
Abstract
The extinct Eopelobates (Eocene of western North America; Eocene–Pliocene of Europe) and Pelobates (Oligocene–Recent of Europe; Recent of northern Africa and the Middle East) are superficially toad-like anurans that are united within the family Pelobatidae mainly on the basis of a unique, tripartite frontoparietal complex. Both genera have a relatively good fossil record consisting of isolated bones, skeletons, and developmental series of tadpoles through adults, all of which are potentially informative for tracing the evolutionary history of the family. Eopelobates is of interest for several reasons. Of the two pelobatid genera, Eopelobates appears earlier in the fossil record (early Eocene vs. late Oligocene) and it is more primitive in lacking many of the features associated with fossoriality in extant Pelobates. The taxonomic composition of Eopelobates has been contentious and at least one putative new species has long been recognised, but never formally named. Here, we provide updated taxonomic accounts for Pelobatoidea, Pelobatidae, Pelobates, and Eopelobates and document development within a series of tadpoles and juveniles of E. bayeri from Bechlejovice (late Oligocene in age), Czech Republic. We also provide updated accounts for the five previously named and currently accepted species of Eopelobates. For the European congeners, E. anthracinus (late Oligocene) and E. bayeri (early Oligocene–middle Miocene) can confidently be regarded as separate species; although the distinction between E. hinschei and E. wagneri (both middle Eocene) is less certain, we provisionally maintain them as separate species. Micro-CT scans for the holotype skeleton of E. grandis (latest Eocene, USA) help resolve some problematic features, most notably showing that the cranial sculpture is of the pit-and-ridge style that is typical for Eopelobates. A sixth congener is named and described based on two skeletons from the middle Eocene portion of the Green River Formation, in Wyoming, USA. We caution that reports of Eopelobates-like anurans from the pre-Eocene of western North America and the early Eocene of India are based on isolated bones that cannot be assigned with confidence to that genus. The presence of Eopelobates in both North America and Europe may be explained by dispersal via the high latitude land bridge that connected those two continents during the late Paleocene through Eocene. The pelobatid fossil record is informative for documenting the nature and timing of changes in cranial features (e.g. ornament patterns, shape of nasals, pattern of frontoparietal–squamosal contact) from the inferred primitive condition seen in most Eopelobates to the more derived condition seen in extant Pelobates, but it is less informative for tracing the evolution of fossoriality, which is a key attribute of extant Pelobates.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
As is typical for many anuran families, Pelobatidae has a complex taxonomic history in which its content, geographic and temporal distribution, diagnostic features, and inferred relationships with other anurans has changed considerably since it was named by Bonaparte in 1850 (e.g. see reviews by Maglia 1998; Frost et al. 2006). Bonaparte (1850) diagnosed Pelobatidae exclusively on non-osteological features, such as vertical pupils and inguinal amplexus, and he included just three extant genera of toad-like frogs within the family: Pelobates from Europe (eastwards to the Ural Mts. and Ural River), northern Africa, and the Middle East; Scaphiopus from North America; and Megophrys from south-eastern Asia. In the same paper, Bonaparte (1850) considered the extant genus Pelodytes from western Europe and Caucasus distinctive enough to be assigned to its own family, which he called Pelodytidae. Those four genera are closely related (e.g. Duellman and Trueb 1994; Henrici 1994; Maglia 1998; Frost et al. 2006; Henrici et al. 2013), to the extent that, in some earlier classifications, they were placed together within the same family (e.g. Boulenger 1910; Noble 1931). The first skeletal features (arciferal shoulder girdle and procoelous vertebral centra) for Pelobatidae sensu Bonaparte (1850) were provided by Cope (1865). In subsequent years, the list of diagnostic features expanded to include, for example, fusion of the sacral vertebra with the urostyle, a keratinized metatarsal spade supported by a well-ossified prehallux, and sculptured dermal bones. The content of Pelobatidae also swelled, as new extant genera and species were named (many of those were Megophrys-like taxa from southern Asia) and as fossil genera and species were recognised. This expanded concept of Pelobatidae survived to the end of the twentieth century (e.g. Duellman and Trueb 1994; Sanchiz 1998), with the family by that time being subdivided into two or three subfamilies and containing more than a dozen genera and nearly 100 species. New insights from molecular analyses (e.g. García-París et al. 2003; Roelants and Bossuyt 2005; San Mauro et al. 2005), from larger scale cladistic analyses that used various combinations of osteological, larval, molecular, and other characters (e.g. Haas 2003; Frost et al. 2006), and from the fossil record (e.g. Henrici and Haynes 2006; Henrici et al. 2013) corroborated the idea that at least some of the previously recognised subfamilial groupings are natural assemblages, but as yet there is little consensus on the contents of those groups, their relative ranks, or their inter-relationships.
Here, we use the name Pelobatidae in a restrictive sense for the more inclusive taxon containing the genera Pelobates Wagler, 1830 and Eopelobates Parker, 1929. These two genera are widely considered to be closely related based on their shared presence of a tripartite frontoparietal complex that is formed by the paired frontoparietals and by a unique posterior bone (the so-called “posterior median element”). This trio of bones appears in later-stage tadpoles. In postmetamorphic individuals, the posterior median element fuses with the frontoparietals, and, in fully grown adults, the frontoparietals also fuse together. Thus, unlike in other anurans, in both Pelobates and Eopelobates the median line of contact between the left and right frontoparietals is prevented by the posterior median bone from reaching the posterior margin of the frontoparietal (e.g. Roček 1981; Maus and Wuttke 2002, 2004; Roček and Wuttke 2010).
The first fossil pelobatids were recognised in the mid-1800s. Pelobates decheni Troschel, 1861 was based on an articulated skeleton from the late Oligocene of Rott, near Bonn, Germany (Troschel 1861; Böhme et al. 1982). Since then, Pelobates fossils have been reported from many localities of middle Oligocene to Pleistocene age in Europe. At present, four exclusively fossil species (P. decheni, P. fahlbuschi, P. praefuscus, and P. sanchizi) and numerous specifically indeterminate occurrences are known (e.g. Sanchiz 1998; Roček and Rage 2000; Roček 2013; this study). The genus also contains four extant species: two in Europe, one in the Middle East, and one in northern Africa (Frost 2014). Three of those species (P. cultripes, P. fuscus, and P. syriacus) also have fossil occurrences (Sanchiz 1998; Roček 2013). The same upper Oligocene strata near Rott that yielded the holotype skeleton of P. decheni later yielded an anuran skeleton sufficiently different from Pelobates that it was described by Parker (1929) as the new genus and species Eopelobates anthracinus. Over the next 60 years, another nine species of Eopelobates were described, largely on the basis of articulated skeletons, from the Eocene–Miocene of Europe, the Eocene of North America, and the Late Cretaceous of Asia (e.g. Sanchiz 1998; Roček and Rage 2000; Roček 2013; this study). Taxonomic revisions have reduced the number of named Eopelobates species to four from Europe and one from North America (e.g. Roček 2013; this study). Specifically indeterminate bones reliably extend the range of the genus forward into the Pliocene in Europe. Elsewhere, putative Eopelobates bones have been reported from the early Eocene of India (Folie et al. 2013) and the Late Cretaceous to Eocene of western North America (e.g. Estes 1970; Golz and Lillegraven 1977; Gardner and DeMar 2013).
Pelobates and Eopelobates both have extensive fossil records consisting of isolated bones, skeletons, and ontogenetic series of tadpoles through to fully metamorphosed adults (e.g. Estes 1970; Špinar 1972; Roček 1981; Sanchiz 1998; Roček and Rage 2000; Maus and Wuttke 2002, 2004; Roček 2013; this study). Yet, for postmetamorphic individuals, the limited number of skeletons and the preponderance of isolated bones available for fossil species of Pelobates makes it challenging to interpret evolutionary trends and relationships within that genus. By contrast, Eopelobates is known by a greater number of skeletons. Each of the six Eopelobates species recognised by us is documented by at least one skeleton and several, such as E. bayeri Špinar, 1952 and E. wagneri (Weitzel, 1938), are known by multiple skeletons, some exquisitely preserved (e.g. Špinar 1952, 1972; Wuttke 2012b).
Despite the relatively large samples of Eopelobates skeletons now available, numerous questions and uncertainties remain, such as: (1) whether two pairs of European species (E. anthracinus vs. E. bayeri; E. hinschei vs. E. wagneri) are truly distinct at the specific level; (2) whether the North American Eocene species E. grandis Zweifel, 1956 can be retained within Eopelobates, despite its reportedly unusual cranial sculpture; (3) whether an unamed, Eopelobates-like anuran from the Green River Formation (Eocene, USA) can be assigned to the genus, as several authors have suggested (e.g. Grande 1984; Henrici 2002, Holman 2003); (4) whether isolated Eopelobates-like bones reported from the Late Cretaceous to Eocene of North America and from the early Eocene of India can confidently be assigned to that genus; and (5) what are the relationhips among the diagnosable species of Eopelobates?
In this paper, we address some of the above points by providing updated taxonomic accounts for Pelobatoidea, Pelobatidae, Pelobates, and Eopelobates. Although the focus of our paper is on Eopelobates, accounts for its sister genus Pelobates and the larger groups to which those genera belong (i.e. Pelobatidae and Pelobatoidea) are necessary to provide some context for Eopelobates and to highlight how it differs from Pelobates. For Eopelobates, we also document development within a series of tadpoles and juveniles of E. bayeri from the upper Oligocene locality of Bechlejovice, Czech Republic. For the species of Eopelobates, we provide taxonomic and, where relevant, descriptive accounts for the five currently accepted species. Our account for E. grandis relies on micro-CT scans (the first published for a fossil pelobatid) that help clarify the pattern of cranial sculpture in the holotype and only available specimen. Additionally, we formally name and describe a sixth species of Eopelobates on the basis of two skeletons from the Green River Formation (middle Eocene in age) of Wyoming, USA. In the final part of our paper, we evaluate problematic reports of Eopelobates-like anurans in the Eocene of India and pre-Eocene of North America, provide a possible explanation for the disjunct distributions of Eopelobates in Europe and North America during the Eocene, and comment on what the fossil record can and cannot tell us about trends in cranial structure and sculpture and in fossoriality within Pelobatidae. Although we do not do so in this paper, much of the information presented here could be useful for attempting a cladistic analysis of Eopelobates and for testing our ideas about palaeobiogeography and evolutionary trends within Pelobatidae.
Materials and methods
Although we are aware that taxa above the species level (genus, family, superfamily, etc.) have no biological meaning, we provide diagnoses for them. Diagnoses for supraspecific taxa and for Eopelobates species are presented in a format that essentially is a list of characters deemed to be important or useful in classification (although not all are necessarily diagnostic except when considered in combination; in contrast, some features appear to have diagnostic value on their own). For ease of use, we have attempted to list the relevant cranial and postcranial characters in a standardised order within each diagnosis. Ideally, such lists of diagnostic features should be limited to those that represent differences between species. In contrast to extant taxa, however, diagnoses of extinct taxa, which are often represented by incomplete specimens, cannot always be complete.
Collectively, we have examined firsthand all of the Eopelobates specimens reported in this study. These include original specimens, plus some peels and replicas. Most of the specimens used in our study were examined visually under dissecting microscopes at low magnifications. The holotype skeleton of Eopelobates grandis was also examined using micro-CT scans. That specimen was scanned by one of us (B-ASB) on a Nikon X-Tek 225 kV cabinet system with a tungsten target and the source set at 95 kV and 90 mA. A total of 3,200 views were collected on the 2,048 × 2,048 detector, averaging 2 frames, with an exposure time of 1 s. Images of the 3D volume rendering were made in VGStudio Max 2.2 using the reconstructed data from the 32-bit float raw file with no downsampling, using the volume rendering algorithm with two light sources and shadows on.
We generally follow the morphological terminology of anuran bones introduced by Bolkay (1919).
Specimens used in this study
Note that unless stated otherwise, all are skeletons of metamorphosed individuals. Eopelobates anthracinus – holotype (NHMUK R4841); counterpart of the same specimen (GPIB-Ro 4029); GPIT 1733; tadpole (NHMUK PV OR49464). Eopelobates bayeri – holotype (NMP Pb 412); paratype (NMP Pb 1694, part, and NMP Pb 1114, counterpart); tadpoles (NMP Pb 227, NMP Pb 228, NMP Pb 350, NMP Pb 418, NMP Pb 430, NMP Pb 434, NMP Pb 439, NMP Pb 407, NMP Pb 440, NMP 443, NMP 444, NMP Pb 445, NMP Pb 449, NMP Pb 450, NMP Pb 1397, NMP Pb 1398, NMP Pb 1401, NMP Pb 1402, NMP Pb 1403, NMP Pb 1404, NMP Pb 1461, NMP Pb 1462, NMP Pb 1463, NMP Pb 1464, NMP Pb 1467, NMP Pb 1474, NMP Pb 1475, NMP Pb 1476, NMP Pb 1477, NMP Pb 1568, NMP Pb 1569, NMP Pb 1577, NMP Pb 1578, NMP Pb 1579, NMP Pb 1695, NMP Pb 1704, NMP Pb 1705, NMP Pb 1716, NMP Pb 1717, NMP Pb 1732, NMP Pb 1733, NMP Pb 1734, NMP Pb 1735, NMP Pb 1736, NMP Pb 1737). New species of Eopelobates holotype (BHM-123); epoxy resin casts or peels of the holotype (FMNH PR 1613, SMM P78.8.29, TMP 2013.05.16); paratype (SMNK PAL 6659a and b, part and counterpart, respectively). Eopelobates grandis – holotype (YPM-PU 16441). Eopelobates hinschei – holotype (GMH 1312, formerly holotype of Halleobatrachus hinschei); GMH 6728 (formerly holotype of Archaeopelobates eusculptus); GMH 6692; GMH 6753. Eopelobates wagneri – holotype (HLMD-Me 1286); paratype (HLMD-Me 4359). Macropelobates linquensis – neotype (IVPP V7700). Macropelobates osborni – holotype (AMNH 6252). Pelobates decheni – holotype (GPIB-Ro 4031). Pelobates cf. decheni – NHMM PW1995/5802a-LS, NHMM PW1997/5040. Pelobates fuscus – DP FNSP 5826, DP FNSP 5895, DP FNSP 5896, DP FNSP 5897, DP FNSP 6326, DP FNSP 6333, DP FNSP 6335, DP FNSP 6339, DP FNSP 6433, DP FNSP 6438. Pelobates varaldii – DP FNSP 6328a. Pelobates indet. – frontoparietal (IZANK 3338). Uldzinia kurochkini – holotype maxilla (PIN 3109/236).
Anatomical abbreviations
F – femur; NF – Nieuwkoop and Faber (1967) developmental stage; SVL – snout-vent length; TF – tibiofibula; V1–V8 – presacral vertebrae numbered anterior to posterior.
Institutional abbreviations
AMNH – American Museum of Natural History, New York, USA; BHM – Black Hills Museum (part of Black Hills Institute [BHI]), Hill City, South Dakota, USA; DP FNSP – Department of Paleontology, Faculty of Natural Sciences, Charles University, Prague, Czech Republic; FMNH – Field Museum of Natural History, Chicago, Illinois, USA; GMH – Geiseltalmuseum, Martin Luther Universität, Halle/Saale, Germany; GPIB – Steinmann-Institut für Geologie, Mineralogie und Paläontologie, Friedrich-Wilhelms-Universität, Bonn, Germany, GPIT – Paläontologisches Institut und Museum für Geologie und Paläontologie, Universität Tübingen, Germany; HLMD – Hessisches Landesmuseum, Darmstadt, Germany; IVPP – Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing, China; IZANK – Institute of Zoology, Academy of Sciences, Kiev, Ukraine; MCZ – Museum of Comparative Zoology, Harvard University, Cambridge, Massachusetts, USA; NHMM – Naturhistorisches Museum, Mainz, Germany; NHMUK – Natural History Museum, London, England; NMP – National Museum, Prague, Czech Republic; PIN – Paleontological Institute, Russian Academy of Sciences, Moscow, Russia; SMM – Science Museum of Minnesota, St. Paul, Minnesota, USA; SMNK – Staatliches Museum für Naturkunde, Karlsruhe, Germany; TMP – Royal Tyrrell Museum of Palaeontology, Drumheller, Alberta, Canada; UALVP – University of Alberta Laboratory for Vertebrate Paleontology, Edmonton, Alberta, Canada; YPM-PU – Yale Peabody Museum, (former Princeton University collection), Yale University, New Haven, Connecticut, USA.
Other abbreviations
Ma – millions of year ago; MN – Mammal Neogene (Neogene portion of European Land Mammal Ages); MP – Mammal Paleogene (Paleogene portion of European Land Mammal Ages); NALMA – North American Land Mammal Age.
