
Abstract
Identifying critical hydrologic parameters that structuring biological communities is crucial for the conservation and restoration of wetland ecosystems. The present study determined the key parameters of water level fluctuations (WLFs) influencing the distribution of a typical wetland plant, Carex, in shallow lakes along the middle and lower reaches of the Yangtze River. The plant community and environmental variables in 13 shallow lakes with different WLFs were investigated during 2011–2015. A total of 14 parameters included in three components of WLFs were identified, and their effects on the distribution of Carex were analyzed. The results showed that distribution of Carex in the studied lakes differed obviously in time and space. WLFs were the critical factors influencing the taxa distribution, whereas soil organic matter, human disturbance, substrate type, and soil moisture were of little importance. Rate of water level change and elevation above water were the main parameters determining the coverage and biomass of Carex during the spring growing season. In the autumn growing season, rate of water level change, elevation above water, and emergence timing were important. These results provide important insights into the conservation and restoration of Carex and other hygrophytes in shallow lakes in this region.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Wetlands are diverse and productive ecosystems that harbor a variety of plants and animals and provide valuable goods and services to human society (Richardson 1994; Sellheim et al. 2016). In the past few decades, most wetlands worldwide have been degraded to various degrees due to hydrological alteration, pollution, habitat loss, and species invasion (Tharme 2003; Van Geest et al. 2005a; Wang et al. 2016a). Among these threats, hydrological alteration is the most important because periodic hydrological processes are widely regarded as the main driving forces in riverine ecosystems (Carter 1986; Poff et al. 1997).
In wetlands and lakes, water level fluctuations (WLFs) are very important factors (Coops et al. 2003; Leira and Cantonati 2008), and water level regulation is considered as a useful tool for vegetation restoration (Coops and Hosper 2002; Van Geest et al. 2005a; Zhang et al. 2014). WLFs can be broken down into five components, i.e. magnitude, frequency, duration, timing, and rate of change (Poff et al. 1997; Webb et al. 2012). Previous studies have shown that plant assemblage is strongly structured by WLFs (Casanova and Brock 2000; Van Geest et al. 2005a; Raulings et al. 2010; Bornette and Puijalon 2011). Plants develop a range of morphological characteristics and life history strategies to adapt to the periodic water regime over a long period of time (Visser et al. 2000; Lytle and Poff 2004). For wetland plants, germination, seedling growth, flowering, and seed dispersal depend on WLFs to varying degrees (Rood et al. 1995; Smith and Brock 2007; Edwards et al. 2011; Yang et al. 2015). Most studies on the effects of WLFs have been conducted on riparian and macrophyte species (Rood et al. 2005; Smith and Brock 2007; Zhang et al. 2013), whereas only very limited evidences are available for hygrophytes (Xu et al. 2015; Yang et al. 2015). Moreover, most studies tended to explore the effects of a single WLF component (e.g. magnitude or rate of change) on the germination and seedling growth of plants, mainly by manipulation experiments (Deegan et al. 2007; Smith and Brock 2007; Edwards et al. 2011; Wang et al. 2016b). Evidence regarding the combined effects of different WLF components remains scarce, especially from field investigations (Casanova and Brock 2000; Riis and Hawes 2002; Zhang et al. 2015).
The Yangtze River floodplain is among the most important ecoregions of the world, and there are thousands of shallow lakes located in this region (Wang et al. 2016a). Aquatic plants are abundant (approximately 400 hygrophytes have been recorded), and they play an important role in maintaining the regional biodiversity and functioning of the ecosystem. However, water regime in this region has been dramatically altered due to river-lake disconnection, hydropower projects, and embankment engineering etc. (Wang and Wang 2009), resulting in serious degradation of hygrophyte communities (Hu et al. 2015). Therefore, understanding the effect of WLFs on the hygrophytes is essential for the conservation and restoration of wetlands in this region.
In the present study, we investigated the effects of WLFs on a typical hygrophyte Carex through large-scale field investigations. Carex is widely distributed in the Yangtze shallow lakes (Hu et al. 2013), and it is closely associated with water birds such as goose, providing food and habitats for these migratory species and other aquatic animals (Guan et al. 2014). Our objective is to determine the key parameters of WLFs structuring the Carex community in shallow lakes. We expect that our results can provide valuable information to aid in the conservation and restoration of Carex and other hygrophytes in shallow lakes in this region.
