
Abstract
Age-related changes in intestinal microbiome composition and function are increasingly recognized as pivotal in the pathophysiology of aging and are associated with the aging phenotype. Diet is a major determinant of gut-microbiota composition throughout the entire lifespan, and several of the benefits of a healthy diet in aging could be mediated by the microbiome. Mediterranean diet (MD) is a traditional dietary pattern regarded as the healthy diet paradigm, and a large number of studies have demonstrated its benefits in promoting healthy aging. MD has also a positive modulatory effect on intestinal microbiome, favoring bacterial taxa involved in the synthesis of several bioactive compounds, such as short-chain fatty acids (SCFAs), that counteract inflammation, anabolic resistance, and tissue degeneration. Intervention studies conducted in older populations have suggested that the individual response of older subjects to MD, in terms of reduction of frailty scores and amelioration of cognitive function, is significantly mediated by the gut-microbiota composition and functionality. In this context, the pathophysiology of intestinal microbiome in aging should be considered when designing MD-based interventions tailored to the needs of geriatric patients.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The intestinal microbiome is the ensemble of microorganisms, predominantly bacteria, living in the gastrointestinal lumen and establishing physiologic interactions with the human body, as well as their theater of activity including a whole set of molecules related to gut and host physiology [1]. In healthy adult subjects, an equilibrium between bacterial species with purported health-promoting properties and opportunistic pathogens is generally present, while significant differences in gut microbial communities can be detected among different subjects [1, 2]. This inter-individual variability depends on a large number of factors, including host genetics, environmental exposures, lifestyle and physiologic status [3, 4]. Diet is perhaps the most influential of these factors, as suggested by metagenome-wide association studies [4, 5].
The process of aging implies a certain degree of disruption of the equilibrium between beneficial, neutral and potentially pathogenic bacteria [6], especially after 70 years old, partly as a consequence of aging of the gastrointestinal and immune system [7], and partly as the result of disease, exposure to drugs, and change of diet and mobility [8]. This imbalance of intestinal microbial communities can negatively influence several aspects of the host physiology and is defined as dysbiosis. Although dysbiosis can occur also in earlier stages of the human life, it is consistently involved in the pathogenesis of several age-related diseases and syndromes and thus increasingly regarded as one of the fundamental pathogenetic drivers of aging [9].
Dysbiosis does not only imply a change in the composition of the microbiome, with increased representation of opportunistic pathogens at the expense of taxa with purported health-promoting activity, but it is also associated with a different microbiome functionality, causing changes in the release of physiologically active compounds [10]. In the last decade, several experimental and clinical studies have suggested that age-related gut-microbiota alterations can negatively influence the pathogenesis of many diseases and conditions with high prevalence in geriatric patients [11], including dementia [12], sarcopenia [13], type 2 diabetes [14], hypertension, and other cardiovascular diseases [15].
In older subjects, inter-individual differences in intestinal microbiota composition are also emphasized in comparison with subjects under 70 years of age [16]. As such, fit individuals who reach extreme ages of life, such as centenarians and supercentenarians, may maintain an intestinal microbiome structure more similar to the one of adult subjects, with good representation of bacterial taxa with beneficial modulatory properties for the body functioning, such as anti-inflammatory and pro-anabolic action [17]. Conversely, the deepest levels of dysbiosis are generally observed in frail multimorbid subjects [18, 19].
These circumstances suggest that maintenance of a good equilibrium in intestinal microbiome should be a goal for promoting successful aging [20]. The administration of live bacteria (i.e., probiotics) or functional components (i.e., prebiotics) and foods has shown limited effectiveness on microbiome structure and clinical endpoints in older individuals, although few studies are available to date [21,22,23]. Dietary interventions, instead, have been emphasized as promising strategies to counteract dysbiosis by inducing generalized and durable rearrangements on microbiome composition and function [24]. Mediterranean diet (MD), in particular, has emerged as the healthy diet paradigm and has been associated with a wide range of beneficial effects on primary and secondary prevention of several non-communicable diseases [25, 26].
In this review, we summarize the current evidence on the interactions between MD and gut microbiome, and their importance for mitigating the pathophysiological processes associated with aging and some of the most relevant age-related conditions. First, we consider the effect of MD on the microbiome of adult subjects; then, we explore the impact of MD on older adults; and last, we comment on the main underlying mechanisms behind the anti-aging effects of MD on the gut microbiome.
Association between Mediterranean diet and microbiome in adult subjects
Observational studies
MD, acknowledged by UNESCO as Intangible Cultural Heritage of Humanity in 2010, is a traditional dietary pattern implying daily consumption of plant foods, including cereals, fruit, vegetables and legumes with olive oil as the main source of added fat, and moderate consumption of fish, seafood, eggs, poultry and milk or dairy products. Read meat and unprocessed sugars, which represent important components of modern Western diets, are generally present in limited amounts in MD [27], as well as parsimonious consumption of wine during mealtimes. Apart from its mere composition, MD also implies socialization and conviviality during meals, seasonality and biodiversity in dietary choices, frequent consumption of traditional and local food products and, overall, a healthy lifestyle with a good balance between physical activity and rest [27].
Adherence to MD pattern can be measured in each individual through specific scores validated by the scientific literature. These scores are generally based on the analysis of semi-quantitative food-frequency questionnaires and on the identification of the frequency of consumption of key foods for MD [28]. Despite the limitations of this approach, including the possibility of recall bias, population-based studies conducted in different countries have shown a correlation between MD adherence scores and fecal microbiome composition [29,30,31,32,33,34,35]. The results of these studies are summarized in Table 1.
Interestingly, in most studies, MD adherence was significantly associated neither with biodiversity, that is, species richness, nor with the overall composition of the intestinal microbial community [29, 33, 34]. Instead, MD seemed more associated with increased representation of bacterial taxa with purported health-promoting activities and metabolic function of bacteria, like Lachnospira, Prevotella, bifidobacteria, Faecalibacterium prausnitzii, Eubacterium rectale [29,30,31,32,33,34,35]. Conversely, Ruminococcus, Oscillospira, Escherichia coli and other members of Enterobacteriaceae were the main microbial taxa associated with Western-style diets [32, 34].
At functional level, MD adherence was associated with different metabolic signatures in blood, urine and fecal samples [36]. In particular, it was associated with increased microbial synthesis of short-chain fatty acids (SCFAs), including acetate, propionate and butyrate, which exhibit an overall anti-inflammatory and pro-anabolic function favoring insulin sensitivity [29,30,31, 33]. Bacterial taxa with known capacity of synthetizing SCFAs, such as F. prausnitzii, Butyrivibrio, and Roseburia, seem to play an important role as mediators of the beneficial metabolic effects of MD [37]. High fiber intake, which is a key component of MD, is in fact able to stimulate the representation of these taxa in intestinal microbial communities, while fibers themselves represent the biochemical substrates for SCFA synthesis [38].
MD adherence was also associated with reduced synthesis of trimethylamine-N-oxide (TMAO) [30, 32], an emerging marker of cardiovascular risk synthetized with the contribution of the gut microbiota [39]. According to a study conducted on 307 male adults, Prevotella copri also resulted as a key mediator of the beneficial effects of MD adherence on biomarkers of cardiometabolic risk, including serum levels of C-reactive protein (CRP), total cholesterol, triglycerides, and glycated hemoglobin [33]. Microbiome mediation in the relationship between MD adherence and reduction of inflammation was also confirmed in groups of patients with chronic inflammatory conditions, such as human immunodeficiency virus (HIV) infection [40], or at risk of inflammatory bowel disease (IBD) [34].
Intervention studies
MD intervention is generally assumed to modify gut-microbiome composition and function not only in subjects with chronic diseases (Table 2), but also in healthy subjects [41,42,43]. The magnitude and the characteristics of these changes, however, are inconsistent across studies.
In a group of 20 healthy male volunteers, Barber et al. showed no major differences in gut-microbiome composition before and after a 2-week MD intervention, but they just demonstrated significant variations in a few bacterial taxa [41]. These findings were probably influenced by the high baseline inter-individual variability in the intestinal microbiome of participants, who responded to the dietary intervention in an individualized manner. However, metabolomic analyses revealed a different urinary metabolic profile after the intervention, with significant variations in the levels of several metabolites of bacterial origin [41]. In another study by Godny et al., a 4-week MD intervention associated with daily physical activity measurement resulted into increased representation of bacterial taxa involved in SCFA synthesis, including F. prausnitzii, Lachnospiraceae and Bifidobacterium spp., with reduced levels of inflammatory biomarkers such as fecal calprotectin [42]. Interestingly, similar results were obtained also by Rejeski et al. in a group of 10 healthy subjects, where a 2-week MD intervention also improved the overall microbiome species richness [43].
The modulatory effects of MD on gut microbiome are potentially useful for the prevention and treatment of several chronic conditions, including obesity, type 2 diabetes, and metabolic syndrome [44]. In 18 overweight subjects with body mass index (BMI) ≥ 25 kg/m2, a 3-month dietary intervention consisting of MD enriched with 40 g/day of high-quality extra-virgin olive oil was associated with improved fecal microbiome biodiversity, significant reduction of myeloperoxidase activity, oxidative stress markers and pro-inflammatory cytokines, and significant increase in adiponectin and plasma anti-inflammatory cytokines such as interleukin-10 (IL-10) [45].