Systematic palaeontology
Pelobatoidea Bonaparte, 1850
Diagnosis (modified from Duellman and Trueb 1994, their “Pelobatidae”; Roček 2013): Eight procoelous presacral vertebrae with imbricate neural arches; atlantal cotyles closely juxtaposed; transverse processes of sacral vertebra dilated; first postsacral vertebra (V10) bears rod-like or dilated transverse processes similar to those of the transverse processes of sacral vertebra, which fuse to transverse processes of sacral vertebra by means of bony webbing in fully grown adults (not known in Megophryidae and Pelodytidae); pectoral girdle arciferal; omosternum cartilaginous, sternum ossified; anterior end of scapula not overlain by clavicle; two tarsalia.
Contained taxa: Pelobatidae (Eopelobates and Pelobates); Scaphiopodidae (Elkobatrachus, Macropelobates, Scaphiopus, Spea, and Tephrodytes); Megophryidae (Borneophrys, Brachytarsophrys, Leptobrachella, Leptobrachium, Leptolalax, Megophrys, Ophryophryne, Oreolalax, Scutiger, and Xenophrys); and Pelodytidae (Aerugoamnis, Miopelodytes, and Pelodytes) (e.g. Henrici et al. 2013; Roček 2013; Frost 2014).
Pelobatidae Bonaparte, 1850
Diagnosis (modified from Rage and Hossini 2000; Roček 2013): Dermal roofing bones always covered with sculpture, which is an integral part of the bone, not a secondary exostosis like, e.g. in Latonia or tropical hylids; frontoparietal azygous (at least in posterior part), with convex posterior margin in metamorphosed individuals, tripartite in larvae and younger metamorphosed individuals (consisting of a pair of frontoparietals plus posterior median element inserted posteriorly between them: Roček 1981, figs. 42, 43, 44, 50; Maus and Wuttke 2004, figs. 3, 4); quadratojugal present; parasphenoid with median keel and posterior median convexity; processus palatinus of maxilla elongate (=palatine fused to maxilla: Roček 1981, fig. 59a, b; Lebedkina 2004, fig. 95a); postchoanal ramus of vomer absent; pterygoid with moderately developed ventral flange on its lateral margin; sternum with elongated, posteriorly tapered ossified stylus; caput humeri shifted laterally, fossa cubitalis open laterally; ilium without dorsal tubercle, iliac shaft without dorsal crest.
Contained genera: Pelobates and Eopelobates.
Stratigraphy and distribution Footnote 1: Late Cretaceous – Type area of Judith River Formation (Campanian), Montana, USA (Sahni 1972); Ellisdale (Campanian), New Jersey, USA (Denton and O’Neill 1998); intertrappean beds (Maastrichtian) of Asifabad and Takli (former Gitti Khadan), near Nagpur, India (Sahni et al. 1982; Gayet et al. 1984; Prasad and Sahni 1987, 2009). Early Eocene – MP 7, Silveirinha, Portugal (Rage and Augé 2003); MP 7, Le Quesnoy, France (Nel et al. 1999). Late Eocene – MP 18, St Neboule, Priabonian, France (Rage and Vergnaud-Grazzini 1978; Rage 1988). Early Oligocene – MP 21, Hoeleden and Hoogbutsel, Belgium (Hecht and Hoffstetter 1962). Middle Miocene – Gürcü Valley, Turkey (Wassersug and Wake 1995); Sarmatian, Karpov Yar, Moldova (Skutschas and Bannikov 2009). Late Miocene – MN 9, Rudabánya, Hungary (Roček 2005). Recent – Europe (east to the Ural Mts. and Ural River), northern Africa, and Middle East.
Remarks: Here we view Pelobatidae in the taxonomical sense of Frost et al. (2006) and include in it only the type genus Pelobates Wagler, 1830 and Eopelobates Parker, 1929 (e.g. Roček 1981, 2013). Several other genera that have been included in Pelobatidae are briefly discussed below.
Uldzinia kurochkini Gubin, 1996 is from the early Oligocene of Mongolia. It is known only by the left holotype maxilla PIN 3109/236 from a medium-sized frog (length of maxilla is 11.8 mm). This taxon was diagnosed by irregular, pitted sculpture on the posterior part of the outer surface of the holotype maxilla, a pointed processus zygomaticomaxillaris having a vertical posterior margin, and a moderately prominent processus posterior (Gubin 1996). However, recent examination of the specimen by one of us (Z.R.) reveals that the entire outer surface of the bone lacks sculpture. Instead, there is only an irregular, groove-like depression running from below the tip of the processus zygomaticomaxillaris and more or less parallel with the margo orbitalis towards the mid-depth of the bone at the level of the processus frontalis, where it disappears. Additionally, there is a triangular depression on the outer surface of the bone between the processus zygomaticomaxillaris and the processus posterior. On the inner surface, the most remarkable features are the absence of both the processus palatinus and the processus pterygoideus. Because the pointed processus zygomaticomaxillaris indicates ligamentous, rather than osseous contact with the lamella alaris of the squamosal, and because contacts with the quadratojugal (if any) and pterygoid also were different from those in Pelobates and Eopelobates, we exclude Uldzinia from Pelobatidae. Earlier, Sanchiz (1998) regarded Uldzinia as incertae sedis within Pelobatoidea.
Macropelobates osborni Noble, 1924 from the early Oligocene of Mongolia and M. linquensis (Yang, 1977) from the middle Miocene (equivalent to MN 4 or MN 5 of the European mammalian chronostratigraphy) of eastern China differ from Pelobates and Eopelobates mainly in having paired frontoparietals (Roček et al. 2011). Comparisons within Pelobatoidea have suggested to some authors that Macropelobates is more similar to Scaphiopus and Spea, rather than to Pelobates and Eopelobates (Noble 1924; Roček 1982). However, other authors have argued for a closer relationship among Macropelobates, Pelobates, and Eopelobates (e.g. Henrici and Haynes 2006; Henrici et al. 2013).
Elkobatrachus brocki Henrici et Haynes, 2006 from the middle Eocene Elko Formation, Nevada, USA, was assigned to Pelobatidae (in a broader sense, corresponding to the original concept by Bonaparte 1850, and to the superfamily Pelobatoidea of Frost et al. 2006). Among other characters, it was diagnosed by the absence of sculpture on dermal skull bones, by having paired frontoparietals without a tectum supraorbitale (=supraorbital flange) and with occipital canal exiting onto dorsal surface of frontoparietal, and by having the urostyle longer than the remainder of the vertebral column. It is obvious that these characters do not fit into the scope of the Pelobatidae as defined in the present paper, and would rather point to Scaphiopodidae. However, several features (urostyle exceeding the length of the vertebral column and provided with two, broad-based, laterally oriented transverse processes; different style of pectoral girdle) that differ greatly from both Pelobatidae and Scaphiopodidae might indicate that Elkobatrachus does not fit into any of these families, at least as they are diagnosed on extant taxa. It should also be noted that, because the carpal and distal tarsal bones are not ossified and were presumably cartilaginous at the time of death, and because the notochord was probably still continuous, Elkobatrachus specimens do not represent fully grown adults (Henrici and Haynes 2006).
Pelobates Wagler, 1830
1830 Pelobates Wagler, Naturl. Syst. Amph., p. 206. – Species typica (by subsequent designation): Bufo fuscus Laurenti, 1768
1832 Cultripes Müller, Tiedem. Zeitschr. Phys. 4: 212, Isis (Oken) 1832: 538. – Species typica: Cultripes minor Müller, 1832 = Pelobates fuscus Wagler, 1830
1866 Didocus Cope, J. Acad. Nat. Sci. Philad. (2)6: 81. – Species typica: Didocus calcaratus (Michahelles, 1830) = Pelobates cultripes (Cuvier, 1829)
1866 Zaphrissa – Cope, J. Acad. Nat. Sci. Philad. (2)6: 77 – Species typica (by monotypy): Zaphryssa eurypelis Cope, 1866 = Pelobates decheni Troschel, 1861
1958 Pseudopelobates Pasteur, C.R. Acad. Sci. Paris 247: 1037. – Species typica (by monotypy): Pelobates transcaucasicus Delwig, 1928 = Pelobates syriacus boettgeri Mertens, 1923
Diagnosis (modified from Rage and Hossini 2000): Sculpture on dermal roofing bones of pit-and-ridge type (in Oligocene and some Miocene forms), pustular (in Pliocene through Recent forms), and transitional between those patterns (in some Miocene forms); anterolateral margin of nasals concave in its anteromedial section, and convex or straight in its posterolateral section; sphenethmoid in fully grown adults completely covered by posterior part of nasals and anterior end of frontoparietal; frontoparietal in contact with squamosal in fully grown adults (separated only in Pelobates fuscus); urostyle short (its posterior end not reaching acetabular part of pelvis); tibiale and fibulare fused at their ends in fully grown adults; spade present in Recent forms (but absent in P. decheni and possibly also in other Oligocene forms for which complete hindlimbs are known); TF equal or shorter than F; F + TF shorter than SVL.
Stratigraphy and distribution Footnote 2: Middle Oligocene – Pelobates sp., Mas-de-Got (MP 22), Itardies (MP 23), Pech Grabit (MP 23), all Quercy, France (de Bonis et al. 1973). Late Oligocene – P. decheni, Rott, MP 30, near Bonn, Germany (Böhme et al. 1982); Enspel, MP 28, Germany (Roček and Wuttke 2010); Pelobates sp., Oberleichtersbach, MP 30, Germany (Böhme 2008); Pelobates sp., Coderet, MP 30, France (Crochet 1972). Early Miocene – P. fahlbuschi, Sandelzhausen, 16.47 or 16.27 Ma, Germany (Böhme 2010); P. sanchizi, Mokrá-Western Quarry, MN 4, Czech Republic (Ivanov 2008). Middle Miocene – P. sanchizi, Mátraszőlős 1, Mátraszőlős 2, and Sámsonháza 3, all MN 6, Hungary (Venczel 2004) and Gratkorn, 12.2–12.0 Ma, Austria (Böhme and Vasilyan 2014); P. cultripes, France (Bailon et al. 1988); Pelobates sp., Sansan, MN 6, France (Rage and Hossini 2000). Early or middle Miocene – Pelobates sp., Bes-Konak, Turkey (Paicheler et al. 1978). Middle–late Miocene – P. cf. sanchizi, Felsötárkány-Felnémet, MN 7 + 8–MN 9, Hungary (Venczel and Hír 2013). Late Miocene – Pelobates, Los Aljezares, Teruel, MN 12, Spain (Sanchíz 1977); Pelobates sp., Gritsev, Sarmatian, Ukraine (Zerova 1985); Pelobates fuscus, Novaya Emetovka, Ukraine (Chkhikvadze 1984). Pliocene – P. fuscus, France (Bailon et al. 1988); Pelobates sp., Arondelli, Italy (Vergnaud-Grazzini 1970); P. fuscus, Pelobates cf. P. fuscus, and Pelobates sp., Węże 1 and 2, both MN 15, Rębielice Królewskie 1, MN 16, Poland (Młynarski 1961, 1962, 1977; Roček 1981; Młynarski et al. 1984; Sanchiz 1998); P. syriacus, Rębielice Królewskie 1, MN 16, Poland (Młynarski 1977; Sanchíz and Mlynarski 1979; Młynarski and Szyndlar 1989); Pelobates cf. fuscus, Ivanovce Csarnotan, Slovakia (Hodrová 1981); Pelobates sp. Çalta, Turkey (Rage and Sen 1976); P. praefuscus, Etulia, Moldavia (Khosatzky 1985). Middle Pleistocene – P. cultripes, Spain (Sanchíz 1983). Late Pleistocene – Pelobates cf. syriacus, Pili 2, Kos Island, Greece (Sanchíz 1984); P. fuscus and Pelobates sp., France (Bailon et al. 1988); Pelobates sp., Šandalja near Pula, Croatia (Paunović 1984); Pelobates sp. and P. fuscus, Zmeevka and Rudnyi, both Belgorodsky Region, Russia (Ratnikov 1988); Pelobates sp., Hortus cave, Hérault France (Rage 1972). Recent – Europe, northern Africa, and the Middle East.
Remarks: Sculpture on dermal roofing bones pustular (Pelobates cultripes, P. fuscus, P. praefuscus, P. syriacus, and P. varaldii: Roček 1981; Khosatzky 1985) or pit-and-ridge (P. decheni: Böhme et al. 1982; Roček and Wuttke 2010; P. fahlbuschi: Böhme 2010; P. sanchizi: Venczel 2004; Pelobates sp. from Oberleichtersbach: Böhme 2008; Pelobates sp. from Sansan: Rage and Hossini 2000). Because sculpture varies within the genus, it can be used only in association with other diagnostic characters. Similarly, the sacro-urostylar articulation is movable in P. decheni, whereas both elements are coalesced in Recent forms; the tibiale and fibulare are free from one another in P. decheni, whereas those bones are fused in extant forms; and a spade is reliably known only in Recent forms.
Eopelobates Parker, 1929
1929 Eopelobates Parker, Ann. Mag. Nat. Hist. 4: 277. – Species typica (by monotypy): Eopelobates anthracinus Parker, 1929
1938 Propelodytes Weitzel, Notizbl. Hessischen Geol. Landesanst. 19: 43. – Species typica (by monotypy): Propelodytes wagneri Weitzel, 1938
1941 Halleobatrachus Kuhn, Nova Acta Leopold. 10: 353. – Species typica (by monotypy): Halleobatrachus hinschei Kuhn, 1941
1941 Eobuffela Kuhn, Nova Acta Leopold. 10: 356. – Species typica (by monotypy): Eobuffela parvula Kuhn, 1941
1941 Parabufella Kuhn, Nova Acta Leopold. 10: 358. – Species typica (by monotypy): Parabufella longipes Kuhn, 1941
1941 Palaeopelobates Kuhn, Nova Acta Leopold. 10: 360. – Species typica (by monotypy): Palaeopelobates geiseltalensis Kuhn, 1941
1941 Archeopelobates Kuhn, Nova Acta Leopold. 10: 361. – Species typica (by monotypy): Archaeopelobates efremovi Kuhn, 1941
1941 Amphignathodontoides Kuhn, Nova Acta Leopold. 10: 364. – Species typica (by monotypy): Amphignathodontoides eocenicus Kuhn, 1941
1941 Germanobatrachus Kuhn, Nova Acta Leopold. 10: 368. – Species typica (by monotypy): Germanobatrachus beurleni Kuhn, 1941
Diagnosis (modified from Roček 2013): Sculpture on dermal roofing bones exclusively of pit-and-ridge type; anterolateral margin of nasals straight along its entire length; sphenethmoid in fully grown adults exposed in a rhomboid gap between posterior part of nasals and anterior margin of frontoparietal; frontoparietal not in contact with squamosal; urostyle long, may reach level of acetabular portion of pelvis; tibiale and fibulare not coalesced (fused only in adults of Eopelobates bayeri and in E. grandis); spade absent; TF equal or longer than F; F + TF exceeding SVL in fully grown adults, but shorter than SVL in juveniles.
