
Abstract
The efficacy of three entomopathogenic nematode (EPN) species, Heterorhabditis indica, Steinernema thermophilum, and S. glaseri, from Meghalaya, India was studied against the larvae of taro leaf beetle, Aplosonyx chalybaeus (Hope) (Coleoptera: Chrysomelidae), under the laboratory conditions. The beetle larvae (grubs) were exposed to 25, 50, 75, 100 and 200 infective juveniles (IJs) of each nematode species for different time periods and they were found to be susceptible to all the EPNs tested. However, the susceptibility of grubs to nematode infection varied according to the dosages of IJs and their exposure periods. Appreciably good performance was achieved by S. glaseri, which showed 100 % mortality of insect larvae in 48 h exposure time. At 48 h of incubation, its LC50 value was 90.3 IJs/larva, which was lower than that of S. thermophilum (115.0 IJs/larva) and H. indica (186.0 IJs/larva), at the same exposure time. All the tested nematode species were also found to reproduce within the host and produced infective juveniles. H. indica, however, showed comparatively more production of IJs per cadaver of infected host (168.9 × 103 IJs/larva), as compared to the other two tested nematode species. The production of IJs per cadaver of infected host by S. thermophilum was recorded to be 82.0 × 103 IJs/larva. In case of S. glaseri, while production of IJs increased initially to 18.9 × 103 IJs/larva at concentration of 100 IJs/larva, it declined thereafter to 14.7 × 103 IJs/larva at the dose of 200 IJs/larva. In conclusion, the evidence obtained in this study suggests that all the three indigenous EPN species are virulent enough to produce 100 % mortality in the last instar larvae of A. chalybaeus. These EPN species thus have potential scope for the management of A. chalybaeus in taro crops.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Taro (Colocasia esculenta (L.) Schott) is an important staple crop in developing countries, especially in African and Southeast Asian countries. It is widely cultivated in South Africa, Asia, Oceania, Central Africa, West Indies and the islands of the Caribbean and Central America (Chandra 1984). Both, the wild and cultivated forms of taro are found in India (Kuruvilla and Singh 1981). All the plant parts, i.e. the leaves, petioles, corms and cormels are eaten in some or other forms in different regions of the world (Chandra 1984; Onwueme 1999). It is also considered as an important source of food during lean period, and feed for livestocks. Taro is susceptible to attack by about two dozens of pathogens and pests, but only a few causes serious reduction in its growth and production (Ooka 1990). In the Northeastern region of India, a chryomelid beetle, Aplosonyx chalybaeus (Hope) (Coleoptera: Chrysomelidae) is a serious pest of taro which damages its roots and corms (Swamy et al. 2002; Sanwal 2008). Elsewhere in the East Asia, these beetles have also been reported as deleterious pest of this crop in Vietnam and the Philippines (Lazell et al. 1991; Szinicz et al. 2005). The adult beetle feeds on the foliage of the host plant and produces several small circular holes. Whereas, the larvae, commonly called chrysomelid corm borer, are found in the soil around the damaged plant, where they feed mainly on corms, making numerous holes and tunnels. The diseased corms possess numerous holes and tunnels that not only make them unmarketable in the local markets, but also unfit for human or livestock consumption. Although chemical insecticides, such as carbofuran, can be used in the management of pest, there are many concerns because of contamination of the corns and adverse effects of chemical pesticides on the human health and environment. Therefore, some suitable and environment friendly methods for the management of pest is urgently required.
