
Abstract
Phosphorus (P) is one of the most important factors regarding plant growth and development. Its deficiency can be compensated by adding chemical fertilizers which are expensive and have an overall negative impact on the ecosystem. Their continuous use leads to the depletion of soil fertility, accumulation of toxic elements which will affect the normal soil micro flora, leading to an imbalance in equilibrium. Phosphate solubilizing microbes (PSM) may be an alternative way to overcome the P deficiency without causing any harm to environment. For this, their biodiversity, mode of action, colonizing ability and their careful relevance should be used as trustworthy components in sustainable agricultural systems. Due to variability in the performance of these microbes, their exploitation in the area of nanotechnology becomes a challenge in sustainable agriculture. In this review, we have discussed the use of PSM as biofertilizers, responsible factors and the strains that are used for the synthesis of different nanoparticles whose implementation will not only promote sustainable agriculture but also be helpful in commercial applicability.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The universal need to increase crop yields from a slowly diminishing and degrading soil resource had drawn significant attention to the agroecosystem around the world. In the coming years, increasing crop production for the growing population in all developing countries, particularly countries like India, will be a major challenge for agro-scientists. The most popular method for improved growth is the use of chemical fertilizers, which reflects a 50 percent boost in crop production, while the other half results in increased soil alkalinity, leading to loss of soil quality and environmental degradation (Lin et al. 2019). These chemically synthesized fertilizers contain heavy acids, like nitric acid, sulfuric acid and hydrochloric acid in their composition that upon accumulation tends to reduce the soil’s useful microbial population, increase soil acidity and ultimately interfere with the growth of plants. However, their continuous use may reduce the population of these beneficial microbes. Their adverse effects on the environment and increasing costs prompted the researchers to use alternate methods to increase crop production that would be environmentally sustainable and cost-effective. Implementations of plant growth-promoting microorganisms (PGPMs) may thus be seen as an effective way of minimizing the usage of chemical fertilizers within the agricultural boundaries (Dobbelaere et al. 2003).
Availability of phosphate in soil
Phosphorus (P) is an essential macronutrient known for biological and physiological development known for the cellular organization, stimulation of root development, formation of seed and in earlier maturation of crops. In nature, two forms of available P are present in the soil, viz., inorganic and organic phosphates. Mostly in some types of soil only 0.1 % is utilized by the plant (Beever and Burns 1980). Still, a large proportion of phosphates remains present in unavailable form (Fig. 1). This inorganic insoluble form of P is apatite, oxyapatite and hydroxyapatite. However, under favorable conditions, they can be solubilized and become available. Depending on the soil properties, phosphate anions (PO43−) are found to be extremely reactive and can be immobilized via precipitation with the Mg2+, Ca2+, Al3+, or Fe3+ cations. A huge amount of P when applied as fertilizer reacts with reactive cations in acidic soil and Ca2+ in calcareous or normal soils via precipitation reaction (Richardson et al. 2011). Richardson (2011) suggested that organic matter should also exist as the main reservoir (30%–80%) of immobilized P mostly in the form of orthophosphoric acid, phytin, inositol phosphates, phospholipids, nucleic acids, etc., degraded by the activity of enzymes like acid phosphatase, phytase and nucleases.

Ground phosphorus balance within the soil
Plant growth-promoting microorganisms (PGPM)
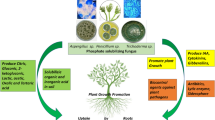
Agricultural activities now take more acceptable and environmentally sustainable approaches, such as the use of plant growth-promoting microbes (PGPMs), as the excessive use of chemical fertilizers and pesticides has adverse effects on the soil ecosystem as well as human health (Leach and Mumford 2011). Various important microbes have been reported to be associated within a plant's rhizospheric region which has beneficial effects on plant growth known as microbe-promoting plant growth (PGPM) (Mustafa et al. 2019). These positive effects are mediated by a series of mechanisms which includes enhancement of plant tolerance against abiotic and biotic stress, improvement in mineral nutrition, nitrogen fixation, phosphate solubilization, iron sequestration, root modification, as well as suppression of soil-borne diseases, synthesis of phytohormones, modulation of plant ethylene levels, and control of phytopathogenic microorganisms (Olanrewaju et al. 2017; Ramamoorthy et al. 2001; Ali et al. 2014; Chabot et al. 1996) as represented in Fig. 2. Microbial strains like Pseudomonas, Rhizobium, Acetobacter, Bacillus amyloliquefaciens, Vibrio proteolyticus, Xanthobacter agilis, Enterobacter aerogenes, E. asburiae, E. taylorae, Aspergillus flavus, Trichoderma harzianum, A. niger, Penicillium rugulosum, etc., were also identified as PGPM (Table 1). Specific bacterial rhizospheric strains such as Rhizobium, Pseudomonas and Bacillus are among the strongest strains with phosphorous solubilizing capacity (Sharma et al. 2013; Antoun and Pre´ Vost 2005). Kaur et al. (2015) stated that the combined inoculation of PSB with rock phosphate solubilization could serve as an alternative to the application of chemical fertilizers in sustainable agricultural systems.

Processes associated with the promotion of plant growth through microbial mediation by direct and indirect mode
Phosphate solubilizing microbes (PSM)
Various microbes such as bacteria, actinomycetes, fungi and algae are capable of solubilizing and mineralizing the P. A variety of rhizospheric bacteria that help to solubilize and mineralize immobilized P are Pseudomonas spp., Bradyrhizobium, Serratia, Agrobacterium spp., Bacillus circulans, Rhodococcus, Sinomonas, Salmonella, Thiobacillus (Chakraborty et al. 2009), Erwinia (Farajzadeh et al. 2012), Azatobacter (Kumar et al. 2012), Bacillus spp., (Raza et al. 2008), Paenibacillus (Rashid et al. 2012), Burkholderia (Badawi et al. 2011), Rhizobium (Kuckey 1983) whereas the fungal strains carrying this ability are Curvularia, Phoma, Alternaria, Glomus, Micromonospora, Arthrobotrys, Aspergillus, Helminthosporium, Oidiodendron, Rhizoctonia, Penicillium, Cephalosporium, Fusarium, Achrothcium, Cladosporium, Cunninghamella, Chaetomium, Mortierella, Sclerotium, Myrothecium, Paecilomyces and many more. Kucey (1983) reported that fungus is claimed to be stronger phosphate solubilizers as compared to bacteria because they do not loss their P dissolving activity when subculturing and can traverse long distances through their hyphen thus producing more acids such as lactic acid, citric acid, gluconic acid, oxalic acid, 2-ketogluconic acid, acetic acid and tartaric acid than bacteria. Recently, actinomycetes have also attracted special attention due to their P solubilizing ability, production of phytohormones, antibiotics and are capable of surviving in extreme conditions, e.g., Actinomyces, Micromonospora and Streptomyces (Sharma et al. 2013; Khan et al. 2007). It has been documented that Kushneria sinocarni, a halophytic bacterium isolated from stressed environments from the Daqiao saltern east coast of China, has also shown to exhibit P solubilization properties (Zhu et al. 2011).
