
Abstract
Biological traits related to the adherence strategy of periphytic algae determine species’ dispersal ability and the degree of protection against physical disturbance, such as strong currents and flood pulse. To help understand how water level can structure species with distinct adherence strategy in a floodplain, we investigated periphytic algal beta diversity in subtropical floodplain lakes (upper Paraná river floodplain) and evaluate the importance of environmental and spatial drivers in groups with limited dispersal and with higher dispersal abilities. The beta diversity was analyzed considering both presence–absence and species density. We used variance partitioning to evaluate the importance of environmental and spatial processes on periphytic algae. Our results registered 181 species, and the class Bacillariophyceae presented the highest species richness and density. Beta diversity did not differ between months with distinct water levels but differed among lakes. Set of species with higher dispersal abilities presented higher values of beta diversity. Environmental variables were the main driver of periphytic algae composition and density, but spatial variables also drive a significant amount of differences in species composition and density. Beta diversity of species with limited dispersal abilities was mainly driven by environmental factors, and no driver was important to species with higher dispersal. The results of beta diversity indicated that nearest lakes may be more environmentally similar that allows the same species to survive, and also allows species to disperse better among them. These results highlight that the inclusion of functional aspects can clarify the understanding of beta diversity patterns.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Understanding the mechanisms that structure the biological diversity through space and time is one of the main goals of ecology (Heino et al. 2015), and remains a great challenge for the operation, management, and conservation of ecosystems and biodiversity (Legendre et al. 2005; Logue et al. 2011; Dray et al. 2012; Heino et al. 2015). Using beta diversity in biodiversity analyzes allows assessing changes in species composition and density between locations or in time (Anderson et al. 2006). Higher variations in species composition are related to an increase in differences of environmental conditions among habitats, and in the spatial distance among habitats (Stegen et al. 2013; Astorga et al. 2014). Differences in environmental conditions among habitats act as filters and may restrict the establishment of species with unsuited traits (Cornwell and Ackerly 2009; Weiher et al. 2011) and allow the persistence only of tolerant species in the habitat (Astorga et al. 2014).
Periphytic algae are a complex aquatic community of organisms that adhere to submerged substrates and go through successional processes. Several abiotic and biotic factors can alter the establishment of these organisms in the community (Stevenson 1996; Graham et al. 2016). All these factors together and the complexity of the community interfere with the understanding of beta diversity patterns. The functional traits of the community are important for protection against losses caused by physical disturbances (Biggs et al. 1998), such as strong currents and flood pulse (Dunck et al. 2016), and can favor the spread of propagules, facilitating dispersal (Algarte et al. 2014, 2017a). The adherence strategy includes the traits of adherence form and the strength of adherence to the substrate (Biggs et al. 1998; Lange et al. 2015; Tapolczai et al. 2016), and the way species adhere to the substrate can culminate in distinct community architecture and organization of species between communities of different sites (beta diversity patterns) (Dunck et al. 2016, 2019). Thus, species that are loosely attached to substrates should be able to easily disperse through sites that are connected, leading to lower beta diversity of loosely attached species among samples since the samples share more species (Algarte et al. 2014, 2017a). Otherwise, species firmly attached to substrates are not great dispersers since they are not able to reach samples far apart, which would lead to high beta diversity of firmly attached species among samples (Algarte et al. 2014, 2017a).
Studies about the beta diversity of periphytic algae have assessed the importance of environmental factors in the entire communities without considering the differences in adherence strategies. The main evaluated factors influencing periphytic algae were artificial eutrophication (Passy and Blanchet 2007; Dunck et al. 2019), physical factors such as substrate roughness (Petsch et al. 2017), and hydrological factors in floodplains (Dunck et al. 2016). The patterns presented in these studies were that beta diversity of periphytic algae was smaller in temperate sites under a human-impacted context (Passy and Blanchet 2007); in tropical streams, the human impacts have not led to a reduction in periphytic algal beta diversity (Dunck et al. 2019); and that higher beta diversity of periphytic algae occurs in more complex substrata (Petsch et al. 2017).
Periphytic algal communities are abundant in floodplains, which are aquatic ecosystems with high heterogeneity and complexity of conditions. In these systems, periphytic algae propagate easily and colonize several submerged substrates by movements in the water column or those generated by seasonal changes in the hydrological regime (Algarte et al. 2009; Ferragut and Bicudo 2009; Rodrigues et al. 2013). These ecosystems have seasonal fluctuations in the water level related to flood pulse (Junk and Wantzen 2004), which are essential factors in the rearrangement of physical characteristics and consequently the organization and dynamics of aquatic ecosystems and communities (Death 2010). Studies have shown that in the upper Paraná river floodplain, when the water level reaches > 4.5, a homogenization occurs in the system, reflecting higher similarity in environmental conditions and communities among the sites from the floodplain (Thomaz et al. 2007). However, in the Paraná River floodplain, a dam has altered the water level fluctuations, controlling seasonal patterns of water level (Roberto et al. 2009). Thus, periphytic algal communities may present different types of spatial and temporal dynamics related to variation in water levels, which reflect changes in the local communities’ organization (Chase et al. 2005).
To help understand the complexity of the periphytic algal communities and how water level can structure species with distinct adherence strategy, we used the presence–absence and density data of the periphytic algal communities adhered to the emergent macrophyte Pontederia azurea Sw from four lakes in the upper Paraná river floodplain between the years 2014 and 2015. We disentangle the entire communities into two groups with a distinct adherence strategy (firmly attached to substrates/limited dispersal, and loosely attached/higher dispersal abilities). We estimated the beta diversity patterns in each case (entire community, only for loosely attached, and only for firmly attached species).
As some assumptions for our study, we use the pattern of distance-decay previously reported for periphyton algae, in which sites closer together share more species than sites that are far apart (Astorga et al. 2012; Wetzel et al. 2012). We also consider the way that water level increases can structure periphytic algae composition, making the composition more similar when water reaches levels high enough to favor dispersion between the river and lakes, while a more heterogeneous composition is present in lower water levels when lakes are not connected (Thomaz et al. 2007). Considering the distinct adherence strategy of periphytic algae and the assumption that species loosely attached to substrates can disperse more, then the lakes in this floodplain would share more species loosely attached, leading to a decrease in beta diversity (Algarte et al. 2014; Jamoneau et al. 2018). Taking the main drivers in the floodplain, we expected that stronger spatial drivers structuring species with limited dispersal would be related to the capacity of these species to firmly attach to the substrate, in this way, these species would not be able to reach sites too far from each other (Biggs et al. 1998; Algarte et al. 2014).
