
Abstract
Vegetation communities in floodplain lakes are adapted to natural water regimes. Construction of sluices or dams between rivers and their floodplain lakes will inevitably lead to great changes in plant communities. In order to evaluate the effects of water level fluctuations (WLFs) on lakeshore vegetation in floodplain lakes of the Yangtze River, a field investigation was conducted in two river-disconnected lakes (Wuchang Lake and Shengjin Lake) and one river-connected lake (Shimen Lake). The results showed that plant species richness was highest in the disconnected Shengjin Lake with intermediate amplitude of WLFs, and lowest in the connected Shimen Lake. Species composition differed among the three lakes and the two disconnected lakes with more similar WLFs exhibiting the greatest similarity. Six plant communities were classified using TWINSPAN. Multivariate analyses showed that the amplitude of WLFs was the most important factor in determining the distribution of lakeshore plants, followed by relative elevation and duration of submergence. Our results provide an eco-hydrological basis for plant restoration in the Yangtze floodplain lakes. We suggest that fluctuating amplitude should be increased in lakes with small WLFs, and small habitats with various hydrological regimes should be created in lakes with large WLFs.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Water regime is the main driving force for the evolution of river floodplain vegetation. Over the long term, plants have gradually adapted to periodic hydrological processes, and have developed a range of morphological, phenological, and life history adaptations (Poff et al., 1997; Lytle & Poff, 2004; Zhang et al., 2014). In shallow lakes, water level fluctuations (WLFs) as a key component of water regime are the major hydrological factors determining the distribution of lakeshore plants (Wilcox & Meeker, 1991; Rills & Hawes, 2002; Leyer, 2005). The shift between low and high water levels can affect species richness and community structure by promoting or inhibiting the germination, growth, and reproduction of plants (Keddy & Reznicek, 1986; Brock & Britton, 1995). Therefore, the natural pattern of WLFs is a critical forcing factor for the development of lakeshore vegetation.
Although the impacts of hydrology on lakeshore vegetation have long been recognized, the role of WLFs in shallow lakes is far from being fully understood (Coops et al., 2003; Abrahams, 2008). There remains a need to determine the relationship between WLFs and lakeshore vegetation, so as to provide information for ecological regulation and management of lakes. The Yangtze floodplain lakes are located in the middle and lower reaches of the Yangtze River, with a total area of 1.57 × 104 km2, accounting for 20% of China’s lake area (Wang & Wang, 2009). Most of these lakes are shallow with a mean water depth of 2 m (Wang & Dou, 1998). Compared with many temperate lakes, they have relatively greater WLFs due to the subtropical monsoon climate that delivers seasonal precipitation and connection with the rising and falling Yangtze mainstem. However, such river–lake connections were blocked by sluices for most lakes during 1950s–1970s. River–lake disconnection has not only changed the connectivity but also altered the WLFs of these lakes. Wang & Wang (2009) preliminarily analyzed the effects of connectivity on different biological groups in the Yangtze floodplain lakes, and found that species richness of fish and macrozoobenthos was significantly higher in river-connected lakes than river-disconnected lakes, but such a tendency was not detected in lake plants. It indicated that the effects of river–lake disconnection on plant diversity were not induced by change of connectivity, but probably by other factors such as alterations of WLFs.
Until now, only a few studies have been carried out to determine the impacts of WLFs on lake plants in the Yangtze floodplain. Most of them were conducted using simulation models (e.g., Yu & Yu, 2009; Zhang et al., 2013) and were focused on one parameter of WLFs (usually water depth). Among the few field studies, most were carried out in a single lake such as the Chaohu Lake (Zhang et al., 2014) and Taihu Lake (Zhao et al., 2012), lacking comparative analyses from lakes with different WLFs. In this study, we chose three different lakes in lower reaches of the Yangtze River to analyze the effects of WLFs on lake plants. We asked: (1) is the plant composition significantly different among the three lakes? and (2) are WLFs the main factors determining the distribution of lakeshore vegetation? We hypothesized that plant composition would be different in response to varied WLFs in the three lakes, and plant diversity would be the highest in lakes with intermediate disturbance of WLFs.
