 20, 25, 30, or 35 ± 1 °C), humidity (60 ± 10 %), and photoperiod (12:12 h). Temperatures and humidity were constantly monitored with thermohygrometers (TH-439; Equitherm®, Rio Grande do Sul, Brazil). Five subsamples, each with ~150 eggs, were used at each temperature. The eggs of each subsample were placed in a 500 ml plastic container with 180 g of artificial diet [19]. This container was placed within a 1000 ml plastic container with vermiculite as substrate for pupation. Experiments were conducted using three generations (F3, F4, and F5) from the stock colony.
The semi-synthetic diet [19] used to feed the larvae included bovine stomach tissue, along with milk powder, brewer’s yeast, casein, nipagin, and agar. We chose this diet because it can be used without major deleterious effects for necrophagous larvae and can substitute for a standard beef diet [20, M.C. Lecheta unpublished results], so did not impair its use in forensic research.
Treatments were checked hourly to determine how long the eggs took to hatch. Because larvae were known to develop at different rates at different temperatures, larvae were observed from hatching to the third instar stage every 6 h at 20–35 °C. Larvae at 15 °C, and third instars at the remaining temperatures, were observed every 12 h. At 10 °C, larvae were observed every 24 h. At each observation, 150 larvae (30 per container) were sampled to determine the larval instar. To follow the developmental time of the pupal stage and adult eclosion, 160 larvae per temperature (32 larvae per subsample) were individually placed in tubes containing vermiculite, after they had completed feeding. These individuals were observed every 24 h to determine the day of pupation, emergence, and mortality. Thereafter, additional observations at 32 ± 1 °C only were used to determine survival of the pupal stage. The methodology was the same as the previous experiment, but only two subsamples were used.
Calculation of the ADH
Minimum development threshold (t 0), thermal constant (K), and rate of development were estimated for each developmental stage and for the total time period it took to develop from egg to adult, using the linear degree-hour model [13, 21]. Minimum development threshold was estimated by linear regression between the development rates (y = 1/time development) and constant temperature (x). The thermal constant was calculated from the equation K = 1/b, where b is the coefficient of the linear regression of the rate of development. If any temperature did not result in complete development, then those temperatures were not used in these calculations, because the degree-hours model requires complete development [13, 22], and with the subsamples pooled by temperature, to avoid pseudoreplication [23].
As subsamples are not independent samples (replicates) we can not use inferential statistics to test any hypothesis [23]. We determined the effect of temperature on S. chlorogaster development time (and its variability) based on the 95 % confidence intervals of the pooled mean at each temperature. In this framework the confidence intervals represents the envelope (range) within which the true mean may lie [24, 25]. All analyses were done on environment R [26].
Results
Development of the immature stages at different controlled temperatures
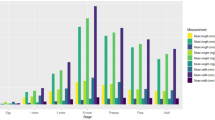
Temperature affected total development time from egg to adult as well as development time to each stage: egg (the CIs of sample means only overlapped for flies reared at 30 and 35 °C, Table 1); first instar (the CIs of sample means only overlapped for flies reared at 25 and 35 °C, Table 1); second instar (the CIs of the sample mean at 35 °C overlapped those for 25 and 30 °C, but the CIs of the latter two temperatures did not overlap, Table 1), third instar (non overlapping CIs for all sample means temperatures, Table 1) and pupae (non overlapping CIs for all sample means temperatures, Table 1).
The time interval of each stage decreased with increasing temperature except at 35 °C. The total period from egg to adult ranged from 14.2 to 95.2 days. The egg-laying to hatching interval ranged from 14 h to 2.7 days. The first instar ranged from 12 h to 3.9 days, second instar from 17.6 h to 5.4 days, and third instar from 2.3 to 16.1 days. The pupal period ranged from 10 to 66.8 days (Table 1).
Pupae reared at intermediate temperatures (15–25 °C) had greater than 96 % survival. At 10 and 30 °C survival was around 80 %, and no survival occurred at 35 °C. Adults did not emerge at 32 ± 1 °C. Thus, the biological upper limit for the complete development of S. chlorogaster is approximately 31 ± 1 °C.
Constant K and estimated minimum development threshold
There is little variation in growth rate at each temperature (as seen by the r 2 values and confidence intervals (Figs. 1, 2) and the regressions clearly show the relationship between development rate (1/D) and temperature. The parameters t 0 and K were 4.77 °C and 336.96 h °C for time to hatch, 10.11 °C and 212.4 h °C for the 1st instar, 7.40 °C and 422.88 h °C for the 2nd instar, 6.77 °C and 1288.56 h °C for 3rd instar, 5.95 °C and 6289.44 h °C for pupation. Complete development t 0 and K were 6.33 °C and 8532.24 h °C. Overall, the estimated minimum development threshold was similar for all stages of development, around 6 °C, with the exception of the first instar larvae (10.11 °C). Parameters estimated from the regressions are found in Table 2.