Stratigraphy and distribution: Late Cretaceous (assorted published identifications, all variations of either definitely assigned to, questionably assigned to, or compared to Eopelobates) – Fruitland Formation (Campanian), New Mexico, USA (Armstrong-Ziegler 1980); Mesaverde Formation (Campanian), Wyoming, USA (e.g. DeMar and Breithaupt 2008); Lance Formation (Maastrichtian), Wyoming, USA (e.g. Estes 1970; Estes and Sanchíz 1982; Gardner 2008); Bug Creek Anthills locality (Maastrichtian or early Paleocene), Hell Creek Formation, Montana, USA (Estes and Sanchíz 1982; Gardner 2008). Early Eocene – Eopelobates aff. E. hinschei, Prémontré, MP 10, France (Duffaud 2000); Eopelobates sp., Vastan Lignite Mine, Ypresian, Gujarat, India (Folie et al. 2013). Middle Eocene – Eopelobates hinschei, Geiseltal, lower Geiseltalium, near Halle, Germany (Estes 1970); Eopelobates wagneri, Grube Messel, lower Geiseltalium, near Darmstadt, Germany (Wuttke 2012b). New species of Eopelobates, Green River Formation, Wyoming, USA (this study). Late Eocene – Eopelobates grandis, Chadron Formation, South Dakota, USA (Zweifel 1956; Estes 1970; Henrici 2002); cf. Eopelobates sp., Mission Valley Formation, California USA (Golz and Lillegraven 1977); Eopelobates cf. E. hinschei, Hordle Cliff, England (Milner et al. 1982); cf. Eopelobates, Headon Hill, Isle of Wight, England (Rage and Ford 1980); cf. Eopelobates, Quercy, France (Crochet et al. 1981). Early Oligocene – E. bayeri, Hoogbutsel, Hoeleden, and Boutersem TGV sites, all MP 21, Belgium (Smith 2003); Eopelobates sp., Sieblos, Germany (Gaudant 1985); Eopelobates sp., Zaisan Basin, Kazakhstan (Chkhikvadze 1985). Late Oligocene – E. anthracinus, Rott, MP 30, near Bonn, Germany (Parker 1929); E. bayeri – Bechlejovice, Czech Republic (Špinar 1952, 1972; Bellon et al. 1998); Eopelobates sp., Oberleichtersbach, MP 30, Germany (Böhme 2008). Early Miocene – Eopelobates sp., Dolnice, MN 4, Czech Republic (Hodrová 1987); Sandelzhausen, MN 5, Germany (Böhme 2010). Middle Miocene – E. bayeri, Devínska Nová Ves, Slovakia (Hodrová 1988). Pliocene – Eopelobates sp., Osztramos 1, MN 14, Hungary (Venczel 2001); Węże 1, MN 15, and Rębielice Królewskie 1, MN 16, both Poland (Młynarski 1961, 1962; Sanchíz and Mlynarski 1979); ?Eopelobates cf. bayeri, Ivanovce Csarnotan, Slovakia (Hodrová 1981); E. cf. bayeri, Gorishnaya Vygnanka, Ukraine (K.A. Tatarinov in Chkhikvadze 1981, 1984).
Remarks: The original diagnosis of the genus Eopelobates given by Parker (1929, pp. 277–278) was as follows: “Maxillary teeth present; bony incrustations on the frontoparietal, squamosal, and maxilla; last two bones broadly in contact behind the orbit. Eight pre-sacral vertebrae, the anterior, at least, with short transverse processes; sacral diapophyses [=transverse processes in our terminology] very widely dilated; coccyx without transverse processes, not fused with the sacrum; ilia not thickened at their symphysis. Scapula twice as long as broad. Tibiale and fibulare free; no pre-hallux or pre-pollex; tibia as long as the femur, the two together much shorter than the head and body. Metacarpals less than half the length of the radio-ulna. Terminal phalanges simple.” In his description of the type and then only known species, Parker (1929) specified that the incrustation (=sculpture in our terminology) is pitted and that the processus zygomaticus of the squamosal forms a broad suture with the maxilla. He was able to differentiate the holotype of Eopelobates anthracinus from Pelobates only by the shorter proximal phalanges and lack of prehallux in the former.
In 1970, Estes revised the generic diagnosis for Eopelobates and mentioned the following additional features (our comments are in parentheses): prominent, elongated sternal style (this occurs in both Pelobates and Eopelobates, so this character is included in our diagnosis of Pelobatidae); approximately subequal orbit and temporal openings (difficult to assess in most specimens); dermal ossification well developed and fused to skull roof (sculpture in both Pelobates and Eopelobates is not a secondary exostosis, but it is an integral part of the bone); skull roof flat or concave dorsally (this is a preservational artifact resulting from compression of skulls during fossilization); ethmoid wide and blunt anteriorly, and with dorsal ethmoid roof over nasal capsules (this depends on degree of ossification of the nasal septum, postnasal walls, and nasal tecta; thus, this should be considered an age-dependent character); prominent, well-ossified processus paraoccipitalis on frontoparietal and occiput (also present in Pelobates); complete maxillary arcade (the quadratojugal is present both in Eopelobates and Pelobates, so this also is a diagnostic character of Pelobatidae).
The taxonomic composition and geographic and temporal distributions of Eopelobates have varied since Parker (1929) first formally named the genus. At its most inclusive, Eopelobates contained about 10 species ranging from the Late Cretaceous to Pliocene of Europe, Asia, and North America (e.g. Duellman and Trueb 1994). At the other end of the spectrum, Sanchiz (1998) restricted the genus to three named European species of Eocene–Miocene age. Of the species previously included within the genus and not synonymised with other congeners, two Asian Late Cretaceous species (E. leptocolaptus Borsuk-Białynicka, 1978 and E. sosedkoi Nessov, 1981) were transferred to the discoglossoid genus Gobiates and one North American Eocene species (E. guthriei Estes, 1970) was transferred to Scaphiopus (see Špinar and Tatarinov 1986; Roček and Nessov 1993; Henrici 2000; Roček 2013). Accounts for the six congeners recognised here are presented below in alphabetical order. All six species are known by slab-style skeletons of varying numbers (one to several dozens) and quality, and four of the species are known from just one locality or area.
Development: Stages were determined after Nieuwkoop and Faber (1967) who provided information about formation of the skeleton, whereas the normal table proposed by Gosner (1960) is mainly restricted to external features. Trueb and Hanken (1992) provided a table comparing equivalent stages of both tables. To avoid potential confusion with Pelobates, which might be expected to have tadpoles similar to those of Eopelobates, development was investigated using tadpoles (see Fig. 1) largely from a single locality and horizon (Bechlejovice, late Oligocene) from which Pelobates is absent.

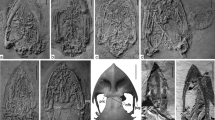
Photographs depicting developmental series of Eopelobates. a The earliest recorded stage, corresponding to about NF stage 51 (NMP Pb 1579), in ventral aspect. b NF stage 51–52 (NMP Pb 439), in ventral aspect. c NF stage 51–52 (NMP Pb 1403), in ventral aspect. d NF stage 51–52 (NMP Pb 1401), in ventral aspect. e NF stage 52–54 (NHMUK PV OR49464), in dorsal aspect; arrow points to posterior median element. f NF stage 52–54 (NMP Pb 1716), in dorsal aspect; arrow points to posterior median element. g Estimated NF stage 55 (NMP Pb 434), in ventral aspect. h NF stage 55, early metamorphosis (NMP Pb 1461), in ventral aspect; arrow points to tibia and fibula. i NF stage 55–56 (NMP Pb 1463), in ventral aspect; upper arrow points to posterior median element and lower arrow points to femur. j NF stage 56, early metamorphosis (NMP Pb 445), in ventral aspect; arrow points to pelvic girdle with hindlimbs. k NF stage 62, advanced metamorphosis (NMP Pb 450), in ventral aspect. l NF stage 52–54, detail of early stage in development of frontoparietal complex in premetamorphic tadpole (NMP Pb 407), in dorsal aspect, showing paired frontoparietals well separated from the small posteromedian element (latter marked by arrow). m Estimated NF stage 55, frontoparietal complex in advanced premetamorphic tadpole (NMP Pb 440), in dorsal aspect, showing paired frontoparietals approaching one another and being contacted along their posteromedian edges by a moderately wide posterior median element (latter marked by arrow). n Estimated NF stage 62, detail of frontoparietal complex in advanced metamorphic tadpole (NMP Pb 430), in dorsal aspect, showing frontoparietals now fused posteriorly and broadly sutured posteriorly to a posterior median element (latter marked by arrow) that has now expanded laterally to form the entire posterior portion of the frontoparietal complex. o NF stage 62, advanced metamorphosis (NMP Pb 1695), in right dorsolateral aspect; arrow points to posterior median element. p NF stage 63, advanced metamorphosis (NMP Pb 449), in ventral aspect, showing well-developed fore limbs. Species identifications: a–d, f–p are Eopelobates bayeri from the late Oligocene of Bechlejovice, Czech Republic; e is E. anthracinus from the late Oligocene of Rott, Germany. Specimens at different magnifications; scale bars 5 mm
The earliest developmental stage roughly corresponds to NF stage 51 (Fig. 1a), in which both frontoparietals are weakly ossified and the posterior median element is not yet formed. The vertebral column consists of eight vertebrae, of which the two most posterior are poorly ossified and the anterior most three are still without transverse processes; presumably, their left and right neural arches are not fused. In NF stages 51–52, the parasphenoid appears, the posteromedial walls of the otic capsules begin to ossify, and the transverse processes of the anterior three vertebrae become apparent; the number of vertebrae is still eight or nine (Fig. 1b, c). In subsequent stages, ossification of the frontoparietal complex (still consisting of only the paired frontoparietals) and the parasphenoid proceeds, together with the otic capsules whose outer wall ossifies; the number of vertebrae increases to 10 or 11 (Fig. 1d). In NF stages 52–54, the posterior median element appears above the tectum synoticum (Fig. 1e, f). As is apparent from Fig. 1f, the posterior median element appears in premetamorphic tadpoles with eight or nine vertebrae and which still have the typical larval mouth. It is also obvious from comparisons of Fig. 1g with Fig. 1h (both tadpoles are at about NF stage 55) that, shortly before or simultaneous with the first signs of ossification of the hindlimbs, the larval mouth apparatus is still positioned close to the anterior end of the head, which indicates that cranial metamorphosis has not yet begun. By this stage, both frontoparietals are well ossified and approach one another, and the otic capsules are now completely ossified. As can be seen in Fig. 1h, the earliest parts of the hindlimbs to ossify are diaphyses of the tibia and fibula, which appear when the vertebral column consists of nine vertebrae. The femur appears slightly later, when the vertebral column consists of 10 ossified vertebrae (Fig. 1i). In this stage, which may be defined as advanced NF stage 55, the posterior median element now contacts the posteromedial margin of the frontoparietals. Shortly afterwards, at about NF stage 56, the iliac shaft begins to ossify, the tibia and fibula fuse with each other, and hindlimb ossification extends distally to the tibiale and fibulare (Fig. 1j; see also Špinar 1972, pl. 176). The whole pelvic girdle and hindlimbs remain well separated from the vertebral column. A preceding stage seen in extant Pelobates fuscus, in which both halves of the pelvic girdle are widely separated from each other (Ročková and Roček 2005, fig. 5D, J, N), is not documented by any of our Eopelobates tadpoles, but it is suggested by Špinar’s (1972) specimen illustrated in his plate 177 (current catalogue numbers for that part and counterpart are NMP Pb 443 and Pb 444). In this stage (NF 56), ossification of the vertebral column extends into the postsacral area, where paired rudiments of the 11th vertebra may be discerned. Also, no cranial bones other than the otic capsules, parasphenoid, and those belonging to the frontoparietal complex can be recognised. It can be inferred from the size of the NF stage 56 individual illustrated in Fig. 1j that the total body length, including the tail, of early metamorphic tadpoles could reach about 100 mm. In late metamorphic tadpoles (NF stages 62–63), in which the hindlimbs are nearly completely ossified and the fore limbs are partly ossified (Fig. 1k, o, p), total body length is shorter (in the individual illustrated in Fig. 1k, it is estimated at about 70 mm). Recently metamorphosed invididuals (NF stage 66; not figured) are even smaller, with SVLs of only about 20–30 mm. In late metamorphic tadpoles, the frontoparietal complex begins to fuse between the parietal portions of the frontoparietals, leaving a wedge-like fontanelle between the frontal portions (Fig. 1n). It is also obvious that the posterior median element increases in width during development, so whereas it is a relatively small in premetamorphic tadpoles (Fig. 1e, f, l), it constitutes the whole posterior part of the frontoparietal complex in late metamorphic individuals (Fig. 1n; see also Špinar 1972, pl. 171–1).
Eopelobates anthracinus Parker, 1929 Fig. 2

Photographs of Eopelobates anthracinus Parker, 1929, from the latest Oligocene of Rott, Germany. a Holotype skeleton (NHMUK R4841), natural mold in dorsal aspect; scale bar 10 mm. b Counterpart of the same skeleton (GPIB-Ro 4029), natural mold in ventral aspect; same magnification as (a). c Detail of skull in holotype (NHMUK R4841), in dorsal view; upper arrow points to left premaxilla and lower arrow points to posterior margin of right nasal; magnification about 2.9× larger than (a). d Detail of pelvic girdle and vertebral column in holotype (NHMUK R4841), in dorsal aspect; upper arrow points to sacro-urostylar joint and lower arrow points to deep groove (represented by a prominent ledge on the imprint) on right transverse process of sacral vertebra; magnification about 2.9× larger than (a). Note that direction of lighting used in these photographs creates the illusion that the bones are preserved in relief; see Estes (1970, fig. 1) and Špinar (1972, pl. 165) for photographs of the holotype that accurately depict the bones preserved as impressions
Diagnosis (modified from Estes 1970; Špinar and Roček 1984; Roček 2013): Cranial sculpture restricted to posterolateral margin of frontoparietal complex, posterolateral part of nasal, posterior part of lamella alaris of squamosal, and posterior half of maxilla; pars facialis of premaxilla almost at midlength of the horizontal part of the bone, pars facialis terminates in a rounded point (Fig. 2c) and its medial margin widely convex; nasals widely separated from each other; lamella alaris of squamosal gradually tapers anteriorly in a point, its margo orbitalis is shallowly concave, contact margin with the processus zygomaticomaxillaris of maxilla is not well defined; urostyle not coalesced with sacral vertebra (Fig. 2b, d); tibiale and fibulare not fused with one another; carpus and distal tarsus not ossified.
Material: Two skeletons of metamorphosed individuals are available from Rott, Germany. Both are natural molds that preserve impressions of bones, along with small amounts of permineralized bone. The holotype (NHMUK R 4841) is a nearly complete skeleton that is exposed in dorsal aspect and was first described by Parker (1929; see also Estes 1970; Špinar 1972). The counterpart (GPIB-Ro 4029) depicts the ventral aspect of the same skeleton; it remained undocumented for over a half-century, until it was recognised and described by Špinar and Roček (1984). The second and less complete skeleton (GPIT 1733) is exposed in ventral aspect and was described by Roček (1995). A tadpole (NHMUK PV OR49464) is also known from Rott.
Stratigraphy and distribution: Latest Oligocene, Rott, near Hennef, Germany (Koenigswald et al. 1992).
Description: Our description is based on the three known metamorphosed specimens. (The tadpole NHMUK PV OR49464 was figured and briefly mentioned above in the “Development” section of the Eopelobates account.) The holotype (NHMUK R 4841: Fig. 2a, c, d) and its counterpart (GPIB-Ro 4029: Fig. 2b) are imprints of the same skeleton exposed in, respectively, dorsal and ventral views; these are from a relatively small individual, having a SVL of about 32 mm according to Parker (1929). GPIT 1733 (Roček 1995, figs 1, 2) is exposed in ventral aspect; although less complete, it is from a larger (SVL = 40 mm: Roček 1995) and, evidently, more mature individual.
The pars facialis of the premaxilla is narrow, terminates dorsally in a rounded point (Fig. 2c), and is deeply concave along its inner surface. Its base joins the dorsal surface of the horizontal portion of the bone (=pars dentalis) at approximately the midpoint of the latter’s width, but slightly closer towards the medial end. Lateral to the base of the pars facialis, about eight tooth positions are preserved. Based on the ventral counterpart, the total number of premaxillary tooth positions is estimated at 12. The maxilla is low below its margo orbitalis, but the anterior end of the bone is comparatively deep and its anterior margin is slightly concave. Posteriorly, the maxilla terminates in a short processus posterior. The lateral (=external) surface of the posterior portion of the maxilla, back from about the anteroposterior midpoint of the margo orbitalis, bears weak sculpture. There are faint horizontal striae closer to the ventral margin of the bone, whereas dorsally and closer to the margo orbitalis the sculpture is pitted. Both the processus frontalis and the processus zygomaticomaxillaris are developed, but neither is especially prominent. The quadratojugal is rather deep, mediolaterally compressed, and bears horizontal striations on its lateral surface similar to those on the processus posterior of the maxilla. Both the ventral and dorsal margins of the quadratojugal are smooth and rounded.
The posterolateral part of the nasal is covered by a pit-and-ridge sculpture that is less pronounced than on the maxilla. The medial margin of the nasal is slightly concave and its processus anterior is rounded. In life the paired nasals were probably widely separated from each other. The processus paraorbitalis is probably narrow and pointed, but the complete shape of the bone cannot be reconstructed from the dorsal imprint. However, the ventral imprint suggests that the anterolateral margin of the nasal is only slightly concave. An imprint of the squamosal on the right side is well preserved on the holotype (Fig. 2c). The posterior part of the lamella alaris is deep, with its dorsal margin moderately extending as a broad processus dorsalis. The anterior part of the lamella alaris, anterior to the level of the processus posterolateralis, tapers anteriorly to a thin, but rounded point; its margo orbitalis is slightly concave and the ventral margin of its processus zygomaticus is correspondingly convex. The lamella alaris is separated from the outer surface of the processus posterolateralis by a distinct, though not especially prominent crista. Externally, the upper part of the posterior portion of the lamella alaris is covered by pitted sculpture, whereas the lower part is smooth.
