
Abstract
To evaluate the influence of human activities on ecosystem respiration (CO2) and CH4 fluxes and determine the seasonal and spatial variations, we measured CO2 and CH4 fluxes at four sampling sites (west side of the seawall, WSS; oilfield, OF; Spartina alterniflora coastal marsh, SCM; aquaculture pond, ACP) in the Yellow River estuary from June to December in 2013. Both CO2 and CH4 fluxes showed seasonal and spatial variations in the Yellow River estuary. The average CO2 fluxes from WSS, OF, SCM and ACP were 125.36, 111.03, 241.97 and −39.49 mg CO2 m−2 h−1, while CH4 fluxes were −0.0110, −0.0165, 0.2012 and 0.0034 mg CH4 m−2 h−1, respectively. Spatial variations of CO2 and CH4 fluxes were mainly affected by vegetation and soil moisture. There were significant relationships between both CO2 fluxes in WSS and SCM and CH4 flux in SCM with temperature. CO2 and CH4 fluxes were mainly affected by the interactions of thermal conditions and other abiotic factors in OF and ACP. Human activities have great effect on greenhouse gas emission, especially in the area where exotic-species S. alterniflora invaded. The construction of seawall blocked sea water transporting into the study area leading to low soil moisture which accelerated CO2 emission. Aquaculture ponds act as an emission of CH4 and consumption of CO2.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Carbon dioxide (CO2) and methane (CH4) are important greenhouse gases (GHG). The concentrations of CO2 and CH4 in atmosphere increased from 280 ppm and 715 ppb in pre-industrial times to 379 ppm and 1,774 ppb in 2005, respectively (IPCC 2007). The levels of CO2 and CH4 have a significant impact on global warming. Therefore, there is a need for quantifying the potential of an individual ecosystem as a source or sink for atmospheric CO2 and CH4 (Purvaja and Ramesh 2001).
Coastal marsh ecosystem is characterized by high temporal and spatial variations including topographic feature, environmental factors, and astronomic tidal fluctuation, and is very sensitive to global climate changes and human activities (Sun et al. 2013). Considerable efforts have been invested in the past two decades to quantify the CO2 and CH4 fluxes in different coastal wetlands (Purvaja and Ramesh 2001; Allen et al. 2007; Cheng et al. 2007; Tong et al. 2012; Sun et al. 2013; Poffenbarger et al. 2011). However, most of the research focused on the emission of GHG from natural wetlands; data of GHG emission from anthropogenic coastal wetland is insufficient. As the development of economy, human activities, such as land-use changes and introduction of invasive alien plants, have more and more impact on coastal wetlands. The phenomenon of transformation of natural coastal wetlands into a harbor, seawall, industrial complex or urban district is very common, this transformation will change the geomorphology of the coastal line and the physical processes of the coastal system permanently, which can result more negative influence on the coastal environment and ecosystem (Bi et al. 2012). Changes in land use have a profound impact on GHG flux. Inubushi et al. (2003) suggested that converting a secondary forest peatland to paddy field increased the annual emissions of CO2 and CH4 to the atmosphere, while transforming the secondary forest to upland decreased the emissions.
Human-induced invasion by exotic-species also have a profound impact on the GHG flux. Invasion by exotic plant species has been considered to be one of the most serious problems for natural ecosystems (Walker and Smith 1997). Spartina alterniflora was introduced to China in 1979, to protect the coastal banks and stabilize the sediment along the eastern coast in Fujian province, Southeast China . Currently, S. alterniflora distributs widely along the east coast of China (Wang et al. 2006a) due to its faster growth rate compared to the native species (Qin and Zhong 1992; Wang et al. 2006b). The coverage of S. alterniflora was approximately 260 ha in six counties by 1985 and increased to more than 112,000 ha by 2,000 (An et al. 2007). Therefore, information on emission of GHG from ecosystem invaded by S. alterniflora is urgently needed, however, studies in this field were mainly conducted at the estuary in the southern part of China (Tong et al. 2012; Cheng et al. 2007; Cheng et al. 2010; Zhang et al. 2010), but information for the estuary in the northern part of China is largely unknown as yet. Thus, it is very important to evaluate how the invasion by exotic-species affects GHG emission in wetlands at estuary area in Northern China.
The Yellow River is well known as a sediment-laden river. Approximately 1.05 × 107 tons of sediment is carried to the estuary and deposited in the delta each year (Cui et al. 2009) resulting in vast area of floodplain and special wetland landscape (Xu et al. 2002; Wang et al. 2004). Typical reclaimed land patterns in the Yellow River estuary included harbor, seawall, salt pans, oilfield, aquaculture ponds and industrial complex. A recent study showed that the area of natural wetlands decreased by 44.5 % in four decades (1976–2008) in the Yellow River estuary, while constructed wetland increased by 1.997 × 104 hm2 in the same period due to rapid development of coastal aquaculture and salt industry (Chen et al. 2011). Wang et al. (2013) pointed that the exploitation of tidal flats resources and construction of artificial ponds related to holothurian culture in the Yellow River delta had become an emerging industry. And the field occupied by holothurian culture covered an area of 1.5 × 104 ha in the Yellow River delta. S. alterniflora was transplanted into Yellow River estuary for three times in 1985, 1987, and 1990. The coverage of S. alterniflora has now reached up to 614.59 hm2 (Zhu et al. 2012). Human activities have more and more impact in Yellow River estuary. Therefore, evaluating the influence of human activities on the emission of GHG is a big necessary in this area.