Mechanisms of phosphorus solubilization
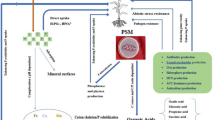
A number of the mechanism has been reported for P solubilization by rhizospheric bacteria and fungi. The ability of PSM to release metabolites like low molecular weight organic acids/inorganic acids (Seshachala and Tallapragada 2012), and decrease the pH (Alori et al. 2017a). A general sketch of P solubilization is represented in Fig. 3. Besides these acid systems, phosphate deprivation also activates the phosphate solubilization system (Gyaneshwar et al. 1999). In addition, releasing root exudates such as organic ligands helps in modification of P concentration within soil (Geelhoed et al. 1999). Consequently, the process responsible for mineralizing and solubilizing organic and inorganic phosphates is discussed as: -

Process of inorganic and organic phosphorus by solubilization and by mineralization means
Solubilization and mineralization of organic P
In soil, organic P constitutes around 35–50%, which is present either in the form of inositol phosphate (soil phytate) or in phosphodiester, phospholipids, nucleic acids, pesticides, antibiotics, detergents additives and flame retardants (Singh and Satyanarayana 2011). These forms are usually resistant toward chemical hydrolysis and thus require conversion into soluble form (Pi, HPO4−, H2PO4−). Halvorson et al. (1990) stated that sink phenomenon could be responsible, claiming that the value of P present in the biomass is significantly related to the decay of P present in the organic compounds. Different classes of enzymes such as phosphomonoesters (phosphatases), non-specific acid phosphatases (NSAPs) whose principal role is to dephosphorylate the phosphoanhydride or phosphodiester bond present in organic compounds.
Solubilization of inorganic phosphates
Organic acid production also plays an important role in inorganic P solubilization which involves pH reduction, enhance cation chelation, competition with P for adsorption sites or complex formation with metallic ions and insoluble P (Fe, Ca, Al). Trolove (2003) stated that the release of organic acids is represented by a direct oxidation mechanism via the pH decrease within the medium. The mechanism behind the production of these organic acids is either due to fermentation of sources presence of organic carbon (e.g., glucose, sucrose) or oxidative respiration (Kimberley et al. 2019). The organic acid formation within the rhizospheric area acidifies its surroundings that ultimately results in the release of PO4− ions (Goldstein and Liu 1987a). The prominent forms of acids are gluconic acid (Goldstein 1994), aspartic acid, lactic acid, tartaric acid (Bar-Yosef et al. 1999), citric acid and oxalic acid (Venkateswarlu et al. 1984) used for solubilization. It is also reported that the ability of phosphate solubilization was done by various methods like chelation /reduction processes and H2S gas production (Kim et al. 1997). Various studies have indicated that inorganic phosphate solubilization in the liquid media by bacterial and a fungal strain is due to carbon dioxide production. Solubilization of Fe+ and Al+ occurs via proton release mechanism in which adsorption of negatively charged P ions occurs. It is tested that proton release decreases P adsorption upon acidification because it increases H2PO4− in relation to HPO42− which have a higher affinity toward reactive soil surfaces. Studies show that it is through rhizospheric microbes having high affinity toward iron uptake system, releasing iron-chelating molecules, i.e., siderophores (Whitelaw 2000).
Genetic engineering of PSM
There is still little information about the genetics of phosphate solubilization (Altomare et al. 1999). However, certain genes involved in organic and mineral phosphate solubilization also need to be isolated and characterized. Microbial biotechnology, genetic engineering and molecular studies have created an entirely new way of modifying a number of rhizobacterial strains which improve their phosphate solubilizing ability. Insertion or over-expression of phosphate solubilizing genes in rhizospheric microbes is a novel approach for improving their ability to work as agricultural biofertilizers.
The gene cloning from the Gram-negative bacteria Erwinia herbicola was reported which encodes for the pyrroloquinoline quinone (PQQ) synthase gene involved in phosphate solubilization (mps). Clones containing those recombinant plasmids showed higher clearing halos with insoluble phosphate within the plates. These findings were the first study on the application of genetic modification to increase the performance of phosphate solubilization from microbial strains that can be used as biofertilizers (Goldstein and Liu 1987b). Additional studies were also carried out to investigate the function of these pqqE gene inhabiting rhizobacteria in the production of organic acid and phosphate solubilization. The pqqE successfully amplified and sequenced in genome of Pantoea sp. as well as Pseudomonas sp. Organic acids such as acetic, malic, gluconic, citric, and succinic acid were produced by these microbes. The effectiveness of phosphate solubilization was also tested by Pantoea sp. in the broth culture that was found as is 311 ± 4 and 204 ± 3 μg ml−1, while Pseudomonas sp. showed 176 ± 3 and 298 ± 5 μg ml−1 of phosphate that is solubilized. The amount of acetic acid and gluconic acid was also greater in the exudates of treated plants. Maximum value was also reported for chlorophyll quality, germination rate, leaf area index, grain weight and P content in plants inoculated with these organisms. In the field trials, grain yield was improved by 10–12 % over non-inoculated control. These findings lead us to the conclusion that organic acids were produced by the rhizobacteria inhabiting the pqqE gene and phosphate was solubilized in vitro and in vivo condition (Tahir et al. 2020).
Similarly, Kim et al. (1997) reported an increase in phosphatase enzyme activity of the recombinant strain was achieved when the gene of napA phosphatase from the rhizospheric bacterium Morganella morganii was transferred into Burkholderia cepacia IS-16 (Ishige et al. 2003). Several genes, such as ushA (nucleotidase), phoA (alkaline phosphatase), phyC (phytase), pst (Pi-specific transporter) and glpQ (glycerophosphoryldiester phosphodiesterase) involved in uptake of PO4− and solubilization of organic phosphate were stimulated during phosphate starvation (Kim et al. 1997; Ishige et al. 2003). Sequences of different phytase genes were also studied for optimal expression in Proteobacteria, synthesized and further engineered into the genomes of three different root-colonizing bacteria. The results showed that engineered strains showed high levels of phytate hydrolysis. Recently, Chryseobacterium sp., a phosphate solubilizing endophytic bacterial strain, shows plant growth promotion and biocontrol activities against Phytophthora blight and anthracnose on pepper using the draft genome sequence that includes genes linked to phosphate solubilization, plant growth promotion and biocontrol features (Fraga et al. 2001). These studies demonstrate that molecular approaches could be used to generate plant-associated strains with novel phosphate solubilizing capabilities (Jeong et al. 2018). Despite that, some hurdles like metabolic machinery dissimilarity and different regulating mechanism among donor and acceptor strains should be solved first to achieve successful gene introduction when using this approach. Last but not least, more studies are required to study better utilization of these PSMs under different environmental conditions and to make them favorable for farmers to boost agricultural production.
Parameters affecting phosphate solubilization
Carbon sources
The carbon source plays a very important role because of the heterotrophic nature of microorganism and of the fact that they solubilize insoluble phosphate by releasing organic acids. In the liquid medium, glucose was found to be the best energy source for microbes. PSM which can solubilize rock phosphate used various types of carbon-rich residues as sources of energy. The amount of soluble phosphate, however, varies with the form of source used for solubilization. Pseudomonas striata were reported to consume glucose, galactose, sucrose and arabinose as their energy while for Aspergillus awamori, the preferred carbon source was sucrose, followed by arabinose, glucose, mannitol, galactose, and xylose (Shulse et al. 2019). A fungus Penicillium digitatum, use glucose, sucrose, mannitol, arabinose, fructose, xylose and galactose and shows maximum P solubilization efficacy.
Nitrogen sources
Microorganisms require nitrogen for the synthesis of amino acids, proteins and nitrogen bases, which may occur either in the form of ammonium salts, nitrite and nitrate, or in the form of amino acids, depending on the type of enzyme produced. Strains of Pseudomonas striata utilized diverse forms of nitrogen sources except for nitrites (Rudresh et al. 2004). A bacteria Staphylococcus occidentalis can make use of nitrogen if present either in ammonium or amino-nitrogen forms as they lack in nitrate-reducing enzymes. Ammonium sulfate and ammonium nitrate were found as the best sources among all the nitrogen sources consumed by microbial strains.
