
Abstract
The solitary ascidian Styela plicata is an introduced species in harbors of temperate and tropical oceans around the world. The invasive potential of this species has been studied through reproductive biology and population genetics but no study has yet examined the microbial diversity associated with this ascidian and its potential role in host ecology and invasiveness. Here, we used 16S rRNA gene tag pyrosequencing and transmission electron microscopy to characterize the abundance, diversity and host-specificity of bacteria associated with 3 Mediterranean individuals of S. plicata. Microscopy revealed low bacterial abundance in the inner tunic and their absence from gonad tissues, while pyrosequencing revealed a high diversity of S. plicata-associated bacteria (284 OTUs from 16 microbial phyla) in the inner tunic. The core symbiont community was small and consisted of 16 OTUs present in all S. plicata hosts. This core community included a recently described ascidian symbiont (Hasllibacter halocynthiae) and several known sponge and coral symbionts, including a strictly anaerobic Chloroflexi lineage. Most recovered bacterial OTUs (79.6 %) were present in single S. plicata individuals and statistical analyses of genetic diversity and community structure confirmed high variability of bacterial communities among host individuals. These results suggest that diverse and variable bacterial communities inhabit the tunic of S. plicata, including environmental and host-associated bacterial lineages that appear to be re-established each host generation. We hypothesize that bacterial communities in S. plicata are dynamic and have the potential to aid host acclimation to new habitats by establishing relationships with beneficial, locally sourced bacteria.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
Ascidians, or sea squirts, are sessile marine invertebrates often found at high densities in harbors and marinas (Turner et al. 1997; Connell 2001; Cohen et al. 2005) and are among the most important marine invaders worldwide (Lambert 2002, 2007; Whitlatch and Bullard 2007). In fact, many cosmopolitan species rely on anthropogenic transport for long-distance dispersal as larval life spans are short, ranging from a few hours in brooders up to a week in oviparous ascidians (Svane and Young 1989). Although the rate of introduction of non-indigenous ascidians has been increasing over recent decades (Lambert 2007), some species relocated centuries ago and have now become ancient introductions whose origins are largely unknown (Lambert 2001; Pineda et al. 2011). While introductions are frequent in ascidians, only few species (e.g., Didemnum vexillum) have become invasive (Bullard et al. 2007; Lambert 2009; Stefaniak et al. 2012). Invasive ascidians are the focus of considerable research due to their threat and disruptive potential towards native communities and overall biodiversity (Lambert 2002, 2009). However, before becoming invasive, introduced species may reside in a given location for decades or centuries, until favorable shifts in local conditions trigger their spread and colonization of new habitats (Blackburn et al. 2011).
The study of ascidian introductions has increased in recent years and focused on acquiring the necessary knowledge to develop appropriate management plans that limit species introductions and their spread from current boundaries (McDonald 2004; Locke 2009; Lejeusne et al. 2011). Studies have targeted a wide range of subjects, including phylogeography and population genetics (López-Legentil et al. 2006a; Rius et al. 2008; Pineda et al. 2011), reproductive biology (Bourque et al. 2007; Shenkar and Loya 2008; Rius et al. 2009; Wong et al. 2011), and larval development and sensitivity (Vázquez and Young 1996; Vázquez and Young 2000; Thiyagarajan and Qian 2003; Bellas 2005; Bellas et al. 2001; Bennett and Marshall 2005; Rius et al. 2010; Pineda et al. 2012a). However, most ascidians do not form independent entities, instead establishing symbiotic associations with diverse microorganisms (termed holobionts), as has been observed in other marine invertebrates. The ecological implications of these microbial symbionts are not well characterized in ascidians, but may contribute to secondary metabolite production (Schmidt et al. 2005) and enhance host metabolism, as occurs in other invertebrate hosts (Mouchka et al. 2010; Webster and Taylor 2012).
In contrast to sponges and corals, microbial associates in ascidians are under-studied, although research in this field is increasing rapidly. These symbiotic microbes are commonly reported in the polysaccharide envelope (tunic) of the ascidian body, a region that surrounds the zooids and is separated from the filtration apparatus (branchial sac) and digestive system where prey microbes are captured and consumed. By far, the most frequently reported and stable association known is between ascidians from the family Didemnidae and the unicellular cyanobacteria Prochloron (Prochlorales) (Carpenter and Foster 2002; Hirose et al. 2004; Hirose and Maruyama 2004; Hirose et al. 2009; Lewin 1978; Münchhoff et al. 2007; Stam et al. 1985; Yokobori et al. 2006). While other bacteria have been reported in ascidians, only a few of these associations have been characterized using molecular approaches (Martínez-García et al. 2007, 2010, 2011; Tait et al. 2007; Münchhoff et al. 2007; López-Legentil et al. 2011; Erwin et al. 2011; Behrendt et al. 2012). Based on these studies, the most abundant bacterial symbionts in ascidians are affiliated with Proteobacteria, in particular Alpha- and Gammaproteobacteria (Tait et al. 2007; Martínez-García et al. 2007; Erwin et al. 2011). In addition, Archaea (Crenarchaeota) were recently reported for the first time in an ascidian host and shown to be involved in nitrification (Martínez-García et al. 2008). Thus, ascidian-associated microorganisms may also play a critical role in the metabolic needs of their host in a given habitat.
Microbial symbionts associated with marine invertebrates have also been implicated in the production of bioactive secondary metabolites that exhibit pharmaceutical properties and have potential biotechnological applications (Schmidt et al. 2005; Bontemps et al. 2010; Erwin et al. 2010). As sessile invertebrates, ascidians are easy prey for predators and are under constant competitive pressure for space, food and light. Consequently, most species have developed a range of mechanisms that include physical (e.g., tunic toughness, camouflage strategies) and chemical defenses (e.g., secondary metabolites) to ensure survival (Pisut and Pawlik 2002; Tarjuelo et al. 2002; López-Legentil et al. 2006b). Recent studies have shown that the animal does not always produce these defensive metabolites and that associated microorganisms can contribute to their synthesis (Schmidt et al. 2005). Such active participation in secondary metabolite production by microbial associates may enhance the chemical defenses of host ascidians, as well as aid in the processing of detrimental substances or the innocuous bioaccumulation of heavy metals. For introduced ascidians, these additional capabilities may provide the host with a competitive edge over native species. However, no study has investigated the microbial diversity in an introduced ascidian, a critical first step in characterizing the relative importance of microbial symbionts in the ecology and long-term establishment of host ascidians in new habitats.
