
Abstract
Under continuous no-till, gypsum has been successfully used to manage soil fertility and improve crop yield. Nitrogen (N) fertilization is critical for crop performance, however, since it is expensive and potentially pollutant, it must be correctly applied. Both subjects have been widely studied, yet there is a lack of information on the interaction between them, especially concerning crops in rotation or succession. The objectives of this study were to evaluate topdressing N fertilization (0, 50, and 100 kg N ha−1) on black oat (Avena strigosa Schreb.) under no-till, inside a long-term gypsum experiment (0, 3, 6, 9, and 12 Mg ha−1). We evaluated black oat regrowth after haylage harvest and nutrient concentration and uptake by aboveground biomass, as well as the effects of this system on the successor crop soybean (Glycine max (L.) Merr.) in terms of leaf nutrient concentration and grain yield over two growing seasons. The gypsum application, even up to 44 and 55 months earlier, presented a long-term effect when associated with N fertilization, causing black oat biomass to increase during the regrowth phase. Nitrogen fertilization increased black oat regrowth biomass, even without gypsum. Greater nutrient uptake by black oat regrowth occurred under higher gypsum rates and N fertilization, which may have been advantageous in the long term for the production system, once nutrient cycling may partially substitute fertilizer-derived nutrients. Soybean yield was not affected by either long-term gypsum or N applied at black oat tillering, even though some leaf nutrient concentrations were influenced.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
In subtropical environments, such as Southern Brazil, black oat (Avena strigosa Schreb.) and soybean (Glycine max (L.) Merr.) are commonly cultivated in succession under no-till. Soybean is grown for grain production, while black oat may be used for cattle grazing or feeding and as a cover crop. To cope with the increasing global demand for food (Ray et al., 2013), improvements in the management practices and yield of these crops are constantly sought. Likewise, nitrogen (N) fertilization and gypsum application have been widely studied and used to manage soil fertility in subtropical conditions, but their interaction has been underexplored, especially in the long term.
In no-till, the soil permanent cover by plant biomass or residues reduces soil and nutrient losses, besides increasing soil organic matter, especially in tropical and subtropical environments (Nunes et al., 2011; Pereira et al., 2009). To avoid soil disturbance, crop residues and soil amendments are not incorporated into the soil by tillage. Thus, nutrients get concentrated near the soil surface, and a vertical fertility gradient builds up over time, with less available basic cations and phosphorus (P) in subsurface soil, which gets more acidic (Schlindwein et al., 2013; Zoca & Penn, 2017) and restrains root growth in depth (Bortoluzzi et al., 2014; Zandoná et al., 2015).
Gypsum application improves the distribution of nutrients in the soil profile, as it contains calcium (Ca2+) and sulfur (S) in a soluble compost (2.5 g L−1) and chemical form (CaSO4) that allows percolation to subsurface layers (Zoca & Penn, 2017). The reaction of CaSO4 with the soil reduces aluminum (Al3+) activity, which alleviates subsurface acidity effects and redistributes basic cations like magnesium (Mg2+) and potassium (K+) from surface to subsurface layers (Caires et al., 2016; Ramos et al., 2013; Vicensi et al., 2020a, b). This change in soil chemical conditions enhances root growth (Caires et al., 2016; Vicensi et al., b) and crop tolerance to water stress (Dalla-Nora & Amado, 2013; Zandoná et al., 2015).
Some studies have reported that gypsum application increased biomass of black oat and other Poaceous crops (Soratto & Crusciol, 2008a; Vicensi et al., 2020a, b), besides improving soil fertility, especially in no-till soils (Caires et al., 2016; Ramos et al., 2013; Soratto & Crusciol, 2008a). In such conditions, N fertilization is also reported to increase black oat biomass (Bassegio et al. 2013; Menezes et al., 2013; Restelatto et al., 2014; Steiner et al., 2009) and quality (Basi et al., 2011), through higher nutrient uptake and concentration and lower C:N rate, which improves nutrient cycling (Restelatto et al., 2015; Santi et al., 2003).
The production of cover and forage crops in winter contributes to sustainable management, as it adds plant diversity to the production systems and organic matter to the soil, promoting nutrient cycling and improving soil quality (Balbinot Junior et al. 2009). Black oat may provide high-quality feed for cattle (Restelatto et al., 2014). Moreover, the plant residues can release significant amounts of N, P, and K, which are majorly available up to 30–40 days from crop management (biomass deposition on the soil surface) (Crusciol et al., 2008), benefiting grain yield of succeeding crops, such as soybean (Caires et al., 2017). When black oat is cultivated for forage in tropical and subtropical production systems, the plants can regrow for at least 40 days before planting the summer crops—sufficient time for a considerable biomass and nutrient accumulation (Vega-García et al., 2020).
Black oat is known for its hardiness and tolerance to poor soil conditions (Silva et al., 2006), having a strong root system (Burr-Hersey et al., 2017; Müller et al., 2001). It is cultivated in no-till areas for crop rotation, grazing, haying, silage, and grain production in Southern Brazil (Ziech et al., 2015). This crop has the potential to accumulate up to 7.8 Mg ha−1 of aboveground biomass (Michelon et al., 2019), or even more, depending on the environment and management practices. It serves as green manure since the absorbed nutrients are cycled, improving nutrient distribution in the soil profile and contributing to the efficiency of continuous no-till (de Faccio Carvalho et al., 2010).
Concerning N rates, in the subtropical areas of Southern Brazil, black oat biomass has been reported to increase with up to 120 kg N ha−1 (Bassegio et al. 2013; Steiner et al., 2009) and even 180 kg N ha−1 (Restelatto et al., 2014; Santi et al., 2003), depending upon production conditions. However, due to the high costs of N fertilization, growers apply none or much lower amounts of N than required, posing a presumable constraint to higher yields. In contrast, the negative effects of N loss processes are well known and there have been researching efforts focusing on N use efficiency (Zhang et al. 2015), so that lower and more environmentally friendly rates of N may be used, in consonance to economic viability and yield levels.
Since gypsum improves root growth of plants (Caires et al., 2016), including black oat (Vicensi et al., 2020b), and N can be easily leached through the soil profile in the nitrate (NO3−) form under normal aeration conditions (Bortolotto et al., 2012; Caires et al., 2015), we hypothesized that gypsum application could have a synergistic effect when associated with N fertilization on black oat regrowth and nutrient absorption, as well as positive effects on succeeding crops, such as soybean (Glycine max (L.) Merrill), the main cash crop in the summer season.
The objectives of this study were to evaluate topdressing N-fertilization on black oat under no-till, inside a long-term gypsum experiment, assessing black oat regrowth after haylage harvest and nutrient concentration and uptake by aboveground biomass, as well as the possible beneficial effects of this system in the successor crop soybean, in terms of leaf nutrient concentration and grain yield, over two growing seasons.
Materials and Methods
The experiment was set up in 2009, in Guarapuava, Paraná, Brazil (25º23′ S, 51º30′ W, altitude 1026 m), where the climate is humid subtropical mesothermic (Cfb—Köppen) (Alvares et al., 2013). The field was cultivated under the continuous no-till system for more than 11 years prior to the growing seasons of 2015 and 2016, which is the period considered for the present study.