Hydrogen ion concentration
Studies show that PSM 's behavior and growth are greatly influenced by pH shift within the liquid medium. Bacteria have a maximum solubilizing pH of 5.5–6 while fungal strains have a range of 5–6. The pH activity of fungal isolates was greatly reduced at pH 7 to 8. In several studies, it was reported that at pH 4.0, fungal strains show maximum solubilization of tri-calcium phosphate.
Temperature
Temperature plays a vital role in the growth and physiological activity of PSMs. Strains of P. striata shows appropriate growth at a range between 25 and 30° C that while for Aspergillus awamori and Penicillium digitatum, 30°C was found as the favorable optimum temperature for P solubilization. The behaviors of certain PSMs were substantially reduced at higher temperatures (35 °C and 40 °C). Wani et al. (2007) stated that 30 °C is a suitable temperature for tricalcium phosphate solubilization by P. digitatum, P. striata and A. Awamori whereas bacterium Bacillus polymyxa can solubilize at a range between 35 and 40°C. It was also observed that the culture medium was found a less acidic at higher temperature.
Aeration
Shake culture exhibited better solubilization of rock phosphate when compared to static culture. It has been noted often that S. occidentals and P. digitatum solubilized less rock phosphate under shake culture. Several studies suggest a persistent rise in phosphate solubilization up to 9–14 days. According to several reports, after 24 hours of incubation, Aspergillus niger and Pseudomonas fluorescence sp. solubilized TCP and reached at maximum on the 7th day and 14th day (Wani et al. 2007).
Humic substances
Humic substances also have an impact on phosphate solubilization. B. megaterium can solubilize insoluble TCP if fulvic acid and sodium humate are present. Thus, the mechanism of phosphate solubilization is directly proportional to the number of humic substances present within the soil. Humic substances behave as strong chelating agents, and this feature of humic substances is of great use increasing the effectiveness of phosphatic fertilizers, which are rendered insoluble via fixation mechanisms (Muratova et al. 2005).
Relevance of PSM as commercial bio-fertilizer
Implementation of chemical fertilizers made farmers satisfied with rising farm yields. Yet gradually there are negative impacts such as water contamination, leaching out, fauna and flora devastation which make crops more vulnerable to microbial infections leading to a decline in soil fertility causing irreparable harm to the biosphere. Then, there comes an existence of biofertilizer, a substance containing living microorganisms which help in plant growth promotion by either increasing the availability of the nutrients or by biological control. The primary biofertilizer sources are bacteria, fungi and cyanobacteria (blue-green algae) as they have a number of capacities that could be applied to enhance agricultural practices as microbes can fight diseases, degrade/convert complex soil compounds into their simpler forms that are further used by plants to develop and grow. This proves that these microbes have a beneficial role in increasing soil fertility by producing essential nutrients for the plant and soil (Souad et al. 2018).
An alternative strategy for the application of these PSMs as inoculants is there use in either in the form of mixed cultures or as co-inoculation in combination with other beneficial microorganisms. For example, the combined effect of microbial strains (Rhizobium+ Bacillus megaterium+ Trichoderma spp.) on nutrient uptake growth, and yield of chickpea showed an increase in nutrient uptake, the number of branches increased in plant height, yield of pea, nodulation and total biomass of chickpea when compared with the individual inoculations or control (Muratova et al. 2005). Co-inoculation of PSB isolates with rock phosphate promote the maize and wheat growth in terms of the height of shoot, dry biomass, the yield of grain and their total P uptake when compared with control (Quan et al. 2008).
In 2003, Nav Shakti Biotech developed a biofertilizer, Phosphobacteria which contains highly biopromoting PSB (Bacillus megaterium) that helps to secrete organic acids and making the unavailable phosphate into the soluble form. A biofertilizer, “Phylazonit-M” which contains a combination of three bacteria, viz., Bacillus megaterium + Azotobacter chrococcum, increases N and P supply to the plants. The primary role of these biofertilizers is to suppress the growth and activities of soil-borne pathogenic microbes in plants.
Since these microbes constitute a massive and crucial reservoir within microbiota, it can act as an innovative alternative to agricultural practices. Not only that, they also reflect the richest range of high biodiversity that helps to sustain biogeochemical cycles and food chains that can be used to grow and conserve food, manage pests, bioleaching, increase soil fertility, generate biofuels, control contaminants and serve as instruments for the therapeutic industry, such as antimicrobial agents, enzyme inhibitors, pesticides, immunomodulators/immunosuppressants and antitumor agents (Suleman et al. 2018; Fatima et al. 2015).
To solve the agricultural problems and to advance the crop biotechnology, there is an essential need to study the biodiversity of these beneficial microbes as the gene responsible for phosphate solubilization gives information related to their biochemical, evolutionary, taxonomical and metabolic and their regulatory networks. Ojuederie et al. (2017) reported that these microbes could also degrade toxic chemical pollutants and help in cleaning of polluted sites proving that these microbes act as self-sustaining and inexpensive ways. Further work is required to research the biodiversity of indigenous PSMs which can degrade various chemical contaminants and make the environment clean. Thus, more efforts have been made to characterize the microbial communities related to PSMs and their affective responses to chemical pollutants, acting as potential degraders and to identify those genes that are involved in particular degradation mechanism.
PSM performance as biocontrol agents
Phytopathogens born from soil are among the biggest issues in sustainable crop growth worldwide. To mitigate the harmful effects of pathogens on crop yields, enormous quantities of poisonous chemicals, particularly pesticides, are employed in conventional agronomic practices, which are incredibly harmful to the environment too (Ojuederie and Babalola 2017). The undesirability of implementing enormous levels of chemicals within the fields results in contamination, thus increasing resistance among soil phytopathogens. This lack of pathogen-resistant agricultural crops has compelled researchers to find alternatives to the issues with pesticides. To this end, biocontrol agents strategies consisting of biological formulations are considered the most promising choice for the use of costly destructive pesticides and the environment. In general, microorganisms with phosphate solubilizing ability were found to synthesize a broad range of metabolites with relevant biocidal and fungicidal capacities. The usage has both direct and indirect effects on plant growth and development. PSM controls pathogens by producing antimicrobial compounds such as phenolics compounds, PAL and flavonoids, antibiotics, cyanide hydrogen, indoleacetic acid (IAA), siderophores and several lytic enzymes, all of which suppress the growth of pathogens in plants (Saud and AL-Ahmadi (February 21st 2019; Köhl et al. 2019). The assessments of Streptomyces isolates against B. glumae (Zaidi et al. 2014) Trichoderma strains against phytopathogens (Suárez-Moreno et al. 2019) and Trichoderma harzianum isolate against Sclerotinia sclerotiorum in soybean (Bononi et al. 2020) were found as an effective biocontrol agent. However, Burkholderia cepacia, Citrobacter freundii, Enterobacter aerogenes, Klebsiella pneumonia, Proteus vulgaris and Acinetobacter lwoffii Alternaria alternate, Macrophomina phaseolina, Fusarium oxysporum and Sclerotium rolfsii were also posse’s biocontrol capacity (Zhang 2016) by exerting antagonistic activities against pathogens by synthesizing antimicrobial peptides, secreting lytic enzymes and inducing systemic resistance against pathogenic microbes (Ullah et al. 2018).