Styela plicata (Lesueur, 1823) is a solitary, oviparous ascidian commonly introduced in harbors and marinas of warm and temperate oceans across the world. In fact, S. plicata has been present in all studied oceans for such a long time that recurrent colonization events and shuffling among populations have blurred the genetic signal needed to determine the origin of this species (Pineda et al. 2011). The successful introduction of S. plicata to new regions has been attributed to the capacity of this species to physiologically adapt to widely fluctuating environments, particularly to changes in temperature and salinity (Sims 1984; Thiyagarajan and Qian 2003; Pineda et al. 2012b). S. plicata can also tolerate highly polluted waters (Naranjo et al. 1996) and grows rapidly until reaching sexual maturity (Sabbadin 1957; Yamaguchi 1975; Sciscioli et al. 1978). The high genetic variability reported in S. plicata may also enable the species to rapidly adapt to new environments (Barros et al. 2009; Pineda et al. 2011). In addition, S. plicata or its associated microbes produce bioactive secondary metabolites, namely the modified octopeptide plicatamide in the hemocytes (Tincu et al. 2000; Tincu et al. 2003), which may contribute to anti-predatory defenses as organic extracts of the gonad deterred consumption by the Caribbean bluehead wrasse (Pisut and Pawlik 2002). These same features may also contribute to the invasive potential of the species although to date its distribution is still mostly restricted to artificial substrata (Pineda 2012, but see Valero-Jiménez et al. 2012).
In this study, we investigated bacterial community structure in 3 individuals of Styela plicata from the Western Mediterranean using 16S rRNA gene tag pyrosequencing and electron microscopy to provide the first characterization of bacteria associated with an introduced ascidian. We incorporated statistical analyses of the diversity, abundance and host-specificity of associated bacteria. In addition, the structure and composition of bacterial associates were assessed and compared to known environmental and host-associated microbes to examine the potential contribution of symbiotic microbes to the ecology and invasive potential of S. plicata.
2 Material and methods
2.1 Sample collection
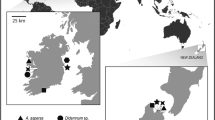
Four individuals of the solitary ascidian Styela plicata (Tunicata; Stolidobranchia; Styelidae) were collected in the harbor of Vilanova i la Geltrú, Spain (41° 12′ 53″ N, 1° 44′ 11″ E) by pulling up ropes (Fig. 1). Immediately after collection, each animal was carefully dissected. From one individual, we cut 2 mm2 pieces from the gonad and inner tunic (polysaccharide envelope) and fixed them separately in 2.5 % glutaraldehyde 2 % paraformaldehyde using filtered seawater as buffer for transmission electron microscopy (TEM). Samples were incubated in the fixative overnight at 4 °C, washed several times in filtered seawater and stored at 4 °C until processed. Gonad sections were examined to assess potential vertical transmission (i.e., parent-to-offspring passage) of bacterial symbionts, which if present would occur during gamete development within the gonads, since S. plicata is an oviparous species that releases sperm and eggs for external fertilization and larval development (Sciscioli et al. 1978; Pineda et al. in press). From the remaining three specimens (individuals SP1, SP2 and SP3), a 4 mm2 piece of the inner tunic (i.e., not in contact with ambient seawater) was carefully dissected with a sterile scalpel to avoid including epibionts or water-borne bacteria, washed several times with filtered seawater, and stored in 100 % ethanol until the DNA was extracted.

The introduced ascidian Styela plicata in the harbor of Vilanova i la Geltrú, NE Spain
2.2 Identification and phylogeny of host ascidians
Morphological identifications of S. plicata samples were complemented with phylogenetic analyses of ascidian 18S rRNA gene sequences, recovered from the eukaryotic data component following sequence sorting by Metaxa (see below). Sequences were processed with Geneious v5.6.3 (Drummond et al. 2012) and aligned using Clustal X (Thompson et al. 1997). To build phylogenetic trees, additional sequences were retrieved from GenBank (see accession numbers and codes in Fig. 2). Neighbor-joining (NJ) and maximum likelihood (ML) analyses were conducted in MEGA v5.05 (Tamura et al. 2011) considering gaps as full deletions. For NJ analyses, the Jukes-Cantor model of nucleotide substitution was used and data were re-sampled using 10,000 bootstrap replicates (Felsenstein 1985). The ML tree was built based on the GTR+I+G (Tavaré 1986) model with substitution rates varying among sites according to an invariant and gamma distribution and re-sampled using 1,000 bootstrap replicates. Bayesian phylogenetic analyses (BI) to calculate the posterior probabilities of branch nodes were conducted with MrBayes 3.1.2 (Ronquist and Huelsenbeck 2003), as implemented in Geneious using the GTR+I+G likelihood model. Monte Carlo Markov Chain length was set to 3 million generations with sampling every 500th generation and a burn-in value of 1,500. The average standard deviation of split frequencies reached values of less than 0.01 after 1,492,000 generations.

Phylogeny of partial 18S rRNA gene sequences from ascidians in the family Styelidae. Sequences obtained in this study are highlighted (bold lettering) and include sample name and code (in parenthesis). Labels on terminal nodes of reference sequences indicate the ascidian species and GenBank accession numbers. Tree topology was obtained from neighbor-joining (NJ) analysis. Individual bootstrap values from NJ and maximum likelihood (ML) analyses and posterior probabilities derived from Bayesian inference (PP) are located on the upper-left of the figure, corresponding to circled numbers on tree nodes. Scale bar represents 0.005 substitutions per site
2.3 Transmission electron microscopy
To construct resin blocks, samples were dehydrated in a graded ethanol series and embedded in Spurr’s resin at room temperature. Semi-thin (5 μm) and ultrathin sections (ca. 60 nm) were cut with a Reichert Ultracut microtome. Ultrathin sections were stained with uranyl acetate and lead citrate for ultrastructural observation (Reynolds 1963). TEM observations were conducted on a JEOL JEM-1010 (Tokyo, Japan) electron microscope coupled with a Bioscan 972 camera (Gatan, Germany). Resin blocks, ultrathin sections and TEM observations were performed at the Microscopy Unit of the Scientific and Technical Services of the University of Barcelona. For both gonad and inner tunic samples, twenty-five squares of the ultrathin sections (96 μm × 96 μm each) were carefully scanned for microbial cells.
2.4 DNA extraction and amplification
DNA extractions were performed using the Power Plant® DNA Isolation Kit (MoBio Laboratories, Carlsbad, CA) and a fragment of the 16S rRNA gene amplified with the primer set pyro926F (5′-AAA CTY AAA KGA ATT GRC GG-3′) and pyro1392R (5′-ACG GGC GGT GTG RC-3) complemented with adaptors B and A, respectively (Roche, Basel, Switzerland). Multiplex identifier (MID) barcodes unique to each sample were attached to the reverse primer. Total reaction volume was 50 μl, including 10 μL of 5×Buffer (containing 5 mM dNTPs and 15 mM MgCl2), 0.4 μL of BSA (10 mg/ml), 0.25 μL (1.25 units) of My Taq DNA Polymerase (Bioline®, London, United Kingdom), 0.2 μL of each primer (100 μM), ca. 10 ng of template DNA and sterile Milli-Q water. PCR conditions were as follows: 1 cycle at 95 °C for 1 min; 35 cycles at 95 °C for 30 s, 60 °C for 30 s, and 72 °C for 45 s, and a final elongation at 72 °C for 10 min. PCR products were visualized on 1 % agarose gels to assess amplification specificity and initial product quantity.