Randomized controlled trials (RCTs) investigating the effects of MD and its variants on the gut microbiome of overweight and obese subjects have also shown beneficial effects, ranging from increased biodiversity to increased representation of SCFA-producing taxa or bacteria involved in branched-chain amino acid metabolism (Table 2) [46,47,48,49]. Interestingly, other healthy dietary patterns not falling within the MD definition, such as the vegetarian [48] or the low-fat high-complex carbohydrate (LFHCC) diet [46], also induced beneficial changes in gut-microbiome composition and function, but these changes were distinct from those induced by a traditional MD pattern. LHFCC diet, for example, determined a more pronounced increase in the representation of F. prausnitzii than MD [46], while vegetarian diet promoted growth of Anaerostipes, Streptococcus, Clostridium, and Odoribacter not observed in MD [48].
The effects of MD on the gut microbiome are emphasized by concomitant involvement in a structured physical exercise program. In a group of subjects with non-alcoholic fatty liver disease (NAFLD), a condition frequently overlapped with obesity, the abundance of Ruminococcaceae, Oscillospiraceae and Lachnospiraceae was much more influenced by the association between aerobic physical exercise and MD than MD alone [50].
The interactions between gut-microbiome modifications induced by MD and type 2 diabetes are less known. In subjects following a MD pattern, an intestinal microbiome signature consisting in high abundance of Prevotella, Saccharibacteria, and Betaproteobacteria was associated with increased risk of developing diabetes [51]. Conversely, a 4-week MD intervention in subjects with type 2 diabetes resulted into an improved bacterial richness of gut microbiota, which exhibited a negative correlation with insulin resistance [52]. However, in a recent RCT comparing the effects of MD with a personalized post-prandial targeting (PPT) diet in subjects with type 2 diabetes (Table 2), MD was associated with only minor modifications of fecal microbiome, such as increased abundance of Bifidobacterium adolescentis, and resulted less effective than PPT diet in inducing metabolically-favorable changes of intestinal microbial communities [53]. Nevertheless, it should be highlighted that here the authors compared two different dietary strategies (general vs. personalized dietary advice) and that further interventions with personalized MD intervention could be of interest.
In subjects with metabolic syndrome (Table 2), however, a long-lasting MD intervention was associated with increases in the fecal microbiome representation of Parabacteroides distasonis, Bacteroides thetaiotaomicron, F. prausnitzii, B. adolescentis and Bifidobacterium longum [54]. In two different analyses of the METADIET randomized, controlled, cross-over trial, Galié et al. showed that MD intervention was associated with increased levels of Lachnospiraceae and Ruminococcaceae and improved insulin sensitivity in patients with metabolic syndrome (Table 2) [55, 56].
The effects of MD interventions on the gut microbiome of adult patients with other chronic illnesses not included in the metabolic syndrome spectrum have been scarcely investigated (Table 2). In a RCT comparing MD with the specific carbohydrate diet in adults with Crohn’s disease, neither dietary approach was superior in reducing parameters of inflammation and disease activity after 6 weeks, and no major differences in post-intervention fecal microbiome composition were detected between study arms [57]. Therefore, the current knowledge on the physiologic effects of the interaction between MD and intestinal microbiome in subjects younger than 70 years old is basically limited to improvements in insulin resistance, glucose and lipid metabolism, and chronic subclinical inflammation related to obesity (Table 2).
Mediterranean diet and gut microbiome in older subjects
Observational studies
Despite the gut-microbiome composition of older individuals is less resilient to stressors and more influenced by a large number of (physio)pathologic conditions, diet remains a major driver of the inter-individual variability observed for this age group. In a cross-sectional analysis of data from the NU-AGE RCT, including 226 Dutch subjects aged between 65 and 79, BMI exhibited a much stronger statistically significant association with overall fecal microbiome composition than frailty phenotype [58]. The main dietary factors explaining microbiome inter-individual variability were, on the one side, consumption of fruits, nuts, grain products, and carbohydrates, which are fundamental parts of the MD pattern, and, on the other side, consumption of processed and red meats, which are consumed only occasionally in MD [58]. The main bacterial taxa showing positive associations with foods typically well represented in MD included F. prausnitzii, E. rectale and Eubacterium biforme, while animal protein-based diets were mainly associated with the abundance of taxa with purported pro-inflammatory action, such as Ruminococcus gnavus and Collinsella [58].
Adherence to MD was associated with increased representation of F. prausnitzii also in a group of 74 older Spanish volunteers, where Clostridium cluster XIVa was identified as another important marker of MD [59]. These features of gut microbiome were also associated with specific metabolic signatures, including increased fecal levels of SCFAs, benzoic and 3-hydroxyphenylacetic acids [59, 60]. These signatures are likely the result of the increased-intestinal microbial metabolism of fibers and (poly)phenols associated with MD, respectively [59, 60]. In a group of older Caribbean Latinos living in the United States, instead, adherence to MD was associated with a distinct cluster of fecal microbiome composition, characterized by different abundance of P. copri and co-occurring bacterial networks [61].
In a group of 17 centenarians and 29 nonagenarians from Sardinia (Italy), adherence to MD was associated with several bacterial taxa in fecal microbiome [62]. However, the taxa positively correlated with MD were different between centenarians (Lactobacillus, Clostridium, and Dorea) and nonagenarians (Bacteroides, Parabacteroides, and Pedobacter), with the only exception of Peptoniphilus [62]. Interestingly, MD was associated with depletion of bifidobacteria only in nonagenarians, but not in centenarians [62]. In fact, the microbial ecology of centenarians is characterized by the persistence of a subdominant fraction of bifidobacteria, which is usually depleted in older individuals who do not reach extreme ages [63, 64].
Intervention studies
The existing intervention studies with MD in older individuals have adapted the traditional MD pattern to the specific needs of aging, targeting, in particular, modulation of inflammation, weight loss, and cognition. Berendsen et al. elaborated a modified MD pattern based on recommended daily allowances (RDAs) of macro- and micronutrients for older subjects, with the aim of modulating inflammaging (i.e., the chronic subclinical activation of inflammatory pathways responsible for the pathogenesis of several chronic illnesses and geriatric syndromes typical of the older age) and maintaining a good balance in protein-energy metabolism [65, 66]. This dietary approach —the so-called NU-AGE diet— was the main intervention studied in the NU-AGE RCT, where 1141 pre-frail and fit subjects aged 65 to 79 years old from five European countries were randomized to receive MD tailored to older people (NU-AGE diet) or a control free diet [65]. It was demonstrated as feasible in pre-frail and non-frail older subjects in the long term, with good adherence scores after 1 year from the initiation of the intervention [67]. The gut-microbiota composition and function was among the endpoints of the NU-AGE study and fecal microbiome profiling of 612 participants showed significant changes in the abundance of several taxa [68]. F. prausnitzii, E. rectale, Roseburia, Blautia, Anaerostipes and Prevotella were the main taxa showing positive association with the NU-AGE dietary intervention, while the abundance of Collinsella, Ruminococcus, Dorea, Blautia and Coprococcus exhibited a negative association [68]. Interestingly, after 1-year intervention, those subjects with deeper degrees of gut-microbiome change, reflected in higher levels of the MD microbiome index, also exhibited significant improvement in measures of inflammation (CRP levels), frailty (Fried score and gait speed) and cognition (verbal fluency, Babcok memory score, and constructional praxis score). Conversely, the subjects in the intervention arm who exhibited reduced variations of fecal microbiome composition experienced less pronounced variations in clinical endpoints [68]. Therefore, the NU-AGE diet intervention modulated gut microbiota in a way that reflected negative associations with inflammation and frailty.
A MD-based intervention with energy restriction resulted into significant modulation of fecal microbiome even after only 15 days in a smaller study conducted in 20 obese older women [69]. However, the observed changes were partly different than those of the NU-AGE RCT, with increased representation of Coprococcus, Oscillospira, Bacteroides, and Akkermansia, while Collinsella was confirmed as negatively associated with the dietary intervention [69]. In the PREDIMED-Plus Study, traditional MD intervention was compared with an energy-restricted MD associated with promotion of physical activity in a group of 343 overweight and obese Spanish subjects aged between 55 and 75 years old [70]. Interestingly, the changes observed in gut microbiota after 1-year follow-up were similar between the two groups, but more pronounced in the energy-restricted MD arm, and included increased representation of SCFA producers from the Lachnospiraceae family [70].
The effects of MD intervention approaches on intestinal microbiome have been recently studied also in age-related neurological conditions. Rusch et al. showed significant changes in fecal microbiome composition, including in particular an increase in the SCFA producer Roseburia, after a 5-week MD intervention in a small group of patients with Parkinson’s disease [71]. These changes were associated with improvements in gastrointestinal symptoms (constipation scores), but no neurologic endpoints were measured in the study.