In Oct. 2009, after the evaluation of soil morphology, clay content, and chemical attributes (Michalovicz et al., 2019), it was classified as very clayey Typic Hapludox (USDA, 1999), corresponding to Latossolo Bruno according to the Brazilian Soil Classification System (Embrapa, 2018), with low acidity in A horizon, low Al3+ levels up to 1.4 m, and no physical limitations (bulk density ≤ 1.1 Mg m−3). The chemical analysis and clay determination of the soil diagnostic layer performed in Nov. 2009 is presented in Table 1. Soil chemical conditions up to 0.8 m just prior to 2015 and 2016 growing seasons were previously published by Vicensi et al. (2020b).
Experimental Design
The experiment was carried out in a randomized block design in a split-plot arrangement, with four replications. The main plots consisted of gypsum rates: 0 (control, G0), 3 (G3), 6 (G6), 9 (G9), and 12 (G12) Mg ha−1 of dry weight, broadcasted on the soil surface and split into three equal annual applications, one third-rate each, in Nov. 2009, Nov. 2010, and Nov. 2011, just after sowing summer crops. These rates were calculated to provide 0, 33, 66, 100, and 133% of the required amount of Ca2+ to reach 60% of Ca saturation on the cation exchange capacity (at pH 7.0) of the A1 horizon of the soil (Michalovicz et al., 2019).
The subplots consisted of N-fertilization rates applied as top-dressings at the beginning of the black oat tillering stage: 0 (control, N0), 50 (N50), and 100 (N100) kg ha−1 of N. The N source used was urea [(CO(NH2)2), 45% of N]. The main plot size was 64 m2 (10 × 6.4 m), and subplots had 21.31 m2 (3.33 × 6.40 m).
Black Oat Crop
The black oat cultivar IAPAR 61 was sown on April 15 and April 17 in the 2015 and 2016 growing seasons, respectively, succeeding soybean in both periods. As additional information, the same variety was sown in the same experimental setup before this study, on April 15, 2014. The seeding rate was 360 seeds per m2, with a row spacing of 0.17 m. The soil was fertilized with 40 kg ha−1 of P2O5 (triple superphosphate) in furrow and 40 kg ha−1 of K2O (potassium chloride) broadcasted on the soil surface after sowing.
The plots were harvested (haylage) at 115 and 120 days after sowing in the 2015 and 2016 growing seasons, and black oat regrew for 30 days before the evaluation. After this period, a 0.51 m2 area on each experimental unit was cut at 0.10 m from the soil surface to evaluate the aboveground biomass. Samples were oven-dried at 50 °C, ground (Wiley), and sieved (0.75 mm). Subsequently, they were analyzed for nitrogen (N), after sulfuric digestion (digestion block), and for phosphorus (P), potassium (K), calcium (Ca), magnesium (Mg), and sulfur (S) after nitric-perchloric digestion (Malavolta et al., 1997). N, P, and S were determined by Kjeldahl distillation, colorimetry, and turbidimetry methods, respectively. Ca and Mg were determined by atomic absorption spectrometry, while K was determined by emission flame photometry. Nutrient uptake by black oat regrowth was estimated by multiplying the nutrient concentration in the aboveground biomass (g kg−1) by the biomass yield (kg ha−1).
Soybean Crop
The soybean cultivar Syngenta 1561 was sown on Nov. 30 and Dec. 4 in the 2014/2015 and 2015/2016 growing seasons, respectively, succeeding black oat in both years. The seeding rate was 37.5 seeds per m2, with a row spacing of 0.4 m. Basic fertilization was performed with 60 kg ha−1 of P2O5 (triple superphosphate) in furrow and 60 kg ha−1 of K2O (potassium chloride) broadcasted on the soil surface after sowing. Using a turfy inoculant on the sowing day, seeds were inoculated with Bradyrhizobium japonicum, with ± 1.2 million viable cells per seed.
When soybean plants reached the R2 growth stage (Fehr & Caviness, 1977), 30 recently fully expanded trifoliate leaves were collected with the petiole in each experimental unit. These samples were oven-dried at 50 °C, ground (Wiley), sieved (0.75 mm), and then, analyzed for the same nutrients and using the same methods as described for black oat biomass samples. At the R8 growth stage, a 4.8 m2 area in the center of each experimental unit was manually harvested to evaluate soybean grain yield. Grain mass was adjusted to 130 g kg−1 grain moisture.
Meteorological Data
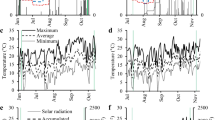
Daily meteorological data were obtained from a meteorological station (SIMEPAR/Brazil) located 200 m away from the experiment (Fig. 1). A sequential water balance proposed by Thornthwaite and Matter (1955) was calculated (Fig. 1a).

Meteorological data from Nov. 2014 to Sep. 2016: A daily rainfall (blue bars), accumulated rainfall (orange line), potential evapotranspiration (ETP, red line), actual evapotranspiration (ETA, black line) across the growing seasons; B daily maximum (TMAX, red), average (TAVG, orange), and minimum (TMIN, light blue) temperatures; C daily total solar radiation energy and solar radiation accumulated over the growing periods. Black arrows point to key dates
Over the black oat regrowth periods, the cumulative rainfall was 46 and 227 mm, and the average temperature was 16.6 and 14.7 °C during the 2015 and 2016 growing seasons, respectively (Fig. 1a, b). The higher frequency of lower minimum temperatures in 2016 evinces the greater occurrence of frosts observed in this growing season (Fig. 1b). The accumulated solar radiation during black oat regrowth was similar in both periods (456 and 413 MJ m−2) (Fig. 1c). The difference of actual evapotranspiration to potential evapotranspiration (ETa—ETp) had a negative balance of 63.5 and 0 mm during black oat cultivation in 2015 and 2016, respectively.
Concerning the soybean cropping seasons, the cumulative rainfall reached 813 and 887 mm, and the average temperature was 21.0 and 21.2 °C during the 2014/2015 and 2015/2016 growing seasons, respectively (Fig. 1a, b). The accumulated solar radiation were 1982 and 1759 MJ m−2 during these periods (Fig. 1c). The balance of ETa—ETp was negative, with values of −54.6 and −204 mm during the 2014/2015 and 2015/2016 soybean growing seasons, respectively (Fig. 1a).
Statistical Analysis
These data were submitted to the two-way analysis of variance (ANOVA) (α = 0.05) in a split-plot scheme. The gypsum treatments were fitted to linear or quadratic regression models, based on the highest coefficient of determination (r2) when significant at p ≤ 0.05 by the F test. N-fertilization treatments were classified by the Tukey test (α = 0.05) when significant. When interaction effects were detected, the degrees of freedom were unfolded.
