
Abstract
Successful wetland restoration depends on the development of the vegetation community post-restoration. Woody vegetation provides functional and structural support to the wetland ecosystem and community development post-restoration dictates restoration outcomes. We investigated basal area, stem density, and species richness of woody vegetation in 40 restored wetlands across West Virginia, USA, ranging in age from 1 to 29 years post-restoration. We aggregated field-collected data into eight indicators at the site scale and investigated stem size distribution to describe the overall woody vegetation community. Generalized linear regression shows native species richness slightly declined as wetland site age increased. In contrast, the total basal area increased over time since restoration. Total stem density did not vary by age. Regardless of age, all sites were dominated by woody vegetation with a stem diameter < 9.1 cm, whereas the frequency of stems > 9.1 cm increased as wetland age increased. This study demonstrates that the development of woody vegetation post-restoration occurs over decades in central Appalachian wetlands and shows the diverse conditions between restoration sites.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Revegetation is a significant component of ecosystem development post-restoration. Specifically, woody vegetation provides structural and functional support to wetlands and is a critical component of post-restoration revegetation, as wildlife (Balcombe et al. 2005a, b; Clipp and Anderson 2014) and macroinvertebrate (Balcombe et al. 2005c; Anderson et al. 2013) communities depend on woody vegetation assemblages. Restoring woody plant species diversity can enhance forested wetland functions such as biomass, carbon, and nitrogen accumulation (Callaway et al. 2003; Davidson et al. 2022). Woody vegetation helps regulate water quantity and improve water quality (Adamus and Brandt 1990). The leftover woody debris that remains after the plant dies plays a vital role in controlling soil temperature, moisture, and subsequent plant growth (Haskell et al. 2012).
Achieving desirable woody vegetation-related monitoring criteria, such as performance standards for wetland mitigation, continually challenges practitioners and managers (Cole and Shafer 2002). Limited or unsuccessful woody vegetation growth can have cascading implications, as some studies suggest that mitigation is not replacing wetland types equally. To achieve appropriate wetland hydrology criteria, practitioners often design projects to retain more water than reference wetlands, resulting in vegetative community shifts (Johnson et al. 2012) at the expense of woody vegetation survival (Morgan and Roberts 2003). Open-water and emergent wetlands are often constructed in place of woody vegetation-dominated wetlands (Cole and Shafer 2002), and projects fail to achieve appropriate vegetative structure (Matthews and Endress 2008). Soil and hydrologic post-restoration monitoring criteria are more often met than vegetative criteria (Mitsch and Wilson 1996; Brown and Veneman 2001; Matthews and Endress 2008).
Additional factors limiting woody vegetation growth and expansion include deer herbivory (Pennington and Walters 2006; Cherefko et al. 2015; Flaherty et al. 2018), variable and unpredictable hydrologic and microtopographic conditions (Bledsoe and Shear 2000; Spencer et al. 2001; Pennington and Walters 2006; Johnson et al. 2012; Diamond et al. 2019), inhospitable physical and chemical soil characteristics (Bledsoe and Shear 2000; Bailey et al. 2007), improper site preparation and soil compaction (Lockhart et al. 2003; Heitmeyer et al. 2013), low survival of planted species (Matthews and Endress 2008), improper species or stock type (Shafer and Roberts 2007; Roquemore et al. 2014), and inappropriate community composition (Matthews et al. 2009a). Woody vegetation also is influenced by factors outside of restoration activities, such as pre-restoration site conditions (Gomez-Aparicio 2009; Heitmeyer et al. 2013), landscape conditions (Matthews et al. 2009b), and the timing and frequency of flooding (McCurry et al. 2010; Johnson et al. 2012).
While the above issues focus on planted stock and planting techniques, natural colonization significantly contributes to post-restoration community development (D’Angelo et al. 2005). Volunteer individuals are more prolific in older sites, while planted individuals dominate younger areas (DeBerry and Perry 2012). In a review of 76 projects in Illinois, 100% of sites achieved naturally revegetating stem criteria (Matthews and Endress 2008). Volunteer species such as black willow (Salix nigra) can have an extremely high density but may have little effect on the survival of planted species (McLeod et al. 2001).
Establishing woody vegetation is vital for restoring wetland ecosystems. Woody vegetation establishment is a regulatory requirement for many compensatory wetland mitigation projects under Sects. 401 and 404 of the Clean Water Act (Hough and Robertson 2009). The slow growth of woody vegetation makes it an excellent indicator of long-term site conditions (Adamus and Brandt 1990). However, this slow growth has also led to many studies questioning the ability to accurately assess ecosystem development within a 5–10-year standard wetland mitigation monitoring period (Mitsch and Wilson 1996; Zedler and Callaway 1999; Matthews et al. 2009a; Robertson et al. 2018). The woody vegetation density increases significantly 15 years following site construction (Cooper et al. 2017), yet it may still take 40 to 50 years to achieve forested wetland conditions (Allen 1997). The 10-year monitoring timeframe to assess woody vegetation as part of successful forested wetland restoration is questionable.
Ecologists measure a variety of variables to quantify woody vegetation changes (Conner and Day 1992; DeBerry and Perry 2004; D’Angelo et al. 2005; Anderson and Mitsch 2008b; Berkowitz 2013, 2019; Walter et al. 2013; Roquemore et al. 2014; Russell and Beauchamp 2017). One commonly used metric is stem diameter at breast height (DBH), which estimates the site’s volume or biomass of woody vegetation. Berkowitz (2019) observed the greatest increases in tree diameter through DBH measurements at 13–20 years and another increase at 25 years post-restoration. However, DBH requires woody vegetation to be at least 1.37 m tall, which limits its utility. Stem area at groundline (SAG) measurements allow shorter stems to be included in biomass estimates (Hudson and Perry Unpublished Report). The SAG is the summed cross-sectional area of measured stems at the groundline. The cumulative SAG measurement describes the proportion of the site covered by woody vegetation. It correlates with biomass accumulation, which provides a woody ecological performance standard linked to wetland function (Hudson and Perry Unpublished Report). The SAG follows a predictable pattern with slow growth during years 2–6, rapid development from years 8–14, and stabilization from ages 16–22 (Hudson and Perry Unpublished Report). Other studies agree that stem area may be a more appropriate metric than stem density (Berkowitz 2013). Research investigating woody vegetation growth can help develop accurate success thresholds for post-restoration evaluation.