In the field of dementia research, ketogenic diets have shown a potential of slowing down cognitive impairment by improving cerebral metabolism [72]. Recently, some authors have proposed to combine the Mediterranean and ketogenic diet approaches into a modified Mediterranean-ketogenic diet (MMKD) [72], which has shown the capacity of improving body composition and reducing levels of cerebrospinal fluid biomarkers of amyloid deposition (Aβ42) and tau protein [73, 74]. In a randomized cross-over trial conducted on 17 subjects with and without mild cognitive impairment, 6-week MMKD intervention induced increased representation of Akkermansia and Slackia and reduced Bifidobacterium and Lachnobacterium, with the family Lachnospiraceae showing significant correlation with cerebrospinal fluid levels of Aβ42 [75]. Interestingly, changes in symbiont intestinal fungal species, the so-called mycobiome, were also observed [76]. At the functional level, MMKD induced changes in bacterial SCFA synthesis, with significant increases in fecal butyrate and propionate levels [75]. In a recent study conducted in 20 subjects with mild cognitive impairment, MMKD induced lower representation of Alistipes, a bacterial species known for its capacity of producing gamma-amino-butyric acid (GABA), and higher representation of Akkermansia, a taxon with GABA-regulating functionality [77]. GABA imbalance is involved in the gut-brain axis and in the pathogenesis of dementia. Therefore, MD diet could influence important aspects of cognitive function by inducing subtle modifications in gut microbiome.
Anti-Aging effects of mediterranean diet mediated by intestinal microbiome: main underlying mechanisms
Microbial synthesis of short-chain fatty acids (SCFAs)
The depletion of bacterial taxa with capacity to synthetize SCFAs, and particularly F. prausnitzii, Roseburia, Butyrivibrio, and Succinivibrio, is a keynote characteristic of the gut microbiome of older individuals with frailty [78,79,80], sarcopenia [81, 82] and cognitive decline [83]. Frailty is associated with reduced fecal levels of butyrate [80], and an inverse correlation has been demonstrated between gut microbial synthesis of butyrate and appendicular lean mass in subjects at risk for sarcopenia [82]. Conversely, fit centenarians generally exhibit higher fecal levels of SCFAs than subjects between 60 and 70 years old [84].
In this scenario, the capacity of MD to stimulate the growth of SCFA-synthetizing bacteria and improve SCFA levels can be extremely important in an anti-aging perspective. However, the functional capacity of gut bacteria to effectively release SCFAs does not depend only on the fiber content of diet, but it also relies on complex cross-feeding interactions among bacteria and on the interaction between bacteria and host [85, 86]. For example, an adequate butyrate production by F. prausnitzii is possible only in presence of a good representation of bifidobacteria in the gut environment [87, 88].
SCFAs, particularly butyrate, exhibit pleiotropic functions for the host [89]. First, they promote gut mucosal integrity and tropism, representing one of the main sources of energy for colonocytes [89]. Second, they exhibit marked capacity to modulate the inflammatory response, which is pivotal for controlling inflammaging and the pathogenesis of several age-related diseases and conditions, including frailty [90, 91]. In a group of Chinese older patients with cognitive impairment related to diabetes, reduced fecal butyrate levels were associated with higher circulating levels of pro-inflammatory cytokines and worse cognitive performance [92]. In animal models of dementia, butyrate administration is in fact able to improve indices of neuroinflammation and cognitive performance via the gut-brain axis [93, 94].
In systemic circulation, butyrate also improves insulin resistance and has an overall pro-anabolic function, which is pivotal in counteracting type 2 diabetes and obesity [95]. At the skeletal muscle level, these actions result into increased protein synthesis and reduced muscle wasting [96], and this is the main reason why butyrate is considered a powerful anti-sarcopenic mediator in the context of the so-called gut-muscle axis [82, 97]. In fact, butyrate also exhibits an inhibitory capacity at the histone deacetylase level, resulting into increased muscle mass in mouse models of cachexia [98].
Reduction of intestinal mucosa permeability
Aging, even with a healthy active pattern, is associated with increased-intestinal permeability, witnessed by elevated levels of the serum biomarker zonulin [99]. According to a recent meta-analysis of case–control studies investigating biomarkers of frailty, serum zonulin levels are in average higher in frail than in healthy older subjects, reflecting a progressive loss of the barrier function of the intestinal mucosa [100]. This condition is associated with loss of skeletal muscle strength, sarcopenia, and functional autonomy in older subjects, either healthy or with chronic conditions such as chronic obstructive pulmonary disease (COPD) and dementia [101,102,103].
Increased intestinal mucosa permeability is associated with increased serum levels of bacterial toxins, including lipopolysaccharide (LPS), and increased presence of bacterial components into systemic circulation [104, 105]. These compounds provide activation of the innate immune response and antigenic stimulation of adaptive immunity, that ultimately result into persistent subclinical inflammation typical of aging with frailty, the so-called inflammaging [106]. Increased LPS toxinemia plays a pivotal role in the pathophysiology of age-related cognitive decline and Alzheimer’s disease, and is believed to represent one of the mainstays of the gut-brain axis dysregulation [107,108,109]. Age-related gut-microbiome alterations are deeply involved in this pathophysiological cascade, since germ-free mice show no development of chronic inflammation during aging and no increased LPS levels, while old mice with dysbiosis exhibit increased circulating levels of pro-inflammatory cytokines and macrophage dysfunction [110].
Higher adherence to MD is inversely associated with biomarkers of gastrointestinal mucosa permeability and with circulating LPS levels, in both adult subjects with chronic illnesses and older individuals [105, 111, 112]. These effects are emphasized with dietary interventions consisting in increased intake of (poly)phenol-rich foods, which are important components of MD [104, 113, 114]. In particular, the MaPLE randomized controlled trial showed that a (poly)phenol-rich food intervention in older subjects caused reduction in the serum levels of zonulin, associated with favorable changes in the gut microbiome including the increase of the relative abundance of F. prausnitzii [113]. Interestingly, these effects were less pronounced in those subjects with higher disruption of gut microbial community structure and with increased-intestinal permeability at baseline [104, 114]. Dysbiosis, in fact, does not only impair the integrity of the intestinal mucosa, but it can also limit the bioavailability of food bioactives contained in MD, especially (poly)phenolic compounds [115], making dietary interventions probably less effective in subjects with higher burden of frailty and associated gut-microbiome alterations.
The precise interactions among components of MD, gut microbiome, and host cells regulating intestinal mucosa permeability have not been elucidated yet. However, recent studies suggest that SCFAs and the bacterial taxa able to synthetize them from dietary fibers may play a central role [116, 117]. In the LIBRE RCT, for example, the baseline levels of SCFAs were independent predictors of the response to MD intervention in terms of intestinal permeability [117]. Therefore, the individual responses to MD diet may be consistently mediated by the preexisting composition and functionality of gut microbiome.
Biotransformation of food bioactives
(Poly)phenols or phenolic compounds are generally abundant in MD, where the intake of fruit and vegetables, wholegrain cereals, nuts, legumes, and extra virgin olive oil is recommended in large amounts [118]. The interaction between dietary (poly)phenols and the gut microbiome is able to generate several bioactive metabolites exhibiting anti-aging effects, especially at the skeletal muscle and central nervous system level [86, 119, 120]. However, the microbiome response to dietary interventions shows a consistent inter-individual variability in terms of production of bioactive compounds [121, 122]. In the case of some polyphenolic subclasses, including ellagitannins, flavanones, isoflavones, flavan-3-ols, prenylflavonoids, avenanthramides, resveratrol, and lignans, some microbiota-related metabotypes have been identified, so that the beneficial effects of these dietary components might be observed only in presence of a particular microbiome composition and functionality [121, 122].
For example, urolithin A, isourolithin A, and urolithin B are metabolites released by the gut microbiome after ingestion of ellagic acid and ellagitannins, polyphenols frequently found in walnuts, pomegranate, and strawberries. Not all the individuals are able to produce these metabolites, and subjects can be classified into the Uro-A or Uro-B metabotype according to their capability to not produce or produce, respectively, urolithin B or isourolithin A, in addition to urolithin A [123]. Conversely, subjects with the Uro-0 metabotype do not show production of (iso)urolithins A or B by the gut microbiome, even after a dietary challenge with foods with high ellagitannin content, and cannot benefit from the biologic functions of urolithins [123]. While isourolithin A and urolithin B have been associated to gut dysbiosis and the prevalence of the Uro-B metabotype increases with aging, urolithin A production and the Uro-A metabotype has been related to a healthier and younger profile [122]. The potential anti-aging effects of urolithin A include improvement of muscle strength and exercise endurance [124, 125], modulation of neuroinflammation and cell apoptosis with improvement in cognitive function [126,127,128], promotion of insulin sensitivity, modulation of lipid metabolism and inflammatory response [129]. Interestingly, in a recent RCT testing the effects of MD in obese subjects, MD was associated with an average increase in the urinary excretion of urolithins, even if the analyses did not consider metabotypes [47]. Similarly, urolithin urinary excretion was significantly associated with visceral adiposity reduction [130] and magnetic resonance-measured hippocampal occupancy score [131] in two distinct RCTs testing the effects of a long MD intervention.
Gut microbiota-derived metabotypes are less known for polyphenol subclasses other than ellagitannins. Hesperidin high-excretors and low-excretors have been identified after dietary intake of flavanones, a polyphenol subclass particularly represented in citrus [121, 122]. Hesperidin exhibits antioxidant, anti-inflammatory and pro-anabolic actions, promoting muscle protein synthesis [132, 133] and reducing amyloid deposition and neuroinflammation in animal models of Alzheimer’s disease [134, 135]. In an intervention study testing the effects of MD in subjects with type 2 diabetes, increased plasma levels of hesperidin and other flavanone derivatives were detected after 12 weeks, with significant reductions in inflammatory biomarkers [136]. Similarly, equol is a bioactive compound released after intestinal biotransformation of soy isoflavone daidzein, but it is produced only by a part of the population harboring a specific microbial profile. Equol exhibited neuroprotective actions against the onset of dementia in vitro [137,138,139], but it was associated in vivo with better cognitive performance only in presence of an equol-producer microbiome metabotype [140].