Results
Black Oat Regrowth
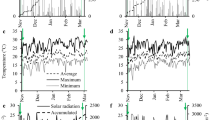
An interaction effect was detected between gypsum and N-fertilization for black oat aboveground biomass after the regrowth period in both growing seasons (Fig. 2). In 2015, the interaction within N-fertilization was as follows: within N0, the regrowth biomass did not differ among gypsum rates; within N50, a quadratic model fitted best the gypsum rate data, where the maximum estimated biomass yield was 2.55 Mg ha−1, at an estimated gypsum rate of 6.26 Mg ha−1, corresponding to 84, 96, 99, 97, and 87% of biomass yield achieved by G0, G3, G6, G9, and G12, respectively; and within N100, a linear model was the better-fitting model for gypsum rates, where the maximum biomass yield achieved was 3.02 Mg ha−1 at a rate of 12 Mg ha−1 of gypsum, corresponding to 73, 94, 84, 94, and 100% of biomass yield achieved by G0, G3, G6, G9, and G12, respectively (Fig. 2a). As for the interaction within gypsum rates, the results were as follows: in G0 N100 = N50 > N0; in G3 N100 > N50 > N0; in G6 N100 = N50 > N0; in G9 N100 = N50 > N0; and in G12 N100 > N50 = N0 (Fig. 2a).

Black oat regrowth biomass 30 days after harvest (at the early heading stage) in (A) 2015 and (B) 2016 growing seasons, in response to the residual effect of gypsum applied between Nov. 2009 and Nov. 2011, and nitrogen rates (N0, N50, and N100, corresponding to 0, 50, and 100 kg ha−1 of N) top-dressed at tillering, in a no-till field in Guarapuava, Paraná State, Brazil. *, ** and ns indicate significant values at p ≤ 0.05 and p ≤ 0.01, and non-significant values, respectively; bars represent the mean significant difference (MSD) by the Tukey test at p < 0.05 to compare N-fertilization treatments
Concerning the 2016 growing season, looking at the interaction within N-fertilization, the interactive effects were as follows: within N0, the regrowth biomass did not differ among gypsum rates; within N50, gypsum rate data fitted a quadratic model, where the maximum estimated biomass yield was 1.55 Mg ha−1, at an estimated gypsum rate of 6.76 Mg ha−1, corresponding to 69, 87, 102, 97, and 80% of biomass yield achieved by G0, G3, G6, G9, and G12, respectively; and within N100, a linear model was the better fit for gypsum rates, where the maximum biomass yield was 1.68 Mg ha−1 with 12 Mg ha−1 of gypsum, corresponding to 85, 89, 91, 106, and 94% of the biomass yield achieved by G0, G3, G6, G9, and G12, respectively (Fig. 2a). Whereas for the interaction within gypsum rates, the results were as follows: within G0 N100 > N50 = N0; within G3 N100 = N50 > N0; within G6 N100 = N50 > N0; within G9 N100 = N50 > N0; and within G12 N100 > N50 = N0.
Nutrient Concentration in Black Oat Biomass
The concentrations of N, P, K, Ca, Mg, and S in black oat regrowth biomass had no interaction between gypsum and N-fertilization in both growing seasons (Table 2). The biomass concentrations of N, P, and K were not influenced by gypsum rates, while Ca, Mg, and S concentrations were affected by gypsum in both growing seasons. Ca and S concentrations increased linearly with gypsum rates in both cultivation periods. Whereas Mg concentration decreased in a linear fashion in response to the increase in gypsum rates in 2015 and 2016 (Table 2). The concentrations of K, Mg, and S were not influenced by N-fertilization. N concentration between N50 and N100 produced no significant effect, and both were higher than N0 in 2015. As for the 2016 growing season, N concentration increased according to the increase in N-fertilization (N100 > N50 > N0). P concentration was lower in N100 than in N0 in 2015. Whereas in 2016, P concentration did not differ between N50 and N100, and both were lower in relation to N0. Ca concentration was higher in N100 in relation to N0 in both growing seasons (Table 2).
Black Oat Nutrient Uptake
The uptake of N, P, K, Ca, Mg, and S by black oat regrowth presented no interaction effect between gypsum and N-fertilization in both growing seasons (Table 3). Mg uptake was not influenced by gypsum rates. N uptake increased linearly with gypsum rates in 2015 and quadratically in 2016. P, Ca, and S uptake increased in a linear fashion with increasing gypsum rates in both growing seasons, and K uptake increased linearly in 2016 (Table 3).
Black oat regrowth uptake of N, P, K, Ca, Mg, and S were higher with N-fertilization in both growing seasons (Table 3). N, Ca, and Mg uptake increased with each further N rate (N100 > N50 > N0) in 2015 and 2016. P, K, and S uptake between N50 and N100 produced no significant effect, and both were higher in relation to N0 in 2015. In 2016, higher levels of P, K, and S were absorbed with each further N rate (N100 > N50 > N0) (Table 3).
Leaf Nutrient Concentration in Soybean at R2
Soybean leaf nutrient concentrations had no interaction effect between gypsum and N-fertilization in both growing seasons (Table 4). Leaf concentrations of N, P, and K were not influenced by gypsum rates, whereas Ca and S concentrations increased linearly with the increase in gypsum rates in both cultivation periods. In contrast, the concentration of Mg decreased linearly in response to the increase in gypsum rates in 2015 and 2016 (Table 4).
The leaf concentration of N, K, Ca, Mg, and S were not influenced by N-fertilization in the two seasons evaluated (Table 4). The concentration of P between N50 and N100 produced no significant effect, and both were lower in relation to N0 in 2015. In 2016, P concentration in N100 was lower than N0, while N50 did not differ from both rates.
Soybean Grain Yield
In both growing seasons, soybean yield was not affected by either gypsum or N-fertilization applied in prior black oat cultivation (Fig. 3). In the 2015/2016 growing season, soybean yield was 17% higher than in 2014/2015.

Soybean yield in response to the residual effect of gypsum rates applied in Nov. 2011 and black oat nitrogen (N) topdressing in (A) 2014/2015 and (B) 2015/2016 growing seasons, in a no-till field in Guarapuava, Paraná State, Brazil. Prior to soybean sowing, black oat was harvested at the early heading stage, regrew for 30 days, and burnt down by herbicide. ns indicates non-significant values; bars represent the mean significant difference (MSD) by the Tukey test at p < 0.05 to compare N-fertilization treatments
Discussion
Black Oat Regrowth
Overall, N-fertilization increased black oat aboveground biomass compared to the control, and in both growing seasons, N100 led to a greater increase than N50 when combined to low (G0 and G3) or high (G12) gypsum rates (Fig. 2a, b). This crop accumulated more aboveground biomass in response to the long-term effect of gypsum depending on N-fertilization rate (Fig. 2a, b). In 2015, the aboveground biomass within N100 increased across gypsum rates in relation to G0 from 16 to 37%. In 2016, the black oat aboveground biomass was 17 to 48% and 4 to 36% higher, within N50 and N100, respectively, across gypsum rates in relation to G0.
Gypsum effects depend on the time course after application and the season conditions. In the same region of Paraná State, the biomass of white oat intercropped with ryegrass was not affected by gypsum applied 3 months prior to evaluation (Silva et al., 2015), possibly due to lack of time for the product to react in the soil. Under water deficit conditions, black oat presented a biomass yield 13% greater than the control in response to 2.1 Mg ha−1 of gypsum applied 10 months before the beginning of the experiment (Soratto & Crusciol, 2008b). Our results showed that gypsum has long-term effects, increasing black oat biomass after 44 and 55 months of the application of the last one-third of the rates (Nov. 2011).