Entomopathogenic nematodes (EPNs) in the families Steinernematidae and Heterorhabditidae are the potential alternatives to chemical control for many economically important insect pests (Georgis 1990; Kaya and Gaugler 1993). The infective third stage juveniles (IJs) of these nematodes, found in the soil, carries mutualistic bacteria (Xenorhabdus spp. and Photorhabdus spp.) in their intestine, for steinernematids and heterorhabditids, respectively (Poinar 1979). Upon finding a susceptible host, the IJs enter the host’s body through spiracles, mouth and anus, and release the specific symbiont bacteria that kill the host by septicemia, usually within 24–48 h (Kaya and Gaugler 1993). Many species of EPNs belonging to both genera have been used with variable success as biocontrol agents against a number of insect pests occupying different habitats. However, considerable success has been achieved against soil-dwelling pests or pests in cryptic habitats (Begley 1990; Klein 1990; Williams and Walters 1999; Tomalak et al. 2005; Valle et al. 2008). Therefore, many studies have highlighted the usefulness of EPNs as suitable biological control agents against soil-dwelling pests (Hominick and Reid 1990). In this connection, researchers have reported that while developing a long-term strategy for the inundative release of a nematode species against pest species, it is always advisable to employ the indigenous strains of EPNs as they are comparatively better adapted to the local climate and also host population, as compared to the exotic EPN strains (Gaugler 1988; Bedding 1990). Further, it has also been argued that an ideal EPN strain must also have a good pathogenicity and should be able to propagate well on a large scale (Bedding 1990). Our recent studies on prevalence of EPNs in Meghalaya (India) have yielded three indigenous strains of nematodes (Lalramliana 2007), which we believe may be potentially effective against local pest species. Therefore, the aim of the present study was to evaluate the efficacy of these three locally isolated EPN strains, namely Heterorhabditis indica Poinar, Karunakar and David, Steinernema thermophilum Ganguly and Singh, and S. glaseri (Steiner) against the last instar larva of taro beetle, Aplosonyx chalybaeus. The virulence of EPN species to the grubs and their reproduction in host species was studied under the laboratory conditions.
Materials and methods
Nematode sources
The three strains of EPNs, H. indica, Karunakar and David, S. thermophilum Ganguly and Singh, and S. glaseri (Steiner), used in this study were isolated from forest soils in Meghalaya. These EPN strains have been characterized in previous studies, using the late instar larvae of the greater wax moth, Galleria mellonella Linnaeus as host (Lalramliana, 2007). For experimental study, the nematodes were reared in the laboratory on late instar larvae of G. mellonella at 25 °C, as described by Woodring and Kaya (1988). The IJs that emerged from wax moth larval cadavers were collected using modified White traps (Kaya and Stock 1997), and stored in darkness at 15 °C in deionized water. Before being used for assay, the IJs were allowed to acclimatize for 1 h at room temperature and their viability was checked by observation of movements under a stereomicroscope.
Insect sources
Last instar larvae of A. chalybaeus were obtained from the experimental farms of Indian Council of Agriculture Research, Shillong, India and maintained in the laboratory on natural diets. The collected larvae were kept for at least 5 days in the laboratory to check, whether or not, there are any other infections before using them for experiments. During this period, larvae were observed for feeding behavior and movements, and only the healthy looking larvae were included in the study.
Larval mortality bioassay
Larval mortality bioassays were carried out in Petri dishes (35 × 10 mm) lined with double layer of Whatman No. 1 filter paper, following the methods of Kaya and Stock (1997). Nematodes in 0.5 ml of deionized water were added to the filter paper in concentrations of 25, 50, 75, 100 and 200 IJs/larva. After 30 min, a single larva of A. chalybaeus was placed in each of the Petri dish. The dishes were sealed with parafilm and maintained in a climatic chamber at 27 ± 2 °C in the dark. For each nematode species and concentration there were eight replicates and the experiment was repeated thrice. Untreated controls were identical to the treatment except that no IJs were added. Larval mortality was checked at every 24 h for up to 120 h. The cause of larval death was confirmed by body colour change of the cadaver which being evident due to the presence of symbiotic bacteria.
Reproduction of EPNs
Last instar larvae of A. chalybaeus were exposed to 25, 50, 75, 100 and 200 IJs concentrations of each EPN in separate Petri dishes and total number of IJs produced per larva for up to a period of 20 days was counted. In brief, the nematode-infected dead insect larvae were removed from dishes, rinsed in deionized water and transferred individually on to White traps for their emergence from the body (White 1927). The larvae were collected daily for up to a period of 20 days, till the emergence of IJs was stopped from insect cadavers and total number of IJs produced per larva was then determined. There were eight replicates for each nematode species and concentration and the experiment was repeated thrice. To each concentration, one Petri dish, prepared as described above but without IJs served as control.
Statistical analysis
All experimental data were analyzed statistically and are presented as mean ± standard error of mean (SEM). The significance of the difference in all bioassay experiments was determined by one way analysis of variance (ANOVA) and Student’s t test. P values <0.05 were accepted as statistically significant. Correlation between the parameters was determined by regression analysis. LC50 values were calculated using Probit analysis with SPSS software.
Results and discussion
The aim of this study was to evaluate the efficacy of three species of EPNs, namely H. indica, S. thermophilum and S. glaseri, against the last instar larvae of A. chalybaeus, a serious pest of taro in the Northeastern region of India. Each nematode species was evaluated for its virulence on the basis of dose and time required by nematode to kill the insect larvae and also on the basis of nematode’s ability to propagate within the body of infected host and produce infective juveniles.