2.5 Pyrosequencing and data analysis
PCR products were sent to Macrogen, Inc. (South Korea) for purification, amplicon library construction and massively parallel 16S rRNA gene tag pyrosequencing using the Roche 454 GS-FLX Titanium system. Pyrosequencing data were deposited as flowgrams (sff file) in the Sequence Read Archive (SRA) of the National Center for Biotechnology Information (NCBI) under the accession number SRA056317. Sequence data from three S. plicata individuals presented herein (SP1 barcode = CGTGT, SP2 barcode = CTAGT, SP3 barcode = CTGAC) represent a portion of the multiplexed pyrosequencing run (1/16th plate). Raw sequence data were processed with stringent filtering and screening criteria to minimize the occurrence of spurious sequences and overestimation of microbial diversity (Huse et al. 2010; Schloss et al. 2011), using the mothur software package (Schloss et al. 2009). Sequences were reverse complemented and adaptor, MID and primer sequences were removed. Initial de-noising and quality filtering removed sequences that contained ambiguous base calls, long homopolymers (> 8 bp), barcode mismatches (> 1 bp), or primer mismatches (> 2 bp). Further, short sequences (<200 bp) and low quality reads (average quality scores less than 35 over 50 bp windows) were discarded. The remaining high-quality dataset was screened for non-target sequences (e.g. eukaryotic 18S rRNA, mitochondria, chloroplast) using Metaxa v1.1 (Bengtsson et al. 2011) to obtain a pool per individual containing only archaeal and bacterial 16S rRNA gene sequences. These sequences were aligned to the GreenGenes database and trimmed to an overlapping alignment space (449 bp). Putatively chimeric sequences were identified with self-reference searching using UChime (Edgar et al. 2011) in mothur and removed from the data set.
High quality sequences from all three S. plicata individuals (n = 791) were assigned to taxonomic groups using a naïve Bayesian classifier and bootstrap algorithm for confidence scoring (Wang et al. 2007), as implemented in mothur (kmer size = 8, iters = 100, cutoff = 60) using the improved GreenGenes taxonomy template (McDonald et al. 2012). Sequences were grouped into operational taxonomic units (OTUs) based on 97 % sequence similarity and mothur’s average neighbor algorithm. The taxonomic assignment of each OTU was constructed by majority consensus (Schloss et al. 2011). Sampling coverage and expected total OTU diversity were calculated using Good’s estimator (Good 1953) and the Chao1 estimator (Chao 1984).
OTU-independent assessments of genetic diversity and differentiation among the bacterial communities in replicate S. plicata hosts were conducted using nonparametric tests of homogeneity of variance (HOMOVA) and analysis of molecular variance (AMOVA; Stewart and Excoffier 1996). In addition, approximate maximum-likelihood trees were constructed in FastTree v2 (Price et al. 2010) and used to assess the distribution of unique lineages and differences in phylogenetic structure of bacterial communities among host individuals, using lineage-sorting (phylogenetic, or P) tests (Martin 2002) and the unweighted UNIFRAC algorithm (Lozupone et al. 2007). OTU-independent statistical analyses were conducted as implemented in the mothur software package.
3 Results
3.1 Host identification and phylogeny
Partial 18S rRNA gene sequences (430 to 434 bp) obtained for the three pyrosequenced individuals of S. plicata (SP1, SP2 and SP3) were identical. The topologies of the ML, NJ and BI trees were congruent so only the phylogeny obtained with NJ is depicted (Fig. 2). All species of Styela (S. montereyensis, S. plicata, S. gibbsii and S. clava) formed a monophyletic and well-supported clade (>93 support in all cases). In fact, all Styela sequences were identical. Two species in the genus Cnemidocarpa formed the most closely related, sister group to the Styela clade (Fig. 2).
3.2 Bacterial ultrastructure
TEM examination revealed that bacterial cells were randomly distributed and extremely rare in the inner tunic of Styela plicata, averaging ca. 35 cells per mm2. All bacterial morphotypes were ovoid to rod-shaped cells (ca. 0.2 μm × ca. 1 μm) always observed in complete isolation and never forming groups or aggregations (Fig. 3a and b). No bacteria were observed interacting with animal cells. The cytoplasmic membrane of bacterial cells appeared to be enclosed by a thin wall and encased riboplasm and nucleoplasm, without a distinct central nucleoplasmic area (Fig. 3d). Peripheral cell regions were rugose, typical of Gram-negative bacteria, but granulated, thus outer membranes were indistinct. No bacterial cells were observed in gonad sections.

Transmission electron microscopy images of bacteria observed in the inner tunic of the solitary ascidian Styela plicata. a Bacterial cell in the tunic, b transverse section, c longitudinal section of a bacterium and, d ultrastructural detail of its mid-region showing a bacterial wall (w), cytoplasmatic membrane (m) and riboplasm (rb)
3.3 Bacterial diversity, structure and host-specificity
The microbiota associated with S. plicata exhibited high diversity, comprised of 284 microbial OTUs (97 % sequence identity). Bacterial OTUs dominated the recovered microbial community, accounting for nearly all OTUs (99.3 %, n = 282), and corresponded to 12 described bacterial phyla and 3 candidate bacterial phyla (Fig. 4). The domain Archaea was represented by only 2 OTUs, each comprised of a single sequence tag (i.e., singleton OTUs) and corresponded to Crenarchaeota (Nitrosopumilus and an unidentified lineage, pMC2A209). High sampling coverage estimates (77.1 %) indicated that the recovered OTUs account for the majority of microbial diversity in S. plicata-associated communities. However, total richness estimates (Chao1) predicted over twice the number of OTUs than represented in our samples (n = 654, confidence interval = 527–848), due to a high prevalence of singleton OTUs (63.7 % of OTUs, n = 181). In fact, rare OTUs (<1 % relative abundance) accounted for the vast majority of microbial OTUs (93.6 %, n = 264) with only 5 OTUs having greater than 3 % relative abundance.