Effectiveness of PSM for enhancing crop production under stress
Due to increasing populations and rising food demands, it has now become vital to mitigate the adverse soil conditions in order to increase agricultural production and, ultimately, meet the food demands. Soil and its related environmental stresses not only impact the residual microbial population species and habitats but also the productivity and yield of plants. The primary stress on the soil is metal contamination, alkalinity/salinity and drought (Kang et al. 2015). Various methods are pursued to solve this problem, and one of those solutions includes leveraging this microbial capacity for the benefit of plants. Significant agronomic features of these PSMs are to produce exopolysaccharide (EPS) (Fatima et al. 2018, 2021), various phytohormones, 1-aminocyclopropane—1 carboxylate
(ACC) deaminase, volatile compounds, enhancement of nutrient uptake, induction of accumulation of osmolytes, upregulation or downregulation of the stress responsive genes or bringing about alterations in root morphology that exaggerate the improvement in soil properties, assist in plant growth, development and biocontrol activities (Meena et al. 2017). There are multiple studies on the enhancement of microbial-mediated plant growth under stress-free conditions while fewer studies are available on their impact under stressful environment.
It demonstrates the microbial ability to improve plant tolerance in stressed soils, and their effects as a mixture on various crops. The key idea here is to integrate the fact that PSM alliance can be used primarily in stressed soil, with the goal of rehabilitating soil conditions to promote crop productivity (Köhl et al. 2019). Bacillus, Pseudomonas spp. (Panwar et al. 2014) and Providencia rettgeri sp. Bradyrhizobium japonicum, (Jiang et al. 2018), Bacillus amyloliquefaciens (Han and Lee 2005), Bacillus megaterium (Shao et al. 2015) and Rhizobium (Thant and Aung 2018) were able to possess strong capacity for dissolving calcium phosphate under saline conditions Bacillus halodenitrificans and Hallobacillus sp. also showed more than 90 % higher root elongation and 17.4 % higher dry weight than untreated control wheat seedlings at 320 mM NaCl stress that suggests a substantial reduction in the detrimental effects of NaCl (Surange et al. 1997). The bacterial species of Pseudomonas fluorescens and Azospirillum brasilense also showed auxin production and inorganic phosphate solubilization under hypersaline conditions (Ramadoss et al. 2013), Pseudoalteromonas sp. showed halo-alkali and thermostable properties proved to have potential industrial activity (Kadmiri et al. 2018). However, Talaromyces aurantiacus and Aspergillus neoniger could survive in acidic environments and both fungi had a considerable ability to release soluble P by decomposing recalcitrant P-bearing compounds in alkaline conditions (Makhdoumi et al. 2015). Same results were found in the case of A. niger (Zhang et al. 2018) and A. terreus strains that had the potential to deal with salinity and drought stress (Li et al. 2015).
Interaction of exogenous phosphorus-solubilizing bacteria with rhizospheric microbial community
Phosphate solubilizing bacteria have made substantial use in the form of biological fertilizers (PSB). However, less understanding of its effect on the general microbial ecosystem has been shown. In several studies, a mixture of PSB was inoculated in conjunction with the manure fertilizer in tomato plant. The growth parameter findings showed that the mixture of PSB and compost could be significantly induced the growth of tomato. Adding PSB could significantly increase pH, available phosphate and many types of trace minerals, both in the rhizosphere and non-rhizosphere soil. The results of quantitative PCR and high-throughput sequencing showed that the rhizosphere-inoculated PSB did not become the dominant strain. However, the structure of the soil microbial species gets modified. Some indigenous bacteria, such as Pseudomonas, have declined in number, while several bacteria have expanded in the population, including members of Anaerolineaceae, Bacillus, Gemmationadaceae and Cytophagaceae. The PSB secreted low-molecular organic compounds and changed soil properties in order to dissolve inorganic phosphorus, which indirectly altered the root system's microbiota community. The results of the durability analysis showed that the soil's properties had a significant effect on the endogenous bacterial population (Liu et al. 2020).
Furthermore, in phosphorus poor soils or soils with rock phosphate, PSM associates well with arbuscular mycorrhizal fungi. Certain phosphatic ions that are accessed and transferred by the arbuscular mycorrhizal fungal hyphae to the plant may be produced by PSM from some of the more sparingly soluble phosphorus reservoirs. In addition, compared to non-mycorrhizal roots, PSM persists better around mycorrhizal roots and works symbiotically with AMF, contributing to increased plant growth, particularly where rock phosphate is added to the soil. In some cases where the soil is phosphate, combined dual inoculation of AMF and PSM has been shown to growth of plants more than inoculation of either microorganism individually. The simpler understanding of this finding is that in abundance, success and survival, the mycorrhizal endophyte may be stimulated. Increased nutrient absorption, especially phosphorus, is the main effect of this mycorrhiza in enhancing plant growth due to the discovery of the existing hyphae of the soil far beyond root-hair region where phosphorus gets reduced. In addition, by PSM inoculation, root exudation and plasticity can alter, which could also influence the growth of AMF. In the root region, the AMF also produces plant hormones and enhances the development of nitrogen-fixing species. However, for the optimum use of these interactions with regard to growth and development, a more comprehensive evaluation of the dynamics among soil microorganisms is required (Alori et al. 2017b).
Canavalia ensiformis, a forage legume popularly used as organic fertilizer, similar studies have been reported. An agronomically effective and sustainable solution applied to economically important crops would be the use of AMF and phosphorus solubilizers. Mycorrhizal colonization, phosphorus availability and ultimate phosphorus solubilizers will be promoted by the interaction of AMF and phosphorus solubilizers. An improvement of 3.07 mg/Kg of soluble P in the soil favored this interaction. The number of nodules varied from 46 to 76. The use of interactions between AMF and PSF increased mycorrhizal colonization, soil phosphorus availability and C growth (Padrón-Rodríguez et al. 2020). AMF and PSB combine synergistically when PSB solubilizes orthophosphate into sparingly accessible phosphorous compounds that can be consumed and transferred to the host plant by AMF. In conjunction with rock phosphate, the co-inoculation of KV with RI showed the biggest improvements in growing plants and tuber inulin quality relative to both unfertilized and fertilized controls. This strong cumulative effect may have had the opportunity for prospective farmers to use this association as a biofertilizer for inulin production in Helianthus tuberosus L (Nacoon et al. 2020).
In Chickpea (Cicer arietinum) several bio-inoculants are encouraged as natural “nutrient” resource. Bio-inoculants diazotroph Pseudomonas palleroniana and a phosphate solubilizer strain of Pseudomonas jesenii were evaluated for their plant growth-promoting potential. Furthermore, denaturing gradient gel electrophoresis (DGGE) and real-time PCR (qPCR) techniques were also implemented to analyze the microbial diversity and dynamics. Comparative agronomical and biochemical crop parameters revealed that organic soils co-inoculated with bio-inoculants and phosphate solubilizers have improvement in morphological and physiological content, respectively, over the control. Furthermore, microbial soil dynamics based on qPCR and DGGE showed the survival of strains until around the end of harvesting time without disrupting the indigenous microflora. Conclusively, their co-inoculation has demonstrated considerable promise for the growth and production of chickpeas under organically controlled soils and would also offer further motivation for their bio-formulation to progress toward zero budget natural farming (Rawat et al. 2019).