Materials and methods
Study area
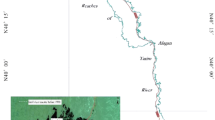
The Shimen Lake, Shengjin Lake, and Wuchang Lake are located in lower reaches of the Yangtze River, Anhui Province (Fig. 1). Climate in this region is subtropical monsoon. Limnological variables of the three lakes are shown in Table 1. The three lakes are close to each other with the shortest linear distance being about 12.8 km between Shimen Lake and Shengjin Lake, and the longest being 26.2 km between Shimen Lake and Wuchang Lake. The surrounding regions of the Shengjin Lake and Wuchang Lake are mainly dominated by farmland, while that of the Shimen Lake is forestland.

Location of the sampling sites in three Yangtze floodplain lakes
Historically, the three lakes were freely connected with the Yangtze mainstem. For flood control and irrigation, Shengjin Lake and Wuchang Lake were regulated through the construction of Huangpen sluice and Zhangshui sluice, respectively. The fluctuating amplitude after the building of sluice was 3.4–7.4 m in Shengjin Lake, and 0.8–5.3 m in Wuchang Lake (Wang & Dou, 1998). Shimen Lake is still connected with the Yangtze mainstem, and has the largest fluctuating amplitude of 10 m. WLFs of the three lakes during the investigation are shown in Fig. 2, and the fluctuating amplitude of Shimen Lake, Shengjin Lake, and Wuchang Lake was 8.58, 5.58, and 2.80, respectively.

Water level fluctuations of the three lakes in 2009
Field sampling
Because late spring and autumn are usually regarded as the best seasons for lakeshore vegetation surveys in this region (Zhao et al., 2012; Zhang et al., 2014), field work was carried out in October, 2009. Seven sampling sites were randomly selected for each lake (Fig. 1). At each site, four to ten plots (1 m × 1 m) were established perpendicular to lakeshore. The plot number and distance between plots depended on the complexity of species composition. Upper limit of the investigation was defined as the appearance of woody species, and the lower limit depended on the distribution of submerged macrophytes (Zhang et al., 2014). A total of 148 plots were used for the three lakes (51 in Wuchang Lake, 65 in Shengjin Lake, and 32 in Shimen Lake). All species were recorded within each plot, and the coverage of each plant was visually estimated using Braun-Blanquet classes: 1 (<5%), 2 (5–25%), 3 (25–50%), 4 (50–75%), 5 (>75%). Horizontal distance to water surface (DWS) and relative elevation above or below the water surface of each plot were measured using meter stick and theodolite (Zhang et al., 2014). Substrate type of the plot was defined as: 1 (clay), 2 (silt), 3 (loam), 4 (sand), 5 (gravel) (Liu, 2008). Conductivity and pH of the substrate were measured by YSI EC 300 and YSI pH100, respectively. Adjoining urban development and roads, grazing, and embankments were considered as disturbance, which was classified as: 1 (light disturbance), 2 (intermediate disturbance), 3 (severe disturbance) (Liu, 2008). Surrounding land use was divided into farmland and forestland. The amplitude of WLFs, duration of submergence, and frequency of submergence were calculated according to the relative elevation of each plot and daily hydrological data. The amplitude was calculated as the difference between highest and lowest water level within a calendar year. Duration was calculated as the sum of days the plot was under submergence, and frequency was the number of times the plot was submerged. Hydrological data of the three lakes were from Anhui Survey and Design Institute of Water Conservancy and Hydropower.