Taking into account both seasons, N-fertilization with N50 and N100 at tillering increased black oat regrowth biomass by 30 and 49%, on average, respectively, in comparison with the control. Another study pointed out that N-fertilization (urea) increased black oat aboveground biomass by 63% with 100 kg N ha−1 and by 77% with 180 kg N ha−1 (Santi et al., 2003). Using the same cultivar of the present study, Steiner et al. (2009) observed that the application of 100 and 120 kg N ha−1 (urea) led to increases of 103 and 124%, respectively, in black oat aboveground biomass. Both studies (Santi et al., 2003; Steiner et al., 2009) assessed the trait at the grain formation phase, whereas this study evaluated the plants 30 days after haylage harvest at the early heading stage, focusing on black oat regrowth for soil cover and nutrient cycling for the successor crop.
Black oat was preceded by soybean in both seasons in this study, and mineralized-N from soybean straw possibly has partially met the black oat N demand, which resulted in a lower relative response to N-fertilization than the observed in other studies, including the same rate of N100. Soybean straw of a medium yield crop may reach 90 kg N ha−1 (Balboa et al., 2018), which will be made available through the mineralization process in the soil, providing N for the following crop (Siqueira-Neto et al., 2010). Black oat would possibly have a greater response to N fertilization if preceded by a crop with a higher C:N ratio, such as corn.
The 41% lower aboveground biomass yield in 2016 compared to 2015, considering the experimental average, may be partly explained by the different meteorological conditions between the growing seasons. In 2016, frost occurred more frequently along with lower average temperature and lower accumulated solar radiation in relation to 2015 (Fig. 1).
Nutrient Concentration in Black Oat Biomass
The concentrations of P, K, Ca, and S in the 2015 growing season showed average decreases of 34, 21, 14, and 20%, respectively, compared to the 2016 growing season. This may be partially explained by the dilution effect from plant growth, as the biomass in 2015 was, on average, 70% higher compared to the one obtained in the 2016 growing season.
The N concentration in 2015 was, on average, 12% higher compared to 2016 (Table 2). In 2016, the temperature during black oat regrowth was, on average, 2.5 °C lower than in 2015 (Fig. 1b), which may have reduced the mineralization rate of soil organic matter and crop residues, decreasing N availability. In this condition, N uptake by plants depends more on N fertilizer, which may have favored the difference between N50 and N100 applied in black oat tillering in 2016. The reduction in P concentration due to N-fertilization rates may be due to a dilution effect from plant growth, as the increase in N-fertilization rates increased biomass produced from black oat regrowth (Fig. 2a, b).
It is possible that the absorption of N and Ca interacted synergistically, as these nutrients presented an equivalent response to N-fertilization. In few weeks after application, N from urea (used in top dressing) is transformed into NO3− in the soil through nitrification (Bremner & Blackmer, 1978). The non-antagonistic interaction between cations and anions promotes simultaneous transport (symport) of both ions through cell membranes, therefore, the absorption of NO3− enables the passive absorption of cations like Ca and K (Andres et al., 2016). N is related to photosynthesis and protein synthesis in plants, while Ca is necessary for cell division and expansion (Yong et al., 2010), so the higher availability of Ca and N, provided by gypsum and N-fertilization, explains the greater black oat growth evaluated by the aboveground biomass.
The reduction in Mg concentration in black oat biomass with the increase in gypsum rate is related to the competitive inhibition of Mg2+ absorption by higher levels of Ca2+ in the soil (Soratto & Crusciol, 2008a). N-fertilization is associated with Mg deficiency in plants, which has the potential to reduce plant development (Soratto & Crusciol, 2008b; Soratto et al., 2010). However, within the N-fertilization rates used in this study, up to 100 kg N ha−1, this effect was not observed in black oat regrowth.
Higher regrowth biomass related to gypsum rates may also be due to higher S availability. In addition, Ca2+ from gypsum benefits nutrient uptake by roots (Siddiqui et al., 2013) and root growth and architecture (Cao et al., 2013), thus contributing to plant growth.
Black Oat Nutrient Uptake
Proving its hardiness (Silva et al., 2006) and strong root system (Burr-Hersey et al., 2017), black oat thrived in a 30-day period of regrowth after harvest and absorbed significant amounts of nutrients from the soil profile, especially of N and K (Table 3). Black oat roots reach 80 cm depth (Vicensi et al., 2020b), and even with only 46 mm of rain during the regrowth period in 2015, more nutrients were absorbed in this growing season by the aboveground biomass than in 2016, when the regrowth period was rainier but also colder.
The increase of black oat regrowth due to the residual effects of gypsum and N-fertilization resulted in greater nutrient uptake by aboveground biomass. Gypsum rates increased the black oat uptake of N, P, K, Ca, and S in both growing seasons (Table 3), even without affecting regrowth biomass concentrations of N, P, and K (Table 2), meaning that biomass yield was determinant. The biomass concentration of Mg was linearly decreased by gypsum rates (Table 2), but this effect was offset by concomitant biomass increase (Fig. 2) and Mg uptake was not affected by gypsum in both growing seasons (Table 3). These results show the importance of crop rotation and winter forage crops for nutrient cycling since sufficient time is given for plant regrowth.
N-fertilization at oat tillering increased the uptake of N, P, K, Ca, Mg, and S during the regrowth phase after harvest, even without affecting Mg and S concentration on the aboveground biomass. In other studies, N-fertilization at rates up to 100 or 120 kg ha−1 has also increased N, P, and K uptake by black oat, proportionally to biomass increase (Melo et al., 2011; Restelatto et al., 2015).
The beneficial outcomes of N-fertilization and residual effects of gypsum to the production system include better soil cover by larger straw deposition and improved nutrient availability to the following crops (Torres et al., 2008), with special advantages for crops with a surface root system (Fan et al., 2016). Nutrients on the aboveground plant residues are deposited on the soil surface for subsequent decomposition. The half-life time for nutrient release from black oat residue was calculated in 15.4, 10, 9, 7.8, and 8.7 days for N, P, K, Ca, and Mg, respectively, and the highest mineralization rate and release of nutrients occurred approximately 30 days after the deposition of the residues (Ferreira et al., 2014).
Leaf Nutrient Concentration in Soybean at R2
Leaf N concentration ranged from 33 to 41 g kg−1 and remained above the leaf sufficiency range suggested by Urano et al. (2006), and below the ranges suggested by Malavolta et al. (1997) and SBCS/NEPAR (2019) (Table 5) in both growing seasons. No effect of black oat N-fertilization was detected and the equivalence in the comparison of N0 with N50 and N100 shows that biological fixation was not affected, providing adequate N supply for soybean. Most of the soybean N is derived from the symbiotic nitrogen fixation (Hungria et al., 2006), and N concentration in soybean leaves is usually insensitive to either residual effects of N fertilizer in predecessor crops or gypsum rates (Caires et al., 2011; Vicensi et al., 2016).