As shown in Fig. 1, the larvae of A. chalybaeus were found to be susceptible to all the three tested EPNs. However, the degree of susceptibility of insect larvae to nematode infection varied according to the doses of their infective juveniles as well as the exposure period. Also, a positive correlation was found between the doses of infective juveniles applied and time of larval mortality for all the three tested EPN species. A moderate larval mortality was observed due to infection with S. thermophilum in 24 h exposure time at 200 IJs/insect (Fig. 1b), whereas, H. indica and S. glaseri caused larval mortality only after 48 h of incubation time (Fig. 1a, c). After 48 h of incubation, both S. thermophilum and S. glaseri, at 200 IJs/insect, caused cent percent larval mortality, while H. indica gave comparatively low larval mortality after first 48 h incubation time. A further increase in incubation time to 72 h led to a complete mortality of insect larvae by S. glaseri only at a dose of 100 IJs/insect. However, at the same dose and incubation time, H. indica gave about 65 % of larval mortality. Larval mortality due to H. indica, however, continued to increase further with an increase in exposure time and 100 % larval mortality was recorded only after 120 h at 100 IJs/insect. The calculated values of LC50 are presented in Table 1. The increase in exposure time resulted in reduction of the values of LC50 for all nematodes. Among the EPNs tested, S. glaseri was the most virulent with minimum values of LC50 at all exposure time. After 48 h of incubation, S. glaseri showed a LC50 value of 90.3 IJs/larva (95 % fiducial limit (FL): 72.5–141.3) which was lower than that of S. thermophilum (115.0, 95 % FL: 92.3–162.7). At the same exposure time, H. indica was comparatively less virulent with the LC50 value of 186.0 IJs/larva (95 % FL: 122.9–212.7). A number of workers have used these assays to adjudge the efficacy of EPNs against various insect pests (Ricci et al. 1996; Sims et al. 1992; Bhatnagar et al. 2004). In the present study, the beetle larvae were found to be susceptible to all the three tested EPNs. Also, it emerged that there exists a positive correlation between the dose of IJs and host mortality for all the three nematodes. Among all nematodes, S. glaseri was found to be the most virulent species. There has been no study to date on the efficacy of entomopathogenic nematodes against the larvae of taro beetle, A. chalybaeus. In general, the findings of the present study on efficacy of EPNs against A. chalybaeus larvae are in agreement with other insect hosts. For example, Hominick and Reid (1990) reported that, besides the intrinsic qualities of the species, EPN efficiency is greatly influenced by its dose. In a similar manner, researchers have also recorded a positive relationship between nematode concentration and host mortality in many previous studies (Forschler and Nordin 1988; Glazer and Navon 1990; Peters and Ehlers 1994). The present findings on virulence of S. glaseri are in tune with Toepfer et al. (2005), who also reported this species to be the most virulent one among all the studied EPNs against western corn rootworm, Diabrotica virgifera virgifera. Further, Karunakar et al. (1999) also recorded almost similar trends in the parasitism of larvae of white grub at different doses of S. glaseri and H. indica. Differences in infectivity between nematode species or strains have been also been documented for many other insect species (Forschler and Nordin 1988; Griffin et al. 1989). This study also confirms the results of previous studies made by Converse and Grewal (1998) and Seth and Barik (2009) who demonstrated that S. glaseri is much more virulent nematode than H. indica. The susceptibility of soil-inhabiting white grubs to S. glaseri was first demonstrated by Glaser (1932). In that study, too, S. glaseri was reported to be more effective than other steinernematids and heterorhabditids against scarab larvae. The pathogenicity of EPNs depends upon many biotic and abiotic factors like host invasion, penetration, reproduction, etc. (Kaya and Gaugler 1993). Thus different nematode species have been found to differ in their pathogenicity against a specific insect host (Forschler and Nordin 1988; Griffin et al. 1989). In many investigations, the virulence of EPNs has also been found to be dependent on other factors such as host invasion and penetration ability of the nematode infective juveniles (Gaugler 1988; Lewis et al. 1992; Glazer et al. 2001). For example, while studying the pathogenicity and penetration of five EPNs against sugarcane internode borer, Sankaranarayanan et al. (2008) reported a higher penetration ability and pathogenicity of S. glaseri than H. indica. A good virulence of S. glaseri, therefore, may also be explained in part due to its efficient penetration ability into the insect host.