Performance of PSM in biosynthesis of metallic nanoparticles
As a strong technology, nanoscience enables us to look at the atomic and molecular levels and build structures on a nanometer scale. Nanotechnology offers agricultural productivity for the larger population of agriculture and food processing, and it promotes and frames the next stage of growth of genetically engineered plants, sustainable agriculture inputs, agricultural chemicals and advanced farming techniques of research and development (Jalili et al. 2020). In recent times, the nanomaterials have been developed using physical and chemical approaches. These methods are expensive and use harmful chemicals under ultraviolet irradiation, laser ablation and aerosol spray during the synthesis of nanoparticles. While such techniques are widely used, there is concern over the use of hazardous chemicals. (Prasad et al. 2017). Microbial synthesis of metallic nanoparticles is being investigated to overcome this problem. The physical solution of lower synthesis limits for the production of nanoparticles needs greater energy usage, and expenses are indeed very high. Industrial solutions, however, are low cost but include the use of hazardous solvents and a trace of chemical waste with hazardous by-product production (Thakkar et al. 2010). Biological approach that includes bacteria, fungi, yeast and viruses therefore provides high production, low-cost and environmentally safe nanoparticles synthesis (Narayanan and Sakthivel 2010).
Microbial nanoparticle synthesis would be a green chemistry method and is regarded an extremely interesting field for potential applications (Mandal 2006). Microbial production with enriched microbial biodiversity, using specific microbial cultivation, can be accomplished under cellular, biochemical and molecular processes, the rate of synthesis and improvement of nanoparticular properties. The relationship between microbial and metals is well recognized in biotechnological applications such as bioleaching, biomineralization, biocorrosion and bioremediation (Mohanpuria et al. 2008). The properties of microbially generated nanostructured mineral particles and metallic nanoparticles are close to those of chemically synthesized nanomaterials. Most unicellular and multicellular organisms, such as bacteria (prokaryotes) and fungi (eukaryotes), make inorganic materials either intracellular or extracellular (Mandal 2006; Gericke and Pinches 2006a). The intracellular processes include the movement of positive ions through the cell wall by contact with negative cell membrane ions. Furthermore, cell wall enzymes reduce the metal ion to nanoparticles, and later, these nanoparticles spread across the bacterial cell wall. The extracellular generation of nanoparticles in the case of fungi is the nitrate reductase-mediated production in the presence of enzyme nitrate, which helps to bio-reduce metal ions into nanoparticles (Duygu et al. 2019; Hulkoti and Taranath 2014). It provides a short description of recent PSM work involving in metallic nanoparticles synthesis.
The synthesis method (GS) of various metallic nanoparticles (MNPs) has reassessed microorganisms for their universal ability to turn heavy metal ions into neutral atoms at no cost to toxic chemicals. Microbial-mediated biosynthesis provides advantages of improved biocompatibility as compared to chemically synthesized MNPs and hence has greater potential for biological as well as agricultural applications. Microbial-mediated biosynthesis includes microorganisms such as fungi, bacteria and algae. They produce MNPs such as gold (Au), silver (Ag), zinc oxide (ZnO), iron (Fe2O3), nickel oxide (NiO) palladium (Pd), copper oxide (CuO), platinum (Pt) and magnesium oxide (MgO) that play a significant role as nanofertilizer (Mandal 2006), nanopesticides, nondiagnostic agents (Fatima et al. 2021), nano-induced resistance, etc. (Fatima et al. 2020). The tolerance of bacteria toward metal ions and formation of metal nanoparticles depends on physical and chemical growth parameters, such as particle size, pH, temperature, culture time, composition of growth medium and growth in light or dark conditions. PSM and different fungal species have also shown their abilities in the reduction of metal ions. However, not a thumb rule, most of the reducing agents sourced from living world also act as capping agents and render MNPs less toxic or more biocompatible. Various examples are given in Table 2.
Thus, in the field of agriculture, nanotechnology has been used to heighten the crop production with quality enrichment by improving farming systems. The emergence of biologically synthesized nanomaterials by PSM and their actions within the frame of sustainable agriculture have revolutionized world agriculture canvass dramatically by novelty, fast growth and enormity to meet the projection of global food demand (Singh and Prasad 2017). Therefore, further research is urgently needed to unravel the behavior and fate of these PSM in altered agriculture inputs and their interaction with biomacromolecules present in living systems and environments.
Conclusion
The inoculation of PSM in soil indicates to be a successful way to turn the insoluble P compounds into the plant-available P type, resulting in improved plant growth, crop production and performance. The most important P solubilizers for increasing the bioavailability of P in the soil are Bacillus, Pseudomonas, Rhizobium, Aspergillus, Penicillium, and AMF. By supplying readily absorbable P type, manufacturing plant hormones and antibiotics such as IAA and GA, PSM causes immediate crop production. PSM promotes plant growth by siderophore development and improves nitrogen fixation ability. In addition, through the synthesis of antibiotics, hydrogen cyanate (HCN), and antifungal metabolites, PSM acts as a disease control toward pathogenic bacteria. There are also possible replacements for inorganic phosphate fertilizers in order to satisfy plant P specifications, improving yields in sustainable farming. An economical and environmentally sustainable approach is their implementation. Further analysis, therefore, is critical in the field study to explore productive biofertilizers-PSM with several growth-stimulating attributes. The diverse form of PSMs has proved an important functional activity in agricultural sector as they constitute an important role by recycling the nutrients, pollutants degraders, waste treatment, a biocontrol agent and as an important source for the production of nanoparticles in the form of nanofertilizers that boost the agricultural crop production in the coming era. However, still there is an urgent need to study these environment-friendly microbes that can solve and show a new way to advance biotechnology. Despite knowing the significance of phosphate solubilizing microbes, still the information related to their diversity is very insufficient. Thus, their evaluation, exploration and exploitation are needed for scientific purposes as well as further work is urgently required to unravel the actions and fate of these PSMs in altered inputs from agriculture and their association with biomacromolecules present in living systems and ecosystems for the betterment of humankind.