The frontoparietal complex is azygous, with no trace of sutures. Nevertheless, along the midline between both frontoparietals there is a distinct, anteroposteriorly oriented depression (Fig. 2c). Although sutures between the posterior median element and the frontoparietals have disappeared during ontogeny, the former element can still be recognised by the prominent convexity along the posterior margin of the frontoparietal complex. The dorsal surface of the frontoparietal complex is covered by a few shallow, but large pits located along the posterolateral margin of the bone, whereas there are only several anteroposteriorly oriented striae covering the anterior part of the bone. The lateral margin of the dorsal, sculptured surface, at the level of the anterior wall of the otic capsule, extends laterally as a widely rounded processus lateralis superior. Between the nasals and frontoparietals is a large rhomboid gap filled with an imprint of a pitted sphenethmoid. Although only the imprint of the medial part of the nasal on the left side is preserved, it is clear that the sphenethmoid is terminated anteriorly by a sharp transverse border between both nasals.
The parasphenoid is well preserved on the ventral counterpart. It has a distinct keel along the anterior part of the processus medialis (=cultriform process). The shape and proportions of the laterally directed processus lateralis (=lateral ala) were illustrated by Špinar and Roček (1984, fig. 3B).
The vertebral column consists of eight presacral vertebrae. The articular facets for the occipital condyles of V1 are separated by a narrow and nearly pointed median process. The transverse processes of V2–V4 are perpendicular and robust, but their exact shapes cannot be restored. The transverse processes of the posterior three presacrals are thin and inclined anteriorly, but their precise extents also cannot be restored. The transverse processes of the sacral vertebra are dilated anteroposteriorly and bear a distinct ledge (represented by a groove on the imprint in the sediment; marked by arrow in Fig. 2d) parallel with the posteromedial margin on the dorsal surface of both transverse processes. The urostyle in the holotype is clearly separated from the sacral centrum and has no transverse processes. By contrast, the urostyle in the more mature GPIT 1733 is fused with the sacral vertebral centrum. The proximal part of the urostyle of both specimens is broad, but because the rest of the bone is fragmentary in the holotype and hidden behind the ilium in GPIT 1733, the complete length of the urostyle cannot be restored in either specimen. Nevertheless, the urostyle appears to have been comparatively short, based on the observation that even though the tips of the iliac shafts are level with the anterior ends of the sacral transverse processes, the posterior tip of the urostyle does not reach the iliac symphysis.
The pectoral girdle is arciferous and the medial end of the coracoid is widely dilated. An imprint of the left scapula is preserved on the right side of the dorsal aspect of the skeleton (Fig. 2a); its posterior margin is concave, whereas its anterior margin is nearly straight. The right scapula is preserved as an imprint on the right side of the ventral counterpart (Fig. 2b) next to the sacral vertebra, where it is accompanied by an imprint of the clavicle. An imprint of the suprascapula also is preserved on the left side; its dorsal margin is concave, with the anterior lobe less prominent than the posterior one. The cleithrum is represented by a deep imprint on the anterior margin of the suprascapula, but provides no significant information. Imprints of the fore limbs are fragmentary, but it seems that the distal parts of the radioulna are unfused. An ossified carpus is absent. The phalangeal formula (inferred from both dorsal and ventral parts) for the manus is 2-?-3-3. The femur is straight and bears a distinct crista femoris on its proximal section. The tibiale and fibulare are free from each other. An ossified distal tarsus is absent. The phalangeal formula for the pes in GPIB-Ro 4029 is estimated to be 2-2-3-4-3 (2-2-3-?-3 in GPIT 1733).
Remarks: Eopelobates anthracinus is the type species of Eopelobates.
Eopelobates bayeri Špinar, 1952
Fig. 3

Photographs of Eopelobates bayeri Špinar, 1952, from the the late Oligocene of Bechlejovice, Czech Republic. a Paratype skeleton (NMP Pb 1694), natural mold in dorsal aspect; same magnification as (b). b Paratype skeleton (NMP Pb 1114; counterpart of NMP Pb 1694), natural mold in ventral aspect; scale bar 10 mm. c Holotype skeleton (NMP Pb 412), natural mold in ventral aspect; scale bar 10 mm. d Detail of skull in paratype (NMP Pb 1694), in dorsal aspect, depicting form and external sculpture of right squamosal and maxilla, both nasals, and frontoparietal complex; magnification about ×3.4 larger than (a). e Detail of pelvic girdle and vertebral column in paratype (NMP Pb 1114), in ventral aspect; arrow points to line of fusion seen in ventral aspect between sacral vertebra and urostyle; magnification about 2.8× larger than (b). f Detail of pelvic girdle and median and posterior portions of vertebral column in paratype (NMP Pb 1694), in dorsal aspect; arrow points to ridge representing posterior margin of sacral neural arch roof; magnification about 2.8× larger than (a)
Diagnosis (modified from Roček 2013): Cranial sculpture well developed on posterior two-thirds of maxilla and on entire nasal, lamella alaris of squamosal, and frontoparietal complex (except along its midline, Fig. 3d, unless this is a preservational artifact); pars facialis of premaxilla close to, but still well separated from the medial end of the horizontal part of that bone and medial margin of that process forms a prominent, rounded outgrowth; nasals in contact with one another along a lengthy median suture with only their short posterior portions divergent and their processus lateralis is broadly rounded; sphenethmoid exposed in a small rhomboid gap between nasals and frontoparietals; processus lateralis inferior of frontoparietal not exceeding the processus lateralis superior (latter may be absent); lamella alaris of squamosal rounded anteriorly, its margo orbitalis straight or almost convex, and its margin for contacting the processus zygomaticomaxillaris of maxilla is well defined and straight (Fig. 3d); urostyle coalesced to sacral vertebra (Fig. 3e); tibiale and fibulare not fused with one another; carpus and distal tarsus calcified or ossified.
Material: Numerous articulated skeletons and tadpoles from Bechlejovice quarry, and isolated bones from various localities (see below). The best preserved and most informative specimens are from the Bechlejovice quarry (late Oligocene), Czech Republic, and include the following: the incomplete holotype skeleton (NMP Pb 412) described by Špinar (1952, 1972; see also Estes 1970); the nearly complete paratype skeleton (NMP Pb 1694, part, and NMP Pb 1114, counterpart) described by Špinar (1972; see also Estes 1970); and a developmental series of several dozen tadpoles (all NMP specimens: see Špinar 1972; this paper, “Specimens used in this study”).
Stratigraphy and distribution: Early Oligocene–middle Miocene of western and central Europe: early Oligocene (MP 21), Hoogbutsel, Hoeleden, and Boutersem TGV sites, all Belgium (Smith 2003); late Oligocene, Bechlejovice, Czech Republic (Bellon et al. 1998; Špinar 1972); early Miocene, Hrabák near Most and Nástup and Merkur mines near Kadaň, all Czech Republic (Špinar 1972); and middle Miocene, Devínska Nová Ves-Bonanza, Slovakia (Hodrová 1988).
Description: Our brief description here is based on two skeletons of metamorphosed individuals from Bechlejovice, Czech Republic, that are preserved as impressions of bone and with some permineralized bone: the holotype (NMP Pb 412: Fig. 3c) and the paratype (NMP Pb 1694, part, and NMP Pb 1114, counterpart: Fig. 3a and b, respectively). Both are from moderate-sized individuals. The holotype has a SVL of 56.5 mm according to Špinar (1952) and the paratype is slightly larger. Špinar (1972, pp. 199–216) provided a more detailed description based on the same two skeletons, plus another five partial skeletons from Bechlejovice.
In the premaxilla, the pars facialis joins the dorsal surface of the horizontal part of the bone closer to the medial than to the lateral end. The medial margin of the pars facialis forms a prominent, widely rounded process, whereas its dorsal end is inclined dorsolaterally (Fig. 3d). The number of observable tooth positions on the premaxilla is 12; there is room for another three on the lateral part of the bone, which suggests the total number of premaxillary tooth positions is about 15. As suggested by the imprint of the right maxilla in NMP Pb 1694, pitted sculpture occurs only on the posterior two-thirds of the bone, whereas the anterior end of the bone is smooth. The estimated number of tooth positions on the maxilla, as suggested by NMP Pb 1114, is 52–55. The quadratojugal is preserved, but because it is not exposed in lateral aspect nothing can be said about its surface texture. The paired nasals contact one another along a long median suture that reaches to the posterior part of those bones. Their processus paraorbitalis tapers to a slender, rounded point. The frontoparietal complex is firmly fused (Fig. 3d) and its lateral margins are widely convex (Fig. 3d); this convexity of the sculptured, dorsal surface represents the processus lateralis superior. The posterolateral corner of the frontoparietal bears a slender and comparatively short processus paraoccipitalis. No processus lateralis inferior can be seen extending on the anterodorsal surface of the otic capsules, unlike in the new species of Eopelobates (see next account). It is unclear if the processus lateralis inferior is truly absent, if it is hidden below the processus lateralis superior, or if it was broken off when the slabs were split; the last scenario might also explain the absence of the prootic/crista parotica. Sculpture on the frontoparietal complex is best developed along the lateral portions. The anteromedian portion of the complex bears an elongate, unsculptured patch that suggests a frontoparietal fontanelle was present at an earlier stage of development. The anterior end of the frontoparietals is indented, suggesting that even in adults there was a rhomboid, exposed part of the sphenethmoid. Although the asymmetrical shape (Fig. 3d) of the sphenethmoid suggests its posterior margin has been partly broken away, it was clearly convex. The sphenethmoid bears prominent anterior and lateral processes, suggesting advanced ossification of the solum nasi and postnasal walls (best seen in NMP Pb 1114, Fig. 3b). The parasphenoid has a distinct keel on the ventral surface of its processus medialis.
The vertebral column consists of eight procoelous presacral vertebrae. V2–V4 are provided with stout, laterally directed transverse processes (Fig. 3f), whereas the four posterior presacrals have thin transverse processes that are inclined anteriorly. The neural arches are moderately imbricate. The transverse processes of the sacral vertebra are strongly dilated anteroposteriorly. The sacral vertebra is separated from the urostyle by the posterior edge of its neural arch (marked by arrow in Fig. 3f), but both are completely fused on the ventral side (Fig. 3e). As can be seen in dorsal aspect, the narrow posteromedial part of the sacral transverse processes is continuous with the urostyle, suggesting it is a derivative of the 10th vertebra. The pectoral girdle is arciferal (Fig. 3e), with both clavicles and coracoids in medial contact, and completed posteriorly by a large ossified sternum. The posterior margin of the scapulae is deeply concave, whereas the anterior margin is almost straight. The carpus is calcified in the holotype (Fig. 3c) or partly ossified; the latter condition being suggested by the presence of one carpal element in the paratype. The phalangeal formula for the manus is 2-2-3-3 (the distal part of fore limb of NMP Pb 1114, formerly 6874a, was illustrated by Špinar 1972, text-fig. 89A, pl. 161). Both ilia are disarticulated and preserved as natural molds of their lateral surfaces in NMP Pb 1114 (Fig. 3e). They were connected by cartilage to the ischia. The tibiale and fibulare are not fused. The distal tarsals are discernible and possibly include a prehallux (Špinar 1972, text-fig. 92). The phalangeal formula for the pes is 2-2-3-4-3.
Remarks: Špinar and Roček (1984) suggested that Eopelobates anthracinus is a juvenile, whereas E. bayeri is an adult of the same species. If so, E. bayeri should be considered a junior synonym of E. anthracinus. That arrangement was advocated by Sanchiz (1998). However, recent re-examination by one of us (Z.R.) of the type material for both species revealed that the available skeletons are all from adults (as evidenced by ossified epiphyses of the long bones) and that the two species can be distinguished reliably by several diagnostic characters (e.g. by shape of the lamella alaris of the squamosal; see our amended Diagnoses for both species).
Eopelobates bayeri has the broadest geographic and temporal range of any named species of Eopelobates, being known from the early Oligocene of Belgium, the late Oligocene–early Miocene of the Czech Republic, and the middle Miocene of Slovakia. By contrast, the other congeners are known from more restricted areas (in some cases just the holotype locality) and temporal intervals. Eopelobates bayeri also is the only species of Eopelobates for which a developmental series of tadpoles is available.
Eopelobates deani sp. nov.Figs. 4, 5


Photographs of Eopelobates deani sp. nov., original holotype skeleton and cast, from the middle Eocene of Wyoming, USA. a Epoxy resin cast (TMP 2013.05.16) of holotype skeleton, in dorsal aspect and lightly coated with ammonium chloride to enhance details and texture; note this is a positive copy (i.e. bones depicted in relief) of the original specimen; same magnification as (b). b Orginal holotype skeleton (BHM-123), natural mold in dorsal aspect and reversed for comparison with a (for photograph of specimen in its original perspective and with some permineralized bone still in place, see Grande 1984, fig. III.1a); scale bar 10 mm. c Detail of skull in epoxy resin cast (TMP 2013.05.16) of holotype, in dorsal aspect and lightly coated with ammonium chloride to enhance details and texture; the juvenile status of this individual is supported, in part, by its frontoparietal complex consisting of three distint components (left and right frontoparietals and posterior median element); arrows point to grooves for arteria orbitonasalis extending across posterolateral corners of frontoparietals. d Detail of skull of original holotype (BHM-123), natural mold in dorsal aspect and reversed for comparison with (c); scale bar 10 mm. e Interpretative line drawing of holotype skeleton of Eopelobates deani sp. nov., based on original holotype skeleton (BHM-123) and cast (TMP 2013.05.16) of holotype. Mt metatarsal, proc. processus, V vertebra


Photographs of Eopelobates deani sp. nov., paratype skeleton from the middle Eocene of Wyoming, USA. a Main slab (SMNK PAL 6659a) preserving nearly complete skeleton, in dorsal aspect; note fossils of dipteran larvae scattered around frog skeleton; scale bar 10 mm. b Counterpart of the same individual (SMNK PAL 6659b), preserving partial skeleton plus imprints of bones, in ventral aspect; scale bar 10 mm. c Detail of skull on main slab (SMNK PAL 6659a), in dorsal aspect; the more adult status of this individual is supported, in part, by its frontoparietal complex having the left and right frontoparietals fused posteriorly and the posterior median element being largely fused to the posterior ends of the frontoparietals; arrow points to groove for arteria orbitonasalis extending across posterolateral corner of right frontoparietal; magnification about 4.4× larger than (a). d Detail of pelvic girdle and median and posterior portions of vertebral column on counterpart slab (SMNK PAL 6659b), in ventral view; magnification about 3.8× larger than (b). e Interpretative line drawing of paratype skeleton of Eopelobates deani sp. nov, based on part and counterpart slabs (SMNK PAL 6659 a and b, respectively). Mc metacarpal, Mt metatarsal, proc. processus, V vertebra
1984 probably a new species of the genus Eopelobates:
Grande, p. 186
1984 possibly Eopelobates sp.: Grande, caption fig. III.1a
1998 “Eopelobates” sp.: Sanchiz, p. 51
1999 probable new species of Eopelobates:
Gardner, p. 457
2000 “Green River pelobatid”: Roček and Rage, table 2
2000 pelobatid anuran: Roček and Rage, caption fig. 22
2002 “Green River Eopelobates”: Henrici, p. 242
2003 Pelobates–Eopelobates-like frog: Holman, p. 104
2013 probable new species of the pelobatid genus
Eopelobates: Henrici et al., p. 295
2013 †“Eopelobates”: Grande, p. 187
Diagnosis: Cranial sculpture weakly developed, consisting of horizontal striae on the anterior part of the maxillae and of low, irregular ridges and pits on the nasals, on the posterior part of the frontoparietal complex, and on lamella alaris of squamosals; base of pars facialis of premaxilla broad and placed medially (occupies medial two-thirds of the horizontal part of the bone) and medial margins of both processes in moderately broad contact across the midline; nasals widely separated from one another, their medial margins divergent posteriorly, and their processus lateralis short, pointed, and directed laterally; sphenethmoid exposed in a large gap between nasals and frontoparietals; processus lateralis inferior of frontoparietal extensive laterally, slender and pointed, its lateral portion is partly separated by a groove for the orbitonasal artery; anterior part of lamella alaris of squamosal is slender and tapered to a sharp point; urostyle separated from sacral vertebra by a joint; tibiale and fibulare not coalesced; carpus and distal tarsus not fully ossified in adults.