Materials and Methods
Studied Lakes
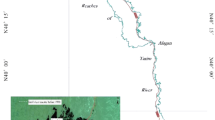
The shallow lakes along the middle and lower reaches of the Yangtze River (108–122°E, 24–34°N) cover an area of ca. 1.6 × 105 km2, accounting for 1/5 of the total lake area in China. Climate in this region is subtropical monsoon. The annual average temperature is 13–20 °C, and the annual precipitation is 800–1600 mm (Wang et al. 2016a; Wang and Wang 2009). Field investigations were carried out in 13 lakes (Fig. 1), including six sub-lakes of Poyanghu Lake with different WLFs. The environmental characteristics of the studied lakes are shown in Appendix Table 4. According to their WLFs characteristics, the studied lakes can be divided into three types, i.e. quasi-natural, intermittent and reservoir-like (Fig. 2) (Zhang 2013). The WLFs of the quasi-natural and intermittent lakes are similar to the water regime of the Yangtze main stem, with the former being larger in amplitude than the latter. The water levels of the reservoir-like lakes are regulated and maintained in a reverse condition.

Locations of the studied lakes along the middle and lower reaches of the Yangtze River

Diagram of the three types of WLFs with standardized water level data
Field Investigations
Carex has two growing seasons each year: the first is from spring to summer (before the flood season), and the second starts after flood recession in autumn (Bernard 1990). Field investigations were therefore carried out in late spring (April–March) and autumn (October–November) during 2011–2015. A total of 88 sampling transects were randomly set, and 417 plots (0.5 m × 0.5 m) were sampled including 124 in spring and 293 in autumn. For each lake, one-three sampling transects were randomly selected. The lower limit of each transect began at the water edge and ended when terrestrial species were encountered (Zhang et al. 2014). In each transect, five-eight plots were set along an elevation gradient of the lakeshore. All species in each plot were recorded and identified according to taxonomic monographs (Zhao and Liu 2009, “Flora Republicae Popularis Sinicae” http://frps.eflora.cn/). The coverage of each species was estimated as the ratio of the projected area of its aboveground parts to the plot area (Li 2000). The total coverage of each plot was calculated as the sum of all species found in the plot. The aboveground parts in the plot were destructively harvested, and the fresh mass was measured with an electronic balance. The occurrence data were further organized as shown in Appendix Table 5.
Five classes of environmental variables were considered including lake morphometric parameters (shoreline development index), nutrition (soil organic matter), substrate (substrate types and soil moisture), human disturbance, and components of WLFs. The shoreline development index was calculated according to the following equation (Aronow 1982): D L = L/2 \( \sqrt{\pi A} \), where L is the shoreline length, and A is the lake area. The values for L and A were extracted from the literatures (Wang and Dou 1998; Zhang 2013; Wang et al. 2016a). Soil organic matter was measured by the K2Cr2O7 titration method after digestion according to standard protocols (Lu 2000; Ding 2013). Substrate types were classified as silt, clay, loam, silt and sand, clay and sand, loam and sand, or sand. Soil moisture were measured with a portable soil moisture meter. Human disturbance was divided into three levels: small (with rare human activities), middle (near to roads or wharfs), and large (with embankment, grazing or cutting).
Calculations of Parameters of WLFs
The WLFs of each lake were quantified using the daily water level data collected from the Hydrology and Water Resource Bureau of Hubei Province (http://www.hbswj.com), Anhui Hydrological Telemetering Information System (http://61.191.22.157/TYFW/InfoQuery/HeDao.aspx), and records of local gauge stations (Dahuchi Lake and Shahuchi Lake). The data for Dahuchi Lake and Shahuchi Lake in 2011 were used to replace the missing data during field investigation in 2012. A total of 14 parameters were quantified belonging to three components of WLFs: magnitude, timing, and rate of change. The elevation above water (E) of each plot was estimated as the perpendicular distance from the center of the plot to the water surface (Zhang et al. 2014) and was treated as a measure of magnitude of WLFs. Monthly E values in February–April and August–October (E-Feb, E-Mar, E-Apr, E-Aug, E-Sep, E-Oct) were calculated to reflect the two growing seasons of Carex. Annual amplitude was also considered as a measure of magnitude, and was calculated as the difference between the minimum and maximum water level for each year (Smith and Brock 2007). Emergence timing (ET) occurred in autumn when water receded and was defined as the first day that the plot emerged. Rate of water level change (R) referred to the daily water level change and was calculated as the absolute difference in water levels between one day and the subsequent day. The R values in February–April (rising) and August–October (declining) (R-Feb, R-Mar, R-Apr, R-Aug, R-Sep, R-Oct) were calculated regarding the two growing seasons. Because of the lack of daily water level data, the amplitude and ET for five lakes (Zhonghuchi Lake, Changhuchi Lake, Zhushihu Lake, Banghu Lake, and Chenhu Lake) were estimated using monthly data, and the E and R values of these lakes could not be calculated.