Development rate (continuous line, with a 95 % confidence interval, and corresponding equations) and duration (in hours, dotted line) of S. chlorogaster, estimated from controlled temperatures of 10–30 °C, for the egg development period (a), first instar (b), and second instar (c). The line of development period is to a third degree polynomial adjusted curve

Development rate (continuous line, with a 95 % confidence interval, and corresponding formulae) and duration (in hours, dotted line) of S. chlorogaster, estimated from constant temperatures of 10–30 °C for the third instar (a), pupa (b), and total period from egg laying to adult (c). The line of development period is to a third degree polynomial adjusted curve
Discussion
We found that S. chlorogaster has well defined development limits and will offer very useful information in forensic cases, especially to estimate PMImin. Within limits, development patterns in S. chlorogaster are similar to those of other Calliphoridae and muscoid dipterans, i.e., higher temperatures (within limits) result in faster development [for example, see 27–34].
In Peru, development time in S. chlorogaster under fluctuating temperatures (13.7–24.8 °C) [16] were similar to those in this study at 15 and 20 °C. With the same source population as this study, developmental period at ambient temperatures (22.2–29.3 °C) [17] resulted in a total development time similar to what we found at 20 °C [17]. In Rio Grande do Sul, Brazil, with an average temperature of 13.6 °C [18], development time was similar to our 15 °C treatment. These discrepancies in development time could result from either the different experimental designs (temperature regimes) or population variations.
In a study conducted by Bonatto using flies that originated from the same site potentially conflicting growth rates at controlled temperatures were described [14]. For example, time intervals for development of S. chlorogaster at 27 °C [14] are comparable to our treatments at 20–25 °C (first instar), 20 °C (second instar), and 15 °C (third instar). Perhaps this is due to the different diets, since Bonatto did not include animal tissue in the diet he used [14]. Because we wished to examine the forensic potential of this species, we suggest that the diet we used was more similar to that of bodies, and thus provides a reasonable estimate of growth rates. While the diet in the study by Bonatto resulted in complete development, it also may have increased development time due to a compensatory response resulting in lower weight gain per unit time. Thus, a greater development time was needed to reach the minimum weight to move to the next stage. In contrast, using the same source population, the period of egg to third instar at 25 °C (beef diet) [15] was very similar to our results at the same temperature.
Determination of the minimum threshold of development and the values of degree-hours or degree-days for each stage of development is an important prerequisite for using the model of accumulated degree-hours (ADH) for the estimates of PMImin [22]. Here we find an overall minimum development threshold of 6 °C for S. chlorogaster. It has been suggested that, in the absence of information on the biology of fly species of forensic interest, one may use the minimum development threshold of 10 °C for tropical species and 6 °C for temperate species [22]. Despite also being found in the tropics, S. chlorogaster is common in colder regions with more temperate climates [12]. So, for this species, using the tropical rather than temperate minimum development threshold will bias PMImin estimation.
The maximum threshold temperature is often ignored in the calculation of accumulated degree-hours, because high temperatures in forensic settings rarely reach a level that would cause lethal effects [35]. Upper temperature limits are rare at crime scenes. However, if temperatures approach the upper limit for an extended time, PMImin will be underestimated [36]. A good example where maximum temperature could be encountered would be in the trunk of a car exposed to full sunlight. Also, even though S. chlorogaster occurs in colder regions, high temperatures (>30 °C) can be common during the summer. Thus, because the upper threshold for S. chlorogaster was 30–31 °C (calculated from development and mortality), the maximum threshold for this species must be understood when estimating the PMImin. In S. chlorogaster, temperatures above 31 °C will increase larval development time and reduce adult eclosion.
The most important result of this study is that we provide a precise model that can be used to estimate PMImin based on age determination of immature stages of S. chlorogaster, since these data can be used to determine the period of activity (sensu Campobasso and Introna [37]). However, based on differences on development time observed between our study and others, is advisable that more studies be conducted to better understand this variation. Additionally, we recommend future experiments on the maximum and minimum thresholds of development on this and other species of forensic importance, to ensure greater reliability of ADD/ADH models used in estimating PMImin. Our results in this study may assist forensic entomologists to make more accurate estimations of PMImin when S. chlorogaster is present.