Holotype: BHM-123 (original number BHI-123), an articulated and nearly complete skeleton preserved in dorsal aspect as a natural mold (i.e. negative impression) in a slab of tan coloured siltstone. Initial preparation at BHI left some poorly preserved bone in the distal portions of the limbs (see Grande 1984, fig. III.1a); subsequent preparation at FMNH removed all traces of bone, resulting in clean impressions of the bones (Fig. 4b, d). Positive copies (i.e. depicting bones in relief) made from the fully cleaned, original specimen include: FMNH PR 1613 (cast); SMM P78.8.29 (peel); and TMP 2013.05.16 (cast; Fig. 4a, c).
Type locality and age: The holotype was recovered from a commercial dimensional stone quarry (one of the G-4 localities in fossil Lake Gosiute; see Grande 1984, fig. I.4 and p. 14) near Farson, Sweetwater County, southeastern Wyoming, USA, in the Laney Member, Green River Formation (Grande 1984, caption fig. III.1a; Robert Farrar, pers. comm. 2013). Litho- and biostratigraphic correlations indicate that the Laney Member is early middle Eocene in age or equivalent to the Bridgerian NALMA (e.g. see Grande 1984, 2013; Krishtalka et al. 1987; Smith et al. 2010, fig. 3). This is corroborated by a radiometric age of about 49 Ma from an ash bed near the top of the Laney Member (Smith et al. 2008, 2010).
Paratype: SMNK PAL 6659a, b (part and counterpart, respectively), a nearly complete, articulated, and permineralized skeleton split horizontally into part and counterpart slabs: most of the skull, girdles and limbs are exposed in dorsal aspect on the part slab (Fig. 5a, c), whereas most of the axial skeleton, lesser portions of the rest of the skeleton and impressions of some bones are exposed in ventral aspect on the counterpart slab (Fig. 5b, d). This specimen was purchased in the 1980s from the commercial fossil dealer Jurgen Henzel (deceased), but without any locality information beyond that it came from the Green River Formation. The consensus among knowledgeable collectors and dealers in Green River Formation fossils is that this specimen likely came from the “mini-fish beds” of the Laney Member, in the Little Colorado Desert area, near Fontanelle, Sweetwater County, Wyoming (Robert Farrar, pers. comm. 2013). If that is correct, the paratype locality is close both geographically and in age to the holotype locality. The occurrence of multiple Diptera (i.e. fly) larvae on the part slab is striking, but uninformative for resolving the provenance of this frog, because dipteran larvae are known from numerous horizons and localities within the Green River Formation, and may be locally abundant (e.g. Bradley 1931, pp. 50–51, pl. 2B; Mason 2011, fig. 1).
Etymology: In honour of James Dean, Sr. (deceased) of Hill City, South Dakota, USA, who discovered the holotype specimen in the 1970s while quarrying for building stone and generously donated the slab to the BHI.
Stratigraphy and distribution: Green River Formation (Laney Member); early middle Eocene or Bridgerian NALMA; Wyoming, USA.
Description of holotype: Our description of the holotype skeleton is based on the original specimen (BHM-123) and on two epoxy resin casts (FMNH PR 1613 and TMP 2013.05.16). A preliminary description of the holotype presented by Roček and Rage (2002, pp. 1361–1363, fig. 22) was based solely on the replica FMNH PR 1613. The original specimen is challenging to interpret, because it is a natural mold depicting the bones as negative impressions. The casts are easier to intepret, because those replicas depict the bones in positive relief as they would have appeared in life. However, details of bone surfaces and some other features are more easily visible in the original. For ease of description and to better reflect the in-life appearance of the skeleton, here we have reversed our photographs of the original specimen (Fig. 4b, d) so that it is depicted in the same perspective as our photographs of the cast (Fig. 4a, c) and interpretive line drawing of the skeleton (Fig. 4e). To be consistent with the accompanying images, in our description “left” and “right” refer to the anatomical left and right sides, rather than to the left and right sides in the original (negative) specimen. For a photograph of the original specimen in its preserved (i.e. negative and not reversed) perspective, see the photograph provided by Grande (1984, fig. III.1a).
The holotype is a nearly complete and mostly articulated skeleton of a small sized, metamorphosed frog preserved in dorsal aspect. As is typical for slab style frog skeletons, dorsoventral compaction has resulted in minor crushing and displacement of some bones. Most of the bones in the skeleton are fully or partially visible. The exceptions are that most of the pectoral girdle is absent (except for the left scapula + cleithrum) and that much of the palatal region and the mandibles are obscured by overlying skull roof and cheek bones. Indistinct staining in the wrist and ankle regions likely represents traces of cartilaginous or weakly ossified bone. Slightly roughened matrix surfaces associated with the skeleton appear to represent traces of soft tissue, as follows: subcircular patches within the orbits are remnants of the eyes, whereas areas alongside the trunk and proximal portions of the hindlimbs represent the body outline. As preserved, the distance between the anterior edges of the premaxillae and the posterior edges of the ischia is 36 mm. Allowing a millimeter or so for rotational displacement of the premaxillae and the pelvis (see below), we estimate a SVL of 35 mm. This relatively small body size, in conjunction with certain other features (e.g. boundaries between three ossifications in frontoparietal complex remain visible; transverse processes on V4 and probably more anterior presacrals do not appear to have been finished in bone [i.e. those ends were cartilaginous; see below]; no epiphyses on either ends of femur and tibiofibula or on the proximal end of humerus; no ossified carpal or tarsal elements) indicate that the skeleton is from an immature, postmetamorphic individual.
As preserved, the skull (Fig. 4c, d) has maximum dimensions of 14 mm long and 16 mm wide and it is approximately triangular in dorsal outline. Those measurements and outline are exaggerated by dorsoventral compaction of the skull and minor displacement of bones. In life the skull would have been more vaulted and the snout and cheek region more broadly rounded in dorsal outline. Dorsoventral compaction is most evident along the skull roof midline, where the medial edges of the left and right frontoparietals are tilted downwards and now lie below their respective lateral edges. The paired nasals lie in a nearly horizontal plane, presumably as a result of their medial portions also having been pushed downwards. The two sets of upper jaw bones are rotated out of their original orientation, but in opposite directions: the premaxillae are rotated anteriorly to expose their inner surfaces, whereas the maxillae are rotated inwards to expose their outer surfaces. On both sides the squamosal and quadratojugal retain their close associations with the adjacent maxilla and, like that bone, have also been rotated inwards and displaced slightly laterally. Bones in the occipital region do not appear to have been compacted or displaced to any extent. Portions of certain underlying skull bones (dentaries, vomers, and pterygoids) also are visible.
The external surfaces of both premaxillae are embedded in matrix, so nothing can be said about that surface. Each premaxilla is relatively narrow and dorsally bears a well-developed pars facialis. This process is relatively broad (i.e. occupies about the medial two-thirds of the bone), moderately tall, and asymmetrically hourglass shaped in outline, being slightly constricted midway along its height and having its lateral edge more deeply concave than the medial. Although the premaxillae are rotated anteriorly out of alignment, in life their partes faciales would have been separated across the midline by a narrow slit between their concave medial margins, but probably were in contact ventrally and dorsally. Both the medial and lateral margins of the inner surface of the pars facialis are thickened, and bracket between them a deep depression (as is the case with extant Pelobates: Roček 1981, fig. 25b). The dorsal end of the pars facialis is moderately expanded laterally and its dorsal edge is shallowly convex in outline. On each premaxilla, a small processus palatinus protrudes posteriorly from the medial end of the horizontal lamina and, in life, the straight medial edges of those processes would have been in median contact. Four tiny and closely spaced teeth are visible along about the lateral half of the crista dentalis on the right premaxilla. Assuming that tooth spacing was consistent across the entire premaxillary tooth row, we estimate that each premaxilla bore about eight teeth. The maxillae are exposed in external (=lateral) aspect. The anteriormost end of each maxilla is partly covered by matrix. Nevertheless, it is clear that each maxilla is moderately elongate (i.e. extending only a moderate distance past the level of the posterior margin of the orbital rim) and shallowly triangular in outline, consisting of a moderately deep preorbital region, an elongate and shallowly concave margo orbitalis, and a posteriorly short and tapered postorbital region. The external surface of the maxilla is weakly sculpted with faint horizontal striae that are best developed across the preorbital portion of the bone, but there is no indication of the pit-and-ridge style sculpture seen on the outer surfaces of the squamosals and nasals. Enough of the ventral edge of the right maxilla is exposed to show that tiny, closely placed teeth are present along at least the anterior three-quarters of the maxilla. None of the maxillary or premaxillary teeth is well enough preserved to determine if they are pedicellate or bicuspid.
Quadratojugals are present on both sides of the skull and exposed in approximately dorsal aspect. On the right side the quadratojugal is displaced slightly posteriorly, but on the left it appears to be preserved in its original articulation with the posterior end of the adjacent maxilla. The exact nature of this articulation is difficult to resolve, but the quadratojugal appears to be sutured to the inner surface of the processus posterior of the maxilla. Both squamosals are preserved in lateral aspect and exhibit the classic, anteriorly tilted and T-shaped outline for that bone, consisting of a posteriorly directed processus posterodorsalis, an anteroventrally directed processus zygomaticus that is pointed and connects ventrally with the complementary processus zygomaticomaxillaris of the maxilla, and a posteroventrally directed, prong-like processus posterolateralis that connects with the quadratojugal. The lamella alaris portion of the bone is narrow (rather than being broadly expanded and plate-like as in some anurans, including Eopelobates grandis) and its external surface is weakly sculpted with low, narrow, and irregular ridges. The squamosal-quadratojugal contact is preserved on the left side, but it is slightly disarticulated on the right side. The squamosal-maxilla contact is preserved only on the right side, where it shows that the connection was moderately long.
The nasals appear to be in essentially their original position relative to the orbits, although they have been forced into a more nearly horizontal plane. In dorsal outline, each nasal is broader than long and in the form of a medially tilted triangle. The paired nasals are widely separated across the midline. Their anteromedial corners approach but do not contact one another, and more posteriorly their medial margins diverge laterally. The orbital and anterolateral margins of the nasals are nearly straight, and enclose between them a processus paraorbitalis that is relatively short and directed laterally. Due to rotation of the maxilla, it is uncertain whether the maxilla and nasal contacted one another. The dorsal surface of the nasal is weakly sculpted with low, irregular ridges similar to those on the squamosal. Between the posteromedial corners of the nasals and the anterior ends of the frontoparietals there is a large, rhomboid gap. We interpret that gap as a fontanelle through which the dorsal surface of the sphenethmoid was exposed. Within that gap and adjacent to the medial margins of the nasals, there are two small, symmetrical protrusions that likely belong to the vomers. The fact that these protrusions are visible suggests that the overlying sphenethmoid was still cartilaginous.
The frontoparietal complex is tripartite, consisting of a pair of left and right frontoparietals and an unpaired, posterior median element that is approximately subpentagonal in outline. Boundaries between the three components are well delineated by narrow grooves, but it is uncertain whether those represent sutures, cracks that formed along sutured or weakly fused contacts during compaction of the skull, or a combination of the two. The junction between the medial edges of the frontoparietals is in the form of a narrow slit. Although this slit may indicate that the left and right frontoparietals were not in direct contact along the skull midline, it is also possible that downwards rotation of the frontoparietals forced their medial edges slightly apart. All three elements dorsally bear faint pit-and-ridge style sculpture: this is broadly distributed across the much of the posterior median element, whereas it is limited to the posteriormost end and lateral portions of the frontoparietals. Along their posterolateral portions, both frontoparietals dorsally bear a deep groove (marked by arrows in Fig. 4c and more prominent on the left side) that represents the canal for the arteriae orbitonasales (see Roček 1981, fig. 28). In dorsal view the frontoparietals are moderately elongate. The portion between the orbits is moderately narrow, whereas the more posterior portion of the bone is broader. In terms of its maximum dimensions, the frontoparietal complex has a width about 80 % of its length. The margo orbitalis of the frontoparietal is anteroposteriorly straight along most of its length, but posteriorly (at the level of the otic capsules) it follows the medial margin of the above mentioned groove; a moderate convexity at the level of the anterior wall of the otic capsule is the processus lateralis superior. The lateral, lower part of the bone that extends onto the anterior surface of the otic capsule is the processus lateralis inferior, well prominent laterally in this species. Undercutting of the matrix along the margo orbitalis in the original specimen indicates that a tectum supraorbitale (=supraorbital ledge) was present, although that ledge does not appear to have been particularly broad in contrast to some other pelobatoids (e.g. Pelobates fuscus: Roček 1981, fig. 27). The posterolateral corners of the bone each bear a processus paraoccipitalis in the form of a moderately elongate, triangular, and posterolaterally directed prong.
At the rear of the skull, the exoccipital and prootic complexes are in their original positions, but it is unclear whether these bones are sutured or fused with one another on either side. These endochondral bones clearly were in at least an intermediate stage of ossification, based on the observations that a narrow bridge of bone forms the dorsal margin of the foramen magnum and, more laterally and ventrally, the bases of the widely spaced occipital condyles are visible.
Some dermal bones belonging to the ventral part of the skull—specifically the dentary and pterygoid—are partially exposed on one or both sides, but their shapes and contacts cannot be reconstructed. No other skull bones (i.e. parasphenoid, septomaxillae, angulars, or columella) can be identified.
The vertebral column is articulated, but it has suffered minor compaction to either side of the midline and a crack runs through the neural arches on V2–V8. Eight presacral vertebrae are present. These bear neural arches that are weakly imbricate (on the anteriormost two vertebrae) to non-imbricate (on the remaining vertebrae) and lack spinal processes. The neural arches of V1 and V2 are slightly larger than those of other presacrals and each minimally overlaps the succeeding vertebra in the series. V2–V4 are provided with robust transverse processes that are relatively uniform along their lengths (i.e. neither tapered nor expanded distally) and are relatively long. The longest transverse processes, on V3, are each subequal in length to the width of the neural arch roof; transverse processes on V2 and V4 are noticeably shorter. Transverse processes on V2 and V3 project laterally and slightly anteriorly, whereas those on V4 project laterally. The truncate lateral ends of transverse processes on V2–V4, which are especially evident on V4, suggest that the ends of those processes were capped in life by cartilage. By contrast, V5–V8 bear transverse processes that are shorter, thinner, tapered, and have bluntly rounded ends that suggest they were not capped by cartilage. Transverse processes on V5 project laterally, whereas transverse processes on the more posterior presacrals are inclined anteriorly. The sacral vertebra consists of a comparatively short centrum and neural arch, and it bears laterally projecting, dilated or wing-shaped transverse processes. Because the iliac shafts on both sides overlay the lateral ends of the sacral transverse processes, it is unknown whether the lateral margins of the sacral transverse processes are straight or laterally convex in dorsal outline and whether they were capped in cartilage. On both sides, an indistinct line originates at the base of the transverse process of the sacral vertebra and extends laterally and slightly posteriorly about two-thirds of the distance across the transverse process, thereby partially bisecting the anterior and posterior portions of each transverse process. A comparable suture or gap has been reported in some pelobatoid individuals (e.g. specimen of Pelobates decheni: Böhme et al. 1982, fig. 4), where it has been interpreted as indicating two centres of origin for the sacral transverse process. A somewhat W-shaped line lies at the boundary between the sacral vertebra and the urostyle. We are inclined to regard that line as indicating the presence of a bicondylar joint, in part, because a similar pattern is seen in the paratype skeleton (see below); however, as noted by one reviewer (A. Henrici, written communication 2014) that W-shaped line might instead be the posterior edge of the sacral neural arch. The anterior section of the urostyle is short and markedly broad. The urostylar shaft is weakly tapered posteriorly. The posterior end of the urostyle is obscured by the pelvic girdle, which has shifted slightly forward. Although the full length of the urostyle cannot be determined, it is clear that this bone is not especially long. Using the relative length ratio proposed by Maglia (1998), the urostyle is relatively short, being considerably less than the combined length of the presacral vertebral series. There is no indication of any processes arising off the anterior portion of the urostyle. The presence of a crest along the urostylar shaft is uncertain, due to compaction of the bone.
Little can be seen for the pectoral girdle, except for a broad and laterally expanded element on the left side. Based on its shape and relatively large size, this appears to be the scapula + cleithrum exposed in dorsal aspect. The pelvis is largely articulated but, as noted above, it has shifted slightly anteriorly; as a result, the posterior end of the urostyle is wedged against the interiliac symphysis and the anterior ends of the iliac shafts lie at about the level of the joint between V6 and V7. The ischia remain in their original articulation with one another and are exposed in dorsal aspect, but the ilia have been slightly splayed apart (i.e. dorsal edges have rotated laterally) and their shafts have been slightly crushed mediolaterally. Both ilia bear a low dorsal prominence and lack an iliac crest. The presence of an oblique (=spiral) groove is unclear, but we see no convincing indication of that structure.