Data Analyses
The relationships between Carex community and environmental variables were assessed using Spearman correlation analyses and curve estimation regression analyses based on transect data. A redundancy analysis (RDA) was carried out to assess the influence of WLFs, soil moisture, and substrate types on Carex community using the plot data. Three species, i.e. C. cinerascens, C. argyi, and C. laticeps, were dominant in terms of frequency of occurrence, and the critical parameters of WLFs for them were further identified. The choice of the linear ordination method was based on the short gradient lengths determined from a preliminary detrended correspondence analysis (DCA). Data analyses were performed with SPSS 19.0 and Canoco 5.0.
Results
Community Characteristics
A total of 9 Carex species were identified, with the dominant species being C. cinerascens and C. argyi (Appendix Table 5). Besides Carex, there were 112 species belonging to 34 families. Carex grew better in spring than in autumn. The coverage and biomass values were 38.8 ± 5.5% (mean ± SE) and 395.0 ± 60.1 g/m2 in spring, and 27.5 ± 3.4% and 364.1 ± 34.6 g/m2 in autumn, respectively. The biomass varied significantly among the studied lakes (Fig. 3). It was the highest in Banghu Lake in spring and Dongtinghu Lake in autumn, with average values of 891.5 ± 292.3 g/m2 and 651.2 ± 147.1 g/m2, respectively. The lowest biomass was found in Chaohu Lake both in spring and in autumn, with average values of 18.2 ± 14.2 g/m2 and 0.6 ± 0.6 g/m2, respectively.

Comparison of biomass of Carex in different lakes
Environmental Analyses
The occurrence, coverage, and biomass of Carex were significantly related with WLFs (Table 1). A significant negative correlation was found between occurrence and type of WLFs (P < 0.01), indicating that Carex rarely occurred in reservoir-like lakes such as Chaohu Lake and Liangzihu Lake. By contrast, occurrences in spring and in autumn, coverage and biomass in autumn had positive correlations with the amplitude of WLFs (P < 0.05), suggesting that higher amplitude of WLFs favored the growth of Carex. Shoreline development index was also important, since it significantly related to coverage and biomass in spring, and biomass in autumn (P < 0.05). However, no significant relationship was detected between the plant community and soil organic matter or human disturbance (Tables 1 and 2).
Key Parameters of WLFs
RDA analysis showed that the cumulative percentage variation of axes 1 and axes 2 was beyond 80% (Table 3), suggesting that environmental variables related to WLFs, soil moisture, and substrate types explained most of the variations in the distribution of Carex. The ordination diagrams of the species and environmental variables are shown in Fig. 4. Among the parameters of WLFs, R-Feb, R-Mar, and R-Apr played a pivotal role in determining the germination and growth of Carex during the spring growing season. E-Feb, E-Mar and E-Apr were of certain importance (Fig. 4a, b). In the autumn growing season, R-Aug, R-Sep, R-Oct, E-Aug, E-Sep, E-Oct and ET were the predominant parameters of WLFs (Fig. 4c, d), whereas soil moisture and substrate types were of little importance.