In this paper, we quantitatively evaluated the variations in the levels of CH4 and CO2 in a typical coastal marsh which was significantly influenced by human activities. The objectives of this study were to: (i) measure the emissions of CO2 and CH4 from exotic S. alterniflora; (ii) determine the relationship between GHG emission and environmental factors; and (iii) estimate the difference of the seasonal change in CO2 and CH4 emissions under different human activities.
Materials and Methods
Site Description
This study was conducted in the Yellow River estuary in Dongying City, Shandong Province, China. The Yellow River estuary has a typical continental monsoon climate with distinct seasons; summer is warm and rainy, and winter is cold. The annual average temperature is 12.1 °C and the frost-free period is 196 d. The average temperatures for spring, summer, autumn and winter are 10.7, 27.3, 13.1, and −5.2 °C, respectively. The mean tidal range of the irregular semidiurnal tide is 0.73 to 1.77 m in the intertidal zone of the Yellow River estuary. Soils in the study area are dominated by intrazonal tide and salt soil. The dissoluble salt content in surface layer (0 to 20 cm) of salt soil is very high (>8 g/kg), and its grain composition is dominated by sand and silt (50–80 %). The average annual evaporation and precipitation are 1962 and 551.6 mm, respectively, with about 70 % of the precipitation occurs in June to August. The main types of vegetation are Sueada salsa, Phragmites australis, Triarrhena sacchariflora, Myriophyllum spicatum, Tamarix chinensis, and Limoninum sinense.
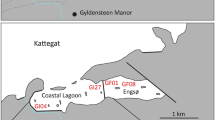
Due to its rich oil and gas resources, Wuhaozhuang region, part of the Yellow River estuary, is significantly affected by human activities. Seawalls were constructed in this region to improve oil production and other economic activities (e.g. aquaculture). S. alterniflora was introduced to Wuhaozhuang in 1990 to protect the seawall from damage. Additionally, Wuhaozhuang is one of important aquaculture farms in Yellow River estuary. We selected four sampling sites in this region, which represented the four typical human-influenced areas in this region, including (a) the west side of the seawall (WSS) (east side of the seawall is the Bohai Sea) (38°01′8.79″N, 118°58′6.84″E); (b) oil field (OF, there are lots of oil wells on the ground) (38°01′8.73″N, 118°57′44.63″E); (c) S. alterniflora coastal marsh (SCM) (38°00′24.8″N, 118°58′23.2″E); and (d) aquaculture pond (ACP) (38°00′26.16″N, 118°58′23.0″E) (Fig. 1).

Sketch of the study area and experimental plots (black star) in the Yellow River estuary
Experimental Design
Fluxes of CO2 and CH4 from WSS, OF, and SCM were measured using static, manual stainless steel chamber and gas chromatography techniques. A stainless steel base with a water groove on the top was inserted into the ground for 20 cm depth in May 2013. A chamber was placed into the groove during measurement; meanwhile water was injected into the groove to build an open-bottom square box. The outside of the chamber was covered with an insulating layer (2 cm thick) to reduce the impact of direct radiative heating during sampling, which can cause very little difference in temperature between the inside and the outside of the chamber (Teiter and Mander 2005; Søvik and Kløve 2007; Jiang et al. 2010; Tong et al. 2010; Sun et al. 2013, 2014). Air inside the chamber was circulated with battery-driven fans installed inside the chamber to make sure that the gas sample was uniform in the chamber. CH4 and CO2 emissions from ACP were measured using floating chambers and gas chromatography techniques. The floating chambers were made of opaque PVC. A flotation gear was installed to make sure that the chamber can float on water. Insulating layer and fans also placed for floating chambers.
Measurements were made in June, August, October, and December of 2013 at the four sites. Each measurement campaign consisted of 12 chambers set up at four positions (three chambers per site). Gas samples were collected at 7:00, 9:30, 12:00, 14:30; and 17:00 h on each sampling date, which have been shown to be the optimum measurement period by Sun et al. (2013, 2014). The gas samples were withdrawn from the headspace of the chamber in a 20-min interval (totally 60 min for each measurement) using a 50 ml syringe equipped with a three-way stopcock. Samples were injected into pre-evacuated packs and taken to the laboratory for determination within 36 h.
The gas samples were determined with gas chromatography (Agilent 7890A, Agilent Co., Santa Clara, CA, USA) equipped with FID. The CH4 was separated from the other gases with a 2 m stainless-steel column, with an inner diameter of 2-mm 13XMS column (60/80 mesh). The CO2 was separated with a 2 m stainless-steel column with an inner diameter of 2 mm Porapak Q (60/80 mesh). The FID operated at 200 °C using high-pure nitrogen as a carrier gas, at a flow rate of 30 ml/min. The column temperatures were maintained at 55 °C for all separations. The greenhouse gas concentrations were quantified by comparing the peak areas of samples against standards. During the gas measurement, standards were analyzed every eight samples of determination to ensure the data quality, the relative standard deviation (RSD) for each sample should below 6 %. The gas flux was calculated using the following equation (Song et al. 2008):
where J is the gas flux (mg m−2 h−1), dc/dt is the slope of the gas concentration curve variation, along with time, M is the mole mass of each gas, P is the atmospheric pressure at the sampling site, T is the absolute temperature during sampling, V 0 , T 0 and P 0 are respectively, the gas mole volume, air absolute temperate and atmospheric pressure under standard conditions, H is the height of chamber above the water surface. The rates of CH4 and CO2 emissions were calculated by fitting the changes in the determined concentrations of CH4 and CO2 over a 60-min period to a linear model. The regression concentration coefficients from linear regressions were rejected when R2 was less than 0.9. Positive values indicate net flux to the atmosphere (efflux), and negative values indicate consumption of atmosphere gases by the soil (influx).