References
Ahmad F, Ahmad I, Khan MS (2005) Indole acetic acid production by the indigenous isolates of azotobacter and flourescent Pseudomonas in the presence and absence of tryptophan. Turk J Biol 29:29–34
Ahmed E, Kalathil S, Shi L, Alharbi O, Wang P (2018) Synthesis of ultra-small platinum, palladium and gold nanoparticles by Shewanella loihica PV-4 electrochemically active biofilms and their enhanced catalytic activities. J Saudi Chem Soc 22:919–929
Ali S, Charles TC, Glick BR (2014) Amelioration of high salinity stress damage by plant growth-promoting bacterial endophytes that contain ACC deaminase. Plant Physiol Biochem 80:160–167
Alori ET, Glick BR, Babalola OO (2017) Microbial phosphorus solubilization and its potential for use in sustainable agriculture. Front Microbiol. https://doi.org/10.3389/fmicb.2017.00971
Alori ET, Glick BR, Babalola OO (2017a) Microbial phosphorus solubilization and its potential for use in sustainable agriculture. Front Microbiol 8:971
Altomare C, Norvell WA, Björkman T, Harman GE (1999) Solubilization of phosphates and micronutrients by the plant growth promoting and biocontrol fungus trichoderma harzianum rifai 1295–22. Appl Environ Microbiol 65:2926–2933
Antoun, H., Pre´ Vost, D., (2005) Ecology of Plant Growth Promoting Rhizobacteria, PGPR: Biocontrol and Bio fertilization. Springer, Dordrecht. 1–38
Asghar HN, Zahir ZA, Arshad M, Khaliq A (2002) Relationship between in vitro production of auxins by rhizobacteria and their growth-promoting activities in brassica junceal. Biol Fertil Soils 35:1–237
Badawi FSF, Amm B, Desoky AH (2011) Peanut plant growth and yield as influenced by co-inoculation with bradyrhizobium and some rhizomicroorganisms under sandy loam soil conditions. Ann Agric Sci 56:17–25
Bar-Yosef B, Rogers RD, Wolfram JH, Richman E (1999) Pseudomonas cepacia-mediated rock phosphate solubilization in kaolinite and montmorillonite suspensions. Soil Sci Soc Am J 63:1703–1708
Beever RE, Burns (1980) Phosphorus uptake, storage and utilization by fungi, advances in botanical. Research 8:127–219
Beveridge T, Murray R (1980) Sites of metal deposition in the cell wall of Bacillus subtilis. J Bacteriol 141:876–887
Bononi L, Chiaramonte JB, Pansa CC (2020) Phosphorus-solubilizing Trichoderma spp from Amazon soils improve soybean plant growth. Sci Rep 10:2858
Chabot R, Antoun H, Cescas MP (1996) Growth promotion of maize and lettuce by phosphate solubilizing rhizobium leguminosarum biovar phaseoli. Plant Soil 184:311–321
Chakraborty U, Chakraborty BN, Basnet M, Chakraborty AP (2009) Evaluation of ochrobactrum anthropi trs-2 and its talc based formulation for enhancement of growth of tea plants and management of brown root rot disease. J Appl Microbiol 107:625–634
Chakraborty J, Mallick S, Raj R (2018) Functionalization of extracellular polymers of pseudomonas aeruginosa n6p6 for synthesis of cds nanoparticles and cadmium bioadsorption. J Polym Environ 26:3097–3108
Chaudhary P, Fatima F, Kumar A (2020) Relevance of nanomaterials in food packaging and its advanced future prospects. J Inorg Organomet Polym 30:5180–5192
Dobbelaere S, Vanderleyden J, Okon Y (2003) Plant growth-promoting effects of diazotrophs in the rhizosphere. CRC Crit Rev Plant Sci 22:107–149
Duygu DY, Erkaya IA, Erdem B (2019) Characterization of silver nanoparticle produced by Pseudopediastrum boryanum (Turpin) E. Hegewald and its antimicrobial effects on some pathogens. Int J Environ Sci Technol 16:7093–7102
Egamberdiyeva D (2005) Plant growth promoting rhizobacteria isolated from a calsisol in semi arid region of uzbekistan: biochemical characterization and effectiveness. J Plant Nutr Soil Sci 168:94–99
Farajzadeh D, Yakhchali B, Aliasgharzad N, Bashir NS, Farajzadeh M (2012) Plant growth promoting characterization of indigenous azotobacteria isolated from soils in Iran. Curr Microbiol 64:397–403
Fatima F, Bajpai P, Pathak N, Singh S, Priya S, Verma SR (2015) Antimicrobial and immunomodulatory efficacy of extracellularly synthesized silver and gold nanoparticles by a novel phosphate solubilizing fungus Bipolaris tetramera. BMC Microbiol 15:52
Fatima F, Verma SR, Bajpai P, Pathak N (2016) Extracellular mycosynthesis of silver nanoparticles and their microbicidal activity. J Glob Antimicrob Re 7:88–92
Fatima F, Pathak N, Verma SR, Bajpai P (2018) Toxicity and immunomodulatory efficacy of biosynthesized silver myconanosomes on pathogenic microbes and macrophage cells. Artif Cells Nanomed Biotechnol 46:1637–1645
Fatima F, Siddiqui S, Khan WA (2020) Nanoparticles as novel emerging therapeutic antibacterial agents in the antibiotics resistant era. Biol Trace Elem Res 2020:1–13
Fatima F, Hashim A, Anees S (2021) Efficacy of nanoparticles as nanofertilizer production: A review. Environ Sci Pollut Res Int 28:1292–1303
Fayaz AM, Balaji K, Kalaichelvan PT (2009) Venkatesan R. Fungal based synthesis of silver nanoparticles—An effect of temperature on the size of particles. Colloids Surf B Biointerfaces 74:123–126
Fraga R, Rodriguez H, Gonzalez T (2001) Transfer of the gene encoding the Nap A acid phosphatase from Morganella morganii to a Burkholderia cepacia strain. Acta Biotechnol 21:359–369
Geelhoed JS, Riemsdijk WHVAN, Findenegg GR (1999) Simulation of the effect of citrate exudation from roots on the plant availability of phosphate adsorbed on goethite. Eur J Soil Sci 50:379–390
Gericke M, Pinches A (2006a) Biological synthesis of metal nanoparticles. Hydrometallurgy 83:132–140
Gericke M, Pinches A (2006b) Microbial production of gold nanoparticles. Gold Bull 39:22–28
Goldstein A. H: (1994). Involvement of the Quinoprotein Glucose Dehydrogenase In The Solubilization Of Exogenous Mineral Phosphates By Gram-Negative Bacteria, Phosphate In Microorganisms: Cellular And Molecular Biology, Washington, DC, 197–203
Goldstein AH, Liu ST (1987a) Molecular cloning and regulation of a mineral phosphate solubilizing gene from erwinia herbicola. Biotechnology 5:72–74
Gyaneshwar P, Pareka LJ, Archana G, Podle PS, Collins MD, Huston RA, Naresh KG (1999) Involvement of a phosphate starvation inducible glucose dehydrogenase in soil phosphate solubilization by enterobacter asburiae. FEMS Microbiol Lett 171:223–229
Halvorson HO, Keynan A, Kornberg HL (1990) Utilization of calcium phosphates for microbial growth at alkaline phosphate. Soil Biol Biochem 22:887–890
Han HS, Lee KD (2005) Physiological responses of soybean—inoculation of Bradyrhizobium japonicum with PGPR in saline soil conditions. Res J Agric Biol Sci 1:216–221
Hulkoti NI, Taranath T (2014) Biosynthesis of nanoparticles using microbes—A review. Colloids Surf B 121:474–483
Ishida K, Cipriano TF, Rocha GM, Weissmüller G, Gomes F, Miranda K, Rozental S (2014) Silver nanoparticle production by the fungus Fusarium oxysporum: nanoparticle characterization and analysis of antifungal activity against pathogenic yeasts. Mem Inst Oswaldo Cruz 109:220–228
Ishige T, Krause M, Bott M, Wendisch VF, Sahm H (2003) The phosphate starvation stimulon of Corynebacterium glutamicum determined by DNA microarray analy ses. J Bacteriol 185:4519–4529