As for leaf P concentration, values ranged from 4.0 to 5.5 g kg−1 and remained above the leaf sufficiency range suggested by Urano et al. (2006), Malavolta et al. (1997), and SBCS/NEPAR (2019) (Table 5) in both growing seasons. Although P concentration in soybean leaves remained above the leaf sufficiency range, it was reduced by N-fertilization rates. As N-fertilization increased P uptake by black oat regrowth, it possibly reduced available P in the soil, and black oat biomass P was not mineralized in time for soybean absorption. Furthermore, as urea-derived N is subjected to nitrification processes, the soil acidification side effect may have also decreased soil available P due to P fixation reactions in the soil (Jones, 2012; Viviani et al., 2010).
Leaf K concentrations ranged from 18 to 20 g kg−1 and remained within the leaf sufficiency range suggested by Urano et al. (2006) and Malavolta et al. (1997), and below the one suggested by SBCS/NEPAR (2019) (Table 5) in both growing seasons. As this last reference range is more recent, changes in the sufficiency limits may be the cause of this variation. Since black oat was harvested for haylage and nutrients were exported from the field and K uptake by black oat regrowth was the second highest (after N), K availability may have been decreased in some soil layers during soybean first stages of growth. K fertilization for soybean was performed with 60 kg ha−1 of K2O broadcasted on the soil surface after sowing. Considering the management conditions, K fertilization rate and application (broadcast or furrow) may be reviewed for the following crops.
Regarding leaf Ca concentrations in the 2015 growing season, values ranged from 6.0 to 7.3 and were below the leaf sufficiency range suggested by Urano et al. (2006) and SBCS/NEPAR (2019), and above the one suggested by Malavolta et al. (1997) (Table 5). In 2016, leaf Ca concentrations had values ranging from 9 to 11 g kg−1 and remained within or above the leaf sufficiency range suggested by Malavolta et al. (1997), Urano et al. (2006), and SBCS/NEPAR (2019) (Table 5). Higher gypsum rates increased Ca concentration linearly in both growing seasons (Table 4) just as occurred for black oat regrowth (Table 2).
For leaf Mg concentrations, numbers from 1.6 to 1.9 g kg−1 were observed and remained below or within the leaf sufficiency range suggested by Urano et al. (2006), and below the ones suggested by SBCS/NEPAR (2019) and Malavolta et al. (1997) (Table 5). The increase of gypsum rates decreased Mg concentration in a linear fashion (Table 4), which also occurred for black oat regrowth (Table 2). Gypsum mobilizes Mg2+ from surface layers to subsurface soil layers (Caires et al., 2011; Vicensi et al., 2020b). In the present study, Mg2+ levels were reduced in the soil up to 80 cm depth in response to gypsum (Vicensi et al., 2020a, 2020b). There is also competitive inhibition of Ca2+ supplied by gypsum over soil Mg2+ and K+ (Jones, 2012). These results show that leaf Mg concentrations were predominantly below the leaf sufficiency ranges for soybean, especially when gypsum rates increased, evidencing that after many years of gypsum application, Mg is limiting crop yield and must be resupplied.
Concerning sulfur, leaf S concentrations ranged from 2.9 to 3.7 g kg−1 and remained within or above the leaf sufficiency range suggested by Malavolta et al. (1997), Urano et al. (2006), and SBCS/NEPAR (2019) in both growing seasons (Table 5). Increasing gypsum rates linearly increased S concentration in soybean leaves (Table 4), following the same pattern observed for black oat regrowth (Table 2). Gypsum provides Ca2+ and SO42−-S to the soil, increasing Ca and S concentrations in plant tissues, as observed in other studies (Caires et al., 2011; Moda et al., 2013).
Our results of leaf nutrient concentrations compared to leaf sufficiency ranges (Malavolta et al., 1997; SBCS/NEPAR, 2019; Urano et al., 2006) evidence that K, Ca, and Mg for the 2014/2015 season and K and Mg in the 2015/2016 season may have limited soybean development and yield.
Among the elements evaluated in the diagnostic leaf (Table 4), only Mg was below or at the lower limit of the sufficiency range (Table 5), suggesting that this nutrient may have limited the performance of soybean, even though these plants showed no deficiency symptoms regarding this nutrient. The critical level of Mg2+ is 1 cmolc dm−3 in soil (SBCS/NEPAR, 2019) and after years of studies performed in this site, without liming and without Mg fertilization, Mg2+ levels were reported below or close to the critical level in gypsum and N treatments (Vicensi et al., 2020b), which corroborates the diagnostic leaf Mg concentration results.
Soybean Grain Yield
Soybean yield was not affected by either the residual effect of gypsum or N application at black oat tillering in both growing seasons. The lower soybean yield in 2014/2015 may be due to excess rainfall in February when soybean was in the reproductive stages. Excessive rainfall coupled with a higher incidence of cloudy days reduce plant photosynthetic activity, restrain nutrient absorption, and increase soybean flower abortion (Kokubun, 2011). In the 2014/2015 growing season, soybean presented lower leaf concentrations of N, P, and Ca than in 2015/2016, which are related to yield potential.
The overall averages of soybean grain yield in this study were 3.55 and 4.15 Mg ha−1 in 2014/2015 and 2015/2016 growing seasons, respectively. Both periods had higher yields than the average of Paraná state on the respective seasons, reaching 3.29 Mg ha−1 for 2014/2015 and 3.73 Mg ha−1 for 2015/2016 (CONAB et al., 2021).
Gypsum commonly has no effect on soybean yield under medium yield potential conditions (Pauletti et al., 2014; Somavilla et al., 2016; Vicensi et al., 2016) and under regular rainfall distribution or irrigation (Caires et al., 2006; Henry et al., 2018; Vicensi et al., 2016). However, in low yield potential conditions (< 2.5 Mg ha−1) with water deficit, gypsum may increase soybean yield (Henry et al., 2018; Schenfert et al., 2020). Under drought, an increase of 11.3% in soybean yield in response to a gypsum application of 2 Mg ha−1 was reported by Zandoná et al. (2015). Further studies are necessary to evaluate the applicability of gypsum in the soybean crop at different levels of magnesium above the critical level.
Soybean has a higher cation exchange capacity of roots when compared to Poaceous crops such as corn (Fernandes & Souza, 2006), which leads to a higher accumulation in the rhizosphere of soil divalent cations, such as Mg2+ and Ca2+, favoring the absorption of these nutrients even under low availability in the soil. This feature may partly explain the different response of this crop to long-term gypsum when compared to black oat.
Conclusions
Our hypothesis that gypsum application associated with N fertilization could have a synergistic effect has been confirmed for black oat regrowth biomass, but not for soybean yield. The gypsum application, even up to 44 and 55 months earlier than the experiment, presented a long-term effect when associated with N fertilization, causing black oat biomass to increase during the regrowth phase. Nitrogen fertilization increased black oat regrowth biomass, even without gypsum. Nutrient uptake levels were greater for black oat regrowth due to higher gypsum rates and N fertilization, which may have been advantageous in the long term for the production system, once nutrient cycling may partially substitute fertilizer-derived nutrients. Despite their effects on soybean foliar diagnosis, soybean yield was not affected by either long-term applied gypsum or by N applied at black oat tillering.