Percentage mortality of A. chalybaeus larvae following exposure to different concentrations of infective juveniles (IJs) of nematodes. a H. indica, b S. thermophilum and c S. glaseri
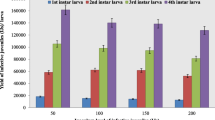
In order to study the reproductive potential of EPNs, the insect larvae were exposed to different concentrations of IJs of the three test nematodes. Following host mortality, the emerging IJs were collected from host cadavers and counted. The data revealed that all the three species were able to invade and propagate within the host and produce infective juveniles (Fig. 2a–c). It was also evident that, except S. glaseri, the other two nematode species exhibited a linear relationship between the concentrations of IJs applied and total number of infective juveniles produced per infected larva. In this study, H. indica produced significantly more number of infective juveniles per insect larva as compared to other tested nematodes (Fig. 2a). At the dose of 200 IJ/larva, the progeny production in H. indica was recorded to be 168.9 × 103 IJs/larva. This was followed by S. thermophilum, which produced 82.0 × 103 IJs/larva (Fig. 2b). In case of S. glaseri though progeny production initially increased with the increase in IJs dose, it declined thereafter to 14.7 × 103 IJs/larva at 200 IJs/larva (Fig. 2c).

Progeny production by insect larvae at different dosages of infective juveniles (IJs) of entomopathogenic nematode species. a H. indica, b S. thermophilum and c S. glaseri. Significant at ** 0.01 and * 0.05 %, (a–d) Mean values shown by different letters are significantly different at 0.05 %
Reproduction and recycling of entomopathogenic nematodes in host play an important role in their persistence in the soil, and also in their overall effectiveness in pest control (Harlan et al. 1971; Georgis and Hague, 1981). A prior knowledge about reproduction and recycling of nematode is considered important in determining the time and dose of subsequent EPN application, which may be useful in reducing the cost of application. The data in this study suggest that following application, all the tested species of nematodes were able to infect and propagate within the insect host and produce infective juveniles. However, the highest production of IJs was obtained with H. indica, which yielded 168.9 × 103 IJs/larva at 200 IJs post exposure time, followed by S. thermophilum which yielded 82.0 × 103 IJs/larva at the same dose and exposure period. In case of S. glaseri, though the production of IJs increased initially with increase in IJs dose, it declined thereafter to 14.7 × 103 IJs/larva at the dose of 200 IJ/larva. Differences in reproduction (i.e., production of F1 generation infective juveniles) between nematode species have been documented for many insect hosts (Ali et al. 2006; Karunakar et al. 1999). The size and behaviour of nematode species may account for differences in nematode ability to reproduce in the host (Loya and Hower 2003). Bhatnagar et al. (2004) also reported that H. bacteriophora being the smaller in size, produces more IJs per cadaver of infected host than S. glaseri, which is larger in size, against the final instar grubs of Maladera insanabilis. Our findings on production of IJs by EPNs are also in agreement with the results of Jothi and Mehta (2006), where H. bacteriophora was recorded to produce comparatively more IJs per infected insect larva than S. glaseri (Stuart et al. 1996). A higher production of IJs by H. indica in the present study may also be attributed, in part, to the fact that heterorhabditids being hermaphroditic are likely to contribute to more progeny than steinernematids which are amphimictic (Poinar 1990; Mannion and Jansson 1992).
In conclusion, the evidence obtained in this study suggests that all the three tested indigenous species of EPNs are virulent enough to produce 100 % mortality to the larvae of A. chalybaeus. Furthermore, all EPNs can also propagate in the infected larva and produce F1 generation infective juveniles. Considering these attributes, it could be suggested that these EPNs have potential use as biocontrol agents for the management of A. chalybaeus in taro crops.