Data analyses
All species were classified into hydrophytes (including emergent, floating, and submerged macrophytes) and hygrophytes according to life form, annuals, and perennials according to life history, sexual reproduction, and combined reproduction (sexual and asexual) according to reproductive strategy. Occurrence frequency of individual species was compared among the three lakes using Chi squared tests. To avoid type I error, the Bonferroni correction (P = 0.05/3 = 0.017) was used to determine the statistically significant differences (Jansson et al., 2000; Liu et al., 2009). One-way analysis of variance (ANOVA) was used to test for difference in coverage, richness, species number of different life histories, life forms, and reproductive strategies among sites in the three lakes. If a significant variance was detected, least significant difference tests (LSD) were conducted to examine the differences between groups. Sørensen coefficient of similarity was used to compare the species composition among the three lakes. All data analyses were done with SPSS 15.0.
Coverage data were analyzed using Two-Way Indicator Species Analysis (TWINSPAN; Hill, 1979) to classify the plants into different groups, which could be conducted by PC-ORD software (McCune & Mefford, 1997). Gradient analysis was used with CANOCO software 4.5 for windows (ter Braak & Smilauer, 1998). The detrended correspondence analysis (DCA) was first carried out to evaluate the type of response model (Vogiatzakis et al., 2009). Based on the results of DCA, the canonical correspondence analysis (CCA) with a unimodal response model was chosen to determine the relationships between species and environmental parameters. The environmental parameters included amplitude of WLFs, duration of submergence, frequency of submergence, relative elevation, DWS, pH, disturbance, conductivity, slope, land use, and substrate type. All data were log transformed to meet the assumption of normality. Species with an occurrence frequency less than 5% were removed prior to the ordination analysis (Bayley & Guimond, 2008). The Monte-Carlo permutation test (P < 0.01) with forward selection was used to determine significant environmental parameters. For simplicity, only significant environmental parameters are shown in the CCA figures.
Results
Species composition
A total of 63 species were recorded in the three lakes (Appendix 1 in ESM). Species number in Shimen Lake, Shengjin Lake, and Wuchang Lake was 21, 43, and 35, respectively. Average species richness per site differed among the three lakes, with higher richness in the disconnected Shengjin Lake compared with the connected Shimen Lake (P = 0.021), but there were no significant differences in coverage (Table 2). The number of hydrophyte species was higher in the two disconnected lakes compared with the connected lake (P = 0.011), while hygrophytes showed no significant differences (Table 2). Analyses of life history and reproductive strategy showed that Shimen Lake had the lowest number of species with annual and perennial life histories, and the fewest species exhibiting both sexual reproduction and combined asexual and sexual reproduction (Table 2).
The Sørensen similarity analyses showed that species were 53.8% similar when comparing the two disconnected lakes (Shengjin and Wuchang), whereas species were 40.6% similar between Shengjin Lake and Shimen Lake, and only 32.1% between Wuchang Lake and Shimen Lake.
Community types
TWINSPAN classification of 148 plots identified six community types after three levels of division (Table 3). The six communities were described as follows:
-
1.
Cynodon dactylon community: the dominant species was Cynodon dactylon, and the common associated species was Cnidium monnieri. The occurrence frequencies of Cynodon dactylon in Wuchang Lake, Shengjin Lake, and Shimen Lake were 0.412, 0.308, and 0.121, respectively, but no significant difference was detected by Chi squared test (Appendix 1 in ESM).
-
2.
Carex brownie community: the dominant species was Carex brownie, and the common associated species were Potentilla supina and Artemisia selengensis. This community only occurred in Shengjin Lake, and the occurrence frequency of Carex brownie was 0.185 (Appendix 1 in ESM).
-
3.
Carex argyi-Artemisia selengensis community: the dominant species were Carex argyi and Artemisia selengensis, and the common associated species was Potentilla supina. Carex argyi only occurred in Shimen Lake. Artemisia selengensis was distributed in Shengjin Lake and Shimen Lake, with frequencies of 0.231 and 0.455, respectively (Appendix 1 in ESM).
-
4.
Phragmites australis community: the dominant species was Phragmites australis, and the common associated species was Artemisia selengensis. The occurrence frequencies of Phragmites australis in Wuchang Lake, Shengjin Lake, and Shimen Lake were 0.039, 0.077, and 0.091, respectively, and no significant difference was detected by Chi squared test (Appendix 1 in ESM).