This research investigates how woody vegetation indicators and community composition vary with time since restoration, ecoregion, and soil compaction to improve our understanding of wetland restoration. We evaluated woody vegetation from study sites that represented a variety of ages. We included volunteer and planted individuals in the analysis to understand the long-term site dynamics post-restoration. This study investigates indicators of woody vegetation in wetlands ranging from 1 to 29 years since restoration and assesses whether they follow a predictable trajectory over time. Woody vegetation growth indicators include (1) species richness, (2) native species richness, (3) wetland indicator status weighted average, (4) abundance weighted floristic quality index, (5) total stem density, (6) shrub stem density, (7) tree stem density, and (8) SAG. In addition, we incorporated the diameter size class to investigate the resiliency and regenerative properties of the community. We hypothesized that species richness, native species richness, tree stem density, and SAG would increase since restoration and that total stem and shrub stem density would decrease in older wetlands. We also hypothesized that increased soil compaction would decrease native species richness, abundance-weighted floristic quality index, and SAG. Post-restoration ecological studies help inform future restoration revegetation approaches and develop effective post-restoration monitoring criteria. We use results from this study to discuss the role and potential of woody vegetation to be used as monitoring and performance standard criteria.
Methods
Study Area
West Virginia is in the Mid-Atlantic region of the United States. Cold winters and warm summers dominate the climate, with annual precipitation ranging from 1,063 mm to 1,180 mm evenly distributed throughout the year (Wilken et al. 2011). The state is dominated by three Level III ecoregions described by the U.S. Environmental Protection Agency: Ridge and Valley, Central Appalachians, and the Western Allegheny Plateau (Woods et al. 1999). The Central Appalachian Ecoregion is a mixed mesophytic forested land cover with harsh terrain (Woods et al. 1999). Comparatively, the Western Allegheny Plateau has mixed mesophytic and oak forests but is less rugged and forested (Woods et al. 1999). The Ridge and Valley ecoregion is lower in elevation and the least rugged and forested but exhibits more diverse ecosystems from varying relief patterns (Woods et al. 1999). A small portion in the easternmost part of the state intersects the Blue Ridge ecoregion and is defined as having forested slopes along narrow ridgelines underlain with metamorphic rock (Woods et al. 1999).
Wetlands represent only 1% of the state’s surface area but are widely distributed across West Virginia (WVDEP and WVDNR Unpublished Report). Most are small and classified as seasonally, temporarily, or permanently flooded (Tiner 1996). Although some states are prime candidates for wetland mitigation due to population growth and increased pressure for development (BenDor and Doyle 2009), West Virginia exhibits a decreasing population. However, anthropogenic land use changes from resource extraction industries, development, and highway construction still negatively impact natural ecosystem attributes and require mitigation to compensate for impacts on wetland resources. Human-induced land use changes and pollution threaten the state’s wetland and aquatic resource integrity (WVDEP and WVDNR Unpublished Report). Numerous federal, state, and local governmental agencies, non-profit conservation agencies, and for-profit private entities facilitate and implement wetland restoration activities within the state.
Study Sites
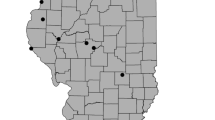
We selected 40 restored wetlands aged 1 to 29 years (x̄ = 9.7, SE = 1.3) for this study based on accessibility and distribution among ecoregions (Fig. 1). Wetlands ranged from 0.20 to 9.5 ha (x̄ = 2.99, SE = 0.39; Appendix A) and varied in elevation from 147 to 1,215 m (x̄ = 495.8 m, SE = 45.5; Appendix B). Wetlands were distributed among all ecoregions (Ridge and Valley (n = 8), Central Appalachians (n = 14), Western Alleghany Plateau (n = 17), and Blue Ridge (n = 1)). Restoration methods varied among study sites and included restoration (n = 5), enhancement (n = 3), establishment (n = 9), or a combination of types, including enhancement and establishment (n = 22) and restoration and enhancement (n = 1). Enhancement improves a specific wetland function (Gwin et al. 1999; USACE and USEPA 2008). Establishment creates a new wetland where one did not exist, and restoration revives a previously existing wetland that became degraded (Gwin et al. 1999; USACE and USEPA 2008). Most study sites were restored for wetland mitigation using mitigation banks (n = 12), in-lieu fee program (n = 8), and permittee-responsible mitigation (n = 13). The remaining study sites were considered voluntary restoration completed by the U.S. Forest Service (n = 3), non-profits (n = 2), a private landowner (n = 1), and the Natural Resources Conservation Service Agriculture Conservation Easement Program (formerly Wetland Reserve Program) (n = 1).

We sampled restored wetlands (n = 40) across four ecoregions in West Virginia, USA. Restored wetlands varied from 1 to 29 years since restoration at the time of field sampling in 2021 (mean ± SE years = 10.3 ± 1.4 years)
Data Collection
We sampled woody vegetation at all sample sites (n = 40) during the 2021 growing season (May – September). Protocols followed DeBerry (2020) and utilized a stratified random sample approach based on wetland class type (Cowardin et al. 1979). Circular plots 100 m2 in area (diameter = 5.6 m) were randomly generated within each wetland class using the ArcGIS Generate Random Points tool to generate plot centroids (DeBerry Unpublished Report). Centroids were buffered at the diameter distance of the plot to avoid overlapping plot areas. While the minimum number of plots per site was four, the number of plots depended on the wetland size, with the total plot area representing at least 2% of the total wetland area to achieve a sufficient sample size (DeBerry Unpublished Report).
Within each plot, we identified all woody vegetation to species, enumerated stems, and measured the stem diameter at the groundline to 0.01 cm using digital calipers. The five largest stems were measured and summed for multi-stem individuals to represent the individual. For live stakes, new shoot growth from the livestake at the base of the stem was measured, as opposed to the diameter of the live stake itself. For plots dominated by a dense monospecific stand, a representative sectional area totaling 1/5th of the plot area was selected, and all individuals were measured, enumerated, and identified. The measured stems in the representative area were multiplied by 5 to estimate the total stems in the entire monospecific stand of the plot. We measured soil penetration resistance at three randomly selected locations within each plot. We used a manual, portable cone soil compaction tester (Dickey-John Corporation, Auburn, Illinois, USA) following the soil-cone penetrometer standards (American Society of Agricultural Engineers 1999). Readings were obtained at 7.6-cm increments up to 45.7 cm in depth (pounds per in2), converted to kilopascals (kPA), and averaged each sampling depth for each site. We cleaned clothing and equipment between sites to avoid the potential spread of invasive species and diseases among wetlands (Bryzek et al. 2022).