RDA ordination diagram of species and environmental variables in spring (a, b) and autumn (c, d). Subst: substrate, E: average elevation above water in a month, e.g. E-Feb represents average elevation above water in February, R: rate of change in a month, e.g. R-Feb represents rate of change in February, ET: emergence timing, SM: soil moisture, C. arg: Carex argyi, C. Lat: Carex laticeps, C. Cin: Carex cinerascens
Discussion
Effects of WLFs
WLFs not only directly affect the growth and reproduction of plants, but also indirectly affect the moisture, oxygen, and substrate that the plants depend on (Keddy and Reznicek 1986; Rood et al. 1995; Coops et al. 2003; Van Geest et al. 2005a; Yu and Yu 2009; Wei et al. 2010). The present study showed that WLFs were the critical factors that determined the distribution of Carex in Yangtze shallow lakes. The magnitude, timing, and rate of change were found to be significantly correlated with the taxa in their different life history stages. The shoreline development index was negatively correlated with Carex, indicating that the shoreline morphology was of certain importance. In this study, lakes with a small shoreline development index usually held a large area of flat inshore region (e.g. Dahuchi Lake, Zhonghuchi Lake etc.), which was beneficial to Carex development. Previous studies have shown that substrate types and nutrients affected plant growth and vegetation succession to some extent (Barko and Smart 1983; Hu et al. 2011). However, they were of little importance in this study mainly because both the substrates and nutrients were quite homogenous among the studied lakes.
Our results showed that Carex biomass differed markedly among the three types of lakes, with the highest values occurring in intermittent lakes and the lowest in reservoir-like lakes. These results suggested that the type of WLFs is very important in shaping the Carex communities in the shallow lakes. The type of WLFs describes the water regime and seasonality of water levels. A reverse condition of the water regime such as that in the reservoir-like lakes is detrimental to Carex as well as other wetland plants. The water levels of intermittent lakes are low and stable before the flood season (Zhang 2013; Wang et al. 2016a), which is beneficial to the germination and growth of these species. By contrast, the reservoir-like lakes usually maintain a higher water level during germination (Zhang et al. 2014; Wang et al. 2016a), exerting detrimental effects on plant development.
Key Parameters of WLFs
Like other wetland plants, Carex adopts life history strategies that are closely correlated with the natural water regime. In different life history stages, the plants are affected by specific components of WLFs. Regarding the magnitude of WLFs, amplitude is an important factor shaping the habitat and plant community (Keddy and Reznicek 1986; Riis and Hawes 2002; Leyer 2005; Van Geest et al. 2005b). Our study revealed that Carex grew better in lakes with a large amplitude (> 3 m). Similar results were also found in other studies in this region (Zhang et al. 2015). Due to the monsoon climate, the highest water level in the Yangtze shallow lakes usually occurs during July–August, when Carex species have already completed their life history in the first growing season. Under these conditions, the taxa could gain competition advantages because a large amplitude would eliminate competing species that cannot tolerate prolonged inundation. In addition, Carex reproduce mainly by belowground rhizomes, and can germinate again when the water recedes (Bernard 1990).
Rate of change in water level was found to be important in both growing seasons of Carex, especially during the germination and fast growing periods. Plants tend to develop a series of morphological adaptions with water level changes (Strand and Weisner 2001; Wu et al. 2012; Zhang et al. 2013), but time is needed for morphological adaptation to occur. In the spring growing season, if the water level rises too fast for Carex to complete any morphological adaptations, the aboveground parts are exposed to inundation stress and even die because of light shortage (Wu et al. 2012) or rapid oxygen depletion (Brock et al. 1987). Also, the rapid decline of the water level in autumn might have detrimental effects on germination and seedling growth due to lack of water. Similar results were also documented in cottonwoods (Populus sp.) and willows (Salix sp.) (Mahoney and Rood 1998; Rood et al. 2005; Amlin and Rood 2002).
Regarding the timing of WLFs, we found that emergence timing in autumn was crucial to Carex development in the autumn growing season. Because Carex cannot germinate underwater, successful germination after water receded depends on the emergence timing and the temperature. If emergence occurs earlier, germination might be prohibited due to water shortage attributed by rapid decrease of water level (Zhang 2013). By contrast, if emergence occurs much later in the autumn growing season, Carex development might be limited by the decreased temperature and inundation stress. The delayed emergence timing in autumn could reduce the growth rate and biomass of Carex, and further result in food shortages of wintering geese in this region (Guan et al. 2014).
Many studies have reported that hygrophytes establish themselves at different elevations in response to WLFs in seasonally flooded wetlands (Lenssen and De Kroon 2005; Leyer 2005). Our results also indicated that elevation above water had significant correlations with the distribution of Carex. In the shallow lakes, elevation per se might have little effects on the germination and growth of hygrophytes, but it is closely related to soil moisture, timing of submergence and emergence, which are crucial for hygrophyte development (Chen et al. 2015; Xu et al. 2015). Soil moisture is proved to be an important factor influencing the germination and growth of wetland plants (Booth and Loheide 2010; Xu et al. 2015; Yang et al. 2015). In the present study, however, no significant relationship was detected between soil moisture and Carex community, probably due to the reason that soil moisture in lakeshore of these shallow lakes was generally high (10%–50%).