Phylum level microbial composition (97 % OTUs) in three individuals of Styela plicata. Proteobacteria OTUs are further divided into classes: Alpha- (α), Beta- (β), Gamma- (γ) and Delta- (δ) and unclassified (uncl.) Proteobacteria. Other = phyla represented by less than 3 sequence tags (Cladithrix, Crenarchaeota, GN04, OP3, Tenericutes)
Comparison of the microbial communities associated with replicate S. plicata individuals revealed similar bacterial composition at the phylum level among hosts. Seven dominant phyla were recovered in all S. plicata individuals and accounted for over 89 % of microbial OTUs per host (Fig. 4). Proteobacteria was the most dominant phylum, accounting for 25.4–50.0 % of OTUs per individual, and consisted primarily of taxa affiliated with Alphaproteobacteria and Gammaproteobacteria, recovered from all host individuals. Deltaproteobacteria and Betaproteobacteria sequences were less common and only recovered in 2 of 3 host individuals. Bacteroidetes was the second most common phylum (19.5–32.7 % of OTUs per host) and included numerous OTUs from the classes Bacteroidia (n = 5), Flavobacteria (n = 30) and Sphingobacteria (n = 33). Planctomycetes represented the third most common microbial phylum (9.8–12.8 % of OTUs per host) and encompassed the nominal classes Phycisphaerae (n = 2) and Planctomycea (n = 19) and the candidate classes: agg27 (n = 7), FFCH393 (n = 1), PW285 (n = 1), and vadinHA49 (n = 1). Sequences affiliated with the phyla Cyanobacteria, Actinobacteria, Firmicutes and Chloroflexi were also present in all S. plicata individuals, but exhibited lower OTU diversity (each accounting for <8 % of OTUs per host).
Despite the broad, phylum-level similarity in microbial communities among S. plicata hosts, most microbial OTUs were present in only a single host individual. Accordingly, OTU-independent statistics revealed significant differences in symbiont structure among the three S. plicata hosts, including genetic diversity, genetic differentiation, phylogenetic lineage distribution and community structure (Table 1). The core symbiont community of S. plicata (i.e., OTUs shared by all host individuals) was comprised of 16 OTUs (Fig. 5). Core OTUs accounted for less than 6 % of all OTUs but were disproportionately abundant, representing 22.0–38.5 % of all bacterial sequence tags. An additional 42 OTUs were shared between 2 of the 3 individuals, while the majority of OTUs (79.6 %, n = 226) were recovered in a single S. plicata individual (Fig. 5), often representing singleton OTUs (80.1 %, n = 181). To assess whether the prevalence of singleton OTUs (63.7 % of all OTUs) was driving the differentiation of bacterial communities among hosts, the genetic and phylogenetic metrics were repeated with singleton OTUs removed from the dataset. Significant differences among hosts were retained for all metrics, with identical results for all pairwise comparisons except for two individuals (SP1 and SP3; Table 1).

Specificity of microbial OTUs in three individuals of Styela plicata (SP1, SP2 and SP3). Pie charts depict the percentage of total sequence tags by host and numbers denote total 97 % OTUs. Black partitions show the core microbial OTUs present in all host individuals. Gray, light gray and barred portions represent OTUs shared between two host individuals. White portions depict OTUs present in a single host individual
Additional analysis of core OTUs revealed high similarity of S. plicata-associated bacteria sequences to known environmental and invertebrate-associated microbes (> 99 % sequence identity, Table 2). Most of the 16 core OTUs matched to seawater (n = 5) or sediment-derived (n = 5) bacteria, including the widespread genera Bacillus and Prochlorococcus, while the remaining core OTUs matched to host-associated bacteria from sponges (n = 2), corals (n = 2), algae (n = 1) and other ascidians (n = 1). Notably, the second most abundant core OTU was affiliated with a strictly anaerobic lineage of Chloroflexi (Anaerolineae).
4 Discussion
This study represents the first investigation of bacteria associated with a widespread introduced ascidian. We examined three individuals of Styela plicata and confirmed their identification and phylogenetic placement by 18S rRNA gene sequence analysis. A combined approach of electron microscopy and pyrosequencing of 16S rRNA gene tags revealed a low abundance yet high diversity of bacteria residing inside this introduced ascidian, consisting of 284 OTUs from 16 microbial phyla. The comparison of three S. plicata individuals revealed conserved microbial composition across hosts at a broad taxonomic level but OTU-level comparisons showed that only a small fraction of microbial OTUs were present in all hosts. Accordingly, statistical analyses confirmed significant differences in the genetic diversity, genetic differentiation and community structure of bacteria among the three host individuals. These results suggest that a wide range of bacteria can establish associations with S. plicata and that these communities are highly variable among individual hosts in the same population.
The phylogenetic signature of bacterial communities associated with S. plicata was conserved at the phylum level across individual hosts and consistent with previous studies of ascidian-associated bacteria. The dominant bacterial taxa in the ascidians Botryllus schlosseri, B. violaceus and Didemnum sp. from the Western Atlantic (Tait et al. 2007), Cystodytes dellechiajei from the Mediterranean (Martínez-García et al. 2007 ) and Didemnum sp. from the Gulf of Mexico (Erwin et al. 2011) were Alphaproteobacteria, Gammaproteobacteria and Bacteroidetes which were also prevalent in the microbiota of S. plicata herein. Additional bacterial phyla present in S. plicata and observed in other ascidians, though less common, were Firmicutes (Moss et al. 2003; Tait et al. 2007; Erwin et al. 2011) and Planctomycetes (Tait et al. 2007; Erwin et al. 2011), the latter of which was particularly abundant and diverse in S. plicata.
In addition to the presence of common ascidian-associated bacterial phyla, S. plicata also hosted bacterial phyla previously reported as rare or absent from the ascidian microbiota. To date, only one published study has utilized pyrosequencing technology to examine the bacteria associated with an ascidian, focusing on the internal and surface microbes associated with the didemnid Lissoclinum patella (Behrendt et al. 2012). Sequences affiliated with Actinobacteria were characterized as rare (0.08–0.29 % of the total community) in the cloacal cavity of the ascidian host, while Chloroflexi were only found on the underside of L. patella colonies (Behrendt et al. 2012). In S. plicata, Actinobacteria and Chloroflexi were both common taxa in the bacterial communities inhabiting the inner tunic of all three S. plicata hosts. The former phylum is of considerable interest, due to the diverse array of bioactive secondary metabolites produced by this bacterial group (Bull and Stach 2007), while the latter is a common component of the sponge microbiota (Schmitt et al. 2011). Less common taxa in S. plicata hosts (present in 1 or 2 individuals) but new to the ascidian microbiota included Acidobacteria, Tenericutes and several candidate phyla (SBR1093, GN04, OP3). The sampling depth afforded by pyrosequencing allows for greater coverage and fuller characterization of complex microbial communities (e.g., Webster et al. 2010), including unprecedented access to extremely rare community members (i.e., the rare biosphere; Sogin et al. 2006). The ecological significance of microbes in the rare biosphere remains uncertain (Pedrós-Alió 2012), although recent experimental evidence suggests that rare microbial taxa can drive important biogeochemical processes (e.g., sulfate reduction) despite their low abundance in environmental assemblages (Pester et al. 2010), and may increase in abundance in response to specific environmental conditions (Sjöstedt et al. 2012). Similar mechanisms for ecologically relevant contributions by rare taxa may exist in host-associated microbial communities and may be of particular importance for widespread host species like S. plicata that thrive in diverse marine environments. Further applications of next-generation sequencing technology to the ascidian microbiota will continue to expand our knowledge of the diverse and so far poorly understood bacterial communities inhabiting ascidian hosts.