In this study, we investigated how changes in the water level of lentic environments from a subtropical floodplain (upper Paraná river floodplain) and different adherence strategy alters beta diversity patterns of periphytic algal communities. Also, we evaluate the importance of environmental and spatial drivers in groups with distinct adherence strategies. We tested the hypotheses that (1) lakes closer to each other would share more species, while sites too far from each other would differ in species composition, thus not sharing many species; (2) the increase in water level leads to a decrease of periphytic algal beta diversity among all lakes; (3) set of species with higher dispersal abilities have lower values of beta diversity; (4) environmental processes are the main forces structuring periphytic algae communities, but spatial processes are a stronger driver of changes in groups of algae with limited dispersal.
2 Materials and methods
Study area
– This study was conducted in the upper Paraná river floodplain, located in Brazilian territory between the states of Mato Grosso do Sul and Paraná, in one undammed stretch of the river (Souza Filho and Stevaux 1997). This stretch is located between the reservoirs of Porto Primavera-SP and Itaipu-PR and is an area of great importance for the preservation and conservation of the local aquatic biodiversity (Agostinho et al. 2004). This floodplain is characterized by highly complex aquatic environments such as canals and lakes, with different degrees of connectivity among them.
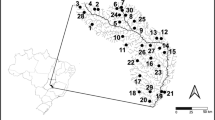
The samplings occurred quarterly during months of low-water levels (May 2014 and 2015, and August 2014) and during months of high-water levels (November and February 2014) in four lakes (Ivaí, São João, Pavão, and Saraiva), that are located along the longitudinal gradient of the Paraná River (Fig. 1). These lakes are connected to the Paraná River and exhibit abundant and rich macrophyte banks, in which Pontederia azurea Sw (= Eichhornia azurea (Sw.) Kunth) is the most abundant macrophyte and was used as a substrate for obtaining periphytic algal communities. We considered each of the five sampled months as a different sample for the development of subsequential analyses, thus, a total of 20 samples (4 lakes × 5 months = 20 samples).

Location of sampling points. Red points represent the Lakes of the upper Paraná river floodplain: 1 Ivai Lake, 2 São João Lake, 3 Pavão Lake and 4 Saraiva Lake
Ivai Lake (23° 16′ 84″ S, 53° 42′ 21.0″ W) has an asymmetrical format with banks of macrophytes in the coastal zone and is located in the right margin of the Paraná River, presenting a short connectivity canal. São João Lake (23° 49′ 20.9″ S, 53° 59′ 16.6″ W), has an oval shape, with approximately 3 km in length and 1.8 m depth, and is situated on the right bank of the respective Paraná river. Ivai Lake is connected to the river through a channel of 2 km in length. Pavão Lake (23° 58′ 31.1″ S, 54° 09′ 49.1″ W) has an irregular-elongated shape, with approximately 1 km in length and with an average width of 20 m. This lake has approximately 2.6 m depth and its connectivity canal with the Paraná river is short and narrow. Saraiva Lake (24° 00′ 79″ S, 54° 08′ 23.3″ W) is considerably larger than the other studied lakes, has an elongated shape with approximately 9.5 km in length with an average width of 180 m and approximately 2.6 m depth.
Limnological variables
– The limnological variables were measured concomitantly with biological sampling in the same sampling points in all samplings. The following limnological variables were measured in the limnetic zone of the lakes with multiparameter Horiba U-50: water temperature (°C), dissolved oxygen (% of saturation and mg L−1), pH, electrical conductivity (µS cm−1), turbidity (NTU). Transparency (m) was estimated using a Secchi disk. Two water samples were taken per lake, thus two subsamples per lake, and filtered through Whatman GF/F filters, under low pressure (0.5 atm) and stored at − 20 °C for later determination of alkalinity (µEq L−1), ammoniacal nitrogen (NH3), orthophosphate (PO4), nitrate (NO3), total nitrogen (TN) and total phosphorus (TP) (μg L−1) according to Mackereth et al. (1978) and Roberto et al. (2009). The data of the Paraná river water level was obtained by the pluvial metric station of Porto Rico. The limnological variables were analyzed and provided by the Laboratory of Limnology of the Nucleus of Research in Limnology, Ichthyology, and Aquaculture (Nupélia) of Maringá State University.
Periphytic algal communities
– The periphytic material was obtained from the sixth or seventh plant internode of submerged mature petioles of Pontederia azurea Sw (= Eichhornia azurea (Sw.) Kunth), totaling two subsamples per lake (Schwarzbold 1990). The plants were sampled on the marginal region of different macrophyte patches located in the littoral zone of the lakes. The petioles of each lake were conditioned in 150 mL Wheaton bottles and kept on ice. The periphytic material was removed from petioles in the laboratory by a stainless-steel blade wrapped in aluminum foil and by jets of distilled water (Algarte et al. 2014). The removed material was fixed and preserved with acetic Lugol’s to 5% (Bicudo and Menezes 2006) and the scrapping area was measured.
Qualitative analyzes of periphytic algae were made through temporary glass slides preparation, observed under an optical binocular microscope with millimeter oculars and clear chamber attached, using the increases to 400 × and 1000 × (Bicudo and Menezes 2006). The identifications were made until the lowest level of identification possible, using taxonomic keys and specialized literature (Anagnostidis and Komárek 1988, and Komárek and Anagnostidis 1989 for Cyanophyceae; Round 1971 and Bicudo and Menezes 2006 for the other classes). We used the Utermöhl method (1958) to quantify the algae, and the counting was conducted in random fields on an inverted Olympus CK2 microscope according to two standardized criteria; until we reached at least 100 individuals of the most common species (cells, colonies, and filaments) of each sample and until a species–accumulation curve reached an asymptote (Ferragut and Bicudo 2012). The density of periphytic algal communities was calculated by the equation proposed by Ros (1979), adapted to the substrate area. The results were expressed per unit of area (ind cm−2).