Both fore limbs are slightly displaced relative to the vertebral column, more so on the right side than the left, but each limb remains articulated. Bones of the fore limbs are preserved in lateral aspect. The fingers are well preserved, with only some minor overlapping, and are exposed in dorsal aspect. No epiphysis is present on the proximal end of the humerus. Otherwise, the humerus and radioulna are unremarkable. The phalangeal formula for the manus is 2-2-3-3. No prepollex or any definitive carpal elements are observed; however, a small and semicircular stain that can faintly be seen on the matrix surface in the region of the left wrist, proximal to the bases of the 3rd and 4th fingers, may represent a cartilaginous or weakly ossified carpal bone. Bones of the hindlimb are preserved in almost perfect articulation and in essentially dorsal aspect. The femur, tibiofibula, tibiale, and fibulare are relatively gracile in build. No epiphyses are present on either ends of the femur or the tibiofibula. Both of those bones are only barely sigmoidal in outline. The femur is shorter than the tibiofibula (F:TF 0.94), and F + TF is markedly shorter than the SVL (0.79). The tibiale and fibulare are not coalesced, although their proximal and distal ends are closely appressed. The only indication of a distal tarsal element is a faint imprint on the right side, between the distal end of the tibiale and the bases of the first and second toes, that may represent the centrale. No prehallux is observed. The phalangeal formula for the pes is 2-2-3-4-3.
Description of paratype: Unlike the above-described holotype, the paratype is a permineralized skeleton preserved in positive relief on two slabs. Most of the skeleton is exposed in dorsal aspect in the part slab (SMNK PAL 6659a: Fig. 5a, c) and the remainder is exposed in ventral aspect in the counterpart slab (SMNK PAL 6659b: Fig. 5b, d). The interpretive line drawing (Fig. 5e) combines information from both slabs. The paratype is from a slightly larger individual, having a SVL of 37 mm (measured as the distance between the anterior edges of the premaxillae and the posterior edges of the ischia), which is only a few millimetres larger than the holotype. This slightly larger body size, coupled with several other age-dependent features (e.g. frontoparietal complex partially fused and its dorsal sculpture slightly more prominent; some carpal elements ossified), indicate that the paratype was a more mature individual.
The paratype skull (Fig. 5c) is dorsoventrally flattened and subtriangular in outline. Probably because the premaxillae remain essentially in their natural position (as opposed to having been rotated forward as in the holotype), the snout is moderately broad and blunt; that likely reflects the in-life profile of the snout. Except for the maxillae, most of the other visible skull bones have largely retained their approximate positions relative to adjacent bones.
The outer surfaces of both premaxillae and maxillae are exposed. Although the premaxillae are in their natural positions, the maxillae are not. The outer surface of the premaxilla is moderately rugose. The pars facialis has a broad base that extends up to the medial margin of the bone. Its medial margin is nearly straight, thus forming a long symphysis extending from the crista dentalis up to the tip of the pars facialis. The lateral margin of the pars facialis extends dorsomedially, meeting the medial margin in a point. The number of tooth positions (well preserved in the right premaxilla) is 16. Both maxillae are broken midway along their lengths. The anterior portions of both maxillae are rotated inwards, so that their outer surfaces now lie nearly in the horizontal plane and the teeth are visible. The posterior portions of both maxillae have either retained their original orientation or may have rotated slightly outwards; regardless, because the crista dentalis along this portion is embedded in matrix, the posterior extent of the maxillary tooth row cannot be determined. The maxillae are long and their external surfaces are smooth, except for shallow, anteroposteriorly elongated pits along the lower edge of the anterior part of the bone. The processus frontalis forms an obtuse angle. The processus palatinus is partly exposed between the maxilla and nasal on the right side. The left maxilla shows that the processus posterior is comparatively short. The processus zygomaticomaxillaris is low and indistinct.
Both quadratojugals are disarticulated from the maxillae. The right quadratojugal is preserved in dorsal aspect. Its dorsal surface is smooth and rounded. The squamosal, although fragmentary, is preserved on the left side, whereas it is less well preserved on the right side. On both sides, the lamella alaris is slender anteriorly and tapers to a point, but its posterior part is crushed and broken away in the right squamosal. The outer surface of the lamella alaris is smooth and confluent with the outer surface of the processus posterolateralis.
Both nasals are preserved in their original positions. Their medial margins are close, but not in contact anteriorly; more posteriorly, their medial margins diverge laterally at a moderate angle. The processus anterior is rounded, whereas the processus paraorbitalis is pointed and directed laterally. The anterolateral margin is nearly straight.
The frontoparietals are separated anteriorly by a narrow, anteroposteriorly oriented slot, which becomes a suture posteriorly. In its posterior section, the suture changes direction and extends posterolaterally to the right. On the opposite side of the bone, a similar suture was probably obliterated. The posterior margin of the bone is convex, suggesting that the unpaired posterior median element was involved. This part of the skull is compressed onto the dorsal surface of the tectum synoticum and, as a result, on both sides the processus paraoccipitalis has been broken; in spite of that, their size and shape can be reconstructed. They are slightly less prominent than the prominentia of the canalis semicircularis posterior. The lateral margins of the frontoparietals extend into a pointed processus lateralis inferior, which adjoins the anterodorsal surface of the prootic, thus forming the posteromedial portion of the margin of the orbit. Because the whole skeleton is dorsoventrally compressed, the processus lateralis superior is not prominent, but it can be recognised by the position and path of the groove for the arteria orbitonasalis that separates the processus lateralis inferior from the main body of the bone (marked by arrow in Fig. 5c). We interpret these grooves as having been dorsally open in life, because there is no obvious indication of broken surfaces alongside the gooves that suggest a bony roof was originally present, but lost when the specimen was split into part and counterpart slabs. Both frontoparietals are covered posteriorly by a pitted sculpture, but they are smooth anteriorly. Impressions of the pits are observable in SMNK PAL 6659b (Fig. 5b).
Anterior parts of both dentaries are exposed by their inner (left) and outer (right) surfaces, and are disarticulated from one another. The pterygoids are crushed and partly exposed in dorsal aspect. The sphenethmoid is exposed between the posterior parts of both nasals and the anterior margins of the frontoparietals; apparently, the nasal septum was not yet ossified. The roof of both otic capsules is broken inside, yet the anterior, lateral, and posterior semicircular canals (not labelled in Fig. 5) remain intact and prominent.
Vertebral centra are procoelous, as can be judged by imprints of the posterior presacrals in SMNK PAL 6659a. There are eight presacrals. V2–V4 bear robust and perpendicular transverse processes, whereas V5–V8 bear thin transverse processes that are inclined anteriorly. V9 is the sacral vertebra and it bears widely dilated transverse processes. The left sacral transverse process seems to originate from the posteriormost presacral vertebra, whereas the right process originates from the sacral vertebra, unless the gap between both is an artifact. The urostyle is long, reaching the symphysis of both ilia, and it is articulated to the sacral vertebra by a bicondylar joint.
Unlike the holotype, the paratype preserves a substantial amount of the pectoral girdle. The scapula is exposed on the right side by its lateral margin, which was completed by cartilage of the suprascapula in the living animal. The suprascapula is represented by a thin, brownish layer, nearly completely preserved on the left side. A well developed cleithrum is preserved on the right side. The sternum has been displaced and is now preserved next to the right iliac shaft in SMNK PAL 6659a. The pelvic girdle is articulated (Fig. 5d), but not especially informative because it is crushed and missing portions of both ilia and ischia.
Both fore limbs are articulated, but have been slightly separated from the pectoral girdle. The humerus and radioulna are preserved on both sides, but except for the distal end of the radioulna, which is sharply truncated perpendicularly, suggesting that the epiphysis was still cartilaginous, these bones provide no significant information. The carpus is preserved in the right fore limb and also partly in the left fore limb. Three elements in the proximal row may be the ulnare (plus pisiforme), the proximal centrale, and the radiale (the last two seem not to be fused). In the distal row, three elements on the ulnar side may be distal carpals 4 to 6, the one element in the middle of the distal row may be centrale 3, and the large element on the radial side may represent the element Y and distal carpals 1 to 3; all identifications based on carpus of advanced larva of Pelobates fuscus (from Schmalhausen 1907, interpretation after Fabrezi and Alberch 1996). The phalangeal formula for the manus is 2-2-3-3. Both hindlimbs are articulated and attached to the body. The tibiale and fibulare are long, slender and separated from one another. The distal tarsus was not ossified. The phalangeal formula for the pes is 2-2-3-4-3.
Remarks: We interpret the holotype and paratype skeletons as being from immature, postmetamorphic individuals because they lack epiphyses on the proximal end of the humerus and on both ends of the femur and tibiofibula, have only a few ossified or weakly ossified carpal (paratype) or tarsal (holotype) elements, have unfinished transverse processes on at least V4, and in the frontoparietal complex the posterior median element is at least partially visible and the three bones forming the complex bear only weak dorsal sculpture. The paratype appears to be from a slightly more mature individual based on its slightly larger body size (SVL about 37 mm versus 35 mm) and in having the posterior median element partially fused with the frontoparietals (versus completely unfused in holotype), slightly more prominent pitted sculpture on the posterior part of the frontoparietals (versus faintly developed in holotype), and ossification of some carpal elements has commenced (versus none ossified in holotype). Even though BHM-123 is the less mature individual, we erect it as the holotype for the following reasons: (1) it clearly exhibits the tripartite frontoparietal complex that is charactistic for Eopelobates (that is less apparent in the paratype due to its slightly more advanced stage of development); (2) its locality information is reliably documented; and (3) that specimen is well entrenched in the literature (e.g. Grande 1984; Henrici 2002; Henrici et al. 2013) as belonging to a probable new species of Eopelobates.
A suite of features seen in both specimens are potentially age-dependent, but among other anurans may also occur in mature adults and, thus, might be taxonomically important for identifying the new species. Those features include: weakly developed cranial sculpture; nasals widely separated; frontoparietals paired or weakly sutured; canals in frontoparietals for arteriae orbitonasales not roofed in bone; poorly ossified sphenethmoid without osseous nasal septum and exposed in a large, rhomboid gap between the nasals and frontoparietals; vertebrae bearing weakly or non-imbricate neural arches that lack spinal processes; sacral-urostyle joint not fused; relatively short posterior limbs; tibiale and fibulare not coalesced; and carpal and tarsal elements not fully ossified. Many of those features are included in our diagnosis for this species, because they help differentiate it from other congeners.
Both specimens share additional features that are not as obviously age dependent and, thus, are also useful for differentiating the new species. Those features include: pars facialis on premaxilla relatively broad, situated medially, and contacts its counterpart from the opposite side in a long median suture; nasals have medial margins divergent posteriorly and their processus paraorbitalis is pointed, short, and directed laterally; anterior part of the lamella alaris of squamosal is slender and tapered in a sharp point; processus lateralis inferior of frontoparietal is long, slender, and pointed and it adjoins the anterodorsal surface of the prootic; and processus paraoccipitalis of frontoparietal is relatively prominent
Compared to the only other currently recognised North American species of Eopelobates—namely E. grandis from the late Eocene of South Dakota (see next account)—E. deani is distinct in its substantially smaller body size, cranial sculpture that is less prominent, consisting of irregular ridges and pits (versus reticulations) and more restricted in its coverage, squamosals that are more gracile and have a narrower lamella alaris, frontoparietals that are relatively narrower between orbital margins and bear a more prominent processus paraoccipitalis, and in having quadratojugals and an unfused tibiale and fibulare (versus quadratojugals may be absent and tibiale + fibulare may be fused distally in E. grandis; see next account).
Besides the above noted differences in frontoparietal sculpture and ossification of the frontoparietal complex and carpal elements, the holotype and paratype individuals differ in three other features: (1) the dorsal end of the pars facialis on the premaxilla is broad and shallowly convex in the holotype, but tapers to a point in the paratype; (2) squamosals are weakly sculpted externally in the holotype, but smooth in the paratype; and (3) the number of tooth positions on the premaxilla is estimated to be 8 in the holotype, but is 16 in the paratype. Considering the number of similarities between the holotype and paratype, we are inclined to attribute those differences to intraspecific variation. The higher premaxillary tooth count in the paratype may also be due, at least partly, to the slightly larger size and more mature age of that individual.
Eopelobates grandis Zweifel, 1956
Fig. 6

Micro-CT scans of Eopelobates grandis Zweifel, 1956, incomplete holotype skeleton (YPM-PU 16441), from the latest Eocene of South Dakota, USA. Note that since this specimen was first described it has been subject to further preparation, the right nasal has been removed, and unfortunately it has suffered some damage (cf. photographs provided by Zweifel 1956, figs. 1, 3; Henrici 2002, figs. 1–3); images here depict the holotype skeleton as of October 2013 and minus the matrix in which it is partially embedded. a, b Entire skeleton, in dorsal (a) and ventral (b) aspects; scale bar (upper centre) 10 mm for these two images only. c Detail of right maxilla in inner view; arrow points to contact area (?weakly developed processus pertygoideus) for ramus maxillaris of pterygoid; magnification about 1.4× larger than (a) and (b). d, e Detail of skull, in dorsal (d) and ventral (e) views; magnification about 1.4× larger than (a) and (b). f Detail of posterior portion of pelvic girdle (same as area marked by dashed rectangle in a), in dorsal aspect; two features in this region suggest the individual was not fully mature, specifically the absence of epiphyses in long bones (marked by arrow at bottom centre) and the exposed spongy bone of ischia that in life would have been covered by cartilage); magnification about 2.6× larger than (a). g Detail of posterior section of presacral vertebral column (same as area marked by dashed rectangle in b), in ventral aspect; arrows point to what we interpret as calcified intervertebral discs between the amphicoelous centra; magnification about 3.5× larger than (b). proc. processus, V vertebra. Images copyright of Yale Peabody Museum
Diagnosis (modified from Henrici 2002): Prominent and extensive pit-and-ridge sculpture covers all dermal bones of the skull, pits may be arranged in loose rows and some adjacent pits may coalesce, but not to the extent of forming elongate grooves; nasals have broadly convex medial margins and probably contacted one another along at least the middle portion of their medial margins; sphenethmoid minimally or perhaps not exposed between posterior parts of nasals and anterior margin of frontoparietal; frontoparietal relatively short and wide, only slightly wider anteriorly than posteriorly, processus lateralis inferior of frontoparietal not exceeding the processus lateralis superior (the latter could be absent), with its posterior margin straight or moderately concave and bearing reduced, spine-like processus paraoccipitalis; anterior part of lamella alaris of squamosal broad, its contact margin with maxilla long and concave (corresponding margin of processus zygomaticomaxillaris on maxillae widely convex); urostyle and sacral vertebra unfused in fully grown individuals; tibiale and fibulare possibly fused at their ends.
Material: The only known specimen is the incomplete, permineralized holotype skeleton (YPM-PU 16441) described by Zweifel (1956) and subsequent authors (Estes 1970; Henrici 2002; this study).
Stratigraphy and distribution: Chadron Formation (Ahern Member); latest Eocene or Chadronian NALMA; South Dakota, USA (Zweifel 1956; Henrici 2002).
Description: The holotype (YPM-PU 16441) and only known specimen has previously been described by Zweifel (1956), Estes (1970), and in considerable detail by Henrici (2002). Our description here is based on micro-CT scans (Fig. 6) of the holotype and it focuses mostly on characters not mentioned by or differing from those reported by previous authors. YPM-PU 16441 is the largest known example of an Eopelobates individual (Table 1). The estimated SVL of 82 mm provided by Henrici (2002) is too low, whereas the value of 110 mm originally proposed by Zweifel (1956) seems more accurate. The higher estimate is supported by our measurement of the specimen as preserved and by the fact that any size estimate needs to take into account that the snout is broken away and that the posterior portion of the vertebral column (V8 and sacral vertebra), urostyle, and pelvic girdle all have been displaced forwards. That anterior displacement is best seen in ventral aspect (Fig. 6b).
Sculpture covers all preserved dermal bones of the roof and cheek (i.e. nasals, frontoparietals, maxillae, and squamosals) and is more extensive in both coverage and prominence than on any other Eopelobates specimen known to us. Micro-CT scans reveal that sculpture is of the classic pit-and-ridge type, consisting of flat bottomed, modest sized and irregular shaped pits that are bordered by narrow ridges. In some areas (e.g. ventral portion of maxilla and anteromedian portion of frontoparietals) the pits are arranged into loose rows and may partially coalesce, but not to the extent that they form elongate and well delineated grooves.