The role of dietary proteins and exercise in older age: a gut microbiome perspective
Nutritional guidelines and clinical recommendations against age-related physical frailty and sarcopenia generally emphasize the importance of increasing protein intake to overcome anabolic resistance [141, 142]. However, intervention studies have shown only mild improvements in muscle mass and strength after increases of daily protein intake up to 1.6 g/kg/day [143]. High-protein diets, especially rich of processed foods of animal origin, are also associated with deleterious consequences for the gut-microbiome composition and functionality in both human and animal experiments [144]. A recent systematic review of RCTs conducted in human beings has shown that higher meat intake is associated with marked reduction of Anaerostipes and Faecalibacterium, two of the main taxa producing SCFAs [145]. In this perspective, ad libitum consumption of animal proteins in the older age may produce conflicting effects on the pathophysiological mechanisms of physical frailty and sarcopenia, resulting in modest clinical benefits [146, 147].
The benefits of high-protein diets on frailty and inflammation, instead, may be more pronounced when the dietary intervention consists in consumption of proteins with high biologic value, such as whey protein [148, 149], or when dietary intervention is associated with regular exercise [150]. In a double-blind placebo-controlled cross-over study, Ford and colleagues demonstrated that a balanced high-protein diet, either in combination with probiotics/synbiotics or alone, was associated with generally favorable changes in the gut-microbiota composition of a group of healthy older women, although the representation of the SCFA producers Roseburia and Anaerostipes was reduced [151]. In healthy young subjects, the increase of lean red meat consumption under controlled conditions was not associated with detrimental consequences for the microbiome structure as well [152].
Protein consumption in MD is balanced and mainly includes vegetal proteins of high biologic value, such as those from legumes and lean meat, rather than proteins from processed red meats. Thus, increasing protein intake maintaining a Mediterranean dietary style may represent an optimal compromise from a gut-microbiota perspective [146, 147].
Furthermore, regular exercise seems to act as an enhancer of the beneficial effects of MD-style diets, both on the gut-microbiome composition and on clinical markers of aging. Multicomponent interventions consisting in regular exercise and tailored nutritional counseling have proven effective in reducing the burden of disability in physically frail older individuals [153, 154]. Exercise represents a powerful beneficial modulator of gut microbiome and can prevent dysbiosis in both adult and older age [155, 156]. However, no comprehensive investigation of the effects of exercise and its interaction with dietary patterns has been conducted in older individuals to date. Interestingly, in the PREDIMED-Plus Study the effects of energy-restricted MD dietary interventions on the gut microbiome of subjects aged 55–75 years old were more pronounced when diet was associated with regular exercise programs [70]. Therefore, multicomponent interventions combining MD or its variants with exercise programs should represent the best alternatives to counteract age-related frailty also from a gut microbiome perspective. Future studies should include also gut-microbiome parameters as endpoints of clinical interventions against frailty.
Conclusion and perspectives
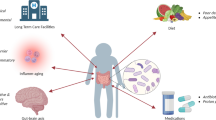
A healthy diet can influence several pathophysiological aspects of aging through mediation of the intestinal microbiome, while Western-style diets may be associated with a tendency toward dysbiosis favoring the pathophysiological processes leading to frailty (Fig. 1). In this context, promoting MD in older individuals can represent an effective strategy to counteract the age-related drift of gut-microbiome composition and function toward dysbiosis and its detrimental consequences. In both adult subjects and older individuals, adherence to the MD pattern is associated with maintenance of a healthy microbiome able to modulate inflammation, anabolic resistance, oxidative stress, and neurodegeneration in a favorable way. Intervention studies have confirmed this interaction among diet, gut microbiome, and host (patho)physiology, but few studies have specifically targeted older individuals and outcomes of geriatric interest. In this regard, the results from the NU-AGE study, suggesting that older individuals respond to MD intervention in an individualized manner by mediation of the intestinal microbiome, are of paramount importance to understand the complex underlying mechanisms linking diet, aging, and its phenotype. Future studies should further investigate the role of MD and its variants in counteracting physical and cognitive decline in the older age, accounting also for the role of the microbiome from a multi-omics perspective.

Overview of the mechanisms linking dietary habits, and particularly Mediterranean diet, to gut-microbiome and pathophysiological aspects of aging (created with Biorender.com)
Data availability
Data sharing not applicable to this article as no datasets were generated or analyzed in the present work.
References
Berg G, Rybakova D, Fischer D et al (2020) Microbiome definition re-visited: old concepts and new challenges. Microbiome 8:103. https://doi.org/10.1186/s40168-020-00875-0
Fassarella M, Blaak EE, Penders J et al (2021) Gut microbiome stability and resilience: elucidating the response to perturbations in order to modulate gut health. Gut 70:595–605. https://doi.org/10.1136/gutjnl-2020-321747
Kurilshikov A, Medina-Gomez C, Bacigalupe R et al (2021) Large-scale association analyses identify host factors influencing human gut microbiome composition. Nat Genet 53:156–165. https://doi.org/10.1038/s41588-020-00763-1
Zhernakova A, Kurilshikov A, Bonder MJ et al (2016) Population-based metagenomics analysis reveals markers for gut microbiome composition and diversity. Science 352:565–569. https://doi.org/10.1126/science.aad3369
Rotschild D, Weissbrod O, Barkan E et al (2018) Environment dominates over host genetics in shaping human gut microbiota. Nature 555:210–215. https://doi.org/10.1038/nature25973
Ling Z, Liu X, Cheng Y et al (2022) Gut microbiota and aging. Crit Rev Food Sci Nutr 62:3509–3534. https://doi.org/10.1080/10408398.2020.1867054
Hohman LS, Osborne LC (2022) A gut-centric view of aging: do intestinal epithelial cells contribute to age-associated microbiota changes, inflammaging, and immunosenescence? Aging Cell 21:e13700. https://doi.org/10.1111/acel.13700
Strasser B, Wolters M, Weyh C et al (2021) The effects of lifestyle and diet on gut microbiota composition, inflammation, and muscle performance in our ageing society. Nutrients 13:2045. https://doi.org/10.3390/nu13062045
López-Otín C, Blasco MA, Partridge L et al (2023) Hallmarks of aging: an expanding universe. Cell 186:243–268. https://doi.org/10.1016/j.cell.2022.11.001
Haran JP, McCormick BA (2021) Aging, frailty, and the microbiome-How dysbiosis influences human aging and disease. Gastroenterology 160:507–523. https://doi.org/10.1053/j.gastro.2020.09.060
Strasser B, Ticinesi A (2023) Intestinal microbiome in normal ageing, frailty and cognition decline. Curr Opin Clin Nutr Metab Care 26:8–16. https://doi.org/10.1097/MCO.0000000000000878
Ticinesi A, Tana C, Nouvenne A et al (2018) Gut microbiota, cognitive frailty and dementia in older individuals: a systematic review. Clin Interv Aging 13:1497–1511. https://doi.org/10.2147/CIA.S139163
Liu C, Cheung WH, Li J et al (2021) Understanding the gut microbiota and sarcopenia: a systematic review. J Cachexia Sarcopenia Muscle 12:1393–1407. https://doi.org/10.1002/jcsm.12784
Gurung M, Li Z, You H et al (2020) Role of gut microbiota in type 2 diabetes pathophysiology. EBioMedicine 51:102590. https://doi.org/10.1016/j.ebiom.2019.11.051
Nesci A, Carnuccio C, Ruggieri V et al (2023) Gut microbiota and cardiovascular disease: evidence on the metabolic and inflammatory background of a complex relationship. Int J Mol Sci 24:9087. https://doi.org/10.3390/ijms24109087
Ghosh TS, Shanhan F, O’Toole PW (2022) Toward an improved definition of a healthy microbiome for healthy aging. Nat Aging 2:1054–1059. https://doi.org/10.1038/s43587-022-00306-9
Santoro A, Ostan R, Candela M et al (2018) Gut microbiota changes in the extreme decades of human life: a focus on centenarians. Cell Mol Life Sci 75:129–148. https://doi.org/10.1007/s00018-017-2674-y