-
5.
Zizania latifolia community: the dominant species was Zizania latifolia, and the common associated species were Salvinia natans and Lemna minor. The Zizania latifolia only occurred in Wuchang Lake and Shengjin Lake, and occurrence frequency had significant difference between lakes (Appendix 1 in ESM).
-
6.
Salvinia natans-Ceratophyllum demersum community: the dominant species were Salvinia natans and Ceratophyllum demersum, and the common associated species was Myriophyllum spicatum. Both dominant species only occurred in Wuchang Lake and Shengjin Lake, but their occurrence frequency was not significantly different (Appendix 1 in ESM).
Environmental analyses
The CCA biplots for samples-environmental variables and species-environmental variables are shown in Figs. 3 and 4. Forward selection analyses and Monte-Carlo permutation tests indicated that the amplitude of WLFs, duration of submergence, relative elevation, DWS, substrate type, pH, and disturbance were the main influencing factors. The eigenvalues of the first two CCA axes were 0.67 and 0.45, and Monte-Carlo permutation tests indicated both axes were significant at P < 0.001. The axes 1 and 2 accounted for 30.3 and 20.5% of the total variance, respectively.

CCA biplots for samples-environmental variables

CCA biplots for species-environmental variables. Myrspi-Myriophyllum spicatum, Cerdem-Ceratophyllum demersum, Salnat-Salvinia natans, Zizlat-Zizania latifolia, Lemmin-Lemna minor, Altphi-Alternanthera philoxeroides, Cararg-Carex argyi, Juneff-Juncus effusus, Artsel-Artemisia selengensis, Phraus-Phragmites australis, Pollap-Polygonum lapathifolium, Potsup-Potentilla supina, Carbro-Carex brownii, Digsan-Digitaria sanguinalis, Cnimon-Cnidium monnieri, Hydsib-Hydrocotyle sibthorpioides var. batrachium, Cyndac-Cynodon dactylon
Among the 7 main factors, amplitude of WLFs was the most important factor determining the distribution of lakeshore vegetation, followed by relative elevation and duration of submergence (Fig. 3). The hydrophytes such as Ceratophyllum demersum, Myriophyllum spicatum, Salvinia natans, and Zizania latifolia mainly were found in habitats with low elevation, long duration of submergence, and small amplitude of WLFs, whereas the hygrophytes such as Carex brownie, Carex argyi, Cynodon dactylon, and Artemisia selengensis were mainly distributed in habitats with high elevation, short duration of submergence, and large amplitude of WLFs (Fig. 4).
Discussion
Periodic fluctuations in water level can significantly affect the distribution and composition of lakeshore plants (Fernandez-Aláez et al., 1999). When fluctuating amplitude decreases, the distribution area of hygrophytes will shrink, and woody species will expand towards the near-shore zone (Keddy & Reznicek, 1986; Hill et al., 1998). On the contrary, increased fluctuating amplitude will eliminate species that are sensitive to prolonged submergence or drought (Van Geest et al., 2005). The present study showed that plant species richness was highest in the disconnected Shengjin Lake with intermediate amplitude of WLFs. This indicated that WLFs other than connectivity was of great importance in shaping lake plant diversity in this region. In connected Shimen Lake, the dramatic WLFs (8.58 m during the study period) limit the establishment of aquatic plants, but the vast riparian zone provides ideal conditions for hygrophytes; Carex argyi is well adapted to this hydrological environment, and became the dominant species with a mean coverage of 72% (Table 3). In disconnected Wuchang Lake, the relatively stable water level leads to the high habitat stability, which may aggravate the interspecific competitions and make the richness of plants low; the distribution area of Zizania latifolia was higher than 30% of the total lake area, and the whole lake demonstrated terrestrialization tendency. In the disconnected Shengjin Lake, the moderate WLFs can be seen as intermediate disturbance, which can strengthen spatial–temporal heterogeneity and decrease interspecific competition, leading to high plant richness.