Environmental data were measured at each site during sampling. Air temperature inside the chamber was measured with a thermometer inserted into the chamber. Soil temperature at 5, 10, 15, 20, and 25 cm depth was measured with five ground thermometers inserted into the corresponding depth. On each sampling date, three soil samples per layer (0 to 10, 10 to 20, and 20 to 30 cm) were collected at each site to determine soil water content. Water temperature was also measured with the thermometer during gas sampling. In August 2013, the aboveground biomass in WSS, OF and SCM was estimated. Three quadrants (50 × 50 cm for OF and SCM, 5 × 5 m for WSS) were selected for biomass measurement at each of the three sites. Biomass was oven-dried (80 °C) to a constant weight.
Statistical Analysis
Statistical analyses were conducted using SPSS 16.0 and Origin 7.5. The results were presented as a mean of replicates, with standard error (SE). Significant differences in GHG emissions and environmental factors between different sites were determined by one-way analysis of variance [ANOVA, followed by Tukey’s Honest Significant Difference (HSD) test]. Correlation analysis was conducted to examine the relationship between fluxes and the measured environmental variables. In all tests, differences were considered significant when p < 0.05.
Results
Plant Growth
Vegetation in WSS is predominated by Tamarix chinensis (>60 %), while that in OF and SCM are Suaeda salsa (>99 %) and S. alterniflora (>99 %), respectively. The coverage and maximum aboveground biomass of T. chinensis, S. salsa, and S. alterniflora are 10, 70, 95 %, and 200.11 ± 15.82 (mean ± SE) g m−2, 376.862 ± 31.50 g m−2, 1281.92 ± 176.93 g m−2, respectively. The aboveground biomass of S. alterniflora was greater than these of T. chinensis and S. salsa.
Variation in CO2 Fluxes
CO2 flux includes respiration from living aboveground and belowground plant parts as well as aerobic and anaerobic microbial activities in the soil column. This CO2 flux can be called ecosystem respiration and is associated with the overall carbon flow of the ecosystem (Nykänen et al. 1998). CO2 fluxes from the four sites ranged from −181.23 to 878.03 mg CO2 m−2 h−1 over the entire sampling period (Fig. 2). Average CO2 fluxes in WSS, OF, SCM and ACP from June to December were 125.36, 111.03, 241.97 and −39.49 mg CO2 m−2 h−1, respectively. All sites except ACP (negative value) released CO2 during the entire experimental period. The CO2 flux rates from WSS, OF, and SCM showed the similar seasonal pattern, initial increase followed by a subsequent fall. The greatest CO2 flux rates from WSS (334.69 mg CO2 m−2 h−1), OF (264.32 mg CO2 m−2 h−1), and SCM (583.07 mg CO2 m−2 h−1) were observed in August while the smallest CO2 flux rates (27.08, 13.41, and 37.22 mg CO2 m−2 h−1, respectively) were observed in December. A significantly greater CO2 flux was observed from SCM than WSS (p = 0.028), OF (p = 0.016), and ACP (p = 0.000). The CO2 flux from ACP varied significantly from June to December, and smaller than that from WSS (p = 0.002), OF (p = 0.006) and SCM (p = 0.000). The greatest consumption was observed from ACP in August (−71.14 mg CO2 m−2 h−1).

Carbon dioxide (CO2) fluxes from WSS (west side of the seawall), OF (Oil field), SCM (Spartina alterniflora coastal marsh) and ACP (aquaculture pond) in different months in the Yellow River estuary
Variation in CH4 Fluxes
CH4 fluxes from the four sites ranged from −0.2390 to 0.5252 mg CH4 m−2 h−1 (Fig. 3). Average CH4 fluxes in WSS, OF, SCM, and ACP from June to December were −0.0110, −0.0165, 0.2012, and 0.0034 mg CH4 m−2 h−1, respectively. The flux rate of CH4 from SCM was significantly greater than that from WSS (p = 0.000), OF (p = 0.000), and ACP (p = 0.000). During the entire experimental period, SCM was a net source of CH4, while both net emission and net consumption of CH4 occurred at other sites. The greatest CH4 flux rate from SCM (0.4107 mg CH4 m−2 h−1) was observed in August, while it was observed in October from OF (0.0157 mg CH4 m−2 h−1) and ACP (0.0165 mg CH4 m−2 h−1). The CH4 flux from WSS varied significantly from month to month during the measurement period, with the greatest consumption being observed in October (−0.0258 mg CH4 m−2 h−1).

Methane (CH4) fluxes from WSS (west side of the seawall), OF (Oil field), SCM (Spartina alterniflora coastal marsh) and ACP (aquaculture pond) in different months in the Yellow River estuary
Environmental Factors
There was no significant difference in the air temperature among the four sites (p > 0.05), and so did in soil temperature among different layers (5, 10, 15 and 20 cm depth) at WSS, OF, and SCM (p > 0.05) (Table 1). Spearman correlation analysis indicated that most of the relationships between CO2 (or CH4) fluxes and air temperature (or soil temperature) from OF and ACP were not significant (p > 0.05) (Table 2). CO2 fluxes from SCM and WSS showed significantly positive relationships with air or soil temperature (p < 0.05). And there was no significant relationship between CH4 flux and air/soil temperature (p > 0.05) at the sampling sites except SCM. In addition, CH4 flux significantly correlated with soil water content (positive, p < 0.05), but not for the CO2 flux.