We assigned all woody species a wetland indicator status (WIS) and a Coefficient of Conservatism (CoC). The WIS quantitatively ranks species’ probability of occurrence in a wetland environment: upland (UPL) = 5, facultative upland (FACU) = 4, facultative (FAC) = 3, facultative wet (FACW) = 2, and obligate (OBL) = 1 (Lichvar et al. 2014). The CoC value is a measure of a species’ fidelity to undisturbed natural communities, as well as their response to anthropogenic disturbance (Spyreas 2019). The CoC ranks species on a scale of 0, very tolerant to disturbance, to 10, intolerant of disturbance (Spyreas 2019), and has been applied to all West Virginia flora (Rentch and Anderson 2006). In addition, we classified all species by origin (native or non-native) and mature life-form physiognomy (vine, tree, or shrub) based on West Virginia Natural Heritage and West Virginia Department of Environmental Protection databases (Bryzek 2022).
Woody vegetation growth indicators were summarized at the plot level and averaged across all plots to represent site-level metrics, including (1) species richness, (2) native species richness, (3) WIS weighted average, (4) abundance weighted floristic quality index (FQI), (5) total woody stem density (stems/ha), (6) tree density (stems/ha), (7) shrub density (stems/ha), and (8) basal area represented as SAG (m2/ha). We calculated WIS-weighted averages using the following equation:
where \( {y}_{1}, {y}_{2}\), etc., are the relative basal area for each species and \( {u}_{1}, {u}_{2}\), etc., is the corresponding WIS for each species (Atkinson et al. 1993; Balcombe et al. 2005d). A lower WIS weighted average shows that wetland-specific vegetation dominates the woody vegetation community (Atkinson et al. 1993). The FQI uses the CoC, a quantitative indicator of a site’s anthropogenic disturbance and ecosystem health (Bell et al. 2017). We calculated an abundance-weighted FQI using the equation FQI= ∑wmC × (√S) where S is the number of woody plant species and the wmC is an abundance-weighted metric computed for each species using the following equation:
where C = Coefficient of Conservatism and a = abundance, defined as the SAG for each species (Minnesota Pollution Control Agency 2012; Spyreas 2016). In addition, we converted measured stem diameters (D) to SAG using the equation: \( SAG=\pi {\left(\frac{D}{2}\right)}^{2}\).
Statistical Analysis
We examined how wetland vegetation indicators varied with site age using regression analysis. The age of the site (years) at the time of field sampling (2021) was the independent variable, and we used the nine vegetation indicators as the dependent variables. The regression data model was dependent on the vegetation indicator. We used a Poisson regression for species richness and native species richness formatted as count data (Gotelli and Ellison 2004: 34) and general linear models for WIS weighted average, abundance weighted FQI, stem densities, and log(SAG) (Gotelli and Ellison 2004: 46). We log-transformed SAG data to meet homoscedasticity and normality assumptions. As additional predictor variables, we incorporated level III ecoregion and site-averaged soil compaction ratings into each vegetation indicator model. Due to multicollinearity among soil compaction ratings at incremental depths (cm) (7.6, 15.2, 22.8, 30.4, 38), we averaged soil compaction across all depths to consolidate one soil compaction reading for each site (Bryzek 2022). We used the glm function in R statistical software, with a specified family for “poisson” for Poisson models (R Core Team 2022). We tested general linear model assumptions, including normal residuals using the Shapiro–Wilk and homoscedastic errors using the Breusch–Pagan tests.
To further assess woody vegetation community organization, we investigated individuals’ relative frequency of diameter size along the age gradient using generalized linear regression (glm function in R; R Core Team 2022). We used the site age (years) as the independent variable and the relative frequency of stems in each diameter size class (1: 0–0.5 cm, 2: 0.51–1.0 cm, 3: 1.1–2.0 cm, 4: 2.1–3.0 cm, 5: 3.1–4.0 cm, 6: 4.1–5.0 cm, 7: 5.1–7.0 cm, 8: 7.1–9.0 cm, and 9: ≥ 9.1) as the dependent variable. We incorporated ecoregion and soil compaction site averages as additional predictor variables. We conducted all statistical tests in R version 4.0.3 and used α = 0.05 (R Core Team 2022).
Results
Species Occurrence and Distribution
We identified 60 unique species from 25 families and tallied 15,783 stems during the 2021 growing season (Table 1). Most species (n = 54, 90%) were native. Smooth alder (Alnus serrulata) was the most common species (19.3%), followed by white meadowsweet (Spiraea alba) (17%), brushy St. John’s wort (Hypericum densiflorum) (11.7%), silky dogwood (Cornus amomum) (9.6%), steeplebush (Spiraea tomentosa) (5.8%), black willow (Salix nigra) (5.2%), silky willow (Salix sericea) (4.2%), silver maple (Acer saccharinum) (3.4%), American sycamore (Platanus occidentalis) (3.3%), buttonbush (Cephalanthus occidentalis) (2.3%), and alderleaf buckthorn (Rhamnus alnifolia) (2.0%). The other 49 species represented less than 2% of measured stems (Bryzek 2022).
In contrast, the most widely distributed species across all study sites and their percentage of sites detected included black willow (72.5%), silky dogwood (70%), buttonbush (60%), smooth alder (52.5%), American sycamore (40%), multiflora rose (40%), black elderberry (Sambucus nigra ssp. canadensis) (35%), and red maple (Acer rubrum) (25%). The other 52 species (86.7%) were documented at less than 10 (25%) study sites. Non-native species included multiflora rose (Rosa multiflora), Asian bittersweet (Celastrus orbiculatus), Autumn olive (Elaeagnus umbellata var. parvifolia), common St. John’s wort (Hypericum perforatum), Morrow’s honeysuckle (Lonicera morrowii), and white willow (Salix alba). The number of plots sampled per wetland ranged from 4 to 19 (x̄ = 7.5, SE = 0.68).