The identification and study of core microbial symbionts are of particular interest due to the ecological relevance and potential functionality of microbial taxa that consistently occur in specific habitats (Shade and Handelsman 2012). The core symbiont communities of S. plicata identified herein represented a mixed assemblage of environmental and host-associated bacteria. While some core symbionts were bacteria common in seawater and thus facultative symbionts (e.g., genus Prochlorococcus), others represented host-associated bacteria and putatively obligate symbionts, such as a sponge-associated Chloroflexi. The presence of bacteria associated with specialized host habitats (e.g., marine sponges and corals) in S. plicata suggests that the tunic microhabitat can support bacterial lineages adapted to host-associated lifestyles. Whether host ascidians also benefit from metabolic activities of these symbiont lineages, as observed in sponge (Thacker and Freeman 2012; Fan et al. 2012) and coral (Mouchka et al. 2010) hosts, remains to be resolved. In addition, the presence of a strictly anaerobic lineage (Chloroflexi, Anaerolineae) in the core microbiota of S. plicata provides evidence for anaerobic microhabitats in the inner ascidian tunic that can support anaerobic metabolism. Indeed, a previous study documented anaerobic conditions in ascidian hosts (Diplosoma spp.) and predicted the proliferation of anaerobic microbes and processes in these microhabitats (Kühl and Larkum 2002). More recently, anaerobic conditions were documented in the cloacal cavity of Lissoclinum patella where resident microbial communities included facultative and obligate anaerobic species (Behrendt et al. 2012). Thus, host ascidians appear to have a range of microhabitats with variable physical and chemical conditions capable of hosting diverse bacterial taxa, including aerobic and anaerobic microbes, as well as facultative symbionts and those adapted to host-associated lifestyles.
Electron microscopy allowed for the visualization of the bacteria in the inner tunic of S. plicata, restricted to a few isolated, ovoid to rod-shaped cell morphotypes, and suggested their absence in reproductive tissues. The abundance of bacterial cells in the inner tunic of S. plicata was extremely low (35 cells per mm2) compared with microbial communities in other invertebrate hosts, for example, the sponge Haliclona caerulea (2 × 103 cells per mm2, Maldonado 2007). Low symbiont densities have been previously reported in ascidians (e.g., Turon et al. 2005) and may result from active host regulation of bacterial communities by phagocytic ‘harvesting’ of symbiont cells for nutritional gains (e.g., Martínez-García et al. 2007). While the identification of microbial cells from electron microscopy is problematic, the size and morphology of some bacterial cells in S. plicata were consistent with a newly described bacterium, Hasllibacter halocynthiae, isolated from the solitary ascidian Halocynthia roretzi (Kim et al. 2012). Further, a core symbiont OTU identified in S. plicata was identical to Hallibacter halocynthiae, suggesting that this newly described bacterium, which represents a new genus in the family Rhodobacteraceae, may be widespread among solitary stolidobranch ascidians. Future studies (e.g., fluorescence in situ hybridization analyses) are required to confirm this finding and provide additional data on the abundance of specific symbiont taxa. The absence of bacteria in the gonads of S. plicata is a first indication that vertical transmission (i.e., direct parent-to-offspring passage) of bacteria is not a strategy employed by this species, although transfer of bacterial symbionts to the progeny has often been observed in colonial ascidian hosts (Hirose 2000; Groepler and Schuett 2003; Moss et al. 2003; Hirose and Fukuda 2006; Martínez-García et al. 2007; Tait et al. 2007; Kojima and Hirose 2010, 2012). The acquisition of bacteria from the environment (i.e., horizontal transmission) appears to be the predominant means of establishing microbial communities in S. plicata, a strategy that may allow for local sourcing of bacterial symbionts across the broad geographic and environmental range of this widespread ascidian. Future studies that target S. plicata individuals from distant populations and distinct environments are required to appropriately address this hypothesis.
The primary objective of the current study was to characterize the bacterial communities associated with the introduced ascidian S. plicata, a critical first step in assessing the potential for these symbiotic relationships to contribute to their hosts’ successful establishment in new habitats. The widespread distribution and prevalence of S. plicata populations in artificial marine habitats has prompted numerous studies on the factors that enable this species to rapidly adapt to new environmental conditions. Here, we show that diverse bacterial communities inhabit S. plicata and hypothesized that the functional capabilities underlying this diversity may contribute to the high tolerance of the host. Further, high variability among individuals and apparent horizontal transmission of microbes suggest that dynamic bacterial communities inhabit S. plicata and that this ascidian has the ability to establish new (and potentially beneficial) symbiotic relationships sourced from local environments with each host generation. Additional study of the biological interactions between S. plicata hosts and their associated bacterial communities is now required to fully assess the ecological implications of these associations and their contribution to host acclimation and survival in new environments.
References
Barros RC, Rocha RM, Pie MR (2009) Human-mediated global dispersion of Styela plicata (Tunicata, Ascidiacea). Aquat Invasions 4(1):45–57
Behrendt L, Larkum AWD, Trampe E, Norman A, Sorensen SJ, Kühl M (2012) Microbial diversity of biofilm communities in microniches associated with the didemnid ascidian Lissoclinum patella. ISME J 6(6):1222–1237
Bellas J (2005) Toxicity assessment of the antifouling compound zinc pyrithione using early developmental stages of the ascidian Ciona intestinalis. Biofouling 21:289–296
Bellas J, Vázquez E, Beirasa R (2001) Toxicity of Hg, Cu, Cd, and Cr on early developmental stages of Ciona intestinalis (Chordata, Ascidiacea) with potential application in marine water quality assessment. Water Res 35:2905–2912