Zhao L, Zhang Y, Wang Y et al (2023) Gut microbiota diversity of hospitalized older adult patients with and without antibiotic-associated diarrhea. Aging Clin Exp Res 35:1541–1555. https://doi.org/10.1007/s40520-023-02436-5
Milani C, Ticinesi A, Gerritsen J et al (2016) Gut microbiota composition and Clostridium difficile infection in hospitalized elderly individuals: a metagenomic study. Sci Rep 6:25945. https://doi.org/10.1038/srep25945
Vaiserman AM, Koliada AK, Marotta F (2017) Gut microbiota: a player in aging and a target for anti-aging intervention. Ageing Res Rev 35:36–45. https://doi.org/10.1016/j.arr.2017.01.001
Prokopidis K, Giannos P, Kirwan R et al (2023) Impact of probiotics on muscle mass, muscle strength and lean mass: a systematic review and meta-analysis of randomized controlled trials. J Cachexia Sarcopenia Muscle 14:30–44. https://doi.org/10.1002/jcsm.13132
Krüger JF, Hillesheim E, Pereira ACSN et al (2021) Probiotics for dementia: a systematic review and meta-analysis of randomized controlled trials. Nutr Rev 79:160–170. https://doi.org/10.1093/nutrit/nuaa037
de la Sánchez y Sánchez BB, Martínez Carrillo BE, Aguirre Garrido JF et al (2022) Emerging evidence on the use of probiotics and prebiotics to improve the gut microbiota of older adults with frailty syndrome: a narrative review. J Nutr Health Aging 26:926–935. https://doi.org/10.1007/s12603-022-1842-4
Wolter M, Grant ET, Boudaud M et al (2021) Leveraging diet to engineer the gut microbiome. Nat Rev Gatroenterol Hepatol 18:885–902. https://doi.org/10.1038/s41575-021-00512-7
Sofi F, Cesari F, Abbate R et al (2008) Adherence to Mediterranean diet and health status: meta-analysis. BMJ 337:a1344. https://doi.org/10.1136/bmj.a1344
Mazza E, Ferro Y, Pujia R et al (2021) Mediterranean diet in healthy aging. J Nutr Health Aging 25:1076–1083. https://doi.org/10.1007/s12603-021-1675-6
Bach-Faig A, Berry EM, Lairon D et al (2011) Mediterranean diet pyramid today. Science and cultural updates. Public Health Nutr 14:2274–2284. https://doi.org/10.1017/S1368980011002515
Zaragoza-Martí A, Cabañero-Martínez MJ, Hurtado-Sánchez JA et al (2018) Evaluation of Mediterranean diet adherence scores: a systematic review. BMJ Open 8:e019033. https://doi.org/10.1136/bmjopen-2017-019033
Gutiérrez-Díaz I, Fernández-Navarro T, Sánchez B et al (2016) Mediterranean diet and faecal microbiota: a transversal study. Food Funct 7:2347–2356. https://doi.org/10.1039/c6fo00105j
De Filippis F, Pellegrini N, Vannini L et al (2016) High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut 65:1812–1821. https://doi.org/10.1136/gutjnl-2015-309957
Mitsou EK, Kakali A, Antonopoulou S et al (2017) Adherence to the Mediterranean diet is associated with the gut microbiota pattern and gastrointestinal characteristics in an adult population. Br J Nutr 117:1645–1655. https://doi.org/10.1017/S0007114517001593
Gallé F, Valeriani F, Cattaruzza MS et al (2020) Mediterranean diet, physical activity and gut microbiome composition: a cross-sectional study among healthy young Italian adults. Nutrients 12:2164. https://doi.org/10.3390/nu12072164
Wang DD, Nguyen LH, Li Y et al (2021) The gut microbiome modulates the protective association between a Mediterranean diet and cardiometabolic disease risk. Nat Med 27:333–343. https://doi.org/10.1038/s41591-020-01223-3
Turpin W, Dong M, Sasson G et al (2022) Mediterranean-like pattern associations with gut microbiome composition and subclinical gastrointestinal inflammation. Gastroenterology 163:685–698. https://doi.org/10.1053/j.gastro.2022.05.037
Rosés C, Cuevas-Sierra A, Quintana S et al (2021) Gut microbiota bacterial species associated with Mediterranean diet-related food groups in a Northern Spanish population. Nutrients 13:636. https://doi.org/10.3390/nu13020636
Almanza-Aguilera E, Urpi-Sarda M, Llorach R et al (2017) Microbial metabolites are associated with high adherence to a Mediterranean diet pattern using a 1H-NMR-based untargeted metabolomics approach. J Nutr Biochem 48:36–43. https://doi.org/10.1016/j.jnutbio.2017.06.001
Vitale M, Giacco R, Laiola M et al (2021) Acute and chronic improvement in postprandial glucose metabolism by a diet resembling the traditional Mediterranean dietary pattern: Can SCFAs play a role? Clin Nutr 40:428–437. https://doi.org/10.1016/j.clnu.2020.05.025
Oliver A, Chase AB, Weihe C et al (2021) High-fiber, whole-food dietary intervention alters the human gut microbiome but not fecal short-chain fatty acids. mSystems 6:e00115-e121. https://doi.org/10.1128/mSystems.00115-21
Tang WHW, Wang Z, Levison BS et al (2013) Intestinal microbial metabolism of phosphatidylcholine and cardiovascular risk. N Engl J Med 368:1575–1784. https://doi.org/10.1056/NEJMoa1109400
Manzano M, Talavera-Rodríguez A, Moreno E et al (2022) Relationship of diet to gut microbiota and inflammatory biomarkers in people with HIV. Nutrients 14:1221. https://doi.org/10.3390/nu14061221
Barber C, Mego M, Sabater C et al (2021) Differential effects of Western and Mediterranean-type diets on gut microbiota: a metagenomics and metabolomics approach. Nutrients 13:2638. https://doi.org/10.3390/nu13082638
Godny L, Reshef L, Sharar Fischler T et al (2022) Increasing adherence to the Mediterranean diet and lifestyle is associated with reduced fecal calprotectin and intra-individual changes in microbial composition of healthy subjects. Gut Microbes 14:e2120749. https://doi.org/10.1080/19490976.2022.2120749
Rejeski JJ, Wilson FM, Nagpal R et al (2022) The impact of a Mediterranean diet on the gut microbiome in healthy human subjects: a pilot study. Digestion 103:133–140. https://doi.org/10.1159/000519445
Dabke K, Hendrick G, Devkota S (2019) The gut microbiome and metabolic syndrome. J Clin Invest 129:4050–4057. https://doi.org/10.1172/JCI129194
Luisi MLE, Lucarini L, Biffi B et al (2019) Effect of Mediterranean diet enriched in high quality extra virgin olive oil on oxidative stress, inflammation and gut microbiota in obese and normal weight adult subjects. Front Pharmacol 10:1366. https://doi.org/10.3389/fphar.2019.01366
Haro C, Montes-Borrego M, Rangel-Zúñiga OA et al (2016) Two healthy diets modulate gut microbial community improving insulin sensitivity in a human obese population. J Clin Endocrinol Metab 101:233–242. https://doi.org/10.1210/jc.2015-3351
Meslier V, Laiola M, Roager HM et al (2020) Mediterranean diet intervention in overweight and obese subjects lowers plasma cholesterol and causes changes in the gut microbiome and metabolome independently of energy intake. Gut 69:1258–1268. https://doi.org/10.1136/gutjnl-2019-320438
Pagliai G, Russo E, Niccolai E et al (2020) Influence of a 3-month low-calorie Mediterranean diet compared to the vegetarian diet on human gut microbiota and SCFA: the CARDIVEG Study. Eur J Nutr 59:2011–2024. https://doi.org/10.1007/s00394-019-02050-0
Rinott E, Meir EY, Tsaban G et al (2022) The effects of the Green-Mediterranean diet on cardiometabolic health are linked to gut microbiome modifications: a randomized controlled trial. Genome Med 14:29. https://doi.org/10.1186/s13073-022-01015-z
Calabrese FM, Disciglio V, Franco I et al (2022) A low glycemic index Mediterranean diet combined with aerobic physical activity rearranges the gut microbiota signature in NAFLD patients. Nutrients 14:1773. https://doi.org/10.3390/nu1409177
Camargo A, Vals-Delgado C, Alcala-Diaz JF et al (2020) A diet-dependent microbiota profile associated with incident type 2 diabetes: from the CORDIOPREV Study. Mol Nutr Food Res 64:2000730. https://doi.org/10.1002/mnfr.202000730
Ismael S, Silvestre MP, Vasques M et al (2021) A pilot study on the metabolic impact of a Mediterranean diet in type 2 diabetes: is gut microbiota the key? Nutrients 13:1228. https://doi.org/10.3390/nu13041228
Ben-Yacov O, Godneva A, Rein M et al (2023) Gut microbiome modulates the effects of a personalised postprandial-targeting (PPT) diet on cardiometabolic markers: a diet intervention in pre-diabetes. Gut 72:1486–1496. https://doi.org/10.1136/gutjnl-2022-329201
Haro C, Garcia-Carpintero S, Alcala-Diaz JF et al (2016) The gut microbial community in metabolic syndrome is modified by diet. J Nutr Biochem 27:27–31. https://doi.org/10.1016/j.jnutbio.2015.08.011
Galié S, García-Gavilán J, Camacho-Barcía L et al (2021) Effects of the Mediterranean diet or nut consumption on gut microbiota composition and fecal metabolites and their relationship with cardiometabolic risk factors. Mol Nutr Food Res 65:2000982. https://doi.org/10.1002/mnfr.202000982