Woody Vegetation Indicator Trajectories
Across all study sites, total species richness ranged from 1 to 19 (x̄ = 7.6, SE = 0.66), and native species richness ranged from 1 to 17 (x̄ = 6.7, SE = 0.56). The site with the lowest species richness was dominated solely by black willow and was 11 years of age, while the highest species richness occurred at a 13-year-old site. We documented at least one invasive species at 23 sites (57.5%). The highest number of invasives recorded at a site was 4 and occurred at one of the oldest sites (29 years old). The average WIS weighted across all sites was 1.9 (SE = 0.06). The average total stem density (stems/ha) was 848.5 (SE = 157.0), while the average shrub density (stems/ha) was 678.2 (SE = 156.8), and tree density (stems/ha) was 170.3 (SE = 33.8). SAG (m2/ha) ranged from 0.016 to 21.2 (x̄ = 2.570, SE = 0.61).
Regression analysis shows mixed results for wetland age’s effect on vegetation indicators (Fig. 2). Native species richness decreased by 0.029 per year (R2 = 0.40, P = 0.042). Meanwhile, SAG increased with site age (R2 = 0.41, P < 0.001). On average, the log SAG increased by 0.110 m2/ha per year when considering both volunteer and planted woody vegetation. Wetland age did not affect total species richness (R2 = 0.36, P = 0.071), FQI (R2 = 0.16, P = 0.15), WIS weighted average (R2 = 0.01, P = 0.86), total stem density (R2 = 0.22, P = 0.80), shrub density (R2 = 0.25, P = 0.99), or tree density (R2 = 0.14, P = 0.35).

Data were obtained from growing season sampling in restored wetlands (n = 40) that ranged from 1 to 29 years old at the time of field sampling in 2021, West Virginia, USA. No significant trends were detected for (a) total species richness, c) wetland indicator status weighted average,d) woody vegetation floristic quality index, e) total stem density, f) shrub stem density, and g) tree stem density along the age gradient. However, (b) native species richness increases with age, and h) log(stem area at groundline) increased with wetland age. The significant regression line is shown in blue with standard error for regression in grey shading
Average site soil compaction was not a significant predictor variable for any vegetation indicator. Seven sites (17.5%) recorded soil penetrometer resistance above the “good” threshold (1–1380 kPa), meaning the soil was compacted according to the soil penetrometer manufacturer’s standards. All other study sites were within the “good” threshold. At each depth, soil compactions ratings, independent of vegetation indicator, were non-significant along the age gradient. The influence of ecoregion varied among vegetation indicators. Compared to the Central Appalachians, the Western Allegheny Plateau ecoregion had a lower native species richness (P = 0.0495; Fig. 3). The Blue Ridge ecoregion was not included in this analysis since only one study site was located in this ecoregion. Ecoregion was not a significant predictor for other parameters.

Level III ecoregion influences (a) woody vegetation species richness and (b) woody vegetation native species richness in restored wetlands, West Virginia, USA. The Ridge and Valley Ecoregion has the highest species richness and native species richness, followed by the Central Appalachians. Standard error bars represent 95% confidence intervals. Data are from growing season field sampling (May – September 2021) in 40 restored wetlands that varied in the time since restoration from 1 to 29 years, West Virginia, USA
Diameters ranged from 0.1 to 104.8 cm (x̄ = 1.2, SE = 0.019). The largest recorded diameter (104.8 cm) was a five-stemmed black willow at a 29-year-old site, where the stem diameter averaged 26.2 cm. Out of all measured stems during the growing season, only 147 (0.93%) had a stem diameter > 10 cm, while 9,158 (42.0%) were < 1 cm. Black willow represented 49% of stems > 10 cm. The relative frequency of diameters ≥ 9.1 cm increased with wetland age (R2 = 0.43, P = 0.008; Fig. 4). Site age (years) did not affect any other diameter classes (Bryzek 2022). All sites, regardless of age, were dominated by small-diameter stems. However, the ecoregion influenced the relative frequency of size diameter classes when diameters were ≥ 5.0 cm (Fig. 5). The relative frequency of stem diameters of 5.1–7.0 cm was lowest in the Central Appalachian ecoregion and highest in the Western Allegheny Plateau. Soil compaction did not influence the relative frequency of size diameter class distribution.

The relative frequency of woody vegetation diameter ≥ 9.1 cm increased as wetland site age increased. Data are from growing season field sampling (2021) in 40 restored wetlands across West Virginia, USA. Restored wetlands varied in the time since restoration from 1 to 29 years at the time of field sampling

Level III ecoregion influences the relative frequency (%) of woody vegetation stem diameter: 5.1–7.0 cm, 7.1–9.0 cm, and ≥ 9.1 cm. Because stem diameters less than 5.1 cm dominated all ecoregions (89–92%), we have omitted their relative frequencies from the graph to highlight differences in the larger stems. Data from growing season field sampling (2021) in 40 restored wetlands across West Virginia, USA, varied in the time since restoration from 1 to 29 years
Discussion
Our approach using a range of ages to assess woody vegetation metrics suggests differences in woody vegetation growth among sites, and differences in site conditions, restoration methodologies, and tree species and densities resulted in poor prediction of all parameters aside from SAG. SAG exhibited a more linear and predictable increase compared to species richness, WIS-weighted averages, abundance-weighted FQI, total stem density, tree density, and shrub density, which did not change based on the time since restoration. Even though these metrics did not change with site age, their non-significant effect is also informative. For example, the WIS weighted average did not change over time, suggesting that although native species richness may decline, the woody vegetation community still matches appropriate wetland hydrology.
Total stem density, shrub density, and tree density metrics did not show a consistent relationship with site age. Our findings mirror other studies that suggest static stem density requirements are not appropriate indicators of wetland change since restoration (Berkowitz 2013; Hudson and Perry Unpublished Report). SAG was a more similar metric between wetlands of similar ages and appeared less variable than stem density. However, Spencer et al. (2001) found opposite results while examining successional processes in restored bottomland forests where similarly aged sites expressed comparable densities.
In our almost 30-year time frame, SAG did not stabilize. The variability of woody vegetation conditions post-restoration complicates model results and demonstrates the difficulty in recommending static thresholds as performance standards. Our empirical justification shows that SAG alone may have limited application for monitoring sites in the first 15 years after restoration. Nonetheless, SAG holds more promise to assess woody plant growth over time compared to other metrics evaluated.