Discussions
Seasonal Variation of CO2 and CH4 Fluxes
CO2 and CH4 emissions varied markedly among different seasons at the four sites (Figs. 2 and 3). Similar variations have been reported in previous studies (Allen et al. 2007; Song et al. 2008; Cheng et al. 2010; Sun et al. 2013). Chen et al. (2010) reported a seasonal variation in CO2 flux in a subtropical mangrove swamp in Hong Kong showed that seasonal variations and the flux in warm seasons was greater than in cold seasons. We also found that CO2 flux in warm seasons (June and August) was significantly greater than that in the cold seasons (October and December) in WSS, OF, and SCM. The significant relationship between CO2 flux and air temperature and soil temperature was consistent with those reported by Cheng et al. (2010) who found that CH4 emissions from S. alterniflora and S. mariqueter soils positively correlated with soil temperature. Similarly, Whalen (2005) observed that seasonal patterns of trace gas emissions were governed by seasonal variability in temperatures affecting water availability, production of substrate precursors, and microbial activity. No significant relationship were found between CO2 flux and air or soil temperature from OF, maybe due in part to the complex interactions of temperature and other biotic/abiotic factors, such as water content.
CH4 flux was not related to temperature significantly expect SCM. Sun et al. (2013) suggested that, in coastal marsh of the Yellow River estuary, seasonal variations in CH4 emission was not affected by seasonal variability in temperatures. In our study, the greatest CH4 emission rate from OF (0.0157 mg CH4 m−2 h−1) and WSS (−0.0258 mg CH4 m−2 h−1) were both observed in October. This result indicated that the influence of temperature on CH4 emission was masked by other biotic/abiotic factors, such as vegetation or soil moisture. Factors affecting CH4 emission are diverse and controlled by the interplay of CH4 production, oxidation, and transport processes (Ding et al. 2004). Kutzbach et al. (2004) reported that the ratio between CH4 production and oxidation is controlled by soil moisture which regulates the relative extent of oxic and anoxic environment within soils. Our result showed that CH4 emissions were positively correlated with soil moisture in WSS, OF, and SCM (Table 2).
CO2 and CH4 fluxes across the air-water interface of ACP had obvious seasonal variations (Figs. 2 and 3). Xing et al. (2005) also pointed that the fluxes of CH4 and CO2 showed strong seasonal dynamics from a shallow hypereutrophic subtropical Lake in China, CH4 emission rate was the greatest in summer, whereas CO2 was adsorbed from the atmosphere in spring and summer, but underwent a large-scale emission in winter. In our study, CO2 flux over the entire sampling period from ACP ranged from −71.14 to −3.96 mg CO2 m−2 h−1, indicating that ACP was a sink for CO2, with the greatest CO2 consumption occurred in August. Previously, 13CDIC and pCO2 measurements suggested that respiration and decomposition of organic sediments were the primary sources of CO2 in the water column (Striegl et al. 2001). Del Giorgio et al. (1999) reported that in the water column of temperate lakes the baseline respiration, fueled by allochthonous C, was independent of phytoplankton production. When primary production was high, baseline respiration was insignificant and algal activity usually dominated the CO2 exchange across air-water interface. Therefore, we consider ACP in our study to be a highly autotrophic water body with high primary production, and the baseline respiration supported by external organic matter was insignificant, which corresponded to the measured CO2 flux that ranged from −71.14 to −3.96 mg CO2 m−2 h−1. Moreover, the insignificant relationship between CO2 flux and air temperature or water temperature (Table 2) confirmed the conclusion that ACP was an autotrophic water body where algal photosynthesis, rather than the mineralization of organic matter, played a more important role in CO2 flux, because an increase in air temperature favored the CO2 production derived from the mineralization of organic matter (Huttunen et al. 2003). Xing et al. (2005) observed that, in a subtropical lake (Donghu, China), exponential relationships between CH4 emission and air, water surface, and sediment surface temperature were observed in a subtropical lake (Donghu, China) (Xing et al. 2005). However, in our study, CH4 emission from ACP was not affected by seasonal variability of temperature. CH4 emission in air-water interface resulted from the balance of two opposing processes: methanogenesis in anoxic conditions and the oxidation of the generated CH4. The production of CH4 is dependent on the concentration of PO4 3− (Schrier-Uijl et al. 2011), phytoplankton primary production (Xing et al. 2005), electron donors and acceptors (Van Bodegom and Scholten 2001). In addition, the CH4 could be oxidized to CO2 during any stage of its travel from the sediment through the water column to the atmosphere (Whiting and Chanton 2001). A large fraction of the unoxidized CH4 was likely to be emitted to the atmosphere by diffusion.