Galié S, García-Gavilán J, Papandreou C et al (2021) Effects of Mediterranean diet on plasma metabolites and their relationship with insulin resistance and gut microbiota in a crossover randomized clinical trial. Clin Nutr 40:3798–3806. https://doi.org/10.1016/j.clnu.2021.04.028
Lewis JD, Sandler RS, Brotherton C et al (2021) A randomized trial comparing the specific carbohydrate diet to a Mediterranean diet in adults with Chron’s disease. Gastroenterology 161:837–852. https://doi.org/10.1053/j.gastro.2021.05.047
Van Soest APM, Hermes GDA, Berendsen AM et al (2020) Associations between pro- and anti-inflammatory gastro-intestinal microbiota, diet, and cognitive functioning in Dutch healthy older adults: the NU-AGE study. Nutrients 12:3471. https://doi.org/10.3390/nu12113471
Gutiérrez-Díaz I, Fernández-Navarro T, Salazar N et al (2017) Adherence to a Mediterranean diet influences the fecal metabolic profile of microbial-derived phenolics in a Spanish cohort of middle-age and older people. J Agric Food Chem 65:586–595. https://doi.org/10.1021/acs.jafc.6b04408
Ruiz-Saavedra S, Salazar N, Suárez A et al (2020) Comparison of different dietary indices as predictors of inflammation, oxidative stress and intestinal microbiota in middle-aged and elderly subjects. Nutrients 12:3828. https://doi.org/10.3390/nu12123828
Maldonado-Contreras A, Noel SE, Ward DV et al (2020) Associations between diet, the gut microbiome, and short-chain fatty acid production among older Caribbean Latino adults. J Acad Nutr Diet 120:2047–2060. https://doi.org/10.1016/j.jand.2020.04.018
Palmas V, Pisanu S, Madau V et al (2022) Gut microbiota markers and dietary habits associated with extreme longevity in healthy Sardinian centenarians. Nutrients 14:2436. https://doi.org/10.3390/nu14122436
Biagi E, Franceschi C, Rampelli S et al (2016) Gut microbiota and extreme longevity. Curr Biol 26:1480–1485. https://doi.org/10.1016/j.cub.2016.04.016
Wu L, Zeng T, Zinellu A et al (2019) A Cross-sectional study of compositional and functional profiles of gut microbiota in Sardinian centenarians. mSystems 4:e00325-19. https://doi.org/10.1128/mSystems.00325-19
Berendsen A, Santoro A, Pini E et al (2013) A parallel randomized trial on the effect of a heathful diet on inflammageing and its consequences in European elderly people: design of the NU-AGE dietary intervention study. Mech Ageing Dev 134:523–530. https://doi.org/10.1016/j.mad.2013.10.002
Santoro A, Pini E, Scurti M et al (2014) Comating inflammaging through a Mediterranean whole diet approach: the NU-AGE project’s conceptual framework and design. Mech Ageing Dev 136–137:3–13. https://doi.org/10.1016/j.mad.2013.12.001
Berendsen AAM, van de Rest O, Feskens EJM et al (2018) Changes in dietary intake and adherence to the NU-AGE diet following a one-year dietary intervention among European older adults-results of the NU-AGE randomized trial. Nutrients 10:1905. https://doi.org/10.3390/nu10121905
Ghosh TS, Rampelli S, Jeffery IB et al (2020) Mediterranean diet intervention alters the gut microbiome in older people reducing frailty and improving health status: the NU-AGE 1-year dietary intervention across five European countries. Gut 69:1218–1228. https://doi.org/10.1136/gutjnl-2019-319654
Cancello R, Turroni S, Rampelli S et al (2019) Effect of short-term dietary intervention and probiotic mix supplementation on the gut microbiota of elderly obese women. Nutrients 11:3011. https://doi.org/10.3390/nu11123011
Muralidharan J, Moreno-Indias I, Bulló M et al (2021) Effects on gut microbiota of a 1-y lifestyle intervention with Mediterranean diet compared with energy-reduced Mediterranean diet and physical activity promotion: the PREDIMED-plus study. Am J Clin Nutr 114:1148–1158. https://doi.org/10.1093/ajcn/nqab150
Rusch C, Neke M, Tucciarone L et al (2021) Mediterranean diet adherence in people with Parkinson’s disease reduces constipation symptoms and changes fecal microbiota after a 5-week single-arm pilot study. Front Neurol 12:794640. https://doi.org/10.3389/fneur.2021.794640
Ellouze I, Sheffler J, Nagpal R et al (2023) Dietary patterns and Alzheimer’s disease: an updated review linking nutrition to neuroscience. Nutrients 15:3204. https://doi.org/10.3390/nu15143204
Neth BJ, Mintz A, Whitlow C et al (2020) Modified ketogenic diet is associated with improved cerebrospinal fluid biomarker profile, cerebral perfusion, and cerebral ketone body uptake in older adults at risk for Alzheimer’s disease: a pilot study. Neurobiol Aging 86:54–63. https://doi.org/10.1016/j.neurobiolaging.2019.09.015
Brinkley TE, Leng I, Register TC et al (2022) Changes in adiposity and cerebrospinal fluid biomarkers following a modified mediterranean ketogenic diet in older adults at risk for Alzheimer’s disease. Front Neurosci 16:906539. https://doi.org/10.3389/fnins.2022.906539
Nagpal R, Neth RJ, Wang S et al (2019) Modified Mediterranean-ketogenic diet modulates gut microbiome and short-chain fatty acids in association with Alzheimer’s disease markers in subjects with mild cognitive impairment. EBioMedicine 47:529–542. https://doi.org/10.1016/j.ebiom.2019.08.032
Nagpal R, Neth BJ, Wang S et al (2020) Gut mycobiome and its interaction with diet, gut bacteria and Alzheimer’s disease markers in subjects with mild cognitive impairment: a pilot study. EBioMedicine 59:102950. https://doi.org/10.1016/j.ebiom.2020.102950
Dilmore AH, Martino C, Neth BJ et al (2023) Effect of a ketogenic and low-fat diet on the human metabolome, microbiome, and foodome in adults at risk for Alzheimer’s disease. Alzheimers Dement. https://doi.org/10.1002/alz.13007
Jackson MA, Jeffery IB, Beaumont M et al (2016) Signatures of early frailty in the gut microbiota. Genome Med 8:8. https://doi.org/10.1186/s13073-016-0262-7
Xu Y, Wang Y, Li H et al (2021) Altered fecal microbiota composition in older adults with frailty. Front Cell Infect Microbiol 11:696186. https://doi.org/10.3389/fcimb.2021.696186
Guo Y, Zhu F, Wang F et al (2022) Distinct serum and fecal metabolite profiles linking with gut microbiome in older adults with frailty. Front Med 9:827174. https://doi.org/10.3389/fmed.2022.827174
Ticinesi A, Mancabelli L, Tagliaferri S et al (2020) The gut-muscle axis in older subjects with low muscle mass and performance: a proof of concept study exploring fecal microbiota composition and function with shotgun metagenomics sequencing. Int J Mol Sci 21:8946. https://doi.org/10.3390/ijms21238946
Lv WQ, Lin X, Shen H et al (2021) Human gut microbiome impacts skeletal muscle mass via gut microbial synthesis of the short-chain fatty acid butyrate among healthy postmenopausal women. J Cachexia Sarcopenia Muscle 12:1860–1870. https://doi.org/10.1002/jcsm.12788
Cattaneo A, Cattane A, Galluzzi S et al (2017) Association of brain amyloidosis with pro-inflammatory gut bacterial taxa and peripheral inflammation markers in cognitively impaired elderly. Neurobiol Aging 49:60–68. https://doi.org/10.1016/j.neurobiolaging.2016.08.019
Cai D, Zhao Z, Zhao L et al (2022) The age-accompanied and diet-associated remodeling of the phospholipid, amino acid, and SCFA metabolism of healthy centenarians from a Chinese longevous region: a window into exceptional longevity. Nutrients 14:4420. https://doi.org/10.3390/nu14204420
Zhang M, Li RW, Yang H et al (2022) Recent advances in developing butyrogenic foods to promote gut health. Crit Rev Food Sci Nutr. https://doi.org/10.1080/10408398.2022.2142194
Ticinesi A, Guerra A, Nouvenne A et al (2023) Disentangling the complexity of nutrition, frailty and gut microbial pathways during aging: a focus on hippuric acid. Nutrients 15:1138. https://doi.org/10.3390/nu15051138
Rios-Covian D, Gueimonde M, Duncan SH et al (2015) Enhanced butyrate formation by cross-feeding between Faecalibacterium prausnitzii and Bifidobacterium adolescentis. FEMS Microbiol Lett 362:fnv176. https://doi.org/10.1093/femsle/fnv176
Kim H, Yeong Y, Kang S et al (2020) Co-culture with bifidobacterium catenulatum improves the growth, gut colonization, and butyrate production of Faecalibacterium prausnitzii: in vitro and in vivo studies. Microorganisms 8:788. https://doi.org/10.3390/microorganisms8050788
Blaak EE, Canfora EE, Theis S et al (2020) Short chain fatty acids in human gut and metabolic health. Benef Microbes 11:411–455. https://doi.org/10.3920/BM2020.0057
Hu T, Wu Q, Yao Q et al (2022) Short-chain fatty acid metabolism and multiple effects on cardiovascular diseases. Ageing Res Rev 81:101706. https://doi.org/10.1016/j.arr.2022.101706