Our aggregated soil compaction results show that soil compaction did not influence SAG trends. Reduced root growth from soil compaction can reduce tree crown and stem diameter development, but the effects may not be apparent for years after planting (Yingling et al. 1979). In addition, woody vegetation species respond differently to soil compaction, where one species may benefit from increased root-soil contact, while the growth of another may be inhibited (Alameda and Villar 2009). Soil compaction can vary across depths depending on the intensity of disturbance, where deeper soil compaction reveals a more extensive disturbance history (Kozlowski 1999).
Besides soil characteristics, invasive species and biotic interactions influence SAG and other woody vegetation metrics. Invasive species colonization and development influence vegetation 5–10 years after restoration (Matthews and Spyreas 2010). Mortality of planted stock (Matthews and Endress 2008), deer herbivory (Pennington and Walters 2006), and beaver (Castor canadensis) through foraging and altering hydrology (Bonner et al. 2009) can affect metrics. Therefore, natural colonization is essential to woody plant community growth post-restoration. While initial active planting of larger-diameter individuals may help restoration sites achieve a basal area like non-restored forests, seedling germination is necessary to achieve a desirable stem density (Niswander and Mitsch 1995).
The dominance of planted vs. volunteered individuals shifts over time as older restored wetlands exhibit more volunteers (DeBerry and Perry 2012). We attempted to recover planting plans for our study sites but were unsuccessful in incorporating them into the analysis (Bryzek 2022). Many site planting specifications were unavailable, and there was no way to determine which individuals had been planted during field sampling because tree shelters were not used at all sites. The lack of available data suggests that more communication and planning are needed to develop and maintain project files to help guide scientific studies that use project implementation and monitoring data. However, we incorporated woody vegetation community resiliency assessments into our analysis. The stem-diameter size frequency analysis showed a high volume of small individuals across all study sites regardless of age, suggesting that natural colonization continually occurs as time since restoration increases. However, stems greater than 9.1 cm increase as wetland age increases, suggesting that the frequency and prevalence of stem diameter sizes may be applicable to track restoration progress.
Mitigation mechanisms are expected to have different ecological outcomes (Campbell et al. 2002). Specifically, mitigation banks are larger and have more concentrated scientific, funding, and regulatory integration (Spieles 2005). In our study, the voluntary restoration and permittee-responsible sites were generally older than the mitigation banks and in-lieu fee program sites, making comparing ecological conditions among different mitigation mechanisms challenging. This shift in the restoration type mirrors policy changes due to the 2008 Final Rule, which established a preference for mitigation banks, followed by in-lieu fee sites and permittee responsible (USACE and USEPA 2008). The geographic distribution of site ages between ecoregions also revealed an interesting trend in West Virginia. Younger sites dominated the Western Allegheny Plateau, reflecting how and where the geographic extent of wetland mitigation changed over time. This uneven distribution of age classes might account for the lower abundance of intermediate-aged stems from planted individuals and the lower native species richness compared to other ecoregions. This may be more influenced by the uneven distribution of site ages across ecoregions than actual differences in woody vegetation growth between ecoregions. Future studies investigating differences among mitigation mechanisms and West Virginia ecoregions may help validate the preferred means for compensation.
Our data indicate that woody vegetation was still developing over the nearly thirty-year span of sites assessed in this study. Our findings suggest woody vegetation may have limited potential to track restoration development within a 10-year monitoring timeframe. Data from a short monitoring period cannot be used to assess or predict long-term ecosystem development (Robertson et al. 2018). Additionally, there was variation between sites of the same age, suggesting some dissimilarity of woody vegetation parameters between similarly aged sites. Relying solely on woody vegetation species richness or stem densities may lead to inaccurate assessments of site development. Including a more diverse suite of metrics that assess the entire woody vegetation community may be needed (DeBerry and Perry 2012). Incorporating basal metrics, like SAG, may improve assessments with stem density and richness data. Incorporating aerial coverage and height data also may lead to improvements.
Metrics that assess other site components, such as soil and hydrologic characteristics, are also needed to describe site development, as hydrology is a driving factor for restoring site function (Hunter et al. 2008). Woody vegetation community expansion post-restoration also may be limited by planting and implementation techniques during the restoration phase and a lack of natural colonization. Studies suggest that implementation approaches can impact wetland development post-restoration for up to 30 years (Moreno-Mateos et al. 2015). In our study, species richness declined over time, but the frequency of small diameter size classes remained high, suggesting that naturally colonizing species are not increasing species diversity. This finding mirrors other studies evaluating woody species diversity development in restored wetlands (Allen 1997; Matthews et al. 2009a).
Conclusion
Managers and regulators should use caution when relying on woody vegetation development as an evaluation metric post-restoration during the first ten years of monitoring. Due to time, energy, and resource constraints, simplified assessments are often used (Cole and Shafer 2002; Spencer et al. 2001). However, reliance on these short-term snapshots may contribute to the long-term loss of woody vegetation structure and function. In addition, ecosystems are dynamic and continuously change over time. Performance standards that assess vegetative structure are difficult to achieve, especially metrics that relate to woody vegetation (Matthews and Endress 2008). Because of many challenges, achieving successful woody vegetation development post-restoration within the pre-defined monitoring timeframe of 5–10 years is trying. Our results show that woody vegetation may take more than two decades to increase SAG substantially. Failure to attain appropriate woody vegetation abundance may result in lost ecosystem functions and services.
Data Availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Adamus P, Brandt K (1990) Impacts on quality of inland wetlands of the United States: a survey of indicators, techniques, and applications of community-level biomonitoring data. Environmental Protection Agency, Washington, D.C. EPA/600/3–90/073 (NTIS PB91113837).