References
Alvares, C. A., Stape, J. L., Sentelhas, P. C., de Moraes, G., Leonardo, J., & Sparovek, G. (2013). Köppen’s climate classification map for Brazil. Meteorologische Zeitschrift. https://doi.org/10.1127/0941-2948/2013/0507
Andres, C., Comoé, H., Beerli, A., Schneider, M., Rist, S., & Jacobi, J. (2016). Cacao nutrition and fertilization. In E. Lichtfouse (Ed.), Sustainable agriculture reviews (pp. 155–202). Springer.
Balbinot-Junior, A. A., Moraes, A., Veiga, M., Pelissari, A., & Dieckow, J. (2009). Crop-livestock system: Intensified use of agricultural lands. Ciência Rural. https://doi.org/10.1590/S0103-84782009005000107 in Portuguese with abstract in English.
Balboa, G. R., Sadras, V. O., & Ciampitti, I. A. (2018). Shifts in soybean yield, nutrient uptake, and nutrient stoichiometry: A historical synthesis-analysis. Crop Science. https://doi.org/10.2135/cropsci2017.06.0349
Basi, S., Neumann, M., Marafon, F., Ueno, R. K., & Sandini, I. E. (2011). Influência da adubação nitrogenada sobre a qualidade da silagem de milho. Pesquisa Aplicada & Agrotecnologia. https://doi.org/10.5777/paet.v4i3.1433
Bortolotto, R. P., Bruno, I. P., Reichardt, K., Timm, L. C., Amado, T. J. C., & Ferreira, A. O. (2012). Nitrogen fertilizer (15N) leaching in a central pivot fertigated coffee crop. Revista Ceres. https://doi.org/10.1590/S0034-737X2012000400006
Bortoluzzi, E. C., Parize, G. L., Korchagin, J., Silva, V. R., Rheinheimer, D. S., & Kaminski, J. (2014). Soybean root growth and crop yield in response to liming at the beginning of a no-tillage system. Revista Brasileira De Ciência Do Solo. https://doi.org/10.1590/S0100-06832014000100026
Bremner, J. M., & Blackmer, A. M. (1978). Nitrous oxide: Emission from soils during nitrification of fertilizer nitrogen. Science. https://doi.org/10.1126/science.199.4326.295
Burr-Hersey, J. E., Mooney, S. J., Bengough, A. G., Mairhofer, S., & Ritz, K. (2017). Developmental morphology of cover crop species exhibit contrasting behaviour to changes in soil bulk density, revealed by X-ray computed tomography. PLoS ONE. https://doi.org/10.1371/journal.pone.0181872
Caires, E. F., Barth, G., & Garbuio, F. J. (2006). Lime application in the establishment of a no-till system for grain crop production in Southern Brazil. Soil and Tillage Research. https://doi.org/10.1016/j.still.2005.06.006
Caires, E. F., Haliski, A., Bini, A. R., & Scharr, D. A. (2015). Surface liming and nitrogen fertilization for crop grain production under no-till management in Brazil. European Journal of Agronomy. https://doi.org/10.1016/j.eja.2015.02.008
Caires, E. F., Maschietto, E. H. G., Garbuio, F. J., Churka, S., & Joris, H. A. W. (2011). Surface application of gypsum in low acidic Oxisol under no-till cropping system. Scientia Agricola. https://doi.org/10.1590/S0103-90162011000200011
Caires, E. F., Sharr, D. A., Joris, H. A. W., Haliski, A., & Bini, A. R. (2017). Phosphate fertilization strategies for soybean production after conversion of a degraded pastureland to a no-till cropping system. Geoderma. https://doi.org/10.1016/j.geoderma.2017.08.032
Caires, E. F., Zardo Filho, R., Barth, G., & Joris, H. A. W. (2016). Optimizing nitrogen use efficiency for no-till corn production by improving root growth and capturing NO3-N in subsoil. Pedosphere. https://doi.org/10.1016/S1002-0160(15)60058-3
Cao, X., Chen, C., Zhang, D., Shu, B., Xiao, J., & Xia, R. (2013). Influence of nutrient deficiency on root architecture and root hair morphology of trifoliate orange (Poncirus trifoliata L. Raf.) seedlings under sand culture. Scientia Horticulturae. https://doi.org/10.1016/j.scienta.2013.07.034
CONAB Companhia Nacional de Abastecimento. (2021). Série histórica—Soja. Retrieved March 28, 2021, from https://www.conab.gov.br/info-agro/safras/serie-historica-das-safras
Crusciol, C. A. C., Moro, E., Lima, E. D. V., & Andreotti, M. (2008). Decomposition rate and nutrient release of oat straw used as mulching in no-till system. Bragantia. https://doi.org/10.1590/S0006-87052008000200024 in Portuguese with abstract in English.
Dalla-Nora, D., & Amado, T. J. C. (2013). Improvement in chemical attributes of Oxisol subsoil and crop yields under no-till. Agronomy Journal. https://doi.org/10.2134/agronj2013.0031
de Faccio Carvalho, P. C., Anghinoni, I., Moraes, A., Souza, E. D., Sulc, R. M., Lang, C. R., Flores, J. P. C., Lopes, M. L. T., da Silva, J. L. S., Conte, O., & de Lima Wesp, C. (2010). Managing grazing animals to achieve nutrient cycling and soil improvement in no-till integrated systems. Nutrient Cycling in Agroecosystems. https://doi.org/10.1007/s10705-010-9360-x
Embrapa [Empresa Brasileira de Pesquisa Agropecuária]. (2018). Sistema Brasileiro de classificação de solos = Brazilian soil classification system (in Portuguese) (5th ed.). Embrapa.
Fan, J., McConkey, B., Wang, H., & Janzen, H. (2016). Root distribution by depth for temperate agricultural crops. Field Crops Research. https://doi.org/10.1016/j.fcr.2016.02.013
Fehr, W. R., & Caviness, C. E. (1977). Stages of soybean development (p. 80). Iowa State University.
Fernandes, M. S., & Souza, S. R. (2006). Absorção de nutrientes. In M. S. Fernandes (Ed.), Nutrição mineral de plantas = Plant mineral nutrition (in Portuguese) (1st ed., pp. 115–152). Sociedade Brasileira de Ciência do Solo.
Ferreira, P. A. A., Girotto, E., Trentin, G., Miotto, A., Melo, G. W. D., Ceretta, C. A., Kaminski, J., Frari, B. K. D., Marchezan, C., Silva, L. O. S., & Faversani, J. C. (2014). Biomass decomposition and nutrient release from black oat and hairy vetch residues deposited in a vineyard. Revista Brasileira De Ciência Do Solo. https://doi.org/10.1590/S0100-06832014000500027
Henry, C. G., Sarzi Sartori, G. M., Gaspar, J. P., Marchesan, E., Hirsh, S. M., Horton, A. P., Espinoza, L., & James, H. (2018). Deep tillage and gypsum amendments on fully, deficit irrigated, and dryland soybean. Agronomy Journal. https://doi.org/10.2134/agronj2015.11.0567
Hungria, M., Campo, R. J., Mendes, I. C., & Graham, P. H. (2006). Contribution of biological nitrogen fixation to the N nutrition of grain crops in the tropics: the success of soybean (Glycine max (L.) Merr.) in South America. In R. P. Singh, N. Shankar, & P. K. Jaiwal (Eds.), Nitrogen nutrition and sustainable plant productivity (pp. 43–93). Studium.