References
Ali SS, Perveza R, Hussain MA, Ahmad R (2006) Susceptibility of three lepidopteran pests to five entomopathogenic nematodes and in vivo mass production of these nematodes. Arch Phytopathol Pfl 41:300–304
Bedding RA (1990) Logistics and strategies for introducing entomopathogenic nematode technology in developing countries. In: Gaugler R, Kaya HK (eds) Entomopathogenic nematodes in biological control. CRC Press, Boca Raton, pp 233–248
Begley JW (1990) Efficacy against insects in habitats other than soil. In: Gaugler R, Kaya HK (eds) Entomopathogenic nematodes in biological control. CRC Press, Boca Raton, pp 215–231
Bhatnagar A, Shinde V, Bareth SS (2004) Evaluation of entomopathogenic nematodes against white grub, Maladera insanabilis Brenske. Int J Pest Manag 50:285–289
Chandra S (1984) Edible aroids. Clarendon Press, Oxford
Converse V, Grewal PS (1998) Virulence of entomopathogenic nematodes to the western masked chafer Cyclocephala hirta (Coleoptera: Scarabaeidae). J Econ Entomol 91:428–432
Forschler BT, Nordin GL (1988) Comparative pathogenicity of selected entomogenous nematodes to the hardwood borers, Prionoxystus roblniae (Lepidoptera: Cossidae) and Megacylletze vobiniae (Coleoptera: Cerambycidae). J Invertebr Pathol 52:343–347
Gaugler R (1988) Ecological considerations in the biological control of soil-inhabiting insects with entomopathogenic nematodes. Agr Ecosyst Environ 24:351–360
Georgis R (1990) Formulation and application technology. In: Gaugler R, Kaya HK (eds) Entomopathogenic nematodes in biological control. CRC Press, Boca Raton, pp 173–191
Georgis R, Hague NGM (1981) A neoaplectanid nematode in the larch sawfly Cephalcia lariciphila (Hymenoptera: Pamphiliidae). Ann Appl Biol 99:171–177
Glaser RW (1932) Studies on Neoaplectana glaseri, a nematode parasite of the Japanese beetle (Papillia japonica). Technical Report 1932, Circular No. 211. New Jersey Department of Agriculture, Trenton
Glazer I, Navon A (1990) Activity and persistence of entomoparasitic nematodes tested against Heliothis avmigeva (Lepidoptera: Noctuidae). J Econ Entomol 83:1795–1800
Glazer I, Alekseev E, Samish M (2001) Factors affecting the virulence of entomopathogenic nematodes to engorged female Boophilus annulatus ticks. J Parasitol 87:808–812
Griffin CT, Simons WR, Smits PH (1989) Activity and infectivity of four isolates of Heterorhabditis spp. J Invertebr Pathol 53:107–112
Harlan DP, Dutky SR, Padgett GR, Mitchell JA, Shaw ZA, Barlett FJ (1971) Parasitism of Neoaplectana dutkyi in white-fringed beetle larvae. J Nematol 3:280–283
Hominick WM, Reid AP (1990) Perspectives on entomopathogenic nematology. In: Gaugler R, Kaya HK (eds) Entomopathogenic nematodes in biological control. CRC Press, Boca Raton, pp 327–345
Jothi DB, Mehta UK (2006) Pathogenicity of three species of EPN against cotton bollworm Helicoverpa armigera Hub. Entomon 31:259–266
Karunakar G, Easwaramoorthy S, David H (1999) Susceptibility of nine lepidopteran insects to Steinernema glaseri, S. feltiae and Heterorhabditis indicus infection. Int J Nematol 9:68–71
Kaya HK, Gaugler R (1993) Entomopathogenic nematodes. Annu Rev Entomol 38:181–206
Kaya HK, Stock P (1997) Techniques in insect nematology. In: Lacey LA (ed) Manual of techniques in insect pathology. Academic Press, San Diego, pp 281–324
Klein MG (1990) Efficacy against soil-inhabiting insect pests. In: Gaugler R, Kaya H (eds) Entomopathogenic nematodes in biological control. CRC Press, Boca Raton, pp 195–214
Kuruvilla KM, Singh A (1981) Karyotypic and electrophoretic studies on taro and its origin. Euphytica 30:405–413
Lalramliana (2007) Faunal composition and distribution of entomopathogenic nematodes and their bioefficacy against major insect pests in Ri-Bhoi District of Meghalaya. PhD thesis, North-Eastern Hill University, Shillong
Lazell JD, Keirans JE, Samuelson GA (1991) The Sulawesi Black Racer, Coluber (Ptyas) dipsas, and a remarkable ectoparasitic aggregation. Pac Sci 45:355–361
Lewis EE, Gaugler R, Harrison R (1992) Entomopathogenic nematode host finding: response to host contact cues by cruise and ambush foragers. Parasitology 105:309–315