Alameda D, Villar R (2009) Moderate soil compaction: implications on growth and architecture in seedlings of 17 woody plant species. Soil Tillage Res 103:325–331. https://doi.org/10.1016/j.still.2008.10.029
Allen JA (1997) Reforestation of bottomland hardwoods and the issue of woody species diversity. Restor Ecol 5:125–134. https://doi.org/10.1046/j.1526-100X.1997.09715.x
American Society of Agricultural Engineers (1999) Soil cone penetrometer. ASAE S313.3 (R2018), St. Joseph, MI. https://elibrary.asabe.org/pdfviewer.aspx?GUID=0B92E720-9285-4CA9-BF4F-FD22F9FF0076
Anderson CJ, Mitsch WJ (2008) Tree basal growth response to flooding in a bottomland hardwood forest in central Ohio. J Am Water Resour Assoc 44:1512–1520.https://doi.org/10.1111/j.1752-1688.2008.00255.x
Anderson JT, Zilli FL, Montalto L, Marchese MR, McKinney M, Park YL (2013) Sampling and processing aquatic and terrestrial invertebrates in wetlands. In: Anderson JT, Davis CA (eds) Wetland Techniques. Volume 2: organisms. Springer, New York, New York, pp 142–195
U. S. Army Corps of Engineers (USACE), and U.S. Environmental Protection Agency (USEPA) (2008) Compensatory Mitigation for Losses of Aquatic Resources. Federal Register 73(70) https://www.ecfr.gov/current/title-40/chapter-I/subchapter-H/part-230/subpart-J
Atkinson RB, Perry JE, Smith E, Cairns J Jr (1993) Use of created wetland delineation and weighted averages as a component of assessment. Wetlands 13:185–193. https://doi.org/10.1007/BF03160879
Bailey DE, Perry JE, Daniel WL (2007) Vegetation dynamics in response to organic matter loading rates in a created freshwater wetland in southeastern Virginia. Wetlands 27:936–950. https://doi.org/10.1672/0277-5212(2007)27[936:VDIRTO]2.0.CO;2
Balcombe CK, Anderson JT, Fortney RH, Kordek WS (2005a) Vegetation, invertebrate, and wildlife community rankings and habitat analysis of mitigation wetlands in West Virginia. Wetl Ecol and Manag 13:517–530. https://doi.org/10.1007/s11273-004-5074-7
Balcombe CK, Anderson JT, Fortney RH, Kordek WS (2005b) Wildlife use of mitigation and reference wetlands in West Virginia. Ecol Eng 25:85–99. https://doi.org/10.1016/j.ecoleng.2005.03.003
Balcombe CK, Anderson JT, Fortney RH, Kordek WS (2005c) Aquatic macroinvertebrate assemblages in mitigated and natural wetlands. Hydrobiologia 541:175–188. https://doi.org/10.1007/s10750-004-5706-1
Balcombe CK, Anderson JT, Fortney RH, Rentch JS, Grafton NW, Kordek WS (2005d) A comparison of wetland plant communities in mitigation and reference wetlands in the Mid-Appalachians. Wetlands 25:130–142. https://doi.org/10.1672/0277-5212(2005)025[0130:ACOPCI]2.0.CO;2
Bell JL, Boyer JN, Crystall SJ, Nichols WF, Pruyn M (2017) Floristic quality as an indicator of human disturbance in forested wetlands of northern New England. Ecol Indic 83:227–231. https://doi.org/10.1016/j.ecolind.2017.08.010
BenDor T, Sholtes J, Doyle MW (2009) Landscape characteristics of a stream and wetland mitigation banking program. Ecol Appl 19:2078–2092. https://doi.org/10.1890/08-1803.1
Berkowitz JF (2013) Development of restoration trajectory metrics in reforested bottomland hardwood forests applying a rapid assessment approach. Ecol Indic 34:600–606. https://doi.org/10.1016/j.ecolind.2013.06.025
Berkowitz JF (2019) Quantifying functional increases across a large-scale wetland restoration chronosequence. Wetlands 39:559–573. https://doi.org/10.1007/s13157-018-1103-9
Bledsoe BP, Shear TH (2000) Vegetation along hydrologic and edaphic gradients in a North Carolina coastal plain creek bottom and implications for restoration. Wetlands 20:126–147. https://doi.org/10.1672/0277-5212(2000)020[0126:VAHAEG]2.0.CO;2
Bonner JL, Anderson JT, Rentch JS, Grafton NW (2009) Vegetative composition and community structure associated with beaver ponds in Canaan Valley, West Virginia, USA. Wetl Ecol Manag 17:543–554. https://doi.org/10.1007/s11273-009-9131-0
Brown SC, Veneman PL (2001) Effectiveness of compensatory wetland mitigation in Massachusetts. USA Wetlands 21(4):508–518. https://doi.org/10.1672/0277-5212(2001)021[0508:EOCWMI]2.0.CO;2
Bryzek JA (2022) Wetland mitigation performance standards: The intersection of science, practice, and policy. Thesis, West Virginia University
Bryzek JA, Noe KL, De Silva S, MacKenzie A, Von Haugg CL, Hartman D, McCall JE, Veselka IVW, Anderson JT (2022) Obligations of researchers and managers to respect wetlands: practical solutions to minimizing field monitoring impacts. Land 11:481. https://doi.org/10.3390/land11040481
Callaway JC, Sullivan G, Zedler JB (2003) Species-rich plantings increases biomass and nitrogen accumulation in a wetland restoration experiment. Ecol Appl 13:1626–1639. https://doi.org/10.1890/02-5144
Campbell DA, Cole CA, Brooks RP (2002) A comparison of created and natural wetlands in Pennsylvania, USA. Wetl Ecol Manag 10(1):41–49. https://doi.org/10.1023/A:1014335618914
Cherefko C, Fridley C, Medsger J, Woody M, Anderson JT (2015) White-tailed deer and balsam woolly adelgid effects on balsam fir in Canaan Valley. Southeast Nat 14(sp 7):218–231. https://doi.org/10.1656/058.014.sp719
Clipp HL, Anderson JT (2014) Environmental and anthropogenic factors influencing salamanders in riparian forests: a review. Forests 5:2679–2702. https://doi.org/10.3390/f5112679
Cole CA, Shafer D (2002) Sect. 404 wetland mitigation and permit success criteria in Pennsylvania, USA, 1986–1999. Environ Manage 30:508–515. https://doi.org/10.1007/s00267-002-2717-4
Conner WH, Day JW Jr (1992) Diameter growth of Taxodium distichum (L.) Rich. And Nyssa aquatica L. from 1979–1985 in four Louisiana swamp stands. Am Midl Nat 127(2):290–299. https://doi.org/10.2307/2426535