Spatial Variation of CO2 and CH4 Fluxes
Vegetation type and species composition affected the carbon dynamics and the formation and emission of the GHG in wetlands (Van Der Nat and Middelburg 2000; Ström et al. 2005). The significant higher fluxes of CO2 and CH4 from SCM than other three sites indicated that the invasion by exotic S. alterniflora had resulted in increase in CO2 and CH4 fluxes sharply, which was consistent with the result of Tong et al. (2012), who found that CH4 cycling in S. alterniflora marshes was high CH4 production in Min River estuary in southern China. The same conclusion was obtained in the Yangtze River estuary (Cheng et al. 2007) and in Jiangsu province (Zhang et al. 2010).
Plants have three main functions in the regulation of CO2 and CH4 emissions. Firstly, plants act as an important source of methanogenic substrate through excreting labile carbohydrates as exudates and root debris. Minoda et al. (1996) and Watanabe et al. (1999) pointed out that substrates derived from plants contributed up to 90 % of the total CH4 emission. Roots can directly regulate most aspects of rhizosphere C flow either by regulating the exudation process itself or by directly regulating the recapture of exudate from soil (Jones et al. 2004). Zhang et al. (2010) found that increase of CH4 emissions was mainly due to a rise in porewater CH4 concentrations in the S. alterniflora mesocosm, therefore concluded that S. alterniflora could fix and allocate more organic carbon, such as root exudates and debris inputs to the soil. Therefore, when compared with S. salsa, the presence of S. alterniflora ensured enhanced CH4 production and emission in wetlands. In our study, the WSS, OF and SCM were predominated by T. chinensis, S. salsa, and S. alterniflora, respectively. Rhizospheres in these sites were different due to different vegetations and that probably affected the emission of CH4 and CO2. It is likely that the slightest change in the chemistry of the soil or physiology of the plant induced rapid shifts in the quantity and quality of the exudative flux (Jones et al. 2004).
Secondly, numerous reports demonstrated that CH4 emission was well correlated with aboveground living biomass of vegetation (Hirota et al. 2007; Tong et al. 2012). In this study, the maximum aboveground biomass of T. chinensis, S. salsa, and S. alterniflora were 200.1131 ± 15.8172, 376.8582 ± 31.5023, and 1281.9200 ± 176.9304 g m−2, respectively. The high aboveground biomass of S. alterniflora matched with the high emission of CO2 and CH4 from the SCM, therefore supporting the conclusion that aboveground live plant biomass was a key factor controlling carbon production and emission. Tong et al. (2012) also found that S. alterniflora could fix and then allocate more carbon to the soil, which in turn resulted in higher CH4 production and emission when compared with native species.
Thirdly, plants act as a conduit for CO2 and CH4 transport through the aerenchyma system, and as a source of oxygen stimulating CH4 oxidation. Using 14C labeling techniques, Christensen et al. (2003) observed that the emission of CH4 was dependent on the amount of vascular plants. Other studies also found that 39.7–90.0 and 48.8–90.0 % of CH4 emission were transported by S. alterniflora and Phragmites australis (Cheng et al. 2007). S. salsa adapts to tidal inundation because the transportation mechanism carries oxygen from aboveground parts to the roots via the aerenchyma (Han et al. 2005). But we found that CO2 and CH4 emissions from OF (dominated by S. salsa) were lower than those from SCM (dominated by S. alterniflora). That may be due to the difference of vegetations in the two sites. S. salsa is a succulent halophytic herb (Song et al. 2009) while S. alterniflora, which belongs to the perennial grass family, has visibly evident lacunae in its stems (Tong et al. 2012), which may explain why the CH4 transport potentials of S. alterniflora was higher than that of S. salsa. However, S. alterniflora has a thick stem and well-developed aerenchyma tissue, which can deliver more oxygen into the rhizosphere and lead to higher rates of CH4 oxidation under S.alterniflora stands. But CH4 emission from S. alterniflora was still higher than others because this effect was outweighed by the higher CH4 production.
Soil moisture in SCM was greater than that in WSS and OF. This was a result of the construction of seawall, which blocked the transport of sea water into the study area and the low coverage of vegetation in WSS and OF under the strong evaporation (evaporation/precipitation ratio, 3.52) condition. Significantly positive impact of moisture on CH4 emission was observed in WSS, OF, and SCM, but the relationship between CO2 fluxes and moisture was not significant (Table 2). In our study, soil moisture in SCM was greater than these in WSS and OF, and was inundated occasionally by tide on the neap tide day. The consumption of CH4 from WSS and OF and the emission from SCM in our study consisted with the conclusion that soil moisture controlled CH4 emission from sites where the water table fluctuates below the soil surface (Christensen 1993). On other hand, low soil moisture make O2 diffuse into soil to oxygen CH4 to CO2.
Comparisons with Other Studies
At present, reports which focused on GHG emission in Yellow River estuary are scarce. Sun et al. (2013) and Zhang et al. (2014) studied GHG emission in natural wetlands and found that fluxes of CH4 and CO2 were −0.0128 mg CH4 m−2 h−1 (Sun et al. 2013) and 20.86 to 45.31 mg CO2 m−2 h−1 (Zhang et al. 2014), respectively (Table 3). Compared with CO2 and CH4 fluxes from natural wetlands where is affected minimally by human mentioned in these two studies, we found that CO2 fluxes recorded from WSS and OF in our study were greater, while CH4 fluxes were close to the reported values. That might due to the construction of seawall leading to low moisture which accelerated CO2 emission. CO2 and CH4 emissions from S. alterniflora marsh (SCM) in our study were also greater compared with that from natural wetland, differences in vegetation may be the main reason. CH4 flux from S. alterniflora marshes in Yellow River estuary was smaller than these from S. alterniflora marshes in Min River estuary (Tong et al. 2012), Yangtze River estuary (Cheng et al. 2010), and Jiangsu province, China (Zhang et al. 2010), which might due to the different latitudes that study areas located. The emission of CH4 and consumption of CO2 in aquaculture pond, similar to the result of Yang et al. (2012), which different from natural water body (lakes, reservoirs) act as a source of CO2 and CH4 (Tremblay et al. 2005; Silvennoinen et al. 2008; Schrier-Uijl et al. 2011; Diem et al. 2012). Aquaculture ponds are significantly influenced by human activities, therefore nutrient substance content and physicochemical property of these will be different from natural water bodies (lakes, reservoirs), which can lead to differences in GHG emissions.