The premaxillae are not preserved. The left nasal remains in place, but it is fractured and its margins are not intact. The right nasal, which originally was present and complete (see Zweifel 1956, fig. 1), has since been detached (see Henrici 2002, fig. 6A) and was not available for our study. The anterolateral margin of the nasal is nearly straight (or slightly concave: see Zweifel 1956, fig. 1) and the medial margin is broadly convex (see Henrici 2002, fig. 6A). Judging by the position of the left nasal as preserved on the skull (Fig. 6d) and the outline drawing of the detached right nasal (Henrici 2002, fig. 6A), in life the paired nasals probably contacted one another across at least the middle portions of their broadly convex medial margins. The dorsal surface of the sphenethmoid is now exposed and is smooth, but it is unclear whether any portion of that bone would have been exposed dorsally between the nasals and the frontoparietals. The frontoparietal is flat dorsally, with its margo orbitalis only slightly concave in dorsal outline and with its processus lateralis superior arising at about the anteroposterior midlength of the bone. The posterolateral corners of the frontoparietals each bear a tiny, spine-like processus paraoccipitalis. The posterior margin of the frontoparietal complex is essentially transverse, with only a hint of a small medial bulge or convexity (see Henrici 2002, fig. 4A) that may represent the fused posterior median element. The existence of a median suture between the frontoparietals is similarly uncertain. No such median suture is apparent in the original photograph and interpretive drawing (see Zweifel 1956, figs 1, 2), yet in his interpretive line drawing Estes (1970, fig. 13A) depicted a nearly straight median suture that completely bisected the entire frontoparietal complex in a manner incompatible with referral to Eopelobates (or Pelobates). In her detailed redescription of the specimen, Henrici (2002, pp. 244–245) stated “A median suture is visible … in a broad, deep groove that extends two-thirds of the length from the anterior end [of the frontoparietal complex]. The groove appears to be the result of preparation to make the median suture more visible. Posteriorly the groove narrows and then disappears near the widest point of the frontoparietal.” Due to the dense sculpture and some adhering matrix, Henrici (2002) was not able to trace the groove more posteriorly. The fact that the groove appeared to have been widened to enhance the suture is a troubling observation may call into question the reality of the suture. Although not noted by Henrici (2002) it is clear from her accompanying photograph and interpretive line drawing (Henrici 2002, figs. 3A, 4A) that the groove or suture is not essentially straight anteroposteriorly, but instead is convex and arcs to the right. Our micro-CT scans reveal that the frontoparietal complex is more fractured than previously realised (cf. Fig. 6a vs. Henrici 2002, figs. 3A, 4) and this raises further doubts about whether any median suture is present or can be reliably recognised.
The maxilla (Fig. 6d) is comparatively short and deep, its pre- and postorbital portions are similar in height, and its postorbital portion is not especially elongate posteriorly. The processus zygomaticomaxillaris is broadly convex dorsally. Micro-CT scans provide the first clear view of the entire inner surface of the maxilla (Fig. 6c). A lamina horizontalis is present and appears to terminate in a lingually directed, triangular, and somewhat trough-like projection (marked by arrow in Fig. 6c) that may represent the processus pterygoideus. The tooth row (estimated at 55–60 tooth positions) extends far behind the posterior termination of the horizontal lamina. It is difficult to assess from the micro-CT scans if the lower margin of the maxilla behind the end of the tooth row is intact and inclined posterodorsally towards the posterior end of the bone or if that inclined portion is due to the posteroventral part of the bone having broken off (Fig. 6c). As preserved, the processus posterior is short and tapers rapidly to a subtle point. Assuming it is not a preservational artifact, the form of the processus posterior suggests there was little or no contact with a quadratojugal (see below). More anteriorly, the maxilla bears a prominent, almost scoop-like processus palatinus that projects at an oblique anterodorsolateral angle well above the dorsal edge of the pars facialis; its apparently truncate free end suggests the process is not complete (see Fig. 6c, e). The structure may represent the palatine coalesced to the maxilla. The lamella alaris of the squamosal is extended lateroventrally, thus partly covering the base of the processus posterolateralis; the tip of this extension represents the posterior end of the squamoso-maxillary suture. Just as the processus zygomaticomaxillaris of the maxilla is broadly convex, the corresponding contact margin on the ventral margin of the squamosal is concave and extends anteriorly and slightly ventrally to terminate in a blunt point.
The small and elongate bone with a flared end (labelled “?” in Fig. 6d, e) that partially overlays the posterior end of the right maxilla was interpreted as the stapes by Zweifel (1956) and as the quadratojugal by Estes (1970) and Henrici (2002). Our micro-CT scans reveal this element is actually a small endochondral bone with a marrow cavity and, thus, it cannot be the quadratojugal because that is a solid dermal bone. This mystery bone may instead be a displaced phalange. No obvious quadratojugal is exposed on the surface or revealed by our micro-CT scans. There is no way to tell whether this bone was lost post-mortem or absent in life as in Scaphiopus. The torus dentigerus of the right vomer is well preserved (Fig. 6e), but the remaining parts of the bone are broken in fragments. The angular is preserved in ventral aspect; its coronoid process is moderately prominent medially.
The vertebral column consists of eight presacrals. Curiously, the vertebral centra are well ossified, with concavities both anteriorly and posteriorly; however, posterior to each centrum there is a transparent lens-like element, which can be interpreted as calcified intervertebral body (marked by arrows in Fig. 6g). The sacral vertebra plus the anterior part of the urostyle was originally exposed dorsally (Zweifel 1956, fig. 1), revealing transverse processes that were greatly dilated posteriorly (Zweifel’s interpretative drawing in his fig. 2 is not accurate). In its current condition, the holotype preserves only a small portion of the left transverse process (Fig. 6f). The observation that the centrum of the sacral vertebra was lost, but the anterior end of the urostyle was not, suggests that those elements were not fused together. It should also be noted that the anterior end of the urostyle above the neural canal is broken and exposes the inner structure of the bone. Zweifel (1956, p. 6, figs. 2, 6) and Henrici (2002, p. 251) reported a pair of short, posterolaterally directed transverse processes on the anterior portion of the urostyle (= “coccyx” in Zweifel’s terminology). It is not clear from our micro-CT scans whether those transverse processes are present; they may have been broken off at some point since Henrici’s (2002) study.
Both coracoids and scapulae are present, but they are fragmentary and partly overlain by other elements. The sternal style is now partly lost, but it was originally well preserved (Zweifel 1956, fig. 3; Henrici 2002, figs 2, 3B). The right humerus of the individual has been twisted along its long axis, so it is now displayed in ventral view on the dorsal surface of the specimen (Fig. 6a; see also Henrici 2002, fig. 1). It is obvious that the articular head has shifted medially, and that the fossa cubitalis was deep and well defined. The ilia are displaced anteriorly and rotated laterally about 90°; on each their shaft is rather strongly convex dorsally and there is no dorsal process. However, the anterior two-thirds of the shaft seems to be provided with a low dorsal crest (also mentioned by Henrici 2002, p. 252) that is paralleled by a shorter, medial ridge, the latter of which does not reach the anterior end of the shaft. There also is a longitudinal depression between the medial ridge and the dorsal crest. Considering that this ridge and crest are present on both ilia, they may be real features rather than an artifact caused by mediolateral compression of the bone. The ilia also appear to lack an oblique groove (Fig. 6a). Dorsal and ventral margins of the ischia were rimmed by cartilage, and both ischia, although coalesced to one another, are disarticulated from the ilia; this may be seen as the evidence that there was no ossified pubis (contra Zweifel 1956, p. 6). The femur and tibiofibula are of the same length (47.2 mm; Zweifel 1956) and the former is strongly sigmoidal in shape (Fig. 6b). Both sides preserve an incomplete tibiale and fibulare, the distal ends of which are in close contact (Fig. 6a, b). Damage along the distal portions of each tibiale and fibulare pair, however, prevents us from determining whether they were in loose contact or coalesced. We suspect the latter is true, based on the observation that despite the shafts of each tibiale and fibulare having been snapped apart postmortem, their distal ends remain in close contact on both sides of the skeleton.
Remarks: Imaging of the holotype skeleton of Eopelobates grandis using micro-CT scans has provided new information about this specimen, but has not been able to decisively resolve the higher level affinities of the species. In some cases, the scans have clarified potentially critical details. A good example is the pattern of cranial sculpture, which is challenging to interpret when visually examining the specimen, because in many places the sculpture is partially obscured by matrix that cannot safely be removed from the bone surfaces (see Henrici 2002, fig. 3A). Based on what she could see of the cranial sculpture, Henrici (2002, p. 243) described it as “extremely reticulated and tubercular” and in her revised diagnosis she listed “dermal sculpture highly tuberculated” as a feature that differentiated E. grandis from other congeners. Our micro-CT scans reveal that cranial sculpture in the holotype of E. grandis is, in fact, of the usual pit-and-ridge pattern seen in members of the genus. The tuberculations seen when visually examining the specimen are simply raised portions of the ridges that project above the surface of the matrix. To cite another example, micro-CT scans showed that the supposed quadratojugal identified by Estes (1970) and Henrici (2002) is hollow and, thus, it cannot be a quadratojugal. However, those same scans did not reveal any other hidden bones that could be the quadratojugal, so we are still left with the uncertainty about whether that bone was present in life. A quadratojugal is present in all other pelobatids (i.e. Pelobates and Eopelobates) but is lacking in scaphiopodids, so the absence of that bone in E. grandis could call into question its higher level affinities. Similarly, the suggestion from the micro-CT scans that the tibiale and fibulare may be coalesced distally, that there is a low dorsal crest on the iliac shaft, and that the centra are amphicoelous could also exclude the species from Eopelobates. Uncertainties over the presence/absence of a quadratojugal and the structure of the tibiale and fibulare cannot be resolved until additional specimens are found. For the time being, we concur with earlier workers (Zweifel 1956; Estes 1970; Henrici 2002) that E. grandis is best retained within Eopelobates.
Eopelobates hinschei (Kuhn, 1941)
Fig. 7

Photographs of Eopelobates hinschei (Kuhn, 1941), from the middle Eocene of Geiseltal, Germany. a, b Referrred specimen (GMH 6728; originally designated as holotype of Archaeopelobates eusculptus Kuhn, 1941), much of skeleton (a) and entire skull (b), both in dorsal aspect and whitened with unknown compound to enhance details and texture; line between posterior halves of left and right frontoparietals and extending ventroposteriorly onto tectum synoticum is a crack, not a suture; in closeup of skull (b) upper arrow points to processus lateralis superior on left side of frontoparietal complex and lower arrow points to processus lateralis inferior on right side of frontoparietal complex; both photographs are from the archive of Z.V. Špinar and neither included a scale or magnification. c, d Holotype (GMH 1312; originally designated as holotype of Halleobatrachus hinschei Kuhn, 1941), entire skeleton (c) and detail of sacral region (d), both in dorsal aspect; scale bar (c) is 10 mm; magnification for (d) about 4× larger than c. A. eusculptus, H. hinschei, and another five species and four genera of anurans named by Kuhn (1941) from Geiseltal were synonymized and assigned by Estes (1970) to Eopelobates as E. hinschei
Diagnosis (modified from Estes 1970; Wuttke 1988): Sculpture well pronounced on nasals (except for narrow strip along posterior margin), on posterior and posterolateral parts of frontoparietal complex, and on posterior two-thirds of maxillae (including processus frontalis), whereas anterior part of frontoparietal complex and of maxillae is covered by anteriorly oriented grooves that become shallower towards the anterior; nasals widely separated from one another (Fig. 7b; contra Estes 1970, fig. 12A); frontoparietals narrow posteriorly, about two times wider anteriorly, with distinct median convexity on the posterior margin and processus lateralis inferior rounded and shifted posteriorly from the level of processus lateralis superior; posterior part of lamella alaris of squamosal thin and rather bent ventrally (GMH 6753: Estes 1970, fig. 20b); tibiale and fibulare not coalesced; posterior margin of scapula deeply concave, anterior margin only slightly concave in its dorsolateral section (GMH 6692: Estes 1970, fig. 9e).
Material: Poorly preserved holotype skeleton (GMH 1312), originally described by Kuhn (1941; see also Estes 1970) as “Halleobatrachus hinschei” and 15 other skeletons (see list provided by Estes 1970, table 1), all from Geiseltal, Germany.
Stratigraphy and distibution: Lower Geiseltalian section (“Oberes Hauptmittel”), MP 13, middle Eocene (ca. 44 Ma), quarries Cecilie III and IV, Geiseltal near Halle, Germany (see Wuttke et al. 2012).
Description: Due to the poorly preserved nature of the available specimens, we refrain from providing a description here. Instead, see descriptive remarks by Kuhn (1941) and Estes (1970) and our photographs of two representative skeletons (Fig. 7).
Remarks: Kuhn (1941) named a number of anuran species and genera based on skeletons from the middle Eocene of Geiseltal, Germany. Estes (1970, p. 306, table 1) argued that of the new taxa named by Kuhn (1941), seven species within six genera pertained to a single species of Eopelobates, a genus that Kuhn (1941) had not identified at Geiseltal. Evidently on the basis of page priority (i.e. Kuhn 1941, p. 353), Estes (1970) choose Halleobatrachus hinschei Kuhn, 1941 as the senior subjective synonym for the Geiseltal Eopelobates species. An unfortunate consequence of that taxonomic reassignment was that the holotype of E. hinschei (Kuhn, 1941) is a poorly preserved specimen (GMH 1312: Fig. 7c, d) in which most of the diagnostic characters for the genus, including those listed by Estes (1970, p. 295) cannot be seen. Holotypes for some of Kuhn’s (1941) other species that were subsumed by Estes (1970) within E. hinschei are better preserved and more informative (e.g. Archaeopelobates eusculptus Kuhn, 1942; holotype GMH 6728: Fig. 7a, b). Estes (1970, p. 306) stated that similarities in proportions of the frontoparietals and hindlimbs suggested E. hinschei and the younger (late Oligocene–middle Miocene) E. bayeri were closely related, but that differences in median fusion of the frontoparietals, proportions of the nasals, and shapes of the squamosals supported recognising E. hinschei and E. bayeri as separate species. Although we agree that E. hinschei is distinct from E. bayeri, it is less clearly different from the paracontemporanous species E. wagneri (Weitzel, 1938) from the middle Eocene of Messel (see next account).
Eopelobates wagneri (Weitzel, 1938)
Diagnosis (modified from Wuttke 2012b): Sculpture developed on narrow posterior part of frontoparietal and along posterolateral margins of its frontal portion (Wuttke 2012b, pl. 5, fig.1), weak and irregular rugosity on orbital section of maxilla and on lamella alaris of squamosal; base of pars facialis of premaxilla positioned midway across horizontal portion of bone (Wuttke 2012b, pl. 3, fig. 1) and process is broader distally than at its base; nasals widely separated and their medial margins are parallel; sphenethmoid exposed in a large gap between nasals and frontoparietal; anterior end of frontoparietal fenestra is exposed (Wuttke 2012b, fig. 4a, c); frontoparietal narrow posteriorly (i.e. more than two times wider anteriorly) and processus lateralis inferior of frontoparietal short, rounded, and shifted far posteriorly from the level of processus lateralis superior; lamella alaris of squamosal slender anteriorly and tapered in a sharp point, markedly constricted in its posterior part (Wuttke 2012b, pl. 5, figs. 3, 4); posterior margin of scapula only slightly more concave than the anterior (Wuttke 2012b, pl. 8, figs. 2, 3); urostyle separated from sacral vertebra by a joint (Wuttke 2012b, pl. 6, fig 3); tibiale and fibulare not coalesced.
Material: Poorly preserved holotype (HLMD-Me 1286) and paratype (HLMD-Me 4359) skeletons described by Weitzel (1938), plus several dozen additional skeletons listed by Wuttke (2012b), all from Messel, Germany.
Stratigraphy and distribution: Messel Formation, MP 11, middle Eocene (ca. 47 Ma), Messel near Darmstadt, Germany (Wuttke 2012b).
Description: None needed, thanks to the detailed description and figures recently provided by Wuttke (2012b).
Remarks: Weitzel (1938) named this species as Propelodytes wagneri and assigned it to Pelodytidae. In his PhD dissertation, Wuttke (1988) transferred the species to Eopelobates and that generic assignment has been accepted since (see summaries by Sanchiz 1998; Wuttke 2012b). Utilising several dozen additional specimens from the type area in the middle Eocene Messel Pit, southwest Germany, Wuttke (2012b) provided a revised diagnosis and more detailed description of E. wagneri. As mentioned above, E. wagneri and E. hinschei are morphologically similar to one another and have similar temporal and geographic distributions (middle Eocene, Germany), with the former known only from Messel and the latter from Geiseltal. The possibility that these may be the same species deserves to be seriously considered; if so, the name E. wagneri has priority. For the time being, we retain these as separate species.