Jones, J. B., Jr. (2012). Plant nutrition and soil fertility manual (2nd ed.). CRC Press. https://doi.org/10.1201/b11577
Kokubun, M. (2011). Physiological mechanisms regulating flower abortion in soybean. In T.-B. Ng (Ed.), Soybean—biochemistry, chemistry and physiology (pp. 541–554). IntechOpen. https://doi.org/10.5772/15694
Malavolta E., Vitti, G. C., Oliveira, S. A. (1997). Evaluation of nutritional status of plants: principles and applications. = Avaliação do estado nutricional das plantas: princípios e aplicações. 2ed. Potafos, Piracicaba, SP, Brazil (in Portuguese)
Melo, A. V., Galvão, J. C. C., Braun, H., Santos, M. M., Coimbra, R. R., da Silva, R. R., et al. (2011). Nutrient uptake and biomass production of black oats in soils under eighteen years of organic and mineral fertilizer. Semina Ciências Agrárias. https://doi.org/10.5433/1679-0359.2011v32n2p411
Menezes, L. F. G., Ronsani, R., Pavinato, P. S., Biesek, R. R., Silva, C. E. K., Martinello, C., Cappellesso, B., & Silveira, M. F. (2013). Production, nutritional value and recovery effciencies and nitrogen utilization of corn silage under different doses of nitrogen fertilizer. Semina Ciências Agrárias. https://doi.org/10.5433/1679-0359.2013v34n3p1353-1362
Michalovicz, L., Müller, M. M. L., Tormena, T. A., Dick, W. A., Vicensi, M., & Meert, L. (2019). Soil chemical attributes, nutrient uptake and yield of no-till crops as affected by phosphogypsum doses and parceling in southern Brazil. Archives of Agronomy and Soil Science. https://doi.org/10.1080/03650340.2018.1505041
Michelon, C. J., Junges, E., Casali, C. A., Pellegrini, J. B. R., Neto, L. R., de Oliveira, Z. B., & de Oliveira, M. B. (2019). Soil attributes and yield of corn cultivated in succession to winter cover crops. Revista De Ciências Agroveterinárias. https://doi.org/10.5965/223811711812019230 in Portuguese with abstract in English.
Moda, L. R., Borges, B. M. M. N., Flores, R. A., Santos, C. L. R., Prado, R. M., & Sousa, J. I. (2013). Gypsum application in the soybeans no-tillage system with and without potassium fertilization. Agro@mbiente. https://doi.org/10.18227/1982-8470ragro.v7i2.948 in Portuguese with abstract in English.
Müller, M. M. L., Ceccon, G., & Rosolem, C. A. (2001). Influence of subsurface soil compaction on the growth of the aerial parts and roots of winter green manure species. Revista Brasileira De Ciencia Do Solo. https://doi.org/10.1590/S0100-06832001000300002 in Portuguese with abstract in English.
Nunes, R. S., Sousa, D. M. G., Goedert, W. J., & Vivaldi, L. J. (2011). Phosphorus distribution in soil as affected by cropping systems and phosphate fertilization management. Revista Brasileira De Ciência Do Solo. https://doi.org/10.1590/S0100-06832011000300022 in Portuguese with abstract in English.
Pauletti, V., Pierri, L., Ranzan, T., Barth, G., & Motta, A. C. V. (2014). Long-term effects of the application of gypsum and lime in a no-till system. Revista Brasileira De Ciência Do Solo. https://doi.org/10.1590/S0100-06832014000200014 in Portuguese with abstract in English.
Pereira, R. G., Albuquerque, A. W., & Madalena, J. A. S. (2009). Influence of the soil management systems on the components of corn and brachiaria decumbens production. Revista Caatinga, 22, 64–71. in Portuguese with abstract in English.
Ramos, B. Z., Toledo, J. P. V. F., Lima, J. M., Serafim, M. E., Bastos, A. R. R., Guimarães, P. T. G., & Coscione, A. R. (2013). Gypsum applications to coffee: influence on calcium, magnesium and potassium contents and pH of the solution of a dystrophic red latosol. Revista Brasileira De Ciência Do Solo. https://doi.org/10.1590/S0100-06832013000400019 in Portuguese with abstract in English.
Ray, D. K., Mueller, N. D., West, P. C., & Foley, J. A. (2013). Yield trends are insufficient to double global crop production by 2050. PLoS ONE. https://doi.org/10.1371/journal.pone.0066428
Restelatto, R., Pavinato, P. S., Sartor, L. R., Einsfeld, S. M., & Baldicera, F. P. (2015). Nitrogen efficiency and nutrient absorption by a sorghum-oats forage succession. Advances in Agriculture. https://doi.org/10.1155/2015/702650
Restelatto, R., Pavinato, P. S., Sartor, L. R., & Paixão, S. J. (2014). Production and nutritional value of sorghum and black oat forages under nitrogen fertilization. Grass and Forage Science. https://doi.org/10.1111/gfs.12076
Santi, A., Amado, T. J. C., & Acosta, J. A. A. (2003). Black oat biomass and nutrient cycling as affected by nitrogen fertilization in soil under no-tillage. Revista Brasileira De Ciência Do Solo. https://doi.org/10.1590/S0100-06832003000600012 in Portuguese with abstract in English.
SBCS/NEPAR—Sociedade Brasileira de Ciência do Solo/ Núcleo do estado do Paraná (2019). Fertilization and liming manual for the state of Paraná. = Manual de adubação e calagem para o estado do Paraná. 2ed. SBCS/NEPAR, Curitiba, PR, Brazil (in Portuguese)
Schenfert, T. A., Ratke, R. F., Casarin, V., Barbosa, J. M., Chaves, D. V., Holanda Neto, M. R. D., Roque, C. G., & Carlos, H. C. V. (2020). Lime and gypsum in the implantation no-till system promote the nutrition and yield of soybean. Journal of Plant Nutrition. https://doi.org/10.1080/01904167.2019.1701028
Schlindwein, J. A., Bortolon, N., Pereira, E. C. F., Bortolon, E. S. O., & Gianello, C. (2013). Phosphorus and potassium fertilization in no till southern Brazilian soils. Agricultural Sciences. https://doi.org/10.4236/as.2013.412A004
Siddiqui, M. H., Al-Whaibi, M. H., Sakran, A. M., Ali, H. M., Basalah, M. O., Faisal, M., Alatar, A., & Al-Amri, A. A. (2013). Calcium-induced amelioration of boron toxicity in radish. Journal of Plant Growth Regulation. https://doi.org/10.1007/s00344-012-9276-6
Silva, M. R., Pelissari, A., Moraes, A. D., Sandini, I. E., Cassol, L. C., Assmann, T. S., & Oliveira, E. B. (2015). Nutrients accumulation and forage production of oat and ryegrass as affected by the application of lime and gypsum on soil surface. Revista De Ciências Agrárias., 38, 346–356. in Portuguese with abstract in English.