Bengtsson J, Eriksson KM, Hartmann M, Wang Z, Shenoy BD, Grelet G-A, Abarenkov K, Petri A, Rosenblad MA, Nilsson RH (2011) Metaxa: a software tool for automated detection and discrimination among ribosomal small subunit (12S/16S/18S) sequences of archaea, bacteria, eukaryotes, mitochondria, and chloroplasts in metagenomes and environmental sequencing datasets. Anton Leeuw 100:471–475
Bennett CE, Marshall DJ (2005) The relative energetic costs of the larval period, larval swimming and metamorphosis for the ascidian Diplosoma listerianum. Mar Freshw Behav Physiol 38(1):21–30
Blackburn TM, Pysek P, Bacher S, Carlton JT, Duncan RP, Jarosik V, Wilson JRU, Richardson DM (2011) A proposed unified framework for biological invasions. Trends Ecol Evol 26(7):333–339
Bontemps N, Bry D, López-Legentil S, Simon-Levert A, Long C, Banaigs B (2010) Structures and antimicrobial activities of pyridoacridine alkaloids isolated from different chromotypes of the ascidian Cystodytes dellechiajei. J Nat Prod 73:1044–1048
Bourque D, Davidson J, MacNair GN, Arsenault G, LeBlanc AR, Landry T, Miron G (2007) Reproduction and early life history of an invasive ascidian Styela clava Herdman in Prince Edward Island, Canada. J Exp Mar Biol Ecol 342:78–84
Bull AT, Stach JEM (2007) Marine actinobacteria: new opportunities for natural product search and discovery. Trends Microbiol 15(11):491–499
Bullard SG, Lambert G, Carman MR, Byrnes J, Whitlatch RB, Ruiz G, Miller RJ, Harris L, Valentine PC, Collie JS, Pederson J, McNaught DC, Cohen AN, Asch RG, Dijkstra J, Heinonen K (2007) The colonial ascidian Didemnum sp. A: current distribution, basic biology and potential threat to marine communities of the northeast and west coasts of North America. J Exp Mar Biol Ecol 342:99–108
Carpenter EJ, Foster RA (2002) Marine cyanobacterial symbioses. In: Rai AN, Bergman B, Rasmussen U (eds) Cyanobacteria in symbiosis. Kluwer Academic Publishers, Netherlands, pp 11–17
Chao A (1984) Nonparametric estimation of the number of classes in a population. Scand J Stat 11:265–270
Cohen AN, Harris L, Bingham BL, Carlton JT, Chapman JW, Lambert CC, Lambert G, Ljubenkov JC, Murray SN, Rao LC, Reardon K, Schwindt E (2005) Rapid Assessment Survey for exotic organisms in southern California bays and harbors, and abundance in port and non-port areas. Biol Invasions 7:995–1002
Connell SD (2001) Urban structures as marine habitats: an experimental comparison of the composition and abundance of subtidal epibiota among pilings, pontoons and rocky reefs. Mar Environ Res 52:115–125
Drummond AJ, Ashton B, Buxton S, Cheung M, Cooper A, Duran C, Field M, Heled J, Kearse M, Markowitz S, Moir R, Stones-Havas S, Sturrock S, Thierer T, Wilson A (2012) Geneious v 5.5.4. Available from www.geneious.com
Edgar RC, Haas BJ, Clemente JC, Quince C, Knight R (2011) UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 27(16):2194–2200
Erwin PM, López-Legentil S, Schuhmann PW (2010) The pharmaceutical value of marine biodiversity for anti-cancer drug discovery. Ecol Econ 70:445–451
Erwin PM, Olson JB, Thacker RW (2011) Phylogenetic diversity, host-specificity and community profiling of sponge-associated bacteria in the northern Gulf of Mexico. PLoS One 6(11):e26806
Fan L, Reynolds D, Liu M, Stark M, Kjelleberg S, Webster NS, Thomas T (2012) Functional equivalence and evolutionary convergence in complex communities of microbial sponge symbionts. Proc Natl Acad Sci 109(27):1878–1887
Felsenstein J (1985) Confidence limits on phylogenies: an approach using the bootstrap. Evolution 39:783–791
Good IJ (1953) The population frequencies of species and the estimation of population parameters. Biometrika 40(3/4):237–264
Groepler W, Schuett C (2003) Bacterial community in the tunic matrix of a colonial ascidian Diplosoma migrans. Helgol Mar Res 57:139–143
Hirose E (2000) Plant rake and algal pouch of the larvae in the tropical ascidian Diplosoma similis: an adaptation for vertical transmission of photosynthetic symbionts Prochloron sp. Zool Sci 17:233–240
Hirose E, Fukuda T (2006) Vertical transmission of photosymbionts in the colonial ascidian Didemnum molle: the larval tunic prevents symbionts from attaching to the anterior part of larvae. Zool Sci 23(8):669–674
Hirose E, Maruyama T (2004) What are the benefits in the ascidian-Prochloron symbiosis? Endocytobiosis Cell Res 15:51–62
Hirose E, Akahori M, Oka AT, Kurabayashi A (2004) Some Prochloron-bearing didemnid ascidians collected from the reef shores of Iriomote Island (Okinawa, Japan). Biol Mag Okinawa 42:7–15
Hirose E, Neilan BA, Scmidt EW, Murakami A (2009) Enigmatic life and evolution of Prochloron and related cyanobacteria inhabiting colonial ascidians. In: Gault PM, Marler HJ (eds) Handbook on cyanobacteria. Bacteriology Research Developments Series. Nova Science Publishers, Inc., pp 161–189
Huse SM, Welch DM, Morrison HG, Sogin ML (2010) Ironing out the wrinkles in the rare biosphere through improved OTU clustering. Environ Microbiol 12(7):1889–1898
Kim SH, Yang HO, Kwon HC (2012) Hasllibacter halocynthiae gen. nov., sp. nov., a nutriacholic acid-producing bacterium isolated from the marine ascidian Halocynthia roretzi. Int J Syst Evol Microbiol 62:624–631
Kojima A, Hirose E (2010) Transfer of prokaryotic algal symbionts from a tropical ascidian (Lissoclinum punctatum) colony to its larvae. Zool Sci 27(2):124–127
Kojima A, Hirose E (2012) Transmission of cyanobacterial symbionts during embryogenesis in the coral reef ascidians Trididemnum nubilum and T. clinides (Didemnidae, Ascidiacea, Chordata). Biol Bull 222:63–73
Kühl M, Larkum AWD (2002) The microenvironment and photosynthetic performance of Prochloron sp. in symbiosis with didemnid ascidians. In: Seckbach J (ed) Cellular origin and life in extreme habitats. Vol. 3: Symbiosis, mechanisms and model systems. Kluwer Acad. Publ, Dordrecht, pp 273–290
Lambert G (2001) A global overview of ascidian introductions and their possible impact on the endemic fauna. In: Sawada H, Tokosawa H, Lambert CC (eds) The biology of ascidians. Springer, Tokyo, pp 249–257
Lambert G (2002) Non-indigenous ascidians in tropical waters. Pac Sci 56:291–298
Lambert G (2007) Invasive sea squirts: a growing global problem. J Exp Mar Biol Ecol 342:3–4
Lambert G (2009) Adventures of a sea squirt sleuth: unraveling the identity of Didemnum vexillum, a global ascidian invader. Aquat Invasions 4(1):5–28
Lejeusne C, Bock DG, Therriault TW, MacIsaac HJ, Cristescu ME (2011) Comparative phylogeography of two colonial ascidians reveals contrasting invasion histories in North America. Biol Invasions 13:635–650