The periphytic algal classification according to adherence strategy in firmly (limited dispersal) or loosely (higher dispersal abilities) attached to the substrate was made following Sládecková and Sládecek (1977) (Electronic supplementary material-ESM1). Periphytic algae with adherence form among mobile and entangled (Biggs et al. 1998) were classified as loosely attached to the substrate, while periphytic algae with adherence form among prostrate, stalked, mucilage tube or heterotrichous (Biggs et al. 1998) were classified as firmly attached to the substrate.
Data analysis
– A principal component analysis (PCA, Legendre and Legendre 1998) was performed to segregate the lakes according to their total environmental variables (turbidity, dissolved oxygen, pH, electrical conductivity, alkalinity, TN, and TP) to evaluate if the lakes and hydrological levels would be characterized by specific environmental variables. The data was previously standardized and the two first axes that had a broken-stick value smaller than the PCA eigenvalues were selected to further interpretation (Legendre and Legendre 1998).
To assess how periphytic algae composition changed across lakes we estimated beta diversity using two indexes: Jaccard (presence–absence matrix) and Bray–Curtis (species densities matrix). The data regarding species’ density was previously logarithmized to avoid discrepant values of species density. Regarding the indexes, we choose to use Bray–Curtis to detect changes in species densities across lakes and water levels, and Jaccard, in its turn, is useful to detect changes in the composition (occurrence) of species (Koleff et al. 2003; Barwell et al. 2015). In this way, we can infer the processes driving changes in species composition as well as changes in the density of these species. We estimated the beta diversity patterns in each case: for the whole community among all lakes and months, for the group of firmly attached among all lakes and months, and for the group of loosely attached among all lakes and months. We conducted six permutational analysis of variance (PERMANOVA; Anderson 2001) to test for differences in species composition among groups of lakes and water levels for the whole community at all sampled months, separately for loosely attached species, and separately for firmly attached species. We also performed six permutational analysis of multivariate dispersions (PERMDISP; Anderson 2006) to test if values of beta diversity differ among lakes and water levels for the whole community among all replicated months, for only loosely attached species, and for only firmly attached species. Whenever a significant difference was found among groups of lakes, we conducted a post hoc test to test for pairwise differences among lakes, using Bonferroni’s correction to estimate P values since we are dealing with multiple comparisons. We also conducted a T test to search for differences in centroid values of beta diversity among species with different adherence strategies. In the T test, our response variable was the beta diversity centroids in each sample of Jaccard’s and Bray–Curtis indexes and our categorical predictor was the adherence strategy of species (loosely and firmly attached). We used 9999 permutations for all tests.
We investigated the importance of environmental and spatial processes on periphytic algae using two datasets. The first dataset was the environmental variables matrix (turbidity, dissolved oxygen, pH, electrical conductivity, alkalinity, TN, and TP; the environment component) and the second was the fluvial distance matrix between sites (spatial component). The environmental variables were previously standardized for further analysis with the environmental component (Legendre and Gallagher 2001). Spatial variables were assessed by submitting the fluvial distance matrix between sites to a principal coordinate of neighbor matrices (PCNM), which generated the spatial filters reflecting the fluvial distance matrix. These spatial filters were used in further analysis as the spatial component (Borcard and Legendre 2002).
We tested the importance of environmental and spatial components as potential drives of beta diversity patterns of periphytic algae species with limited dispersal (firmly attached to substrates) and with higher dispersal abilities (loosely attached to substrates). The standardized environmental variables and spatial filters were previously submitted to forward selection to identify which variables of these two datasets were related to the response matrixes (Jaccard’s and Bray–Curtis’ beta diversity of periphytic algae species with limited dispersal and with higher dispersal abilities) (Blanchet et al. 2008). P values under 0.05 and maximum values of adjusted R2 equal to the one generated by partial distance-based redundancy analysis (dbRDA) across beta diversity and environmental variables were used as a threshold in the forward selection. The selected environment and spatial variables were used in the subsequent analysis as the environment and spatial components.
We performed three variance partitioning (Borcard et al. 1992) to identify the unique and shared contributions of the environment (as the environment standardized variables selected in the forward selection) and spatial component (as the spatial filters selected in the forward selection) on Jaccard’s and Bray–Curtis’ beta diversity indexes obtained for three different matrixes: (1) considering all species in all lakes and months (thus, for the 20 samples); (2) considering only loosely attached species in all lakes and months; and (3) considering only firmly attached species in all lakes and months. Although we sampled four lakes only, we accounted for the spatial and seasonal variation within lakes, increasing our sampling range width (Fischer 2019), and we also corrected our data following the adjusted R2 method unbiased by sampling size, proposed by Peres-Neto et al. (2006). Thus, we choose to include variance partitioning even with our small dataset, given that we adopted such precautions. The adjusted R2 of the unique fractions from the environment and spatial components were accessed through a dbRDA, in which we checked for responses in the distance matrixes (i.e., Bray–Curtis and Jaccard’s indexes; Borcard and Legendre 2002). This allows us to search for the effect of a component while controlling the other (i.e., identifying the significance of the environment component in the beta diversity matrix while controlling the influence of the spatial component) (Montag et al. 2018). The dbRDA were then tested through analysis of variance (ANOVA; 10,000 permutations). All analyses were performed in the software R (R Development Core Team 2014), using the packages “vegan” (Oksanen et al. 2013), “adespatial” (Dray et al. 2019), and ade4 (Dray and Dufour 2007).
3 Results
The water level variation and limnological conditions
– The variation of the hydrologic regime of the Paraná river during the study period was characterized by the occurrence of short pulses (< 3.5 m). Increases in the water levels were registered from November 2014 until April 2015, with values between 2.5 and 3.0 m. From the 5 months that we sampled, November 2014 and February 2015 were considered the months with higher water levels of the study (> 2.5 m), while May 2014 and 2015, and August 2014 were considered months of lower water levels (< 2.5 m; Fig. 2).