Silva, P. R. F., Argenta, G., Sangoi, L., Strieder, M. L., & Silva, A. A. (2006). Management strategies of winter cover crops to maize grown in succession in no-till system. Ciência Rural. https://doi.org/10.1590/S0103-84782006000300049 in Portuguese with abstract in English.
Siqueira-Neto, M., Piccolo, M. D. C., Venzke Filho, S. D. P., Feigl, B. J., & Cerri, C. C. (2010). Mineralization and denitrification of soil nitrogen under no-tillage system. Bragantia. https://doi.org/10.1590/S0006-87052010000400019 in Portuguese with abstract in English.
Somavilla, L., Pinto, M. A. B., Basso, C. J., Da-Ros, C. O., Silva, V. R., Brun, T., & Santi, A. L. (2016). Response of soybean and corn to soil mechanical intervention and agricultural gypsum application to the soil surface. Semina Ciências Agrárias. https://doi.org/10.5433/1679-0359.2016v37n1p95
Soratto, R. P., & Crusciol, C. A. C. (2008a). Chemical soil attributes as affected by lime and phosphogypsum surface application in a recently established no-tillage system. Revista Brasileira De Ciência Do Solo. https://doi.org/10.1590/S0100-06832008000200022 in Portuguese with abstract in English.
Soratto, R. P., & Crusciol, C. A. C. (2008b). Nutrition and grain yield of black oat as affected by surface application of lime and phosphogypsum at the establishment of no-tillage system. Revista Brasileira De Ciência Do Solo. https://doi.org/10.1590/S0100-06832008000200026 in Portuguese with abstract in English.
Soratto, R. P., Crusciol, C. A. C., & de Mello, F. F. C. (2010). Yield components of rice and bean cultivars as affected by lime and phosphogypsum applied in soil surface. Bragantia. https://doi.org/10.1590/S0006-87052010000400023 in Portuguese with abstract in English.
Steiner, F., Fey, R., Zoz, T., & Costa, L. (2009). Biomass production and C/N ration of black oat grown under nitrogen sources and doses. Global Science and Technology., 02, 29–37. in Portuguese with abstract in English.
Thornthwaite, C. W., & Matter, J. R. (1955). The water balance. Publications in climatology. Drexel Institute of Technology.
Torres, J. L. R., Pereira, M. G., & Fabian, A. J. (2008). Cover crops biomass production and its residues mineralization in a Brazilian no-till Oxisol. Pesquisa Agropecuária Brasileira. https://doi.org/10.1590/S0100-204X2008000300018 in Portuguese with abstract in English.
Urano, E. O. M., Kurihara, C. H., Maeda, S., Vitorino, A. C. T., Gonçalves, M. C., & Marchetti, M. E. (2006). Soybean nutritional status evaluation. Pesquisa Agropecuária Brasileira. https://doi.org/10.1590/S0100-204X2006000900011
USDA [United States Department of Agriculture]. (1999). Soil taxonomy: A basic system of soil classifications for making and interpreting soil surveys. US Department of Agriculture.
Vega-García, J. I., López-González, F., Estrada-Flores, J. G., Flores-Calvete, G., Prospero-Bernal, F., & Arriaga-Jordán, C. M. (2020). Black oat (Avena strigosa Schreb.) grazing or silage for small-scale dairy systems in the highlands of central Mexico. Part I. Crop and dairy cow performance. Chilean Journal of Agricultural Research. https://doi.org/10.4067/S0718-58392020000400515
Vicensi, M., Lopes, C., Koszalka, V., Umburanas, R. C., Vidigal, J. C. B., de Ávila, F. W., & Müller, M. M. L. (2020b). Soil fertility, root and aboveground growth of black oat under gypsum and urea rates in no till. Journal of Soil Science and Plant Nutrition. https://doi.org/10.1007/s42729-020-00211-3
Vicensi, M., Lopes, C., Koszalka, V., Umburanas, R. C., Kawakami, J., Pott, C. A., & Müller, M. M. L. (2020a). Gypsum rates and splitting under no-till: soil fertility, corn performance, accumulated yield and profits. Journal of Soil Science and Plant Nutrition. https://doi.org/10.1007/s42729-019-00157-1
Vicensi, M., Müller, M. M. L., Kawakami, J., Nascimento, R., Michalovicz, L., & Lopes, C. (2016). Do rates and splitting of phosphogypsum applications influence the soil and annual crops in a no-tillage system? Revista Brasileira De Ciência Do Solo. https://doi.org/10.1590/18069657rbcs20150155
Viviani, C. A., Marchetti, M. E., Vitorino, A. C. T., Novelino, J. O., & Gonçalves, M. C. (2010). Phosphorus availability in two clayey oxisols and its accumulation in soybean as a function of the increase in pH. Ciência e Agrotecnologia. https://doi.org/10.1590/S1413-70542010000100007 in Portuguese with abstract in English.
Yong, J. W. H., Ng, Y. F., Tan, S. N., & Chew, A. Y. L. (2010). Effect of fertilizer application on photosynthesis and oil yield of Jatropha curcas L. Photosynthetica. https://doi.org/10.1007/s11099-010-0026-3
Zandoná, R. R., Beutler, A. N., Burg, G. M., Barreto, C. F., & Schmidt, M. R. (2015). Gypsum and lime increase soybean and maize yield and decrease drought stress. Pesquisa Agropecuária Tropical, 45, 128–137. in Portuguese with abstract in English.
Zhang, X., Davidson, E. A., Mauzerall, D. L., Searchinger, T. D., Dumas, P., & Shen, Y. (2015). Managing nitrogen for sustainable development. Nature, 528(7580),51–59. https://doi.org/10.1038/nature15743
Ziech, A. R. D., Conceição, P. C., Luchese, A. V., Balin, G. C., & Garmus, T. G. (2015). Soil protection by winter-cycle cover crops in South Brazil. Pesquisa Agropecuária Brasileira. https://doi.org/10.1590/S0100-204X2015000500004 in Portuguese with abstract in English.
Zoca, S. M., & Penn, C. (2017). An important tool with no instruction manual: A review of gypsum use in agriculture. Advances in Agronomy. https://doi.org/10.1016/bs.agron.2017.03.001
Acknowledgements
We acknowledge the Universidade Estadual do Centro-Oeste (Paraná) for supporting this study, the staff of the Departamento de Agronomia and the Laboratório de Solos e Nutrição de Plantas, especially to Luis Henrique de Lima.
Funding
Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (Coordination for the Improvement of Higher Education Personnel, CAPES—Brazil)—Financial Code 001; Conselho Nacional de Desenvolvimento Científico e Tecnológico (National Counsel of Technological and Scientific Development, CNPq—Brazil)—Grant No. 312024/2018-5, and Fundação Araucária (Araucária Foundation from Paraná State—Brazil).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declared that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
Vicensi, M., Umburanas, R.C., da Rocha Loures, F.S. et al. Residual Effect of Gypsum and Nitrogen Rates on Black Oat Regrowth and on Succeeding Soybean under No-Till. Int. J. Plant Prod. 15, 431–445 (2021). https://doi.org/10.1007/s42106-021-00149-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42106-021-00149-1