Key Points
-
1.
We show that the development of S. chlorogaster, a key species for forensic entomology in South America, occurs within well defined upper and lower limits that should be incorporated in PMI estimates.
-
2.
Sarconesia chlorogaster showed little variation in growth rate at each temperature and the time interval of each stage decreased with increasing temperature except at 35 °C. The total period from egg to adult ranged from 14.2 to 95.2 days.
-
3.
Pupae reared at intermediate temperatures had greater than 96 % survival. At 10 and 30 °C survival was around 80 %, and no survival occurred at 35 °C. Adults did not emerge at 32 ± 1 °C.
-
4.
The biological upper limit for the complete development of S. chlorogaster is approximately 31 °C and the estimated minimum development threshold is 6.33 °C. The constant K was 8532.24 h °C.
-
5.
Overall we provided accurate models that can be a useful tool to estimate PMImin based on immature stages of S. chlorogaster.
Change history
13 November 2020
A Correction to this paper has been published: https://doi.org/10.1007/s12024-020-00331-3
References
Amendt J, Campobasso CP, Gaudry E, Reiter C, LeBlanc HN, Hall MJR. Best practice in forensic entomology—standards and guidelines. Int J Legal Med. 2007;121:90–104.
Amendt J, Richards CS, Campobasso CP, Zehner R, Hall MJR. Forensic entomology: applications and limitations. Forensic Sci Med Pathol. 2011;7:379–92.
Gunn A. Essential forensic biology. Chichester: Wiley; 2006.
Moura MO, de Carvalho CJB, Monteiro-Filho ELA. A Preliminary analysis of insects of medical-legal importance in Curitiba, State of Paraná. Mem Inst Oswaldo Cruz. 1997;92:269–74.
Centeno N, Maldonado M, Oliva A. Seasonal patterns of arthropods occurring on sheltered and unsheltered pig carcasses in Buenos Aires Province (Argentina). Forensic Sci Int. 2002;126:63–70.
Horenstein MB, Arnaldos MI, Rosso B, García MD. Estudio preliminar de la comunidad sarcosaprófaga em Córdoba (Argentina): aplicación a la entomología forense. An Biol. 2005;27:191–201.
Souza ASB, Kirst FD, Krüger RF. Insects of forensic importance from Rio Grande do Sul state in southern Brazil. Rev Bras Entomol. 2008;52:641–6.
Horenstein MB, Linhares AX, Ferradas BR, García D. Decomposition and dipteran succession in pig carrion in central Argentina: ecological aspects an their importance in forensic science. Med Vet Entomol. 2010;24:16–25.
Vairo KP, Corrêa RC, Lecheta MC, Caneparo MF, Mise KM, Preti D, et al. Forensic use of a subtropical blowfly: the first case indicating minimum post-mortem interval (mPMI) in Southern Brazil and first record of Sarconesia chlorogaster from a human corpse. J Forensic Sci. 2015;60:S257–60.
de Carvalho CJB, Ribeiro PB. Chave de identificação das espécies de Calliphoridae (Diptera) do sul do Brasil. Rev Bras Parasitol Vet. 2000;9:169–73.
James MT. Family calliphoridae. In: Papavero N, editor. A catalogue of the Diptera of the Americas South of the United States. São Paulo: Museu de Zoologia da Universidade de São Paulo; 1970. P. 102.1–28.
Dear JP. A revision of the Toxotarsinae (Diptera, Calliphoridae). Pap Avulsos Zool. 1979;32:145–82.
Ikemoto T, Takai K. A new linearized formula for the law of total effective temperature and the evaluation of line-fitting methods with both variables subject to error. Environ Entomol. 2000;29:671–82.
Bonatto SR. Ciclo de vida de Sarconesia chlorogaster (Wiedemann) (Diptera, Calliphoridae, Toxotarsinae), criada sob condições de laboratório em dieta artificial. Rev Bras Zool. 1996;13:685–706.
Wells JD, Lecheta MC, Moura MO, LaMotte LR. An evaluation of sampling methods used to produce insect growth models for postmortem interval estimation. Int J Legal Med. 2015;129:405–10.
Greenberg B, Szyska ML. Immature stages and biology of fifteen species of Peruvian Calliphoridae (Diptera). Ann Entomol Soc Am. 1984;77:488–517.
Queiroz SMP, de Almeida JR, de Carvalho CJB, Dudas L. Bionomia de Sarconesia chlorogaster (Wiedemann, 1830) (Diptera, Calliphoridae) em Curitiba, Paraná, Brasil. An Soc Entomol Bras. 1985;14:105–10.