Daily variation of hydrometric levels of the Paraná river during the study period (May/2014 to May/2015). The arrows indicate the days of sampling of the periphytic algal community (08-09-11/05/2014, 06-07-08/08/2014, 04-05-08/11/2014, 03-04-08/02/2015, and 06-07-10/05/2015)
The main variability in the limnological variables occurred among lakes (spatial scale) instead of the sampling months (Fig. 3). The PCA axes selected through the broken-stick method were axes two and three, which summarized 41.14% of the variability of the limnological variables (Electronic supplementary material-ESM2). The second axis was correlated mainly with total phosphorus (r = 0.637) and total nitrogen (r = 0.622; Electronic supplementary material-ESM2). The third axis was correlated mainly with pH (r = − 0.873). The PCA representation demonstrated a distinction between the lakes concerning its nutrients (P and N; Fig. 3). The majority of samples in Ivaí and Saraiva lakes showed higher values of total nutrients than Pavão and São João (Fig. 3). We also observed a pattern where the majority of the sites sampled in months of low-water levels showed lower pH values (more acidic), while most samples in months of high-water levels showed the highest values of pH (more basic) (Fig. 3).

Representation of the principal components analysis (PCA). Ivaí lake (circles), Pavão lake (squares), São João lake (diamond), and Saraiva lake (triangle). Non-filled shapes represent samples in the low-water levels, while black-filled shapes represent samples in the high-water levels. TP total phosphorus; NT total nitrogen
Periphytic algal communities
– Periphytic algae were represented by 181 taxa, within nine classes (51 Bacillariophyceae, 42 Chlorophyceae, 36 Zygnemaphyceae, 17 Cyanophyceae, 16 Euglenophyceae, 10 Xanthophyceae, 7 Oedogoniophyceae, one Chlamydophyceae, and one Chrysophyceae). Bacillariophyceae was the richest class, followed by Chlorophyceae and Zygnemaphyceae. May (144) and February (131) of 2015 were the richest months. Bacillariophyceae was also the most abundant class in organism density, followed by Oedogoniophyceae and Cyanophyceae. August and May of 2014 presented the highest values of algal density and February of 2015 the lowest density. Pavão Lake showed the highest values of periphytic algae density over time, especially in low water levels (Electronic supplementary material-ESM1).
Species composition and beta diversity
– For the whole community, considering the Jaccard index, species composition differed among the lakes (Table 1), but did not differ among months with distinct hydrological levels, as returned by the PERMANOVA (Table 1; Electronic supplementary material-ESM2). For the Bray–Curtis index, species composition also differed among the lakes (Table 1) but did not differ among months with higher and lower hydrological levels (Table 1; Electronic supplementary material-ESM2). The pairwise comparisons revealed that species composition (for Jaccard as well as for Bray–Curtis) were not different between the closest lakes (Pavão and Saraiva; Table 1). The most isolated lake (Ivaí) was consistently different from São João (the intermediate lake) but varies in differences depending on the index when we compared with Pavão and Saraiva, the most distant lakes from Ivaí (Table 2). Regarding differences in the variance of beta diversity within lakes (PERMDISP), there was no difference among lakes, nor hydrological levels (Fig. 4) for any of the beta diversity indexes (Table 1).

Boxplots of distance to centroid values of Jaccard (a) and Bray–Curtis (b) beta diversity indexes between months of different water levels (< 2.5 and > 2.5 m). Central lines in the boxplots represent mean distances to centroids, thus beta diversity, boxes are its confidence interval, and spreads are its minimum and maximum values
When we conducted the analyses to identify differences in species composition between groups of distinct adherence strategy, we found that loosely and firmly attached species are both different among lakes for Jaccard’s index as well as for Bray–Curtis, but not for months of different water level (Table 1; Electronic supplementary material-ESM2). The pairwise comparisons varied in differences among lakes depending on the index: for the Jaccard index, the closest lakes (Pavão and Saraiva) were not different from each other regarding loosely attached species’ composition, but it was different for firmly attached (Table 1). As for the Bray–Curtis index, the closest lakes differed regarding loosely attached species’ density but did not for firmly attached (Table 1). Species composition in the most isolated lake (Ivaí) was different from the ones found in the intermediate lake (São João) for both groups of distinct adherence strategy considering the Jaccard index but differed only for loosely attached species when we considered the Bray–Curtis index (Table 1). Yet, Ivaí was different from Pavão, a distant lake, but was not different from Saraiva, the neighbor lake of Pavão (Table 2). Regarding differences in the variance of beta diversity within the lakes (PERMDISP), there was no difference among different lakes, nor hydrological levels for neither groups of distinct adherence strategy or beta diversity indexes (Table 1). When we tested for differences in distances to centroid values of beta diversity between loosely and firmly attached species, we found that these two groups with distinct adherence strategy were different from each other regarding its beta diversity (Jaccard: T = 8.58, DF = 1, P < 0.01; Bray–Curtis: T = 9.39, DF = 1, P < 0.01); Loosely attached species had higher values of beta diversity than firmly attached species in Jaccard’s index as well as in Bray–Curtis’ index, indicating that loosely attached species’ composition and density changes more among lakes and months than the composition and density of firmly attached species (Fig. 5).

Boxplots of distance to centroid values of Jaccard (a) and Bray–Curtis (b) beta diversity indexes between groups with distinct adherence strategies. Central lines in the boxplots represent mean distances to centroids, thus beta diversity, boxes are its confidence interval, and spreads are its minimum and maximum values
Importance of environmental and spatial drivers on periphytic algae
– Alkalinity, total phosphorus, total nitrogen, conductivity, and water turbidity were environmental variables often selected in the forward selection (Table 2). In terms of spatial variables, the PCNM axes representing the smaller distances (axes 2 and 3; see Borcard and Legendre 2002) were always selected for species with limited dispersal and the whole community, while the axis representing the greater distances (axis 1; Borcard and Legendre 2002) was often selected for species with higher dispersal. All environmental and spatial variables selected for the whole community and by groups with distinct adherence strategies are in Table 2.
When we considered the whole community, we found that both beta diversity indexes were mainly driven by environmental (Jaccard: adjusted R2 = 0.082, F = 1.858, P = 0.001; Bray–Curtis: adjusted R2 = 0.105, F = 2.516, P = 0.001; Table 2), but a lower significant part of beta diversity’s variance was also explained by spatial variables (Jaccard: adjusted R2 = 0.045, F = 1.883, P = 0.002; Bray–Curtis: adjusted R2 = 0.064, F = 2.319, P = 0.001; Table 2). The way beta diversity varies according to environmental differences and geographical distance among lakes was clear: The more two sites are different from each other concerning its environmental variables, the more different these two sites were regarding species’ composition and density (Fig. 6a); the same is true for geographical distance: the more distant two sites are from each other, the more they differ in species’ composition and density (Fig. 6b).