Implications for Further Studies and Conservation
Regarding to the effects of WLFs, previous studies have usually conducted by simulation experiments. Water depth and soil moisture are the most concerned because the manipulation of other parameters such as timing and magnitude are difficult and even impossible. In the present study, we revealed the effects of each component of WLFs on the distribution of Carex through filed investigations. Our results provided some new insights compared to previous simulation experiments. For example, we revealed the importance of different parameters of WLFs during the different life history stages of Carex. In addition, several experimental manipulations indicated that a moderate amplitude was beneficial to aquatic plants. However, we found that Carex preferred a large amplitude under natural conditions. Moreover, simulation experiments have shown that inundation can inhibit the germination and growth of Carex (Wang et al. 2007; Yang et al. 2015), whereas we observed that prolonged inundation after the first growing season was necessary for the taxa to reduce interspecific competitions in shallow lakes. Since both simulation experiments and field investigations have pros and cons, a better approach would be to combine them together in the further studies.
In the present study, we determined the effects of different parameters of WLFs on the distribution of Carex by large scale field investigations. Our results can provide important implications considering the conservation and restoration of Carex and other hygrophytes in shallow lakes. Because a reservoir-like type of WLFs is harmful to vegetation development, it is important to keep the water regime in a natural or quasi-natural condition. The present study also revealed that rate of change and emergence timing were key hydrologic parameters determining the distribution of Carex. Water level regulation in this region should take into consideration of these two factors regarding the restoration of Carex and other hygrophytes.
References
Amlin NM, Rood SB (2002) Comparative tolerances of riparian willows and cottonwoods to water-table decline. Wetlands 22:338–346
Aronow S (1982) Shoreline development ratio. In: Schwartz ML (ed) Beaches and coastal geology. Hutchinson Ross Publishing Company, Pennsylvania, pp 754–755
Barko JW, Smart RM (1983) Effects of organic matter additions to sediment on the growth of aquatic plants. Journal of Ecology 71:161–175
Bernard JM (1990) Life history and vegetative reproduction in Carex. Canadian Journal of Botany 68:1441–1448
Booth E, Loheide SP (2010) Soil moisture versus depth-to-water-level: which is better for predicting plant composition in a restored floodplain wetland? Dissertation, American Geophysical Union
Bornette G, Puijalon S (2011) Response of aquatic plants to abiotic factors: a review. Aquatic Sciences 73:1–14
Brock TCM, Van der Velde G, Van de Steeg HM (1987) The effects of extreme water level fluctuations on the wetland vegetation of a nymphaeid-dominated oxbow lake in The Netherlands. Arcl-iivfflr Hydrolliologie Beilzcfte/Ergubeziisse der Lininologitr 27:57–73
Carter V (1986) An overview of the hydrologic concerns related to wetlands in the United States. Canadian Journal of Botany 64:364–374
Casanova MT, Brock MA (2000) How do depth, duration and frequency of flooding influence the establishment of wetland plant communities? Plant Ecology 147:237–250
Chen XS, Li YF, Xie YH, Deng ZM, Li X, Li F, Hou ZY (2015) Trade-off between allocation to reproductive ramets and rhizome buds in Carex brevicuspis populations along a small-scale elevational gradient. Scientific Reports 5:12688
Coops H, Hosper SH (2002) Water-level management as a tool for the restoration of shallow lakes in the Netherlands. Lake and Reservoir Management 18:293–298
Coops H, Beklioglu M, Crisman T (2003) The role of water-level fluctuations in shallow lake ecosystems-workshop conclusions. Hydrobiologia 506(1–3):23–27
Deegan BM, White SD, Ganf GG (2007) The influence of water level fluctuations on the growth of four emergent macrophyte species. Aquatic Botany 86:309–315