Lewin RA (1978) Distribution of symbiotic didemnids associated with prochlorophytes. In: Proceedings of the International Symposium of marine biogeography and evolution in the Southern Hemisphere, Auckland, New Zealand. NZ DSIR Information Series 137, pp 365–369
Locke A (2009) A screening procedure for potential tunicate invaders of Atlantic Canada. Aquat Invasions 4(1):71–79
López-Legentil S, Turon X, Planes S (2006a) Genetic structure of the star sea squirt, Botryllus schlosseri, introduced in southern European harbours. Mol Ecol 15:3957–3967
López-Legentil S, Turon X, Schupp P (2006b) Chemical and physical defenses against predators in Cystodytes (Ascidiacea). J Exp Mar Biol Ecol 332:27–36
López-Legentil S, Song B, Bosch M, Pawlik JR, Turon X (2011) Cyanobacterial diversity and a new Acaryochloris-like symbiont from Bahamian sea-squirts. PLoS One 6(8):e23938
Lozupone CA, Hamady M, Kelley ST, Knight R (2007) Quantitative and qualitative β diversity Mmeasures lead to different insights into factors that structure microbial communities. Appl Environ Microbiol 73(5):1576–1585
Maldonado M (2007) Intergenerational transmission of symbiotic bacteria in oviparous and viviparous demosponges, with emphasis on intracytoplasmically-compartmented bacterial types. J Mar Biol Assoc UK 87:1701–1713
Martin AP (2002) Pylogenetic approaches for describing and comparing the diversity of microbial communities. Appl Environ Microbiol 68(8):3673–3682
Martínez-García M, Díaz-Valdés M, Wanner G, Ramos-Esplà A, Antón J (2007) Microbial community associated with the colonial ascidian Cystodytes dellechiajei. Environ Microbiol 9(2):521–534
Martínez-García M, Stief P, Díaz-Valdés M, Wanner G, Ramos-Esplá A, Dubilier N, Antón J (2008) Ammonia-oxidizing Crenarchaeota and nitrification inside the tissue of a colonial ascidian. Environ Microbiol 11:2991–3001
Martínez-García M, Díaz-Valdés M, Antón J (2010) Diversity of pufM genes, involved in aerobic anoxygenic photosynthesis, in the bacterial communities associated with colonial ascidians. FEMS Microbiol Ecol 71:387–398
Martínez-García M, Koblízek M, López-Legentil S, Antón J (2011) Epibiosis of oxygenic phototrophs containing chlorophylls a, b, c, and d on the colonial ascidian Cystodytes dellechiajei. Microb Ecol 61:13–19
McDonald J (2004) The invasive pest species Ciona intestinalis (Linnaeus, 1767) reported in a harbour in southern Western Australia. Mar Pollut Bull 49:868–870
McDonald D, Price MN, Goodrich J, Nawrocki EP, DeSantis TZ, Probst A, Andersen GL, Knight R, Hugenholtz P (2012) An improved GreenGenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J 6:610–618
Moss C, Green DH, Pérez B, Velasco A, Henríquez R, McKenzie JD (2003) Intracellular bacteria associated with the ascidian Ecteinascidia turbinata: phylogenetic and in situ hybridisation analysis. Mar Biol 143:99–110
Mouchka ME, Hewson I, Harvell CD (2010) Coral-associated bacterial asemblages: current knowledge and the potential for climate-driven impacts. Integr Comp Biol 50(4):662–674
Münchhoff J, Hirose E, Maruyama T, Sunairi M, Burns BP, Neilan BA (2007) Host specificity and phylogeography of the prochlorophyte Prochloron sp., an obligate symbiont in didemnid ascidians. Environ Microbiol 9(4):890–899
Naranjo SA, Carballo JL, Garcia-Gomez JC (1996) Effects of environmental stress on ascidian populations in Algeciras Bay (southern Spain). Possible marine bioindicators? Mar Ecol Prog Ser 144:119–131
Pedrós-Alió C (2012) The rare bacterial biosphere. Annu Rev Mar Sci 4:449–466
Pester M, Bittner N, Deevong P, Wagner M, Loy A (2010) A ‘rare biosphere’ microorganism contributes to sulfate reduction in a peatland. ISME J 4:1591–1602
Pineda MC (2012) A global wanderer: Biology, phylogeography and resilience of the introduced ascidian Styela plicata. PhD Thesis, University of Barcelona, Barcelona, pp 1–223
Pineda MC, López-Legentil S, Turon X (2011) The whereabouts of an ancient wanderer: global phylogeography of the solitary ascidian Styela plicata. PLoS One 6(9):e25495
Pineda MC, McQuaid CD, Turon X, López-Legentil S, Ordoñez V, Rius M (2012a) Tough adults, frail babies: an analysis of stress sensitivity across early life-history stages of widely introduced marine invertebrates. PLoS One 7(10):e46672
Pineda MC, Turon X, López-Legentil S (2012b) Stress levels over time in the introduced ascidian Styela plicata: the effects of temperature and salinity variations on hsp70 gene expression. Cell Stress Chaperones 17:435–444
Pineda MC, López-Legentil S, Turon X (in press) Year-round reproduction in a seasonal sea: Biological cycle of the introduced ascidian Styela plicata in the Western Mediterranean. Marine Biology. doi:10.1007/s00227-012-2082-7
Pisut DP, Pawlik JR (2002) Anti-predatory chemical defenses of ascidians: secondary metabolites or inorganic acids? J Exp Mar Biol Ecol 270:203–214
Price MN, Dehal PS, Arkin AP (2010) FastTree 2—Approximately maximum-likelihood trees for large alignments. PLoS One 5(3):e9490
Reynolds ES (1963) The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J Cell Biol 17:208–212
Rius M, Pascual M, Turon X (2008) Phylogeography of the widespread marine invader Microcosmus squamiger (Ascidiacea) reveals high genetic diversity of introduced populations and non-independent colonizations. Divers Distrib 14:818–828
Rius M, Pineda MC, Turon X (2009) Population dynamics and life cycle of the introduced ascidian Microcosmus squamiger in the Mediterranean Sea. Biol Invasions 11:2181–2194
Rius M, Turon X, Dias GM, Marshall DJ (2010) Propagule size effects across multiple life-history stages in a marine invertebrate. Funct Ecol 24:685–693
Ronquist F, Huelsenbeck JP (2003) MrBayes 3: bayesian phylogenetic inference under mixed models. Bioinformatics 19(12):1572–1574
Sabbadin A (1957) Il ciclo biologico di Ciona intestinalis (L.), Molgula manhattensis (De Kay) e Styela plicata (Lesueur) nella laguna Veneta. Arch Oceanogr Limnol XI:1–29
Schloss PD, Westcott SL, Ryabin T, Hall JR, Hartmann M, Hollister EB, Lesniewski RA, Oakley BB, Parks DH, Robinson CJ, Sahl JW, Stres B, Thallinger GG, Van Horn DJ, Weber CF (2009) Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl Environ Microbiol 75(23):7537–7541
Schloss PD, Gevers D, Westcott SL (2011) Reducing the effects of PCR amplification and sequencing artifacts on 16S rRNA-based studies. PLoS One 6(12):e27310