Scatterplots showing the relationship between Jaccard (black circles) and Bray–Curtis (gray squares) beta diversity indexes and a environmental variables (turbidity and total phosphorus; represented by a Euclidean distance between lakes) and b fluvial distance (km) between lakes. The lightest gray circles and darkest gray squares in graph b are the distances between São João lake to Pavão lake (56 km) which is almost the same as from Saraiva lake to São João lake (55.7 km), so we changed the colors of the distances for better visualization of the two distances
Concerning the drivers of beta diversity of groups with distinct adherence strategy, the Bray–Curtis index for species with limited dispersal (firmly attached) was not driven by environmental (adjusted R2 = 0.033, F = 1.265, P = 0.176; Table 2) nor spatial (adjusted R2 = 0.031, F = 1.317, P = 0.164; Table 2) components, but the Jaccard index was driven by both (Purely environment: adjusted R2 = 0.143, F = 2.032, P = 0.001; Purely space: adjusted R2 = 0.104, F = 2.063, P = 0.002; Table 2). Regarding species with higher dispersal (loosely attached), both of the indexes (Bray–Curtis and Jaccard) did not show any significant part of its variance explained by the environment (Bray–Curtis: adjusted R2 = 0.016, F = 1.147, P = 0.199; Jaccard: adjusted R2 = 0.006, F = 1.054, P = 0.374; Table 2) or space (Bray–Curtis: adjusted R2 = 0.016, F = 1.147, P = 0.199; Jaccard: adjusted R2 = 0.006, F = 1.054, P = 0.374; Table 2).
4 Discussion
Our results demonstrated, when we considered the whole community, that the closest lakes (Pavão and Saraiva) showed a similar species composition, but these lakes varied regarding beta diversity depending on the group of adherence strategy addressed, partially corroborating our first hypothesis. Yet, periphytic algal beta diversity did not differ between months with distinct water levels, which did not support the hypothesis that increases in water level lead to a decrease of periphytic algal beta diversity. Our third hypothesis concerning species with higher dispersal presenting lower values of beta diversity was not corroborated, and we found the opposite pattern, where the set of species with higher dispersal (loosely attached to the substrate) presented higher values of beta diversity. Related to our fourth hypothesis, we showed that the changes in the composition of species with limited dispersal (firmly attached) were mainly driven by environmental factors, but space also plays an important role, which partially confirms our hypothesis. We found no apparent driver of changes in species composition or density (Jaccard and Bray–Curtis indexes) for species with higher dispersal. The limnological variation in our study demonstrated that most upriver sites (Ivaí lake) and most downriver sites (Saraiva lake) presented higher nutrient availability, and that pH tended to change from more acidic in the low-water level to more basic in the high-water level.
Periphytic algal communities and beta diversity patterns
– Based on our predictions of species composition, for the whole community, some lakes differed regarding their species composition and density, in which the geographically closest lakes (Pavão and Saraiva) did not differ from each other. These results indicate that these two lakes may be environmentally structured in a similar way that allows the same species to survive in both lakes, and, since these two lakes are the closest together, species should be able to disperse within them easier than too far apart lakes, such as Ivaí (Wetzel et al. 2012). Indeed, Ivaí lake has a distinct composition than Pavão (a distant lake from Ivaí) and São João (an intermediate lake), although no differences were found between Ivaí and Saraiva (also a distant lake from Ivaí). These results corroborate our predictions that sites closer to each other share more species among them, even though the results for some sites apart from each other (specifically Ivaí and Saraiva) are fuzzy regarding changes in species composition and density.
The higher water level did not lead to lower beta diversity of periphytic algae. The water level in these months did not reach levels of homogenization (> 4.5 m), indeed, our sampling was conducted in a period of abnormal drought, when the highest water level reached was close to 3 m, which might have led to such results. In this way, the strong temporal variation documented for the Paraná floodplain may have not affected the periphytic algal beta diversity, as pointed for the organization of other aquatic communities in this floodplain (Thomaz et al. 2007). Dunck et al. (2016) also did not detect the importance of seasonal differences and wide variation in water level in periphytic algal beta diversity. The possible explanation is that the regulation of the water discharge regime imposed by hydroelectric dams in this floodplain provides unpredictable disturbances (Dunck et al. 2016). Such disturbance can be an important factor driving local colonization and extinction events, and override the expected effect of natural flood pulse regimes on aquatic communities (Padial et al. 2014). But the fact that beta diversity differed among lakes, reinforces the importance of local environmental factors and geographical distance, and the prolonged time of drought, including very low water level. Differences in species composition may be results of local environmental filters as well as geographical distance among lakes, which may restrict the occurrence of species to the local environmental conditions and sites far apart from each other. Yet, a higher variety of environmental conditions provides more variety of niches, leading to variation in species composition among localities, as well as sites too far from each other should vary more in species composition since some species are unable to disperse to such far sites (Leibold et al. 2004).
The inclusion of species adherence strategies in the evaluation of beta diversity patterns can clarify the understanding of the drivers of beta diversity patterns because these traits may facilitate or not dispersal. Our prediction of lower beta diversity for loosely attached was not corroborated. These results indicated that even with the capacity of easy spread of mobile and entangled species, some factor still provides the establishment of more distinct flora, that could be environmental drivers, as pointed out for beta diversity of diatom guilds (Jamoneau et al. 2018). However, the patterns of beta diversity of set of species with higher dispersal in the floodplain were not drive by space nor by the environment. These loosely attached species can be easily displaced from the substrate and be passively drifted away, yet, some species are motile and can actively disperse to other sites (Algarte et al. 2014). So, the lack of relationship between space and beta diversity of the set of species with higher dispersal was expected; since these species have great dispersal abilities, they are not limited by space and can reach other sites (as long as they are connected; Algarte et al. 2009; Wetzel et al. 2012). Surprisingly, the environmental variables evaluated in this study did not have any influence in changes of composition and density of loosely attached, what may indicate that other processes, not account for in our study, could be driving these changes, such as the hydrological characteristics of the flood itself (Fonseca and Rodrigues 2005; Algarte et al. 2006).