Conclusion
Human activities have profound impact on greenhouse gas emission in the Yellow River estuary. Exotic-specie S. alterniflora invasion significantly increased CO2 and CH4 emissions due to its strong gas transportation capacity and excreting large amounts of substrates for methanogens. The construction of seawall blocked sea water transporting into the study area leading to low soil moisture which accelerated CO2 emission. Aquaculture pond was a source of CH4 and a sink of CO2. However, the results of this study are preliminary and need to be validated with further studies. More investigations and long-term measurements (including year-to-year variations) on CO2 and CH4 exchanges between ecosystem and atmosphere are needed in order to gain a better understanding of human activities on CO2 and CH4 emissions in the Yellow River estuary.
References
Allen DE, Dalal RC, Rennenberg H, Meyer RL, Reeves S, Schmidt S (2007) Spatial and temporal variation of nitrous oxide and methane flux between subtropical mangrove sediments and the atmosphere. Soil Biology Biochemistry 39:622–631. doi:10.1016/j.soilbio.2006.09.013
An SQ, Gu BH, Zhou CF, Wang ZS, Deng ZF, Zhi YB, Li HL, Chen L, Yu DH, Liu YH (2007) Spartina invasion in China: implications for invasive species management and future research. Weed Research 47:183–191. doi:10.1111/j.1365- 3180.2007.00559.x
Bi XL, Liu FQ, Pan XB (2012) Coastal projects in China: from reclamation to restoration. Environmental Science Technology 46:4691–4692. doi:10.1021/es301286d
Chen GC, Tam NFY, Ye Y (2010) Summer fluxes of atmospheric greenhouse gases N2O, CH4 and CO2 from mangrove soil in South China. Science of Total Environment 408:2761–2767. doi:10.1016/j.scitotenv.2010.03.007
Chen J, Wang SY, Mao ZP (2011) Monitoring wetland changes in Yellow River delta by remote sensing during 1976–2008. Progress in Geography 30:585–592
Cheng XL, Peng RH, Chen JQ, Luo YQ, Zhang QF, An SQ, Chen JK, Li B (2007) CH4 and N2O emissions from Spartina alterniflora and Phragmites australis in experimental mesocosms. Chemosphere 68:420–427. doi:10.1016/j. chemosphere. 2007.01.00
Cheng XL, Luo YQ, Xu Q, Lin GH, Zhang QF, Chen JK, Li B (2010) Seasonal variation in CH4 emission and its 13C-isotopic signature from Spartina alterniflora and Scirpus mariqueter soils in an estuarine wetland. Plant and Soil 327:85–94. doi:10.1007/s11104-009-0033-y
Christensen TR (1993) Methane emission from Arctic tundra. Biogeochemistry 21:117–139. doi:10.1007/BF00000874
Christensen TR, Panikov N, Mastepanov M, Joabsson A, Stewart A, Öquist M, Sommerkorm M, Reynaud S, Svensson B (2003) Biotic controls on CO2 and CH4 exchange in wetlands–a closed environment study. Biogeochemistry 64:337–354. doi:10.1023/A:1024913730848
Cui BS, Yang QC, Yang ZF, Zhang KJ (2009) Evaluating the ecological performance of wetland restoration in the Yellow River delta, China. Ecological Engineering 35:1090–1103. doi:10.1016/j.ecoleng.2009.03.022
Del Giorgio PA, Cole JJ, Caraco NF, Peters RH (1999) Linking planktonic biomass and metabolism to net gas fluxes in northern temperate lakes. Ecology 80:1422–1431. doi:10.1890/0012-9658(1999) 080
Diem T, Koch S, Schwarzenbach S, Wehrli B, Schubert CJ (2012) Greenhouse gas emissions (CO2, CH4, and N2O) from several perialpine and alpine hydropower reservoirs by diffusion and loss in turbines. Aquatic Sciences 74:619–635. doi:10.1007/s00027-012-0256-5
Ding WX, Cai ZC, Tsuruta H (2004) Diel variation in methane emissions from the stands of Carex lasiocarpa and deyeuxia angustifolia in a cool temperate freshwater marsh. Atomospheric Environment 38:181–188. doi:10.1016/j. atmosenv. 2003. 09.066
Han N, Shao Q, Lu CM, Wang BS (2005) The leaf tonoplast V-H+-ATPase activity of a C3 halophyte Suaeda salsa is enhanced by salt stress in a Ca-dependent mode. Journal of Plant Physiology 162:267–274
Hirota M, Senga Y, Seike Y, Nohara S, Kunii H (2007) Fluxes of carbon dioxide, methane and nitrous oxide in two contrastive fringing zones of coastal lagoon, Lake Nakaumi, Japan. Chemosphere 68:597–603. doi:10.1016/j.chemosphere. 2007. 01.002
Huttunen JT, Alm J, Liikanen JS, Larmola T, Hannar T, Sivola J, Martikainen PJ (2003) Fluxes of methane, carbon dioxide and nitrous oxide in boreal lakes and potential anthropogenic effects on the aquatic greenhouse gas emissions. Chemosphere 52:609–621. doi:10.1016/S0045-6535(03)00243-1