Loya LJ, Hower AA Jr (2003) Infectivity and reproductive potential of the Oswego strain of Heterorhabditis bacteriophora associated with life stages of the clover root curculio, Sitona hispidulus. J Invertebr Pathol 83:63–72
Mannion CM, Jansson RK (1992) Comparison of ten entomopathogenic nematodes for control of sweet potato weevil (Coleoptera: Apionidae). J Econ Entomol 85:1642–1650
Onwueme I (1999) Taro cultivation in Asia and Pacific. RAP Publication 1999/16. Food and Agriculture Organization Department of United Nations, Regional Office for Asia and the Pacific, Bangkok
Ooka JJ (1990) Taro diseases. In: Hollyer JR, Sato DM. (Eds). Proceedings of taking Taro into the 1990s: a Taro conference. Aug 17 1989.Hilo, University of Hawaii,Honolulu, pp 51–59
Peters A, Ehlers RU (1994) Susceptibility of leatherjackets (Tipula paludosa and Tipula oleracea; Tipulidae; Nematocera) to the entomopathogenic nematode Steinernema feltiae. J Invertebr Pathol 63:163–171
Poinar GO Jr (1979) Nematodes for biological control of insects. CRC Press, Boca Raton, p 270
Poinar GO Jr (1990) Taxonomy and biology of Steinernematidae and Heterorhabditidae. In: Gaugler R, Kaya HK (eds) Entomopathogenic nematodes in biological control. CRC Press, Boca Raton, pp 23–61
Ricci M, Glazer I, Campbell JF, Gaugler R (1996) Comparison of bioassays to measure the virulence of different entomopathogenic nematodes. Biocontrol Sci Technol 6:235–245
Sankaranarayanan C, Somasekhar N, Singaravelu B, Shanmugasundaram M (2008) Pathogenicity of entomopathogenic nematodes to sugarcane internode borer, Chilo sacchariphagus indicus Kapur (Lepidoptera: Crambidae). J Biol Control 22:167–171
Sanwal SK (2008) Underutilized vegetable and spice crops. Agrobios, Delhi
Seth RK, Barik TK (2009) Assessment of infective behaviour and reproductive potential over successive generations of entomopathogenic nematodes, Steinernema glaseri (Rhabditida: Steinernematidae), reared within radiosterilized host larvae, towards Spodoptera litura (Lepidoptera: Noctuidae). Biocontrol Sci Technol 19:111–125
Sims SR, Downing AA, Pershing JC (1992) Comparison of assays for determination of entomogenous nematode infectivity. J Nematol 24:271–274
Stuart RJ, Lewis EE, Gaugler R (1996) Selection alters the pattern of emergence from the host cadaver in the entomopathogenic nematode, Steinernema glaseri. Parasitology 113:183–189
Swamy TMS, Sriram S, Byju G, Misra RS (2002) Tropical tuber crops production in northeastern India: pests, diseases and soil fertility constraints. J Root Crops 28:64–68
Szinicz G, Martin K, Sauerborn J (2005) Abundance of selected insect species in natural and agricultural habitats of a tropical upland (Leyte, Philippines). Agr Ecosyst Environ 111:104–110
Toepfer S, Gueldenzopha C, Ehlers R-U, Kuhlmanna U (2005) Screening of entomopathogenic nematodes for virulence against the invasive western corn rootworm, Diabrotica virgifera virgifera (Coleoptera: Chrysomelidae) in Europe. Bull Entomol Res 95:473–482
Tomalak M, Piggott S, Jagdale GB (2005) Glasshouse applications. In: Grewal PS, Ehlers R-U, Shapiro-Ilan DI (eds) Nematodes as biocontrol agents. CABI Publishing, Oxon, pp 147–166
Valle EED, Dolinski C, Souza RM (2008) Dispersal of Heterorhabditis baujardi LPP7 (Nematoda: Rhabditida) applied to the soil as infected host cadavers. Int J Pest Manag 54:115–122
White GF (1927) A method for obtaining infective nematode larvae from cultures. Science 66:302–303
Williams EC, Walters KFA (1999) Foliar application of entomopathogenic nematode Steinernema feltiae against leaf miners on vegetables. Biocontrol Sci Techn 10:61–70
Woodring JL, Kaya HK (1988) Steinernematid and heterorhabditid nematodes: a handbook of biology and techniques. Southern Cooperative Series Bulletin 331. Arkansas Agricultural Experiment Station, Arkansas
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Yadav, A.K., Lalramliana Efficacy of indigenous entomopathogenic nematodes from Meghalaya, India against the larvae of taro leaf beetle, Aplosonyx chalybaeus (Hope). J Parasit Dis 36, 149–154 (2012). https://doi.org/10.1007/s12639-012-0139-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12639-012-0139-7