Importance of environmental and spatial drivers on periphytic algae
– When we considered the whole community, the beta diversity pattern of both indexes was driven mainly by the environment (especially conditions of turbidity and total phosphorus), but a smaller yet significant portion of the beta diversity pattern was driven by space (especially smaller distances). This reinforces that periphytic algae composition, as well as its density, are structured by the environmental characteristics of the lakes, specifically, differences in turbidity among lakes can select different species since higher turbidity leads to higher light limitation in the lake, which selected only for tolerant species to survive in these light deficient systems (Thomaz et al. 2007; Xiao et al. 2011). In another way, different phosphorus concentrations among lakes can also select for a different composition and density patterns of species since phosphorus is one of the main nutrients that limit algae population growth, thus, lakes that have a lower concentration of phosphorus may select only species tolerant to phosphorus deficiency (Xiao et al. 2011; Bozelli et al. 2015). In this context, the more different two lakes are regarding their environmental characteristics, the more different they will be regarding their species composition and density. Yet, space plays an important but smaller role in driving periphytic algae composition and density. Our analyses revealed a pattern of distance-decay, previously documented for the biological group studied (Wetzel et al. 2012), in which the more distant geographically two sites are, then the more different their composition and density of periphytic algae will be. This pattern was also described for stream fish (Benone et al. 2018), tropical trees (Condit 2002), diatoms, and macroinvertebrates (Astorga et al. 2012).
Regarding the drivers of beta diversity for groups with distinct adherence strategies, we found that space does play an important role in structuring changes in the composition of species with limited dispersal, reflecting their poor dispersal capacity, but environmental conditions also drive a similar amount of changes in composition. Periphytic algal species firmly attached to substrates are less likely to be displaced from the substrate they live on, which can be a useful adaptation in current waters and floodplains to avoid being passively drifted away from an ideal habitat (Algarte et al. 2014). But this adaptation also implies that these species are not able to reach sites too far from where they live, resulting in higher variation in species composition and density among distant sites (Soininen et al. 2007; Jamoneau et al. 2018). Yet, environmental conditions act as filters for species with limited dispersal, selecting species adapted to survive in specific conditions of nutrient availability, water conductivity, and alkalinity. Thus, beyond being limited by their poor dispersal ability, periphyton species firmly attached to substrates are yet limited by environmental conditions to establish themselves in the habitat. Although changes in the occurrence of species with limited dispersal between sites are driven by space and environment, changes in their densities are not. In a way, changes in density may be driven by other variables that we did not consider in this study, such as herbivory, as shown by Dunck et al. (2018) with an experimental approach demonstrating that different herbivores may structure periphyton density in different ways.
Concluding remarks
– In summary, we showed that lakes closer to each other present similar patterns of beta diversity, but higher water levels did not lead to lower periphytic algal beta diversity. Yet, local environmental factors act as filters in beta diversity differences among lakes, while spatial filters influence periphytic algae communities in a pattern of distance-decay, in which the more distant two sites are from each other, the more different they will be regarding species’ composition and density. We also demonstrate that species with distinct adherence strategies differ in beta diversity pattern in the floodplain, in which species with limited dispersal (firmly attached to substrates) are driven by environmental and spatial factors, while no apparent driver (neither environmental nor spatial variables approached in this study) was important to species with higher dispersal (loosely attached to substrates). These results highlight the importance of analyzing functional aspects of periphytic algal to understand beta diversity patterns. We also indicate that the inclusion of more traits related to life form and adherence strategy can clarify the understanding of the beta diversity patterns of periphytic algae.
References
Agostinho AA, Thomaz SM, Gomes LC (2004) Threats for biodiversity in the floodplain of the Upper Paraná River: effects of hydrological regulation by dams. Ecohydrol Hydrobiol 4:255–256
Algarte VM, Moresco C, Rodrigues L (2006) Algas do perifíton de distintos ambientes na planìcie de inundação do alto rio Paraná. Acta Sci Biol Sci 28:243–251. https://doi.org/10.4025/actascibiolsci.v28i3.216
Algarte V, Siqueira N, Murakami E, Rodrigues L (2009) Effects of hydrological regime and connectivity on the interannual variation in taxonomic similarity of periphytic algae. Braz J Biol 69:609–616. https://doi.org/10.1590/s1519-69842009000300015
Algarte VM, Rodrigues L, Landeiro VL, Siqueira T, Bini LM (2014) Variance partitioning of deconstructed periphyton communities: does the use of biological traits matter? Hydrobiologia 722:279–290
Algarte VM, Siqueira NS, Ruwer DT, Osório NC, Rodrigues L (2017a) Richness of periphytic algae and its relationship with hydrological attributes. Braz J Bot 40:1–6
Anagnostidis K, Komárek J (1988) Modern approach to the classification system of Cyanophytes, 3: Oscillatoriales. Algol Stud 80:327–472
Anderson MJ (2001) A new method for non-parametric multivariate analysis of variance. Austral Ecol 26:32–46
Anderson MJ (2006) Distance-based tests for homogeneity of multivariate dispersions. Biometrics 62:245–253