Jalili B, Bagheri H, Azadi S (2020) Identification and salt tolerance evaluation of endophyte fungi isolates from halophyte plants. Int J Environ Sci Technol 17:3459–3466
Jeong JJ, Sang MK, Pathiraja D, Park B, Choi IG, Kim KD (2018) Draft genome sequence of phosphate-solubilizing Chryseobacterium sp. strain ISE14 biocontrol plant growth promoting rhizobacterium isolated from cucumber. Prokaryotes 6:12–18
Jiang H, Qi P, Wang T, Wang M, Chen M, Chen N, Pan L, Chi X (2018) Isolation and characterization of halotolerant phosphate-solubilizing microorganisms from saline soils. Biotech 8:461
Kadmiri IM, Chaouqui L, Azaroual SE (2018) Phosphate-solubilizing and auxin-producing rhizobacteria promote plant growth under saline conditions. Arab J Sci Eng 43:3403–3415
Kang SM, Radhakrishnan R, Lee IJ (2015) Bacillus amyloliquefaciens subsp. plantarum GR53, a potent biocontrol agent resists Rhizoctonia disease on Chinese cabbage through hormonal and antioxidants regulation. World J Microb Biot 31:1517–1527
Kaur G, Reddy M (2015) Effects of phosphate-solubilizing bacteria, rock phosphate and chemical fertilizers on maize-wheat cropping cycle and economics. Pedosphere 25:428–437
Khalid A, Arshad M, Zahir ZA (2001) Factor affecting auxin biosynthesis by wheat a rice rhizobacteria. Pak J Soil Sci 21:11–18
Khan MS, Zaidi A, Wani PA (2007) Role of phosphate-solubilizing microorganisms in sustainable agriculture - a review. Agron Sustain Dev 27:29–43
Kim KY, Mcdonald GA, Jordan D (1997) Solubilization of hydroxyapatite by enterobacter agglomerans and cloned escherichia coli in culture medium. Bio Fertil Soils 24:347–352
Kim DY, Kadam A, Shinde S, Saratale RG, Patra J, Ghodake G (2018) Recent developments in nanotechnology transforming the agricultural sector: a transition replete with opportunities. J Sci Food Agric 98:849–864
Kimberley D, Schneider JR, Thiessen M, Francis Z, Keith D, Tandra R, Derek DF, Lynch H, O’Halloran IP, Henry F (2019) Options for improved phosphorus cycling and use in agriculture at the field and regional scales. J Environ Quality 48(5):1247–1264
Köhl J, Kolnaar R, Ravensberg WJ (2019) Mode of action of microbial biological control agents against plant diseases: relevance beyond efficacy. Front Plant Sci 10:845
Kuckey (1983) Phosphate-solubilizing bacteria and fungi in various cultivated and virgin alberta soils. Canad J Soil Sci 63:671–678
Kumar P, Dubey RC, Maheshwari DK (2012) Bacillus strains isolated from rhizosphere showed plant growth promoting and antagonistic activity against phytopathogens. Microbiol Res 167:493–499
Leach AW, Mumford JD (2011) Pesticide environmental accounting: a decision-making tool estimating external costs of pesticides. J Verbrauch Lebensm 6:21–26
Li X, Luo L, Yang J, Li B, Yuan H (2015) Mechanisms for solubilization of various insoluble phosphates and activation of immobilized phosphates in different soils by an efficient and salinity-tolerant Aspergillus niger strain An2. Appl Biochem Biotech 175:2755–2768
Lin W, Lin M, Zhou H, Wu H, Li Z, Lin W (2019) The effects of chemical and organic fertilizer usage on rhizosphere soil in tea orchards. PLoS ONE 14:e0217018
Liu J, Qi W, Li Q (2020) Exogenous phosphorus-solubilizing bacteria changed the rhizosphere microbial community indirectly. Biotech 10:164
Makhdoumi A, Dehghani-Joybari Z, Mashreghi M (2015) A novel halo-alkali-tolerant and thermo-tolerant chitinase from Pseudoalteromonas sp. DC14 isolated from the Caspian Sea. Int J Environ Sci Technol 12:3895–3904
Mandal D (2006) The use of microorganisms for the formation of metal nanoparticles and their application. Appl Microbiol Biotechnol 69:485–492
Meena KK, Sorty AM, Bitla UM, Choudhary K, Gupta P, Pareek A, Singh DP, Prabha R, Sahu PK, Gupta VK, Singh HB, Krishanani KK, Minhas PS (2017) Abiotic stress responses and microbe-mediated mitigation in plants: the omics strategies. Front Plant Sci 8:172
Mohanpuria P, Rana NK, Yadav SK (2008) Biosynthesis of nanoparticles: technological concepts and future applications. J Nanopart Res 10:507–517
Mukherjee P, Ahmad A, Mandal D, Senapati S, Sainkar SR, Khan MI, Parishcha R, Ajaykumar PV, Alam M, Kumar R (2001) Fungus-mediated synthesis of silver nanoparticles and their immobilization in the mycelial matrix: a novel biological approach to nanoparticle synthesis. Nano Lett 1:515–519
Mukherjee P, Senapati S, Mandal D, Ahmad A, Khan MI, Kumar R, Sastry M (2002) Extracellular synthesis of gold nanoparticles by the fungus Fusarium oxysporum. Chem Bio Chem 3:461–463
Muratova AYU, Turkovskaya OV, Antonyuk LP, Makarov OE, Pozdnyakova LI, Ignatov VV (2005) Oil-oxidizing potential of associative rhizobacteria of the genus azospirillum. Microbiol 74:210–215
Mustafa S, Kabir S, Shabbir U (2019) Plant growth promoting rhizobacteria in sustainable agriculture: from theoretical to pragmatic approach. Symbiosis 78:115–123
Nacoon S, Jogloy S, Riddech N (2020) Interaction between phosphate solubilizing bacteria and arbuscular mycorrhizal fungi on growth promotion and tuber inulin content of helianthus tuberosus L. Sci Rep 10:4916
M. M. Naik , M. S. Prabhu , S. N. Samant , P. M. Naik and S. Shirodkar (2017), Thalassas: an International Journal of Marine Sciences, 33 , 73–80.
Nair B, Pradeep T (2002) Coalescence of nanoclusters and formation of submicron crystallites assisted by Lactobacillus strains. Cryst Growth Des 2:293–298
Narayanan KB, Sakthivel N (2010) Biological synthesis of metal nanoparticles by microbes. Adv Coll Interface Sci 156:1–13
Nicomrat D, Tharajak J, Kanthang P (2016) Microbial comparison in synthesizing gold nanoparticles for harvesting gold in wastewater system. Appl Mech Mater 848:52–55
Ojuederie OB, Babalola OO (2017) Microbial and plant-assisted bioremediation of heavy metal polluted environments: a review. Int J of Env Res Pub Health 14(12):1504
Olanrewaju OS, Glick BR, Babalola OO (2017) Mechanisms of action of plant growth promoting bacteria. World J Microbiol Biotechnol 33:197
Padrón-Rodríguez L, Arias-Mota RM, Medel-Ortíz R, de la Cruz-Elizondo Y (2020) Interaction with arbuscular mycorrhizal and phosphate solubilizer fungi in Canavalia ensiformis (Fabaceae). Botanical Sci 98:278–287
Panwar M, Tewari R, Nayyar H (2014) Microbial Consortium of Plant Growth-Promoting Rhizobacteria Improves the Performance of Plants Growing in Stressed Soils: An Overview. In: Khan M, Zaidi A, Musarrat J (eds) Phosphate Solubilizing Microorganisms. Springer, Cham
Prasad R, Bhattacharyya A, Nguyen QD (2017) Nanotechnology in sustainable agriculture: recent developments, challenges, and perspectives. Front Microbiol 8:1014
Quan X, Jian YL, Jian-Min I, HuoYan ZHOU, ChangWen WANG, Xiao QC (2008) Enhancement of phosphorus solubility by humic substances in ferrosols. Pedosphere 18:533–538
Raliya R (2013) Rapid, low-cost, and ecofriendly approach her for iron nanoparticle synthesis using Aspergillus oryzae TFR9. J Nanoparticles Article ID 141274:1–4
Raliya R, Biswas P, Tarafdar JC (2015) TiO2 nanoparticle biosynthesis and its physiological effect on mung bean (Vigna radiata L.) Biotechnol. Rep 5:22–26