Ding QZ (2013) Impact of water level fluctuations on substrate environments of lakeshore vegetation. University of Chinese Academy of Sciences, Beijing (In Chinese)
Edwards AL, Lee DW, Richards JH (2011) Responses to a fluctuating environment: effects of water depth on growth and biomass allocation in Eleocharis cellulosa Torr. (Cyperaceae). Canadian Journal of Botany 81:964–975
Guan L, Wen L, Feng DD, Zhang H, Lei GC (2014) Delayed flood recession in central Yangtze floodplains can cause significant food shortages for wintering geese: results of inundation experiment. Environmental Management 54:1331–1341
Hu XZ, Xu QJ, Jin XC, Jiang LJ (2011) Summary on the interaction between sediment and aquatic macrophytes. Journal of Biology 28:73–76 (In Chinese)
Hu DD, Ouyang HK, Dai ZH (2013) Investigation on community characteristics and α-diversity of Carex cinerascens meadow steppe in Poyang Lake wetland. Pratacultural Science 30:844–848 (In Chinese)
Hu ZP, Ge G, Liu CL (2015) Cause analysis and early warning for wetland vegetation degredation in Poyang Lake. Resources and Environment in the Yangtze Basin 24:381–386 (In Chinese)
Keddy PA, Reznicek AA (1986) Great lakes vegetation dynamics: the role of fluctuating water levels and buried seeds. Journal of Great Lakes Research 12:25–36
Leira M, Cantonati M (2008) Effects of water-level fluctuations on lakes: an annotated bibliography. Hydrobiologia 613:171–184
Lenssen JPM, De Kroon H (2005) Abiotic constraints at the upper boundaries of two Rumex species on a freshwater flooding gradient. Journal of Ecology 93:138–147
Leyer I (2005) Predicting plant species’ responses to river regulation: the role of water level fluctuations. Journal of Applied Ecology 42:239–250
Li B (2000) Composition and structure of biotic community. In: Li B, Yang C, Lin P (eds) Ecology. High Education Press, Beijing, pp 112–149 (In Chinese)
Lu RK (2000) Soil agricultural chemical analysis method. China Agriculture Science and Technique Press, Beijing (In Chinese)
Lytle DA, Poff NL (2004) Adaption to natural flow regimes. Trends in Ecology & Evolution 19:94–100
Mahoney JM, Rood SB (1998) Stream flow requirements for cottonwood seedling recruitment in integrative model. Wetlands 18:634–645
Poff NL, Allan JD, Bain MB, Karr JR, Prestegaard KL, Richter BD, Sparks RE, Stromberg JC (1997) The natural flow regime. BioScience 47:769–784
Raulings EJ, Morris K, Roache MC et al (2010) The importance of water regimes operating at small spatial scales for the diversity and structure of wetland vegetation. Freshwater Biology 55:701–715
Richardson CJ (1994) Ecological functions and human values in wetlands: a framework for assessing forestry impacts. Wetlands 14:1–9
Riis T, Hawes I (2002) Relationships between water level fluctuations and vegetation diversity in shallow water of New Zealand lakes. Aquatic Botany 74:133–148
Rood SB, Mahoney JM, Reid DE, Zilm L (1995) Instream flows and the decline of riparian cottonwoods along the St. Mary River, Alberta. Canadian Journal of Botany 73:1250–1260
Rood SB, Samuelson GM, Braatne JH, Gourley CR, Hughes FMR, Mahoney JM (2005) Managing river flows to restore floodplain forests. Frontiers in Ecology and the Environment 3:193–201
Sellheim KL, Vaghti M, Merz JE (2016) Vegetation recruitment in an enhanced floodplain: ancillary benefits of salmonid habitat enhancement. Limnologica-Ecology and Management of Inland Waters 58:94–102
Smith RGB, Brock MA (2007) The ups and downs of life on the edge: the influence of water level fluctuations on biomass allocation in two contrasting aquatic plants. Plant Ecology 188:103–116
Strand JA, Weisner SEB (2001) Morphological plastic responses to water depth and wave exposure in an aquatic plant (Myriophyllum spicatum). Journal of Ecology 89:166–175
Tharme RE (2003) A global perspective on environmental flow assessment: emerging trends in the development and application of environmental flow methodologies for rivers. River Research and Applications 19:397–441