Anderson MJ, Ellingsen KE, McArdle BH (2006) Multivariate dispersion as a measure of beta diversity. Ecol Lett 9:683–693
Astorga A, Oksanen J, Luoto M, Soininen J, Virtanen R, Muotka T (2012) Distance decay of similarity in freshwater communities: do macro- and microorganisms follow the same rules? Global Ecol Biogeogr 21:365–375
Astorga A, Death R, Death F, Paavola R, Chakraborty M, Muotka T (2014) Habitat heterogeneity drives the geographical distribution of beta diversity: the case of New Zealand stream invertebrates. Ecol Evol 4:2693–2702
Barwell LJ, Isaac NJB, Kunin WE (2015) Measuring β-diversity with species abundance data. J Anim Ecol 84:1112–1122. https://doi.org/10.1111/1365-2656.12362
Benone NL, Ligeiro R, Juen L, Montag LFA (2018) Role of environmental and spatial processes structuring fish assemblages in streams of the eastern Amazon. Mar Freshw Res 69:243–252. https://doi.org/10.1071/MF17103
Bicudo CEM, Menezes M (2006) Gêneros de algas de águas continentais do Brasil: chaves para identificação e descrições. RiMa, São Carlos
Biggs BJF, Stevenson RJ, Lowe RL (1998) A habitat matrix conceptual model for stream periphyton. Arch Hydrobiol 143:21–56
Blanchet FG, Legendre P, Borcard D (2008) Forward selection of explanatory variables. Ecology 89:2623–2632
Borcard D, Legendre P (2002) All-scale spatial analysis of ecological data by means of principal coordinates of neighbour matrices. Ecol Model 153:51–68. https://doi.org/10.1016/j.catena.2016.01.006
Borcard D, Legendre P, Drapeau P (1992) Partialling out the spatial component of ecological variation. Ecology 73:1045–1055. https://doi.org/10.2307/1940179
Bozelli RL, Thomaz SM, Padial AA, Lopes PM, Bini LM (2015) Floods decrease zooplankton beta diversity and environmental heterogeneity in an Amazonian floodplain system. Hydrobiologia 753:233–241
Chase JM, Amarasekare P, Cottenie K, Gonzalez A, Holt RD, Holyoak M, Hoopes MF, Leibold MA, Loreau M, Mouquet N, Shurin JB, Tilman D (2005) Competing theories for competitive metacommunities. In: Holyoak M, Leibold MA, Holt RD (eds) Metacommunities: spatial dynamics and ecological communities. The University of Chicago Press, Chicago, pp 335–354
Condit R (2002) Beta-diversity in tropical forest trees. Science (80-) 295:666–669. https://doi.org/10.1126/science.1066854
Cornwell WK, Ackerly DD (2009) Community assembly and shifts in plant trait dis-tributions across an environmental gradient in coastal California. Ecol Monogr 79:109–126
Death RG (2010) Disturbance and riverine benthic communities: what has it con-tributed to general ecological theory? River Res Appl 26:15–25
Dray S, Dufour A (2007) The ade4 package: implementing the duality diagram for ecologists. J Stat Softw 22:1–20. https://doi.org/10.18637/jss.v022.i04
Dray S, Pélissier R, Couteron P, Fortin MJ, Legendre P, Peres-Neto PR, Bellier E, Bivand R, Blanchet FG, De Cáceres M, Dufour AB, Heegaard E, Jombart T, Munoz F, Oksanen J, Thioulouse J, Wagner HH (2012) Community ecology in the age of multivariate multiscale spatial analysis. Ecol Monogr 82:257–275
Dray S, Bauman D, Blanchet G, Borcard D, Clappe S, Guenard G, Jombart T, Larocque G, Legendre P, Madi N, Wagner HH (2019) Multivariate multiscale spatial analysis. R package version 0.3–7
Dunck B, Algarte VM, Cianciaruso MV, Rodrigues L (2016) Functional diversity and trait–environment relationships of periphytic algae in subtropical floodplain lakes. Ecol Indic 67:257–266
Dunck B, Amaral DC, Fernandes UL, Santana NF, Lopes TM, Rodrigues L (2018) Herbivory effects on the periphytic algal functional diversity in lake ecosystems: an experimental approach. Hydrobiologia 816:231–241
Dunck B, Felisberto SA, Nogueira IS (2019) Effects of freshwater eutrophication on species and functional beta diversity of periphytic algae. Hydrobiologia 837:195–204. https://doi.org/10.1007/s10750-019-03971-x
Ferragut C, Bicudo DC (2009) Efeito de diferentes níveis de enriquecimento por fósforo sobre a estrutura da comunidade Perifítica em represa oligotrófica tropical (São Paulo, Brasil). Braz J Biol 32:571–585
Fischer MM (2019) Quantifying the uncertainty of variance partitioning estimates of ecological datasets. Environ Ecol Stat 26:351–366
Ferragut C, Bicudo DC (2012) Effect of N and P enrichment on periphytic algal community succession in a tropical oligotrophic reservoir. Limnology 13:131–141
Fonseca IA, Rodrigues L (2005) Comunidade de algas perifíticas em distintos ambientes da planície de inundação do alto rio Paraná. Acta Sci Biol Sci 27:21–28. https://doi.org/10.4025/actascibiolsci.v27i1.1354
Graham LE, Graham JM, Wilcox LW, Cook ME (2016) Algae, 3rd edn. LJLM Press, LLC, Madison
Heino J, Melo AS, Tadeu S, Soininen J, Valanko S, Bini LM (2015) Metacommunity organisation, spatial extent and dispersal in aquatic systems: patterns, processes and prospects. Freshw Biol 60:845–869
Jamoneau A, Passy SI, Soininen J et al (2018) Beta diversity of diatom species and ecological guilds: response to environmental and spatial mechanisms along the stream watercourse. Freshw Biol 63:62–73. https://doi.org/10.1111/fwb.12980
Junk WJ, Wantzen KM (2004) The flood pulse concept: new aspects, approaches,and applications—an update. In: Welcomme R, Petr T (eds) Proceedings of the 2nd large river symposium (LARS). RAP Publication, Bangkok, pp 117–149
Koleff P, Gaston KJ, Lennon JJ (2003) Measuring beta diversity for presence–absence data. J Anim Ecol 72:367–382. https://doi.org/10.1046/j.1365-2656.2003.00710.x
Komárek J, Anagnostidis K (1989) Modern approach to the classification system of Cyanophytes, 4: Nostocales. Algol Stud 82:247–345
Lange K, Townsend CR, Matthaei CD (2015) A traitbased framework for stream algal communities. Ecol Evol. https://doi.org/10.1002/ece3.1822
Legendre P, Gallagher ED (2001) Ecologically meaningful transformations for ordination of species data. Oecologia 129:271–280
Legendre P, Legendre L (1998) Numerical ecology. Elsevier Science, Amsterdam
Legendre P, Bocard D, Peres-Neto PR (2005) Analyzing beta diversity: partitioning the spatial variation of community composition data. Ecol Monogr 75:435–450