Lake area has often been considered as an important factor influencing the plant richness, because more species may appear in large lakes. Although lake area is smallest in the connected Shimen Lake, average species number per site of the disconnected Shengjin Lake was still significantly higher than the Shimen Lake (Table 2). Results about the large connected Poyang Lake also indicated that the dramatic WLFs were the main reasons for low species richness (Li et al., 2005). Otherwise, some results about the temperate lakes showed that species richness had a parabolic relationship with the amplitude of WLFs, but the optimum value was different. For example, Van Geest et al. (2005) suggested an optimum amplitude of 0.4–0.6 m to maximize species richness, while Hill et al. (1998) thought the optimum value should be 1–2 m. Although this study supports the intermediate disturbance hypothesis, the optimum amplitude in Yangtze floodplain lakes is likely higher than temperate lakes because of the local adaptations to these dramatic WLFs.
Annuals have shorter life history compared with the perennials, suggesting they can better adapt to the dramatic WLFs (Liu et al., 2006). However, both the species number and percentage of annuals was lowest in Shimen Lake. This may be due to their different life history strategies. Most annuals are classified as r-strategists, having low resistance to adverse external environment, while the perennials are K-strategists with enhanced self-protection abilities (MacArthur & Wilson, 1967). For example, perennial Carex argyi in Shimen Lake can tolerate long-term flooding through temporary dormancy. The similarity was greatest between the two disconnected lakes, and lowest between Wuchang Lake and Shimen Lake. It indicates that the species similarity mainly depended on the similarity of WLFs. The seed bank is another factor affecting the similarity, but the three lakes are close enough to each other, the seeds could disperse freely through hydrochory before the building of sluices. Thus, presence or absence of seed banks was unlikely the main factor determining the similarity.
Among the six communities, only Phragmites australis and Cynodon dactylon were dominant in all three lakes. This suggested that both species have strong tolerance to WLFs. Some studies have also shown that both species can adapt to long-term drought and flooding (Cao et al., 2008; Wang et al., 2008; Hong et al., 2011). Zizania latifolia was only present in the two disconnected lakes, and its occurrence frequency was significantly higher in Wuchang Lake than Shengjin Lake indicating a need for stable water levels in this species. Otherwise, both Alternanthera philoxeroides and Lemna minor exhibited similar tendencies as Zizania latifolia (Appendix 1 in ESM). The former is an invasive species, and the decrease in WLFs may increase its distribution. The latter is a floating species, and stable water levels may be more conducive to its survival and reproduction. Carex brownie and Carex argyi only occurred in Shengjin Lake and Shimen Lake, respectively. This may be because they respond positively to large WLFs. Both species have adaptations that allow them to complete their life history in late spring and enter a period of temporary dormancy during summer flooding period.
CCA analyses showed that the amplitude of WLFs was the most important factor determining the lakeshore vegetation, followed by relative elevation and duration of submergence. Fluctuating amplitude can directly affect the relative elevation of plants (Leyer, 2005) and further influence the duration of submergence. In this study, we found that species tended to shift from lower to higher elevations with the increase of fluctuating amplitude (Fig. 5). This was consistent with the results of Leyer (2005). Low elevation species occurred at higher elevations when WLFs were increased, indicating their increased submergence at lower elevations. On the contrary, species shifted from higher to lower elevations to avoid permanent drought (Leyer, 2005). Frequency of submergence had no significant effect on the distribution of lakeshore vegetation, mainly because it is low in lakes compared with the rivers.

Comparison of relative elevation of plants in the three lakes (only occurrence frequency >5% and occurred in at least two lakes are shown)
In the Yangtze floodplain lakes, the loss of plant diversity is being given more and more attentions. In this study, we found that WLFs were the main factors determining the distribution of lakeshore plants, and the maximum plant diversity occurred in lakes with intermediate fluctuating amplitude. These results provide significant implications for the restoration of lake plants. Most lakes in this region are controlled by sluices, so their water levels can be ecological regulated. In lakes with small WLFs (such as the Wuchang Lake), the fluctuating amplitude should be increased to improve plant diversity and prevent lake terrestrialization. At the same time, creating small habitats with various hydrological regimes is important for plants restoration in lakes with large WLFs (such as the Shimen Lake).