Cooper DJ, Kacynski KM, Sueltenfuss J, Gaucherand S, Hazen C (2017) Mountain wetland restoration: the role of hydrologic regime in plant introductions after 15 years in the Colorado Rocky Mountains, U.S.A. Ecol Eng 101:46–59. https://doi.org/10.1016/j.ecoleng.2017.01.017
Cowardin LM, Carter V, Golet FC, LaRoe ET Classification of wetland and deepwater habitats of the United States. FWS/OBS-79/31. U. S. Fish and Wildlife Service, Washington DC (1979) USA. https://files.dnr.state.mn.us/lands_minerals/northmet/permit_to_mine/wrp/08_cowardin_et_al_1979_classification_of_wetlands.pdf
D’Angelo EM, Karathanasis AD, Sparks EJ, Ritchey SA, Wehr-McChesney SA (2005) Soil carbon and microbial communities at mitigated and late successional bottomland forest wetlands. Wetlands 25:162–175. https://doi.org/10.1672/0277-5212(2005)025[0162:SCAMCA]2.0.CO;2
Davidson SJ, Dazé E, Byun E, Hiler D, Kangur M, Talbot J, Finkelstein SA, Strack M (2022). The unrecognized importance of carbon stocks and fluxes from swamps in Canada and the USA. Environ Res Lett 17 053003https://doi.org/10.1088/1748-9326/ac63d5/meta
DeBerry DA (2020) Vegetation sampling concepts for compensatory mitigation sites. Wet Sci Pract 37:174–182
DeBerry DA, Perry JE (2004) Primary succession in a created freshwater wetland. Castanea 69:185–193. https://doi.org/10.2179/0008-7475(2004)0690185:PSIACF2.0.CO;2
DeBerry DA, Perry JE (2012) Vegetation dynamics across a chronosequence of created wetland sites in Virginia, USA. Wetl Ecol Manag 20:521–537. https://doi.org/10.1007/s11273-012-9273-3
Diamond JS, McLaughlin DL, Slesak RA, Stovall A (2019) Pattern and structure of microtopography implied autogenic origins in forested wetlands. Hydrol Earth Syst Sci 23:5069–5088. https://doi.org/10.5194/hess-23-5069-2019
Flaherty KL, Rentch JS, Grafton WN, Anderson JT (2018) Timing of white-tailed deer browsing affects wetland plant communities. Plant Ecol 219:313–324. https://doi.org/10.1007/s11258-018-0797-3
Gomez-Aparicio L (2009) The role of plant interactions in the restoration of degraded ecosystems: a meta-analysis across life-forms and ecosystems. J Ecol 97:1202–1214. https://doi.org/10.1111/j.1365-2745.2009.01573.x
Gotelli NJ, Ellison AM (2004) A primer of ecological statistics, vol 1. Sinauer Associates, Sunderland
Gwin SE, Kentula ME, Shaffer PW (1999) Evaluating the effects of wetland regulation through hydrogeomorphic classification and landscape profiles. Wetlands 19:477–489. https://doi.org/10.1007/BF03161687
Haskell D, Flaspohler EDJ, Webster CR, Meyer MW (2012) Variation in soil temperature, moisture, and plant growth on lakeshore restoration sites. Rest Ecol. https://doi.org/10.1111/j.1526-100X.2010.00730.x. 20113-121
Heitmeyer M, Fredrickson L, Laubhan M, Nelson F, Pogue G, Helmers D, King W (2013) Wetland design and development. In: Anderson JT, Davis CA (eds) Wetland Techniques. Volume 3: applications and management. Springer, New York, New York, pp 69–120
Hough P, Robertson M (2009) Mitigation under Sect. 404 of the Clean Water Act: where it comes from, what it means. Wetl Ecol Manage 17:15–33. https://doi.org/10.1007/s11273-008-9093-7
Hunter RG, Faulkner SP, Gibson KA (2008) The importance of hydrology in restoration of bottomland hardwood wetland functions. Wetlands 28:605–615. https://doi.org/10.1672/07-139.1
Johnson YB, Shear TH, James AL (2012) Identifying ecohydrological patterns in natural forested wetlands useful to restoration design. Ecohydrol 5:368–379. https://doi.org/10.1002/eco.227
Kozlowski TT (1999) Soil compaction and growth of woody plants. Scand J for Res 14:596–619. https://doi.org/10.1080/02827589908540825
Lichvar RW, Butterwick M, Melvin NC, Kirchner WN (2014) The national wetland plant list: 2014 update of wetland ratings. Phytoneuron 2014–41:1–42. https://cwbi-app.sec.usace.army.mil/nwpl_static/data/DOC/lists_2014/National/National_2014v1.pdf
Lockhart B, Keeland B, McCoy J, Dean T (2003) Comparing regeneration techniques for afforestating previously farmed bottomland hardwood sites in the Lower Mississippi Alluvial Valley, USA. Forestry (Lond) 76:169–180. https://doi.org/10.1093/forestry/76.2.169
Matthews JW, Endress AG (2008) Performance criteria, compliance success, and vegetation development in compensatory mitigation wetlands. Environ Manage 41:130–141. https://doi.org/10.1007/s00267-007-9002-5
Matthews JW, Spyreas G (2010) Convergence and divergence in plant community trajectories as a framework for monitoring wetland restoration progress. J Appl Ecol 47:1128–1136. https://doi.org/10.1111/j.1365-2664.2010.01862.x
Matthews JW, Spyreas G, Endress AG (2009a) Trajectories of vegetation-based indicators used to assess wetland restoration progress. Ecol Appl 19:2093–2107. https://doi.org/10.1890/08-1371.1
Matthews JW, Peralta AL, Flanagan DN, Baldwin PM, Soni A, Kent AD, Endress AG (2009b) Relative influence of landscape vs. local factors on plant community assembly in restored wetlands. Ecol Appl 19:2108–2123. https://doi.org/10.1890/08-1836.1
McCurry JR, Gray MJ, Mercker DC (2010) Early growing season flooding influence on seedlings of three common bottomland hardwood species in western Tennessee. J Fish Wildl Manag 1:11–18. https://doi.org/10.3996/JFWM-015
McLeod KW, Reed MR, Nelson EA (2001) Influence of a willow canopy on tree seedling establishment for wetland restoration. Wetlands 21:395–402. https://doi.org/10.1672/0277-5212(2001)021[0395:IOAWCO]2.0.CO;2
Minnesota Pollution Control Agency (2012) Rapid Floristic Quality Assessment Manual, vol wq–bwm2–02b. Minnesota Pollution Control Agency, St. Paul, MN.https://conservationresearchinstitute.org/files/minnesota_rapid_assessment_manual.pdf