Inubushi K, Furukawa Y, Hadi A, Purnomo E, Tsuruta H (2003) Seasonal changes of CO2, CH4 and N2O fluxes in relation to land-use change in tropical peatlands located in coastal area of South Kalimantan. Chemosphere 52:603–608. doi:10.1016/ S0045- 6535(03)00242-X
IPCC (2007) Climate change 2007: the scientific basis. Cambridge University Press, New York
Jiang C, Yu G, Fang H, Cao G, Li Y (2010) Short-term effect of increasing nitrogen deposition on CO2, CH4 and N2O fluxes in an alpine meadow on the Qinghai-Tibetan Plateau, China. Atmospheric Environment 44:2920–2926. doi:10.1016/j.atmosenv.2010.03.030
Jones DL, Hodge A, Kuzyakov Y (2004) Plant and mycorrhizal regulation of rhizodeposition. New Phytologist 163:459–480. doi:10.1111/j.1469-8137. 2004.01130.x
Kutzbach L, Wagner D, Pfeiffer EM (2004) Effect of microrelief and vegetation on methane emission from wet polygonal tundra, Lena Delta, Northern Siberia. Biogeochemistry 69:341–362. doi:10.1023/B:BIOG.0000031053.81520.db
Magenheimer JF, Moore TR, Chmura GL, Daoust RJ (1996) Methane and carbon dioxide flux from a macrotidal salt marsh, Bay of fundy, New Brunswick. Estuaries 19:139–145. doi:10.2307/1352658
Minoda T, Kimura M, Wada E (1996) Photosynthates as dominant source of CH4 and CO2 in soil water and CH4 emitted to the atmosphere from paddy fields. Journal of Geophysical Research 101:21091–21097. doi:10.1029/96JD01710
Nykänen H, Alm J, Silvola J, Tolonen K, Martikainen PJ (1998) Methane fluxes on boreal peatlands of different fertility and the effect of long-term experimental lowering of the water table on flux rates. Global Biogeochemical 12:53–69. doi:10.1029/97GB02732
Poffenbarger HJ, Needelman BA, Megonigal JP (2011) Salinity influence on methane emissions from tidal marshes. Wetlands 31:831–842. doi:10.1007/s13157-011-0197-0
Purvaja R, Ramesh R (2001) Natural and anthropogenic methane emission from coastal wetlands of south India. Environmental Management 27:547–557. doi:10.1007/s002670010169
Qin P, Zhong CX (1992) Applied studies on Spartina. Ocean Press, China. Rounsevell MDA, Reay DS (2009) Land use and climate change in UK. Land Use Policy 26: s160-s169. doi: 10.1016/j.landusepol.2009.09.007
Schrier-Uijl AP, Veraart AJ, Leffelaar PA, Berendse F, Veenendaal EM (2011) Release of CO2 and CH4 from lakes and drainage ditches in temperate wetlands. Biogeochemistry 102:265–279. doi:10.1007/s10533-010-9440-7
Silvennoinen H, Liikanen A, Rintala J, Martikainen PJ (2008) Greenhouse gas fluxes from the eutrophic Temmesjoki River and its Estuary in the Liminganlahti Bay (the Baltic Sea). Biogeochemistry 90:193–208. doi:10.1007/s10533-008-9244-1
Song CC, Zhang JB, Wang YY, Wang YS, Zhao ZC (2008) Emissions of CO2, CH4 and N2O from freshwater in northeast of China. Journal of Environmental Management 88:428–436. doi:10.1016/j.jenvman.2007.03.030
Song J, Chen M, Feng G, Jia YH, Wang BS, Zhang FS (2009) Effect of salinity on growth, ion accumulation and the roles of ions in osmotic adjustment of two populations of Suaeda salsa. Plant and Soil 314:133–141. doi:10.1007/s11104-008-9712-3
Søvik AK, Kløve B (2007) Emission of N2O and CH4 from a constructed wetland in southeastern Norway. Science of the Total Environment 380:28–37. doi:10.1016/j.scitotenv.2006.10.007
Striegl RG, Kortelainen P, Chanton JP, Wickland KP, Bugna GC, Rantakori M (2001) Carbon dioxide partial pressure and 13C content of north temperate and boreal lakes at spring ice melt. Limnology and Oceanography 46:941–945
Ström L, Mastepanov M, Christensen TR (2005) Species-specific effects of vascular plants on carbon turnover and methane emissions from wetlands. Biogeochemistry 75:65–82. doi:10.1007/s10533-004-6124-1
Sun ZG, Wang LL, Tian HQ, Jiang HH, Mou XJ, Sun WL (2013) Fluxes of nitrous oxide and methane in different coastal Suaeda salsa marshes of the Yellow River estuary, China. Chemosphere 90:856–865. doi:10.1016/j.chemosphere. 2012. 10.004
Sun ZG, Wang LL, Mou XJ, Jiang HH, Sun WL (2014) Spatial and temporal variations of nitrous oxide flux between coastal marsh and the atmosphere in the Yellow River estuary of China. Environmental Science and Pollution Research 21:419–433. doi:10.1007/s11356-013-1885-5
Teiter S, Mander Ü (2005) Emission of N2O, N2, CH4, and CO2 from constructed wetlands for wastewater treatment and from riparian buffer zones. Ecological Engineering 25:528–541. doi:10.1016/j.ecoleng.2005.07.011