References
Abrahams, C., 2008. Climate change and lakeshore conservation: a model and review of management techniques. Hydrobiologia 613: 33–43.
Bayley, S. & J. K. Guimond, 2008. Effects of river connectivity on marsh vegetation community structure and species richness in Montana floodplain wetlands in Jasper National Park, Alberta, Canada. Ecoscience 15: 377–388.
Brock, M. & D. Britton, 1995. The role of seed banks in the revegetation of Australian temporary wetlands. In Wheeler, B., S. Shaw, W. Fojt & A. Robertson (eds), Restoration of Temperate Wetlands. Wiley, England: 183–188.
Cao, Y., G. X. Wang & D. Zhang, 2008. Effects of drought stress on the growth and chlorophyll fluorescence of reed seedlings. Arid Land Geography 31: 862–869. (In Chinese).
Coops, H., M. Beklioglu & T. L. Crisman, 2003. The role of water-level fluctuations in shallow lake ecosystems—workshop conclusions. Hydrobiologia 506–509: 23–27.
Fernandez-Aláez, C., M. Fernandez-Aláez & E. Bécares, 1999. Influence of water level fluctuation on the structure and composition of the macrophyte vegetation in two small temporary lakes in the northwest of Spain. Hydrobiologia 415: 155–162.
Hill, M. O., 1979. TWINSPAN, a Fortran Program for Arranging Multivariate Data in an Ordered Two-Way Table by Classification of the Individuals and Attributes. Cornell University Press, Ithaca, New York.
Hill, N. M., P. A. Keddy & I. C. Wisheu, 1998. A hydrological model for predicting the effects of dams on the shoreline vegetation of lakes and reservoirs. Environment Management 22: 723–736.
Hong, M., Q. S. Guo, B. H. Nie, Y. Kang, S. X. Pei, J. Q. Jin & X. F. Wang, 2011. Responses of Cynodon dactylon population in hydro-fluctuation belt of Three Gorges Reservoir area to flooding-drying habitat change. Chinese Journal of Applied Ecology 22: 2829–2835. (In Chinese).
Jansson, R., C. Nilsson, M. Dynesius & E. Andersson, 2000. Effects of river regulation on river-margin vegetation: a comparison of eight boreal rivers. Ecological Applications 10: 203–224.
Keddy, P. A. & A. A. Reznicek, 1986. Great lakes vegetation dynamics: the role of fluctuating water levels and buried seeds. Journal of Great Lakes Research 12: 25–36.
Leyer, I., 2005. Predicting plant species’ responses to river regulation: the role of water level fluctuations. Journal of Applied Ecology 42: 239–250.
Li, W., Y. Cheng & G. H. Liu, 2005. Resource and environmental characteristics of the Poyang Lake, a large lake interconnected with the Changjiang River-Aquatic macrophytes. In Cui, Y. B. & Z. J. Li (eds), Fishery Resources and Conservation of Environment in Lakes of the Changjiang River Basin. Science Press, Beijing: 156–171.
Liu, W. Z., 2008. Prediction of wetland vegetation dynamics in the Danjiangtou Reservoir Region and its role in controlling nonpoint source pollution. Ph.D thesis, Wuhan Botanical Garden, Chinese Academy of Sciences, Wuhan, China. (In Chinese).
Liu, W. Z., Q. F. Zhang & G. H. Liu, 2009. Seed bank of a river-reservoir wetland system and their implications for vegetation development. Aquatic Botany 90: 7–12.
Liu, Y., H. C. Guo, F. Zhou, Z. Wang & K. Huang, 2006. Role of water level fluctuation on aquatic vegetation in lakes. Acta Ecologica Sinica 26: 3117–3126. (In Chinese).
Lytle, D. A. & N. L. Poff, 2004. Adaptation to natural flow regimes. Trends in Ecology and Evolution 19: 94–100.