Hildebrand CB, Lichatz R, Pich A et al (2023) Short-chain fatty acids improve inflamm-aging and acute lung injury in old mice. Am J Physiol Cell Mol Physiol 324:L480–L492. https://doi.org/10.1152/ajplung.00296.2022
Du Y, Li X, An Y et al (2022) Association of gut microbiota with short-chain fatty acids and inflammatory cytokines in diabetic patients with cognitive impairment: a cross-sectional, non-controlled study. Front Nutr 9:930626. https://doi.org/10.3389/fnut.2022.930626
Matt SM, Allen JM, Lawson MA et al (2018) Butyrate and dietary soluble fiber improve neuroinflammation associated with aging in mice. Front Immunol 9:1832. https://doi.org/10.3389/fimmu.2018.01832
Zhou Y, Xie L, Schröder J et al (2023) Dietary fiber and microbiota metabolite receptors enhance cognition and alleviate disease in the 5xFAD mouse model of Alzheimer’s disease. J Neurosci 43:6460–6475. https://doi.org/10.1523/JNEUROSCI.0724-23.2023
Canfora EE, Jocken JW, Blaak EE (2015) Short-chain fatty acids in control of body weight and insulin sensitivity. Nat Rev Endocrinol 11:577–591. https://doi.org/10.1038/nrendo.2015.128
Prokopidis K, Chambers E, Ni Lochlainn M et al (2021) Mechanisms linking the gut-muscle axis with muscle protein metabolism and anabolic resistance: implications for older adults at risk of sarcopenia. Front Physiol 12:770455. https://doi.org/10.3389/fphys.2021.770455
Ticinesi A, Lauretani F, Milani C et al (2017) Aging Gut microbiota at the cross-road between nutrition, physical frailty, and sarcopenia: is there a gut-muscle axis? Nutrients 9:1303. https://doi.org/10.3390/nu9121303
Walsch ME, Bhattacharya A, Sataranatarajan K et al (2015) The histone deacetylase inhibitor butyrate improves metabolism and reduces muscle atrophy during aging. Aging Cell 14:957–970. https://doi.org/10.1111/acel.12387
Qi Y, Goel R, Kim S et al (2017) Intestinal permeabolity biomarker zonulin is elevated in healthy aging. J Am Med Dir Assoc 18:810.e1-810.e4. https://doi.org/10.1016/j.jamda.2017.05.018
Rashidah NH, Lim SM, Neoh CF et al (2022) Differential gut microbiota and intestinal permeability between frail and healthy older adults: a systematic review. Ageing Res Rev 82:101744. https://doi.org/10.1016/j.arr.2022.101744
Li C, Li Y, Wang N et al (2022) Intestinal permeability associated with the loss of skeletal muscle strength in middle-aged and older adults in rural area of Beijing. China Healthcare 10:1100. https://doi.org/10.3390/healthcare10061100
Karim A, Muhammad T, Ustrana S et al (2021) Intestinal permeability marker zonulin as a predictor of sarcopenia in chronic obstructive pulmonary disease. Respir Med 189:106662. https://doi.org/10.1016/j.rmed.2021.106662
Karim A, Iqbal MS, Muhammad T et al (2022) Elevated plasma zonulin and CAF22 are correlated with sarcopenia and functional dependency at various stages of Alzheimer’s disease. Neurosci Res 184:47–53. https://doi.org/10.1016/j.neures.2022.08.004
Gargari G, Taverniti V, Del Bò C et al (2022) Higher bacterial DNAemia can affest the impact of a polyphenol-rich dietary pattern on biomarkers of intestinal permeability and cardiovascular risk in older subjects. Eur J Nutr 61:1209–1220. https://doi.org/10.1007/s00394-021-02680-3
Pastori D, Carnevale R, Nocella C et al (2017) Gut-derived serum lipopolysaccharide is associated with enhanced risk of major adverse cardiovascular events in atrial fibrillation: effect of adherence to Mediterranean diet. J Am Heart Assoc 6:e005784. https://doi.org/10.1161/JAHA.117.005784
Di Giosia P, Stamerra CA, Giorgini P et al (2022) The role of nutrition in inflammaging. Ageing Res Rev 77:101596. https://doi.org/10.1016/j.arr.2022.101596
Qu L, Li Y, Liu F et al (2023) Microbiota-gut-brain axis dysregulation in Alzheimer’s disease: multi-pathway effects and therapeutical potential. Aging Dis. https://doi.org/10.14336/AD.2023.0823-2
Inaba T, Yamashiro K, Kurita N et al (2023) Microbial lipopolysaccharide-induced inflammation contributes to cognitive impairment and white matter lesion progression in diet-induced obese mice with chronic cerebral hypoperfusion. CNS Neurosci Ther 29:200–212. https://doi.org/10.1111/cns.14301
Kalyan M, Tousif AH, Sonali S et al (2022) Role of endogenous lipopolysaccharides in neurological disorders. Cells 11:4308. https://doi.org/10.3390/cells11244038
Thevaranjan N, Puchta A, Schulz C et al (2017) Age-associated microbial dysbiosis promotes intestinal permeability, systemic inflammation, and macrophage dysfunction. Cell Host Microbe 21:455–466. https://doi.org/10.1016/j.chom.2017.03.002
Baratta F, Pastori D, Bartimoccia S et al (2020) Poor adherence to Mediterranean diet and serum polysaccharide are associated with oxidative stress in patients with non-alcoholic fatty liver disease. Nutrients 12:1732. https://doi.org/10.3390/nu12061732
André P, Pais de Barros JP, Merle BMJ et al (2021) Mediterranean diet and prudent diet are both associated with low circulating esterified 3-hydroxy fatty acids, a proxy of LPS burden, among older adults. Am J Clin Nutr 114:1080–1091. https://doi.org/10.1093/ajcn/nqab126
Del Bò C, Bernardi S, Cherubini A et al (2021) A polyphenol-rich dietary pattern improves intestinal permeability, evaluated as serum zonulin levels, in older subjects: the MaPLE randomised controlled trial. Clin Nutr 40:3006–3018. https://doi.org/10.1016/j.clnu.2020.12.014
Peron G, Gargari G, Meroño T et al (2021) Crosstalk among intestinal barrier, gut microbiota and serum metabolome after a polyphenol-rich diet in older subjects with “leaky gut”: the MaPLE trial. Clin Nutr 40:5288–5297. https://doi.org/10.1016/j.clnu.2021.08.027
Hidalgo-Liberona N, González-Domínguez R, Vegas E et al (2020) Increased intestinal permeability in older subjects impacts the beneficial effects of dietary polyphenols by modulating their bioavailability. J Agric Food Chem 68:12476–12484. https://doi.org/10.1021/acs.jafc.0c04976
Mörkl S, Lackner S, Meinitzer A et al (2018) Gut microbiota, dietary intakes and intestinal permeability reflected by serum zonulin in women. Eur J Nutr 57:2985–2997. https://doi.org/10.1007/s00394-018-1784-0
Seethaler B, Nguyen NK, Basrai M et al (2021) Short-chain fatty acids are key mediators of the favorable effects of the Mediterranean diet on intestinal barrier integrity: data from the randomized controlled LIBRE trial. Am J Clin Nutr 116:928–942. https://doi.org/10.1093/ajcn/nqac175
Castro-Barquero S, Lamuela-Raventós RM, Doménech M et al (2018) Relationship between Mediterranean dietary polyphenol intake and obesity. Nutrients 10:1523. https://doi.org/10.3390/nu10101523
Ticinesi A, Mancabelli L, Carnevali L et al (2022) Interaction between diet and microbiota in the pathophysiology of Alzheimer’s disease: focus on polyphenols and dietary fibers. J Alzheimers Dis 86:961–982. https://doi.org/10.3233/JAD-215493
Ticinesi A, Nouvenne A, Cerundolo N et al (2023) Accounting gut microbiota as the mediator of dietary (poly)phenols on skeletal muscle in aging. Nutrients 15:2367. https://doi.org/10.3390/nu15102367
Narduzzi L, Agulló V, Favari C et al (2022) (Poly)phenolic compounds and gut microbiome: new opportunities for personalized nutrition. Microbiome Res Rep 1:16. https://doi.org/10.20517/mrr.2022.06
Cortés-Martín A, Selma MV, Tomás-Barberán FA et al (2020) Where to look into the puzzle of polyphenols and health? The postbiotics and gut microbiota associated with human metabotypes. Mol Nutr Food Res 64:e1900952. https://doi.org/10.1002/mnfr.201900952
García-Villalba R, Giménez-Bastida JA, Cortés-Martín A et al (2022) Urolithins : a comprehensive update on their metabolism, bioactivity, and associated gut microbiota. Mol Nutr Food Res 66:e2101019. https://doi.org/10.1002/mnfr.202101019
Liu S, D’Amico D, Shankland E et al (2022) Effect of urolithin A supplementation on muscle endurance and mitochondrial health in older adults: a randomized clinical trial. JAMA Netw Open 5:e2144279. https://doi.org/10.1001/jamanetworkopen.2021.44279
Singh A, D’Amico D, Andreux PA et al (2022) Urolithin A improves muscle strength, exercise performance, and biomarkers of mitochondrial health in a randomized trial in moddle-aged adults. Cell Rep Med 3:100633. https://doi.org/10.1016/j.xcrm.2022.100633
Kujawska M, Jourdes M, Kurpik M et al (2019) Neuroprotective effects of pomegranate juice against Parkinson’s disease and presence of ellagitannins-derived metabolite-urolithin A-in the brain. Int J Mol Sci 21:202. https://doi.org/10.3390/ijms21010202
DaSilva NA, Nahar PP, Ma H et al (2019) Pomegranate ellagitannin-gut microbial-derived metabolites, urolithins, inhibit neuroinflammation in vitro. Nutr Neurosci 22:185–195. https://doi.org/10.1080/1028415X.2017.1360558