Leibold MA, Holyoak M, Mouquet N, Amarasekare P, Chase JM, Hoopes MF, Holt RD, Shurin JB, Law R, Tilman D, Loreau M, Gonzalez A (2004) The metacommunity concept: a framework for multi-scale community ecology. Ecol Lett 7:601–613
Logue JB, Mouquet N, Peter H, Hillebrand H (2011) Empirical approaches to metacommunities: a review and comparison with theory. Trends Ecol Evol 26:482–491
Mackereth FYH, Heron JG, Talling J (1978) Water analysis: some revised methods for limnologist. Freshw Biol Assoc 36:1–120
Montag LFA, Winemiller KO, Keppeler FW, Leão H, Benone NL, Torres NR, Prudente BS, Begot TO, Bower LM, Saenz DE, Lopez-Delgado EO, Quintana Y, Hoeinghaus DJ, June L (2018) Land cover, riparian zones and instream habitat influence stream fish assemblages in the eastern Amazon. Ecol Freshw Fish 00:1–13
Oksanen JF, Blanchet G, Kindt R, Legendre P, Minchin PR, O´Hara RB, Simpson GL, Solymos MP, Stevens HH, Wagner H (2013) Vegan: community ecology package. R package version 2.0–9. http://cran.r-project.org/package=vegan
Padial AA, Ceschin F, Declerck SAJ, De Meester L, Bonecker CC, Lansac-Tôha FA, Rodrigues L, Rodrigues LC, Train S, Velho LFM, Bini LM (2014) Dispersal ability determines the role of environmental, spatial and temporal drivers of metacommunity structure. PLoS One 9:e111227
Passy S, Blanchet FG (2007) Algal communities in human-impacted stream ecosystems suffer beta diversity decline. Divers Distrib 13:670–679
Peres-Neto PR, Legendre P, Dray S, Borcard D (2006) Variation Partitioning of Species Data Matrices: Estimation and Comparison of Fractions. Ecology 87:2614–2625
Petsch DK, Schneck F, Melo A (2017) Substratum simplification reduces beta diversity of stream algal communities. Freshw Biol 62:205–213. https://doi.org/10.1111/fwb.12863
R Development Core Team (2014) R: a language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, Austria. http://www.R-project.org/. Accessed 11 Nov 2017
Roberto MC, Santana NF, Thomaz SM (2009) Limnology in the Upper Paraná River floodplain: large-scale spatial and temporal patterns, and the influence of reservoirs. Braz J Biol 69:717–725
Rodrigues L, Algarte VM, Siqueira NS, Machado EMN (2013) Fatores envolvidos na distribuição e abundância do perifíton e principais padrões encontrados em ambientes da planície de inundação. In: Schwarzbold A, Burliga AL, Torgan LC (eds) Ecologia do Perifíton. RiMA, Sao Carlos, pp 131–145
Ros J (1979) Práticas de ecologia. Barcelona: Ed. Omega, p 181
Round FE (1971) The taxonomy of the Chlorophyta, 2. J Br Phycol Soc 6:235–264
Schwarzbold A (1990) Métodos ecológicos aplicados ao estudo do perifíton. Acta Limnol Bras 3:545–592
Sládecková A, Sládecek V (1977) Periphyton as indicator of the reservoir water quality II—pseudo-periphyton. Arch Hydrobiol 19:176–191
Soininen J, McDonald R, Hillebrand H (2007) The distance decay of similarity in ecological communities. Ecography 30:3–12. https://doi.org/10.1111/j.0906-7590.2007.04817.x
Souza Filho EE, Stevaux JC (1997) Geologia e geomorfologia do complexo rio Baia, Curutuba, Ivinheima. In: Vazzoler AEAM, Agostinho AA, Hahn NS (eds) A Planície de Inundação do Alto Rio Paraná. EDUEM NUPÉLIA, Maringá, p 460
Stegen J, Freestone A, Crist T, Anderson M, Chase J, Comita L, Cornell H, Davies K, Harrison S, Hurlbert A, Inouye B, Kraft N, Myers J, Sanders N, Swenson N, Vellend M (2013) Stochastic and deterministic drivers of spatial and temporal turnover in breeding bird communities. Glob Ecol Biogeogr 22:202–212
Stevenson RJ (1996) An introduction to algae ecology in freshwater benthic habi-tats. In: Stevenson RJ, Bothwell ML, Lowe RL (eds) Algal ecology. Academic Press, San Diego, pp 3–30
Tapolczai K, Bouchez A, Stenger-Kovács C, Padisák J, Rimet F (2016) Trait-based ecological classifications for benthic algae: review and perspectives. Hydrobiologia 776:1–17
Thomaz SM, Bini LM, Bozelli RL (2007) Floods increase similarity among aquatic habitats in river-floodplain systems. Hydrobiologia 579:1–13. https://doi.org/10.1007/s10750-006-0285-y
Utermӧhl H (1958) Zur Vervollkommnung der quantitative phytoplankton-methodic. Mitt Int Ver Theor Angew Limnol 9:1–38
Weiher E, Freund D, Bunton T, Stefanski A, Lee T, Bentivenga S (2011) Advances, challenges and a developing synthesis of ecological community assembly theory. Philos Trans R Soc Lond B Biol Sci 366:2403–2413
Wetzel CE, de Bicudo DC, Ector L et al (2012) Distance decay of similarity in neotropical diatom communities. PLoS One 7:10–11. https://doi.org/10.1371/journal.pone.0045071
Xiao LJ, Wang T, Hu R et al (2011) Succession of phytoplankton functional groups regulated by monsoonal hydrology in a large canyon-shaped reservoir. Water Res 45:5099–5109. https://doi.org/10.1016/j.watres.2011.07.012
Acknowledgements
We thank the Coordination for the Improvement of Higher Education Personnel (CAPES) for granting K. L. Adame with a master scholarship, B. Dunck with a postdoctoral scholarship during the study development, and L. Colares also thanks to the Coordination for the Improvement of Higher Education Personnel (CAPES) for granting current master scholarships. We also thank the National Council for Scientific and Technological Development (CNPq) for granting L. Rodrigues with productivity support; the LongTerm Ecological Research (CNPq-PELD-Brazil) and the Research Center in Limnology, Ichthyology, and Aquaculture (Nupélia) for technical and logistical support during the conduction of this study.
Author information
Authors and Affiliations
Contributions
KLA and LR contributed to the study conception and design. KLA quantified the biological material. KLA, BD, and LC analyzed the results and wrote the manuscript. All authors reviewed and complemented the paper.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Adame, K.L., Colares, L., Rodrigues, L. et al. Dispersal increases beta diversity in periphytic algae communities of subtropical floodplain lakes. Braz. J. Bot 44, 273–285 (2021). https://doi.org/10.1007/s40415-020-00688-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40415-020-00688-3