MacArthur, R. H. & E. O. Wilson, 1967. The Theory of Island Biogeography. Princeton University Press, Princeton, USA.
McCune, B. & M. J. Mefford, 1997. Multivariate Analysis of Ecological Data (PC-ORD). MjM Software, Gleneden Beach, Oregon.
Poff, N. L., J. D. Allan, M. B. Bain, J. R. Karr, K. L. Prestegaard, B. D. Rigther, R. E. Sparks & J. C. Stromberg, 1997. The natural flow regime: a paradigm for river conservation and restoration. BioScience 47: 769–784.
Rills, T. & I. Hawes, 2002. Relationships between water level fluctuations and vegetation diversity in shallow water of New Zealand lakes. Aquatic Botany 74: 133–148.
ter Braak, C. J. F. & P. Smilauer, 1998. CANOCO reference manual and user’s guide to CANOCO for window: software for canonical community ordination (version 4). Microcomputer Power, Ithaca, New York.
Van Geest, G. J., H. Wolters, F. C. J. M. Roozen, H. Coops, R. M. M. Roijackers, A. D. Buijse & M. Scheffer, 2005. Water-level fluctuations affect macrophyte richness in floodplain lakes. Hydrobiologia 539: 239–248.
Vogiatzakis, I. N., G. Kazakis & D. Ghosn, 2009. Macrophyte community struture and species occurrence in relation to environmental determinants in the ephemeral aquatic habitats of Gavdos, Greece. Hydrobiologia 630: 127–138.
Wang, H. F., B. Zeng, Y. Li, P. Qiao, X. Q. Ye & F. L. Luo, 2008. Effects of long-term submergence on survival and recovery growth of four riparian plants species in the Three Gorges Reservoir region, China. Journal of Plant Ecology 32: 977–984. (In Chinese).
Wang, H. Z. & H. J. Wang, 2009. Ecological effects of river-lake disconnection and restoration strategies in the midlower Yangtze River. In Wang, Z. Y. (ed.), Ecological Management on Water and Sediment in the Yangtze River Basin. Science Press, Beijing: 379–396. (In Chinese).
Wang, S. M. & H. S. Dou, 1998. Lakes of China. Science Press, Beijing. (In Chinese).
Wilcox, D. A. & J. E. Meeker, 1991. Disturbance effects on aquatic vegetation in regulated and unregulated lakes in northern Minnesota. Canada Journal of Botany 69: 1542–1551.
Yu, L. F. & D. Yu, 2009. Responses of the threatened aquatic plant Ottelia alismoides to water level fluctuaions. Fundamental and Applied Limnology 174: 295–300.
Zhang, X. K., X. Q. Liu & Q. Z. Ding, 2013. Morphological responses to water level fluctuations of two submerged macrophytes, Myriophyllum spicatum and Hydrilla verticillata. Journal of Plant Ecology 6: 64–70.
Zhang, X. K., X. Q. Liu & H. Z. Wang, 2014. Developing water level regulation strategies to restore the macrophytes of a large river-lake disconnected lake, China. Ecological Engineering 68: 25–31.
Zhao, D. H., H. Jiang, Y. Cai & S. Q. An, 2012. Artificial regulation of water level and its effect on aquatic macrophyte distribution in Taihu Lake. PLoS One 7: e44836.
Acknowledgments
We thank Bo Deng and Yuning Li for their helps in the field investigation. We also thank Dr Tim Jardine and two anonymous reviews for their invaluable comments on this manuscript. This research was supported by National Natural Science Foundation of China (41371054 and 41001117) and the Ministry of Water Resources’ Special Funds for Scientific Research on Public Causes (201101062).
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: Boping Han
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Zhang, X., Liu, X. & Wang, H. Effects of water level fluctuations on lakeshore vegetation of three subtropical floodplain lakes, China. Hydrobiologia 747, 43–52 (2015). https://doi.org/10.1007/s10750-014-2121-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-014-2121-0