Gong Z, Huang J, Xu B et al (2019) Urolithin A attenuates memory impairment and neuroinflammation in APP/PS1 mice. J Neuroinflamm 16:62. https://doi.org/10.1186/s12974-019-1450-3
D’Amico DD, Andreux PA, Valdés P et al (2021) Impact of the natural compound urolithin A on health, disease, and aging. Trends Mol Med 27:687–699. https://doi.org/10.1016/j.molmed.2021.04.009
Zelicha H, Kloting N, Kaplan A et al (2022) The effect of high-polyphenol Mediterranean diet on visceral adiposity: the DIRECT PLUS randomized controlled trial. BMC Med 20:327. https://doi.org/10.1186/s12916-022-02525-8
Kaplan A, Zelicha H, Meir AY et al (2022) The effect of a high-polyphenol Mediterranean diet (Green-MED) combined with physical activity on age-related brain atrophy: the Dietary Intervention Randomized controlled trial polyphenols unprocessed study (DIRECT PLUS). Am J Clin Nutr 115:1270–1281. https://doi.org/10.1093/ajcn/nqac001
Imperatrice M, Cuijpers I, Troost FJ et al (2022) Hesperidin function as an ergogenic aid by increasing endothelial function and decreasing exercise-induced oxidative stress and inflammation, thereby contributing to improved exercise performance. Nutrients 14:2955. https://doi.org/10.3390/nu14142955
Martínez Noguera FJ, Alcaraz PE, Vivas JC et al (2021) 8 weeks of 2S-hesperidin supplementation improves muscle mass and reduces fat in amateur competitive cyclists: randomized controlled trial. Food Funct 12:3872–3883. https://doi.org/10.1039/d0fo03456h
Thenmozhi AJ, Raja TRW, Janakiraman U et al (2015) Neuroprotective effects of hesperidin on aluminium chloride induced Alzheimer’s disease in Wistar rats. Neurochem Res 40:767–776. https://doi.org/10.1007/s11064-015-1525-1
Javed H, Vaibhav K, Ahmed ME et al (2015) Effect of hesperidin on neurobehavioral, neuroinflammation, oxidative stress and lipid alteration in intracerebroventricular streptozotocin induced cognitive impairment in mice. J Neurol Sci 348:51–59. https://doi.org/10.1016/j.jns.2014.10.044
Al-Aubaidy HA, Dayan A, Deseo MA et al (2021) Twelve-week mediterranean diet intervention increases citrus bioflavonoid levels and reduces inflammation in people with type 2 diabetes mellitus. Nutrients 13:1133. https://doi.org/10.3390/nu13041133
Mayo B, Vázquez L, Flórez AB (2019) Equol: a bacterial metabolite from the daidzein isoflavone and its presumed beneficial health effects. Nutrients 11:2231. https://doi.org/10.3390/nu11092231
Sekikawa A, Wharton W, Butts B et al (2022) Potential protective mechanisms of S-equol, a metabolite of soy isoflavone by the gut microbiome, on cognitive decline and dementia. Int J Mol Sci 23:11921. https://doi.org/10.3390/ijms231911921
Johnson SL, Kirk RD, DaSilva NA et al (2019) Polyphenol microbial metabolites exhibit gut and blood-brain barrier permeability and protect murine microglia against LPS-induced inflammation. Metabolites 9:78. https://doi.org/10.3390/metabo9040078
Igase M, Igase K, Tabara Y et al (2017) Cross-sectional study of equol producer status and cognitive impairment in older adults. Geriatr Gerontol Int 17:2103–2108. https://doi.org/10.1111/ggi.13029
Dent E, Morley JE, Cruz-Jentoft AJ et al (2018) International clinical practice guidelines for sarcopenia (ICFSR): screening, diagnosis and management. J Nutr Health Aging 22:1148–1161. https://doi.org/10.1007/s12603-018-1139-9
Chen LK, Arai H, Assantachai P et al (2022) Roles of nutrition in muscle health of community-dwelling older adults: evidence-based expert consensus from Asian working group for sarcopenia. J Cachexia Sarcopenia Muscle 13:165–1672. https://doi.org/10.1002/jcsm.12981
Nunes EA, Colenso-Semple L, McKellar SR et al (2022) Systematic review and meta-analysis of protein intake to support muscle mass and function in healthy adults. J Cachexia Sarcopenia Muscle 13:795–810. https://doi.org/10.1002/jcsm.12922
Mervant L, Tremblay-Franco M, Olier M et al (2023) Urinary metabolome analysis reveals potential microbiota alteration and electrophilic burden induced by high red meat diet: results from the French NutriNet-Santé cohort and an in vivo intervention study in rats. Mol Nutr Food Res 67:e2200432. https://doi.org/10.1002/mnfr.202200432
Wang Y, Uffelman CN, Bergia RE et al (2023) Meat consumption and gut microbiota: a scoping review of literature and systematic review of randomized controlled trials in adults. Adv Nutr 14:215–237. https://doi.org/10.1016/j.advnut.2022.10.005
Prokopidis K, Cervo MM, Grandham A et al (2020) Impact of protein intake in older adults with sarcopenia and obesity: a gut microbiota perspective. Nutrients 12:2285. https://doi.org/10.3390/nu12082285
Ni Lochlainn M, Bowyer RCE, Steves CJ (2018) Dietary protein and muscle in aging people: the potential role of gut microbiome. Nutrients 10:929. https://doi.org/10.3390/nu10070929
Lin CC, Shih MH, Chen CD et al (2021) Effects of adequate dietary protein with whey protein, leucine, and vitamin D supplementation on sarcopenia in older adults: an open-label, parallel-group study. Clin Nutr 40:1323–1329. https://doi.org/10.1016/j.clnu.2020.08.017
Ticinesi A, Meschi T, Lauretani F et al (2016) Nutrition and inflammation in older individuals: focus on vitamin D, n-3 polyunsaturated fatty acids and whey proteins. Nutrients 8:186. https://doi.org/10.3390/nu8040186
Negm AM, Lee J, Hamidian R et al (2022) Management of sarcopenia: a network meta-analysis of randomized controlled trials. J Am Med Dir Assoc 23:707–714. https://doi.org/10.1016/j.jamda.2022.01.057
Ford AL, Nagulesapillai V, Piano A et al (2020) Microbiota stability and gastrointestinal tolerance in response to a high-protein diet with and without a prebiotic, probiotic, and synbiotic: a randomized, double-blind, placebo-controlled trial in older women. J Acad Nutr Diet 120:500-516.e10. https://doi.org/10.1016/j.jand.2019.12.009
Wang Y, Lindemann SR, Cross TWL et al (2023) Effects of adding lean read meat to a U.S.-style healthy vegetarian dietary pattern on gut microbiota and cardiovascular risk factors in young adults: a crossover randomized controlled trial. J Nutr 153:1439–1452. https://doi.org/10.1016/j.tjnut.2023.03.013
Pahor M, Guralnik JM, Anton SD et al (2020) Impact and lessons from the lifestyle interventions and independence for elders (LIFE) clinical trials of physical activity to prevent mobility disability. J Am Geriatr Soc 68:872–881. https://doi.org/10.1111/jgs.16365
Bernabei R, Landi F, Calvani R et al (2022) Multicomponent intervention to prevent mobility disability in frail older adults: andomized controlled trial (SPRINTT project). BMJ 377:e068788. https://doi.org/10.1136/bmj-2021-068788
Ticinesi A, Lauretani F, Tana C et al (2019) Exercise and immune system as modulators of intestinal microbiome: implications for the gut-muscle axis hypothesis. Exerc Immunol Rev 25:84–95
Mailing LJ, Allen JM, Buford TW et al (2019) Exercise and the gut microbiome: a review of the evidence, potential mechanisms, and implications for human health. Exerc Sports Sci Rev 47:75–85. https://doi.org/10.1249/JES.0000000000000183
Funding
Open access funding provided by Università degli Studi di Parma within the CRUI-CARE Agreement. Open access funding provided by Università degli Studi di Parma within the CRUI-CARE Agreement.
Author information
Authors and Affiliations
Contributions
Andrea Ticinesi contributed to concept and design of the review, literature search and analysis, interpretation, and preparation of the manuscript. Antonio Nouvenne contributed to concept and design of the review, and manuscript draft. Nicoletta Cerundolo contributed to literature search and analysis. Alberto Parise contributed to literature search and analysis. Pedro Mena contributed to concept and design of the review and manuscript draft. Tiziana Meschi provided supervision and contributed to study concept and design.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose in relation with the present manuscript.
Ethical approval
Not applicable—this work does not report original research involving human subjects.
Human and animal rights
This article does not contain any studies with human participants or animals performed by any of the authors.
Informed consent
For this type of study, formal consent is not required.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ticinesi, A., Nouvenne, A., Cerundolo, N. et al. The interaction between Mediterranean diet and intestinal microbiome: relevance for preventive strategies against frailty in older individuals. Aging Clin Exp Res 36, 58 (2024). https://doi.org/10.1007/s40520-024-02707-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s40520-024-02707-9