Van Geest GJ, Coops H, Roijakers RMM, Buijse AD, Scheffer M (2005a) Succession of aquatic vegetation driven by reduced water-level fluctuations in floodplain lakes. Journal of Applied Ecology 42:251–260
Van Geest GJ, Wolters H, Roozen FCJM, Coops H (2005b) Water-level fluctuations affect macrophyte richness in floodplain lakes. Hydrobiologia 539:239–248
Visser EJW, Bögemann GM, Van de Steeg HM, Pierik R, Blom CWPM (2000) Flooding tolerance of Carex species in relation to field distribution and aerenchyma formation. The New Phytologist 148:93–103
Wang SM, Dou HS (1998) Records of Chinese lakes. Science Press, Beijing (In Chinese)
Wang HZ, Wang HJ (2009) Ecological effects of river-lake disconnection and restoration strategies in the midlower Yangtze River. In: Wang ZJ, Shao DG, Shao XJ, Lu YJ, Duan XH (eds) Ecological management on water and sediment in the Yangtze River basin. Science Press, Beijing, pp 379–396 (In Chinese)
Wang L, Hu JM, Song CC, Yang T (2007) Effects of water level on the rhizomatic germination and growth of typical wetland plants in Sanjiang plain. Journal of Applied Ecology 18:2432–2437 (In Chinese)
Wang HZ, Liu XQ, Wang HJ (2016a) The Yangtze River floodplain: threats and rehabilitation. American Fisheries Society Symposium 84:263–291
Wang MZ, Liu ZY, Luo FL, Lei GC, Li HL (2016b) Do amplitudes of water level fluctuations affect the growth and community structure of submerged macrophytes? PloS One 11:e0146528
Webb JA, Wallis EM, Stewardson MJ (2012) A systematic review of published evidence linking wetland plants to water regime components. Aquatic Botany 103:1–14
Wei H, Cheng SP, Wu ZB (2010) Effects of hydrological characteristics on aquatic plants. Modern Agricultural Science and Technology 7:1–16 (In Chinese)
Wu XD, Wang GX, Wei HN (2012) Growth responses of Hydrilla verticillata to increasing water levels. Journal of Lake Science 24:384–390 (In Chinese)
Xu XL, Zhang Q, Tan ZQ, Li YL, Wang XL (2015) Effects of water-table depth and soil moisture on plant biomass, diversity, and distribution at a seasonally flooded wetland of Poyang Lake, China. Chinese Geographical Science 25:739–756
Yang Y, Cao Y, Zhang SJ (2015) Effects of soil moisture regime on rhizomatic germination and young shoot growth of Carex cinerascens. Journal of Ecology and Rural Environment 31:180–187 (In Chinese)
Yu LF, Yu D (2009) Responses of the threatened aquatic plant Ottelia alismoides to water level fluctuations. Fundamental and Applied Limnology 174:295–300
Zhang XK (2013) Water level fluctuation requirements of plants in the Yangtze floodplain lakes. University of Chinese Academy of Sciences, Beijing (In Chinese)
Zhang XK, Liu XQ, Ding QZ (2013) Morphological responses to water-level fluctuations of two submerged macrophytes, Myriophyllum spicatum and Hydrilla verticillata. Journal of Plant Ecology 6:64–70
Zhang XK, Liu XQ, Wang HZ (2014) Developing water level regulation strategies for macrophytes restoration of a large river-disconnected lake. Ecological Engineering 68:25–31
Zhang XK, Liu XQ, Wang HZ (2015) Effects of water level fluctuations on lakeshore vegetation of three subtropical floodplain lakes, China. Hydrobiologia 747:43–52
Zhao JR, Liu YL (2009) Aquatic plant. Huazhong University of Science and Technology Press, Wuhan (In Chinese)
Acknowledgements
This work was supported by the National Natural Science Foundation of China (41371054) and World Wild Fund for Nature (WWF). We would like to thank Xiaoke Zhang for help in field investigations and data analyses, Yajing He for preparation of the map, and Gordon Bradley for his useful suggestions on writing. We also thank Dr. Marinus L. Otte and two anonymous reviewers for their constructive comments.
Author information
Authors and Affiliations
Corresponding author
Appendix
Appendix
Rights and permissions
About this article

Cite this article
Yuan, S., Yang, Z., Liu, X. et al. Key Parameters of Water Level Fluctuations Determining the Distribution of Carex in Shallow Lakes. Wetlands 37, 1005–1014 (2017). https://doi.org/10.1007/s13157-017-0934-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13157-017-0934-0