Schmidt EW, Nelson JT, Rasko DA, Sudek S, Eisen JA, Haygood MG, Ravel J (2005) Patellamide A and C biosynthesis by a microcin-like pathway in Prochloron didemni, the cyanobacterial symbiont of Lissoclinum patella. Proc Natl Acad Sci 102(20):7315–7320
Schmitt S, Deines P, Behnam F, Wagner M, Taylor MW (2011) Chloroflexi bacteria are more diverse, abundant, and similar in high than in low microbial abundance sponges. FEMS Microbiol Ecol 78:497–510
Sciscioli M, Lepore E, Tursi A (1978) Relationship between Styela plicata (Les.) (Tunicata) settlement and spawning. Mem Biol Mar Oceanogr VIII(III):65–75
Shade A, Handelsman J (2012) Beyond the Venn diagram: the hunt for a core microbiome. Environ Microbiol 14(1):4–12
Shenkar N, Loya Y (2008) The solitary ascidian Herdmania momus: native (Red Sea) versus non-indigenous (Mediterranean) populations. Biol Invasions 10:1431–1439
Sims LL (1984) Osmorregulatory capabilities of three macrosympatric stolidobranch ascidians, Styela clava Herdman, S. plicata (Lesueur), and S. montereyensis (Dall). J Exp Biol Ecol 82:117–129
Sjöstedt J, Koch-Schmidt P, Pontarp M, Canbäck B, Tunlid A, Lundberg P, Hagström A, Riemann L (2012) Recruitment of members from the rare biosphere of marine bacterioplankton communities after an environmental disturbance. Appl Environ Microbiol 78(5):1361–1369
Sogin ML, Morrison HG, Huber JL, Welch DM, Huse SM, Neal PR, Arrieta JM, Herndl GJ (2006) Microbial diversity in the deep sea and the underexplored “rare biosphere”. Proc Natl Acad Sci 103(92):12115–12120
Stam WT, Boele-Bos SA, Stulp BK (1985) Genotypic relationships between Prochloron samples from different localities and hosts as determined by DNA-DNA reassociations. Arch Microbiol 142:340–341
Stefaniak L, Zhang H, Gittenberger A, Smith K, Holsinger K, Lin S, Whitlatch RB (2012) Determining the native region of the putatively invasive ascidian Didemnum vexillum Kott, 2002. J Exp Mar Biol Ecol 422–423:64–71
Stewart CN, Excoffier L (1996) Assessing population genetic structure and variability with RAPD data: application to Vaccinium macrocarpom (American Cranberry). J Evol Biol 9:153–171
Svane IB, Young CM (1989) The ecology and behaviour of ascidian larvae. Oceanogr Mar Biol Annu Rev 27:45–90
Tait E, Carman M, Sievert SM (2007) Phylogenetic divrsity of bacteria associated with ascidians in Eel Pond (Woods Hole, Massachusetts, USA). J Exp Mar Biol Ecol 342:138–146
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S (2011) MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 28(10):2731–2739
Tarjuelo I, López-Legentil S, Codina M, Turon X (2002) Defence mechanisms of adults and larvae of colonial ascidians: patterns of palatability and toxicity. Mar Ecol Prog Ser 235:103–115
Tavaré S (1986) Some probabilistic and statistical problems in the analysis of DNA sequences. In: Miura RM (ed) Some mathematical questions in biology—DNA sequence analysis. American Mathematics Society, Providence, pp 57–86
Thacker RW, Freeman CJ (2012) Sponge-microbe symbioses: recent advances and new directions. Adv Mar Biol 62:57–112
Thiyagarajan V, Qian P-Y (2003) Effect of temperature, salinity and delayed attachment on development of the solitary ascidian Styela plicata (Lesueur). J Exp Mar Biol Ecol 290:133–146
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG (1997) The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 25(24):4876–4882
Tincu JA, Craig AG, Taylor SW (2000) Plicatamide: a lead to the biosynthetic origins of the tunichromes? Biochem Biophys Res Commun 270:421–424
Tincu JA, Menzel LP, Azimov R, Sands J, Hong T, Waring AJ, Taylor SW, Lehrer RI (2003) Plicatamide, an antimicrobial octapeptide from Styela plicata hemocytes. J Biol Chem 278(15):13546–13553
Turner SJ, Thrush SF, Cummings VJ, Hewitt JE, Wilkinson MR, Williamson RB, Lee DJ (1997) Chamges in epifaunal assemblages in response to marina operations and boating activities. Mar Environ Res 43(3):181–199
Turon X, López-Legentil S, Banaigs B (2005) Cell types, microsymbionts, and pyridoacridine distribution in the tunic of three color morphs of the genus Cystodytes (Ascidiacea, Polycitoridae). Invertebr Biol 124(4):355–369
Valero-Jiménez CA, Pérez-Portela R, López-Legentil S (2012) Characterization of novel microsatellite markers from the worldwide invasive ascidian Styela plicata. Conserv Genet Resour 4:559–561
Vázquez E, Young CM (1996) Responses of compound ascidian larvae to haloclines. Mar Ecol Prog Ser 133:179–190
Vázquez E, Young CM (2000) Effects of low salinity on metamorphosis in estuarine colonial ascidians. Invertebr Biol 119(4):433–444
Wang Q, Garrity GM, Tiedje JM, Cole JR (2007) Naïve bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl Environ Microbiol 73(16):5261–5267
Webster NS, Taylor MW (2012) Marine sponges and their microbial symbionts: love and other relationships. Environ Microbiol 14(2):335–346
Webster NS, Taylor M, Behnam F, Lücker S, Rattei S, Whalan S, Horn M, Wagner M (2010) Deep sequencing reveals exceptional diversity and modes of transmission for bacterial sponge symbionts. Environ Microbiol 12(8):2070–2082
Whitlatch RB, Bullard SG (2007) Introduction to the Proceedings of the 1st International Invasive Sea Squirt Conference. J Exp Mar Biol Ecol 342:1–2
Wong NA, McClary D, Sewell MA (2011) The reproductive ecology of the invasive ascidian, Styela clava, in Auckland Harbour, New Zealand. Mar Biol 158:2775–2785
Yamaguchi M (1975) Growth and reproductive cycles of the marine fouling ascidians Ciona intestinalis, Styela plicata, Botrylloides violaceus, and Leptoclinum mitsukurii at Aburatsubo-Moroiso Inlet (Central Japan). Mar Biol 29:253–259
Yokobori S, Kurabayashi A, Neilan BA, Maruyama T, Hirose E (2006) Multiple origins of the ascidian-Prochloron symbiosis: molecular phylogeny of photosymbiotic and non-symbiotic colonial ascidians inferred from 18S rDNA sequences. Mol Phylogenet Evol 40(1):8–19
Acknowledgments
This research was funded by the Marie Curie International Reintegration Grant FP7-PEOPLE-2010-RG 277038 within the 7th European Community Framework Program, the Spanish Government projects CTM2010-17755 and CTM2010-22218, and the Catalan Government grant 2009SGR-484 for Consolidated Research Groups.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Erwin, P.M., Carmen Pineda, M., Webster, N. et al. Small core communities and high variability in bacteria associated with the introduced ascidian Styela plicata . Symbiosis 59, 35–46 (2013). https://doi.org/10.1007/s13199-012-0204-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13199-012-0204-0