Mitsch WJ, Wilson RF (1996) Improving the success of wetland creation and restoration with know-how, time, and self‐design. Ecol Appl 6:77–83. https://doi.org/10.2307/2269554
Moreno-Mateos D, Meli P, Vara‐Rodríguez MI, Aronson J (2015) Ecosystem response to interventions: lessons from restored and created wetland ecosystems. J Appl Ecol 52:1528–1537. https://doi.org/10.1111/1365-2664.12518
Morgan KL, Roberts TH (2003) Characterization of wetland mitigation projects in Tennessee. USA Wetlands 23:65–69. https://doi.org/10.1672/0277-5212(2003)023[0065:COWMPI]2.0.CO;2
Niswander SF, Mitsch WJ (1995) Functional analysis of a two-year old created in-stream wetland: hydrology, phosphorus retention, and vegetation survival and growth. Wetlands 15:212–225. https://doi.org/10.1007/BF03160701
Pennington MR, Walters MB (2006) The response of planted trees to vegetation zonation and soil redox potential in created wetlands. For Ecol Manag 233:1–10. https://doi.org/10.1016/j.foreco.2006.04.026
Rentch JS, Anderson JT (2006) A floristic quality index for West Virginia wetland and riparian plant communities. West Virginia Agricultural and Forestry Experiment Station Bulletin 730, Morgantown, WV. 65 pages https://researchrepository.wvu.edu/cgi/viewcontent.cgi?article=1620&context=wv_agricultural_and_forestry_experiment_station_bulletins
Robertson M, Galatowitsch SM, Matthews JW (2018) Longitudinal evaluation of vegetation richness and cover at wetland compensation sites: implications for regulatory monitoring under the Clean Water Act. Wetl Ecol Manag 26:1089–1105. https://doi.org/10.1007/s11273-018-9633-8(0123456789().,-volV)(0123456789().,-volV)
Roquemore JD, Hudson HW III, Atkinson RB, Perry JE (2014) Survival and growth of seven tree species from three stocktypes planted in created wetlands in Loudoun County, Virginia. Ecol Eng 64:408–414. https://doi.org/10.1016/j.ecoleng.2014.01.001
Russell KN, Beauchamp VB (2017) Plant species diversity in restored and created Delmarva Bay wetlands. Wetlands 37:1119–1133. https://doi.org/10.1007/s13157-017-0945-x
Shafer DJ, Roberts TH (2007) Long-term development of tidal mitigation wetlands in Florida. Wetl Ecol Manag 16:23–31. https://doi.org/10.1007/s11273-007-9044-8
Spencer DR, Perry JE, Silberhorn GM (2001) Early secondary succession in bottomland hardwood forests of southeastern Virginia. Environ Manage 27:559–570. https://doi.org/10.1007/s002670010170
Spieles DJ (2005) Vegetation development in created, restored, and enhanced mitigation wetland banks of the United States. Wetlands 25:51–63. https://doi.org/10.1672/0277-5212(2005)025[0051:VDICRA]2.0.CO;2
Spyreas G (2016) Scale and sampling effects on floristic quality. PLoS ONE 11:e0160693. https://doi.org/10.1371/journal.pone.0160693
Spyreas G (2019) Floristic quality assessment: a critique, a defense, and a primer. Ecosphere 10:e02825. https://doi.org/10.1002/ecs2.2825
Tiner RW (1996) Current status of West Virginia’s wetlands: results of the national wetland inventory. U.S. Fish and Wildlife Service, Ecological Services, Region 5, Hadley, MA. 44 pp. plus Appendices. https://www.fws.gov/wetlands/Documents//Current-Status-of-West-Virginias-Wetlands.pdf
Walter WD, Godsey LD, Garrett HE, Dwyer JP, Van Sambeek JW, Ellersieck MR (2013) Survival and 14-year growth of black, white, and swamp white oaks established as bareroot and RPM®-containerized planting stock. North J Appl for 30:43–46. https://doi.org/10.5849/njaf.11-047
Wilken E, Nava FJ, Griffith G (2011) North American Terrestrial Ecoregions-Level III. Commission for Environmental Cooperation, Montreal, Canada. http://www.cec.org/files/documents/publications/10415-north-american-terrestrial-ecoregionslevel-iii-en.pdf
Woods AJ, Omernik JM, Brown DD Level III and IV ecoregions of Delaware, Maryland, Pennsylvania, Virginia, and, Virginia W (1999) U.S. Environmental Protection Agency, Corvallis, OR. https://www.nrc.gov/docs/ML0832/ML083220457.pdf
Yingling EL, Keeley CA, Little S, Burtis J Jr (1979) Reducing damage to shade and woodland trees from construction activities. Arboric J 5:97–105
Zedler JB, Callaway JC (1999) Tracking wetland restoration: do mitigation sites follow desired trajectories? Restor Ecol 7:69–73. https://doi.org/10.1046/j.1526-100X.1999.07108.x
Acknowledgements
Funding support was provided by the United States Environmental Protection Agency (Grant # CD-96383001-0). Thanks to the West Virginia Department of Environmental Protection (WVDEP), the Division of Forestry and Natural Resources at West Virginia University, and the James C. Kennedy Waterfowl and Wetlands Conservation Center at Clemson University for additional support. Special thanks to Elizabeth Byers (WVDEP) for her input and guidance. We appreciate the landowners and managers that allowed wetland access.
Funding
This work was supported by the United States Environmental Protection Agency (Grant # CD-96383001-0).
Author information
Authors and Affiliations
Contributions
Walter Veselka IV and James T. Anderson designed the study. Jessica A. Bryzek collected the data. Jessica A. Bryzek and Christopher T. Rota analyzed the data. Jessica A. Bryzek wrote the first draft of the manuscript with substantial input from James T. Anderson. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic Supplementary Material
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Bryzek, J.A., Veselka IV, W., Rota, C.T. et al. Woody Vegetation Indicators vary with time Since Wetland Restoration. Wetlands 43, 89 (2023). https://doi.org/10.1007/s13157-023-01735-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13157-023-01735-x