Krüger RF, Kirst FD, Souza ASB. Rate of development of forensically-important Diptera in southern Brazil. Rev Bras Entomol. 2010;54:624–9.
Estrada DA, Grella MD, Thyssen PJ, Linhares AX. Taxa de desenvolvimento de Chrysomya abiceps (Wiedemann) (Diptera: Calliphoridae) em dieta artificial acrescida de tecido animal para uso forense. Neotrop Entomol. 2009;38:203–7.
Rabêlo KCN, Thyssen PJ, Salgado RL, Araújo MSC, Vasconcelos SD. Bionomics of two forensically important blowfly species Chrysomya megacephala and Chrysomya putoria (Diptera: Calliphoridae) reared on four types of diet. Forensic Sci Int. 2011;210:257–62.
Kipyatkov VE, Lopatina EB. Intraspecific variation of thermal reaction norms for development in insects: new approaches and prospects. Entomol Rev. 2010;90:163–84.
Higley LG, Haskell NH. Insect development and forensic entomology. In: Byrd JH, Castner JL, editors. Forensic entomology: the utility of arthropods in legal investigations. 2nd ed. Boca Raton: CRC Press; 2010. p. 389–406.
Hurlbert SH. Pseudoreplication and the design of ecological field experiments. Ecol Monogr. 1984;54:187–211.
Tryon WW. Evaluating statistical difference, equivalence, and indeterminacy using inferential confidence intervals: an integrated alternative method of conducting null hypothesis statistical tests. Psychol Methods. 2001;6:371–86.
Steidl RJ, Thomas L. Power analysis and experimental design. In: Scheiner SM, Gurevitch J, editors. Design and analysis of ecological experiments. 2nd ed. New York: Oxford University Press; 2001. p. 14–36.
R Development Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. 2011. ISBN 3-900051-07-0. http://www.R-project.org.
Grassberger M, Reiter C. Effect of temperature on Lucilia sericata (Diptera: Calliphoridae) development with special reference to the isomegalen- and isomorphen-diagram. Forensic Sci Int. 2001;120:32–6.
Grassberger M, Reiter C. Effect of temperature on development of the forensically important holarctic blow fly Protophormia terraenovae (Robineau-Desvoidy) (Diptera: Calliphoridae). Forensic Sci Int. 2002;128:177–82.
Donovan SE, Hall MJR, Turner BD, Moncrieff CB. Larval growth rates of the blowfly, Calliphora vicina, over a range of temperatures. Med Vet Entomol. 2006;20:106–14.
Richards CS, Villet MH. Factors affecting accuracy and precision of thermal summation models of insect development used to estimate post-mortem intervals. Int J Legal Med. 2008;122:401–8.
Richards CS, Villet MH. Data quality in thermal summation development models for forensically important blowflies. Med Vet Entomol. 2009;23:269–76.
Boatright SA, Tomberlin JK. Effects of temperature and tissue type on the development of Cochliomyia macellaria (Diptera: Calliphoridae). J Med Entomol. 2010;47:917–23.
Gosselin M, Charabidze D, Frippiat C, Bourel B, Gosset D, Rasmont P. Development time variability: adaptation of Régnière’s method to the intrinsic variability of Belgian Lucilia sericata (Diptera, Calliphoridae) population. J Forensic Res. 2010;1:109.
Tarone AM, Picard CJ, Spiegelman C, Foran DR. Population and temperature effects on Lucilia sericata (Diptera: Calliphoridae) body size and minimum development time. J Med Entomol. 2011;48:1062–8.
Wilson LT, Barnett WW. Degreedays: an aid in crop and pest management. Calif Agric. 1983;37:4–7.
Gennard DE. Forensic entomology: an introduction. Chichester: Wiley; 2007.
Campobasso C, Introna F. Reply: commentary on letter to the editor from Jeffrey Wells. J Med Entomol. 2014;51:492–4.
Acknowledgments
We thank Claudio J. Von Zuben, Marcio R. Pie, Rodrigo F. Krüger, David R. Luz, and Kevin A. Williams who provided helpful comments on a previous version of the paper. This research was partially supported by grants from the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (process 312357/2006 and process 307947/2009-2) to M.O.M and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) to M.C.L. James J. Roper provided a careful review of the English language.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Lecheta, M.C., Thyssen, P.J. & Moura, M.O. The effect of temperature on development of Sarconesia chlorogaster, a blowfly of forensic importance. Forensic Sci Med Pathol 11, 538–543 (2015). https://doi.org/10.1007/s12024-015-9727-z
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12024-015-9727-z