Tong C, Wang WQ, Zeng CS, Marrs R (2010) Methane (CH4) emission from a tidal marsh in the Min River estuary, southeast China. Journal of Environmental Science and Health Part A 45:506–516
Tong C, Wang WQ, Huang JF, Gauci V, Zhang LH, Zeng CS (2012) Invasive alien plants increase CH4 emissions from a subtropical tidal estuarine wetland. Biogeochemistry 111:677–693. doi:10.1007/s10533-012-9712-5
Tremblay A, Therrien J, Hamlin B, Wichmann E, LeDrew LJ (2005) GHG emissions from boreal reservoirs and natural aquatic ecosystems. In greenhouse gas emissions-fluxes and processes. Springer, Berlin, pp 209–232
Van Bodegom PM, Scholten J (2001) Microbial processes of CH4 production in a rice paddy soil: model and experimental validation. Geochimica et Cosmochimica Acta 65:2055–2066. doi:10.1016/S0016-7037(01)00563-4
Van Der Nat FJ, Middelburg JJ (2000) Methane emission from tidal freshwater marshes. Biogeochemistry 49:103–121. doi:10.1023/A:1006333225100
Walker LR, Smith SD (1997) Impacts of invasive plants on community and ecosystem properties. In assessment and management of plant invasions. Springer, New York, pp 69–86
Wang FY, Liu RJ, Lin XG, Zhou JM (2004) Arbuscular mycorrhizal status of wild plants in saline-alkaline soils of the Yellow River delta. Mycorrhiza 14:133–137. doi:10.1007/s00572-003-0248-3
Wang Q, An SQ, Ma ZJ, Zhao B, Chen JK, Li B (2006a) Invasive Spartina alterniflora: biology, ecology and management. Acta Phytotaxonomica Sinica 44:559–588. doi:10.1360/aps06044
Wang Q, Wang CH, Zhao B, Ma ZJ, Luo YQ, Chen JK, Li B (2006b) Effects of growing conditions on the growth of and interactions between salt marsh plants: implications for invasibility of habitats. Biological Invasions 8:1547–1560. doi:10.1007/s10530-005-5846-x
Wang YH, Yang JM, Zhang ML, Song XJ, Wang WJ, Sun GH (2013) The study of biodiversity of micro-phytoplankton in sea cucumber aquaculture ponds of Yellow River delta. Qceanologia Et Limnologia Sinica 44:415–420
Watanabe A, Takeda T, Kimura M (1999) Evaluation of origins of CH4 carbon emitted from rice paddies. Journal of Geophysical Research: Atmospheres (1984–2012) 104:23623–23629. doi:10.1029/1999JD900467
Whalen SC (2005) Biogeochemistry of methane exchange between natural wetlands and the atmosphere. Environmental Engineering Science 22:73–94. doi:10.1089/ees.2005.22.73
Whiting GJ, Chanton JP (2001) Greenhouse carbon balance of wetlands: methane emission versus carbon sequestration. Tellus B 53:521–528. doi:10.1034/j.1600-0889.2001.530501.x
Xing YP, Xie P, Yang H, Ni LY, Wang YS, Rong KW (2005) Methane and carbon dioxide fluxes from a shallow hypereutrophic subtropical Lake in China. Atmospheric Environment 39:5532–5540. doi:10.1016/j.atmosenv.2005.06.010
Xu XG, Guo HH, Chen XL, Lin HP, Du QL (2002) A multi-scale study on land use and land cover quality change: the case of the Yellow River delta in China. GeoJournal 3:177–183. doi:10.1023/A:1025175409094
Yang P, Tong C, He QH, Huang JF (2012) Diurnal variations of greenhouse gas fluxes at the water-air interface of aquaculture ponds in the Min River estuary. Environmental Science 33:4194–4204
Zhang YH, Ding WX, Cai ZC, Valerie P, Han FX (2010) Response of methane emission to invasion of Spatina alterniflora and exogenous N deposition in the coastal salt marsh. Atmospheric Environment 44:488–4594. doi:10.1016/j. atmosenv. 2010.08.012
Zhang LH, Song LP, Zhang LW, Shao HB (2014) Diurnal dynamics of CH4, CO2 and N2O fluxes in the saline-alkaline soils of the Yellow River delta, China. Plant Biosystems-An International Journal Dealing with all Aspects of Plant Biology. doi:10.1080/11263504.2013.870937
Zhu SW, Pan XL, Li XQ, Liu C (2012) Effects of exotic Spartina anglica on ecological environment of the Yellow River delta. Shandong Agricultural Sciences 44(73–75):83
Acknowledgments
The authors would like to acknowledge the financial support of the National Basic Research Program of China, (No. 2013CB430401).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Song, H., Liu, X. Anthropogenic Effects on Fluxes of Ecosystem Respiration and Methane in the Yellow River Estuary, China. Wetlands 36 (Suppl 1), 113–123 (2016). https://doi.org/10.1007/s13157-014-0587-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13157-014-0587-1