Ramadoss D, Lakkineni VK, Bose P, Ali S, Annapurna K (2013) Mitigation of salt stress in wheat seedlings by halotolerant bacteria isolated from saline habitats. Springerplus 2:6
Ramamoorthy V, Viswanathan R, Raguchander T, Prakasanv SR (2001) Induction of systemic resistance by plant growth promoting rhizobacteria in crop plants against pests and diseases. Crop Protect 20:1–11
Rashid S, Charles TC, Glick BR (2012) Isolation and characterization of new plant growth promoting bacterial endophytes. Appl Soil Ecol 61:217–224
Rawat N, Sharma M, Suyal DC, Singh DK, Joshi D, Singh P, Goel R (2019) Psyhcrotolerant Bio-inoculants and Their Co-inoculation to Improve Cicer arietinum Growth and Soil Nutrient Status for Sustainable Mountain Agriculture. J Soil Sci Plant Nutr 19:639–647
Raza W, Yang W, Shen QR (2008) Paenibacillus Polymyxa: antibiotics, hydrolytic enzymes, and hazard assessment. J Plant Pathol 90:419–430
Richardson AE, Simpson RJ (2011) Soil microorganisms mediating phosphorus availability. Plant Physiol 156:989–996
Rudresh DL, Shivaprakash MK, Prasad RD (2004) Prasad: effect of combined application of Rhizobium, phosphate solubilizing bacterium and Trichoderma spp. on growth, nutrient uptake and yield of chickpea (cicer aritenium L.). Appl Soil Ecol 28:139–146
Mona Saud AL-Ahmadi (February 21st 2019). Pesticides, Anthropogenic Activities, and the Health of Our Environment Safety, Pesticides - Use and Misuse and Their Impact in the Environment, Marcelo Larramendy and Sonia Soloneski, IntechOpen,
Senapati S, Ahmad A, Khan MI, Sastry M, Kumar R (2005) Extracellular biosynthesis of bimetallic Au-Ag alloy nanoparticles. Small 1:517–520
Seshachala U, Tallapragada P (2012) Phosphate solubilizers from the rhizosphere of Piper nigrum L. in Karnataka. India Chil J Agric Res 72:397–403
Shao J, Xu Z, Zhang N, Shen Q, Zhang R (2015) Erratum to: contribution of indole-3-acetic acid in the plant growth promotion by the rhizospheric strain Bacillus amyloliquefaciens, SQR9. Biol Fertil Soils 51:331–331
Sharma SB, Sayyed RZ, Trivedi MH, Gobi TA (2013) Phosphate solubilizing microbes: sustainable approach for managing phosphorus deficiency in agricultural soils. Springerplus 2:587
Shulse CN, Chovatia M, Agosto C, Wang G, Hamilton M, Deutsch S, Yoshikuni Y, Blow MJ (2019) Engineered root bacteria release plant-available phosphate from phytate. Appl Environ Microbiol 85:01210–01219
Singh A, Prasad SM (2017) Nanotechnology and its role in agro-ecosystem: a strategic perspective. Int J Environ Sci Technol 14:2277–2300
Singh B, Satyanarayana T (2011) Microbial phytases in phosphorus acquisition and plant growth promotion. Physiol Mol Biol Plants 17:93–103
Souad Z, Ahmed AB, Lakhdar B (2018) Efficacy of Bacillus subtilis and Trichoderma harzianum combination on chickpea Fusarium wilt caused by F. oxysporum f. sp. ciceris. Arch Phytopathol Plant Protect 51:217–226
Srivastava N, Srivastava M, Mishra PK, Ramteke PW (2016) Application of ZnO nanoparticles for improving the thermal and ph stability of crude cellulase obtained from Aspergillus fumigatus AA001. Front Microbiol 7:514
Suárez-Moreno ZR, Vinchira-Villarraga DM, Vergara-Morales DI, Castellanos L, Ramos FA, Guarnaccia C, Degrassi G, Venturi V, Moreno-Sarmiento N (2019) Plant-growth promotion and biocontrol properties of three streptomyces spp isolates to control bacterial rice pathogens. Front Microbiol 10:290
Suleman M, Yasmin S, Rasul M, Yahya M, Atta BM, Mirza MS (2018) Phosphate solubilizing bacteria with glucose dehydrogenase gene for phosphorus uptake and beneficial effects on wheat. PloS one 13(9):e0204408
Surange S, Wollum AG, Kumar N (1997) Characterization of Rhizobium from root nodules of leguminous trees growing in alkaline soils. Can J Microbiol 43:891–894
Syed A, Ahmad A (2012) Extracellular biosynthesis of platinum nanoparticles using the fungus Fusarium oxysporum. Colloids Surf B Biointerfaces 97:27–31
Tahir M, Naeem MA, Shahid M, Khalid U, Farooq ABU, Ahmad N, Waqar A (2020) Inoculation of pqqE gene inhabiting Pantoea and Pseudomonas strains improves the growth and grain yield of wheat with a reduced amount of chemical fertilizer. J Appl Microbiol 129:575–589
Thakkar KN, Mhatre SS, Parikh RY (2010) Biological synthesis of metallic nanoparticles. Nanomed Nanotechnol Biol Med 6:257–262
Thant S, Aung NN (2018) Phosphate solubilization of Bacillus megaterium isolated from non-saline soils under salt stressed conditions. J Bacteriol Mycol Open Access 6:335–341
Trolove SN, Hedley MJ, Kirk GJD, Bolan NS, Loganathan P (2003) Progress in selected areas of rhizosphere research on p acquisition. Aust J Soil Res 41:471–499
Ullah A, Mushtaq H, Ali U (2018) Screening, isolation, biochemical and plant growth promoting characterization of endophytic bacteria. Microbiol Curr Res 2:62–68
Venkateswarlu B, Rao AV, Raina P, Ahmad N (1984) Evaluation of phosphorus solubilization by microorganisms isolated from arid soil. J Indian Soc Soil Sci 32:273–277
Vigneshwaran N (2007) Biological synthesis of silver nanoparticles using the fungus Aspergillus flavus. Mater Lett 61:1413–1418
Wani PA, Khan MS, Zaidi A (2007) Synergistic effect of the inoculation with nitrogen-fixing and phosphate-solubilizing rhizobacteria on performance of field-grown chickpea. J Plant Nutr Soil Sc 170:283–287
Whitelaw MA (2000) Growth promotion of plants inoculated with phosphate solubilizing fungi. Adv Agron 69:99–151
Zaidi A, Ahmad E, Khan MS (2014) Role of Phosphate-Solubilizing Microbes in the Management of Plant Diseases. In: Khan M, Zaidi A, Musarrat J (eds) Phosphate Solubilizing Microorganisms. Springer, Cham
Zhang F (2016) Biocontrol potential of Trichoderma harzianum isolate T-aloe against Sclerotinia sclerotiorum in soybean. Plant Physiol Biochem 100:64–74
Zhang Y, Chen FS, Wu XQ, Luan FG, Zhang LP, Fang XM (2018) Isolation and characterization of two phosphate-solubilizing fungi from rhizosphere soil of moso bamboo and their functional capacities when exposed to different phosphorus sources and pH environments. PLoS ONE 13:e0199625
Zhu FG, Qu L, Hong X, Sun X (2011) Isolation and characterization of a phosphate solubilizing halophilic bacterium Kushneria sp. Ycwa18 from DaQiao salt Ern on the coast of the yellow sea of china. Evid Based Complement Alternat Med 615032:1–6
Acknowledgements
The authors are highly thankful to Chancellor, Integral University for his support and encouragement.
Funding
No funding is received to carry out this work.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declares that they have no competing of interest.
Additional information
Communicated by Samareh Mirkia.
Rights and permissions
About this article

Cite this article
Fatima, F., Ahmad, M.M., Verma, S.R. et al. Relevance of phosphate solubilizing microbes in sustainable crop production: a review. Int. J. Environ. Sci. Technol. 19, 9283–9296 (2022). https://doi.org/10.1007/s13762-021-03425-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13762-021-03425-9