It is worth noting that Messel has also produced a second, more problematic pelobatid. Lutetiobatrachus gracilis (Wuttke in Sanchiz 1998; see Wuttke 2012a) can be assigned to Pelobatidae on the basis of the following features: frontoparietals fused posteriorly and their posterior margin is convex in dorsal outline, the latter of which suggests the presence of a posterior median element; sacral transverse processes broadly dilated; and ilium without a dorsal tubercle. Resolving the generic and specific affinities of L. gracilis is complicated by the fact that the taxon is known by just one juvenile skeleton. The juvenile status of that holotype skeleton is indicated by it having frontoparietals that lack dorsal sculpture and are separated anteriorly by a median suture, separate quadratojugals and quadrates, the anteriormost end of the urostyle showing signs of weak segmentation, and in lacking epiphyses on the humeri and radioulnae. A suite of features (frontoparietals broader anteriorly than posteriorly, triangular nasals, and femur shorter than tibiofibula) suggest possible relations to Eopelobates, but at present that possibility can neither be verified nor refuted.
Comments on fossil occurrences and evolutionary trends in Eopelobates and Pelobates
The geologically oldest records of Pelobatidae in Europe are from the earliest Eocene (MP 7) at Dormaal, Belgium (Duffaud 2000), at Le Quesnoy (formerly called Creil), Rians, and perhaps Meudon, all in France (Nel et al. 1999; Duffaud 2000; Rage and Roček 2003), and at Silveirinha, Portugal (Antunes and Russell 1981; Rage and Augé 2003). All are represented by disarticulated and fragmentary bones and, as suggested by slightly younger (latest early Eocene or MP 10) frontoparietals and other cranial bones from Prémontré, France (Duffaud 2000), at least some of those early Eocene specimens belong to Eopelobates. The geologically oldest articulated pelobatid skeletons are from the middle Eocene of Germany and belong to two named species of Eopelobates: E. wagneri from Messel and E. hinschei (slightly younger than E. wagneri) from Geiseltal (e.g. Estes 1970; Sanchiz 1998; Wuttke 2012b; this study).
The earliest occurrences of unequivocal pelobatids in North America are less certain. Two points are important when considering the North American record: (1) earlier reports viewed pelobatids in a broader sense (i.e. sensu Duellman and Trueb 1994) than we do here, and (2) all pre-Eocene reports of pelobatids on the continent (see summary of occurrences by Gardner and DeMar 2013, tables 1, 4–7) are based on isolated and usually incomplete bones that may be pelobatid- or pelobatoid-like in their general structure, but are not especially diagnostic for either group. These caveats apply particularly to reports founded entirely on isolated, incomplete ilia from the Upper Jurassic Morrison Formation, Wyoming, USA (Evans and Milner 1993) and from the Upper Cretaceous Judith River Formation (late Campanian), Montana, USA (Sahni 1972) and Ellisdale Formation (undifferentiated Campanian), New Jersey, USA (Denton and O’Neill 1998); none of those iliac specimens can reliably be assigned to Pelobatidae. Other reports of pre-Eocene “pelobatids” in North Amercia are founded on isolated ilia and skull bones attributed to Eopelobates or an Eopelobates-like frog from the following units in the western USA: Fruitland Formation (late Campanian), New Mexico (Armstrong-Ziegler 1980); Mesaverde Formation (middle to late Campanian), Wyoming (e.g. DeMar and Breithaupt 2008); Lance Formation (late Mastrichtian), Wyoming (e.g. Estes 1970; Estes and Sanchíz 1982; Gardner 2008); and the Bug Creek Anthills locality (mixed late Maastrichtian and earliest Paleocene) in the Hell Creek Formation, Montana (Estes and Sanchíz 1982; Gardner 2008). Estes (1964, pp. 57–60) was the first to tentatively report pelobatid-like bones from the North American Cretaceous, in his monograph on non-mammalian vertebrates from the Lance Formation. Later in his revision of Eopelobates, he (Estes 1970, pp. 315–316) identified those specimens as “?Eopelobates sp.” The practice of identifying isolated frog skull bones bearing reticulate or pit-and-ridge style sculpture from the North American Late Cretaceous as “?Eopelobates sp.”, “cf. Eopelobates sp.”, or similar has been widely followed since, if only because it is a convenient way for grouping and identifying isolated bones that bear some resemblance to those of unequivocal Tertiary age Eopelobates, rather than necessarily indicating membership within or a close relationship to that genus.
The best described and figured of the problematic Eopelobates-like fossils from the North American latest Cretaceous and earliest Paleocene are from the Lance Formation and from Bug Creek Anthills in the Hell Creek Formation. These include ilia (e.g. Estes and Sanchíz 1982, fig. 5A–D; Gardner 2008, fig. 13.3H, I) and cranial bones, mostly maxillae but also some nasals, frontoparietals, and squamosals (e.g. Gardner 2008, fig. 13.3A–G). Examination by one of us (Z.R.) of the maxillary specimens listed by Gardner (2008, p. 232) as “cf. Eopelobates sp.” indicates that at least two specimens (AMNH 25002: Gardner 2008, fig. 13.3A, B; AMNH 27205: unfigured) differ from unequivocal Tertiary age Eopelobates in having sculpture that is more pustular or vermicular (rather than pit-and-ridge style), in details of the inner surface, and in having a peculiar canal within the orbital margin. That canal enters the maxilla on the inner surface of the processus zygomaticomaxillaris and exits on the posterior portion of the processus frontalis; in the latter region, the groove remains well separated from the groove for the nasolacrimal duct. In those features, the two maxillary specimens are more similar to referred maxillae (e.g. MCZ 3626: Estes 1969, fig. 2c, d; UALVP 40156: Gardner 2008, fig. 13.1N, O) of the incertae sedis anuran Scotiophryne pustulosa Estes, 1969 from the latest Cretaceous and early Paleocene of North America, although they lack the tightly packed, almost bead-like pustular sculpture that is characteristic of Scotiophryne. The incomplete frontoparietal (UALVP 40164: Gardner 2008, fig. 13.3F, G) listed by Gardner (2008, p. 232) for “cf. Eopelobates sp.” preserves most of its lateral margin, but its frontoparietal incrassation, which is a bony patch on the inner (ventral) surface of the bone that provides information on the shape of fenestrae in the roof of the braincase and has some taxonomically informative variability, is indistinct. That differs from the condition in an uncatalogued frontoparietal from the latest early Eocene (MP 10) of Prémontré, France, that preserves a prominent and pelobatid-like incrassation suggesting only one large fenestra that is not divided by taeniae (Duffaud 2000, fig. 105). The frontoparietal from Prémontré resembles that of E. hinschei (middle Eocene, Germany), which suggests that species may have occurred earlier in the Eocene; unfortunately, the Prémontré frontoparietal is now lost (J.-C. Rage, personal communication 2013). Returning to the problematic North American latest Cretaceous and Paleocene specimens, all reported Eopelobates-like ilia (e.g. Gardner 2008, fig. 13.3H, I) consistently display a generalised pelobatid-like morphology (e.g. no dorsal tubercle or dorsal crest, weakly developed oblique groove crossing the dorsal margin onto the medial surface of the bone), but those features are not particularly informative because they are widespread among Mesozoic anuran ilia from North America (see Roček et al. 2010), including Scotiophryne and the earliest (Early Jurassic) anuran Prosalirus. Other reports of Eopelobates or Eopelobates-like frogs from the North American Late Cretaceous (e.g. Armstrong-Ziegler 1980; DeMar and Breithaupt 2008) deserve to be treated with similar caution.
At present, the only unequivocal records for Eopelobates and for Pelobatidae (in the sense that we view that family; i.e. Eopelobates and Pelobates) in North America are the three specimens documented in our paper, specifically the holotype and paratype skeletons of E. deani from the early middle Eocene of Wyoming, USA, and the holotype skeleton of E. grandis from the latest Eocene of South Dakota, USA. For the sake of completeness, it should also be noted that there are two unverified reports of Eopelobates-like frogs from other Eocene age formations in the western USA. The first is a sacral vertebra from the Bridger Formation (early middle Eocene or Bridgerian NALMA), Wyoming, that was reported by Hecht (1959, p. 131, pl. 57, figs 6–9) and favorably compared by him to Eopelobates. Estes (1970, p. 308) accepted that generic identification. Although none of us has personally examined the relevant specimen (AMNH 3832), judging from the published photographs there does not seem to be any compelling osteological basis for that generic assignment. The second report is contained in a faunal list (Golz and Lillegraven 1977, table 1) in which “cf. Eopelobates sp.” was reported from the Mission Valley Formation (late Eocene or Uintan NALMA) of southern California. The relevant specimens have neither been described nor have we seen them, so their identification remains uncertain.
The Eopelobates deani occurrences are approximately contemporaneous with the earliest records of the genus in Europe. Assuming there is not a lengthy but undiscovered pre-Eocene record for Eopelobates in North America or Europe and taking into account pre-Eocene palaeogeographic reconstructions for those continents, it seems unlikely that Eopelobates spread from North America to Europe, or vice versa, before the late Paleocene. The only terrestrial connection then available between those two continents was the high latitude North Atlantic land bridge (the so-called DeGeer route or passage) that extended across present day Greenland during the late Paleocene–early Eocene (West and Dawson 1978; Beard and Dawson 1999; Smith et al. 2006; Solé and Smith 2013). That route could have been used not only by endothermic mammals and non-flying birds but, as evidenced by fossils of insects, salamanders, pond turtles, squamates, and alligators (Estes and Hutchison 1980; Buffeteaut 1997; Archibald and Makarkin 2006; Petrulevičius et al. 2007; Archibald et al. 2011; Gamble et al. 2011; Eberle and Greenwood 2012; Sullivan et al. 2012), also by ectothermic animals thanks to extended intervals of warmer annual mean temperatures, such as occurred during the Paleocene-Eocene Thermal Maximum that lasted about 100,000 years. Various lines of evidence gathered from early–middle Eocene age localities that today are well above the Arctic Circle suggest milder temperates, with winter values at or just above freezing and summer values of 20 °C (or higher), and high precipitation in that region during the Eocene (Eberle and Greenwood 2012). This high latitude route has previously been invoked to explain the palaeobiogeographical histories of certain salamander families (e.g. Naylor 1981; Milner 1983; Duellman and Trueb 1994) and it seems no less reasonable for frogs such as Eopelobates.
A peculiar anomaly in the reported distribution of Eopelobates is an ilium from the early Eocene (Ypresian) Vastan Lignite Mine in Gujarat, India, that Folie et al. (2013) recently identified as belonging to an indeterminate species of Eopelobates. As already noted by Roček (2013, p. 403) that generic identification is suspect for two reasons. First, ilia have not previously been used to diagnose Eopelobates and it is not clear that the features listed by Folie et al. (2013) are reliably diagnostic. Second, the presence of Eopelobates in India during the early Eocene, roughly simultaneous with the earliest occurrences of Eopelobates in Europe and North America, is at odds with well accepted palaeogeographical reconstructions in which India and Europe were separated during and prior to the early Eocene by the Turgai Strait. We note, however, that intermittent land routes across the Turgai Strait during the Paleocene and Eocene have been suggested as routes for mammals, turtles, and snakes to move between Asia and Europe and that non-marine vertebrate assemblages on those continents during the Eocene are now looking to be less dissimilar than previously thought (e.g. Godinot and Lapparent de Broin 2003; Hooker and Dashzeveg 2003; Smith et al. 2006, 2007; Rage et al. 2008; Rana et al. 2008). Even if such a route was available for Eopelobates to move from Europe into India, the ilium from Vastan Lignite Mine is insufficient for establishing Eopelobates at that locality. In our opinion, the only reliable materials for making such an identification are cranial elements, ideally associated or articulated.
Eopelobates has not been recorded in North America in post Eocene times, but in Europe it persisted until the Pliocene, as evidenced by its last records from Poland, Slovakia, and Ukraine (see above). The geologically oldest occurences of Pelobates fossils (reported as cf. Pelobates) are disarticulated bones from the middle Oligocene (MP 22–23) localities in France (de Bonis et al. 1973). Thus, both genera overlapped in Europe from the middle Oligocene until the latest Pliocene, which is an interval of about 25 million years. During that time, characteristic features of Eopelobates were gradually, but at different rates, modified into those of Pelobates. Among the first features to be transformed were that the frontoparietal established a postorbital bridge with the squamosal and that the anterolateral margin of the nasals changed from straight to sinusoid. Both of those features are seen in articulated skeletons of Pelobates decheni from the late Oligocene (Böhme et al. 1982, fig. 2; Roček and Wuttke 2010, fig. 7c). The concavity in the anterior section of the anterolateral margin of the nasal in Pelobates implies possible changes in this area, such as extension of the external naris. In the postcranial skeleton, an early change is a shortening of the urostyle. That feature also is seen in Pelobates decheni (Roček and Wuttke 2010, fig. 6a, b, h).
On the other hand, other features were retarded. Sculpture in the earliest Pelobates (see P. decheni Fig. 8b, c) resembled that of Eopelobates (Fig. 8a) in being of the pit-and-ridge style and in having low relief. A transitional pattern that combines pit-and-ridge and pustular sculpture (Fig. 8d) is seen only during the Miocene; see also P. fahlbuschi from the early Miocene of Sandelzhausen, Germany (Böhme 2010, fig. 5Y) and P. sanchizi from the middle Miocene of Hungary (Böhme 2010, fig. 5X; Venczel 2004, text-fig. 6B). Exclusively pustular sculpture seems to have appeared only in the Pliocene (Hodrová 1985) and continues to the present (Fig. 8e). Within Pelobatidae a bony spade also may have evolved rather late, although that is not certain because recognising that feature requires articulated postcranial material and that is sorely lacking for most fossil species of Pelobates. A bony spade has never been found in Eopelobates or in the only fossil Pelobates species (P. decheni) for which articulated postcranial material is available. All extant species of Pelobates have a bony spade.

Representative pelobatid frontoparietal complexes, showing hypothetical transformation series in cranial sculpture. a–c Inferred primitive pit-and-ridge pattern, typical of Eocene–Pliocene Eopelobates and Oligocene–Miocene Pelobates: a Eopelobates hinschei (Kuhn, 1941), middle Eocene, Geiseltal, Germany (GMH 6728); b Pelobates cf. P. decheni Troschel, 1861, late Oligocene, Enspel, Germany (NHMM PW1995/5802a-LS); c Pelobates cf. P. decheni Troschel, 1861, late Oligocene, Enspel, Germany (NHMM PW1997/5040). d Inferred intermediate pattern that combines pit-and-ridge with pustular sculpture, seen only in some Miocene Pelobates: Pelobates sp., late Miocene (middle Sarmatian, MN 9a), Gritsev, Ukraine (IZANK 3338). e Inferred derived pustular sculpture, seen in Pliocene–Recent Pelobates: P. varaldii ♀, Recent, Mamora, Morocco (DPFNSP 6328a). Not to scale
Fossoriality in fossil Pelobates potentially can be inferred from a suite of other skeletal features that tend to characterise burrowing anurans, but for various reasons none of those helps establish when fossoriality appeared in the genus. Because the urostyle is a structure from which muscles important for burrowing take their origin (Přikryl et al. 2009), fusion of this element with the sacral vertebra is typical of frogs that have lost the ability to jump in favor of hindlimb burrowing. The urostyle-sacral joint is fused in adults of all Recent species of Pelobates, whereas it is a movable joint in skeletons of the late Oligocene P. decheni and, judging by complementary surfaces on isolated sacral vertebrae and urostyles, also during the middle Miocene (e.g. P. sanchizi: Venczel 2004, text-fig. 7E, F, L, M). Another feature often associated with burrowing is fusion of the tibiale and fibulare. Such fusion occurs in all extant species of Pelobates, but it is unknown for fossil species of the genus (including P. decheni) and, with the possible exception of Eopelobates grandis (see above account for that species), it is also unknown for Eopelobates. It is worth noting that fusion of the tibiale and fibulare does not appear to be a prerequisite to burrowing. For example, the pelobatoid Macropelobates linquensis from the Miocene of China is a large-bodied species that bears well-developed bony spades and on that basis likely was a burrower, yet its tibiale and fibulare are unfused (Roček et al. 2011). Extant species of Pelobates also have the tibiofibula shorter than the femur and F + TF shorter than SVL; however, these ratios do not necessarily indicate fossoriality, because similar ratios are seen in non-fossorial bufonids. It should also be noted that the prehallux, formed by two elements, is a characteristic feature of anuran hindlimbs, and is not restricted to burrowers. In fossorial anurans, including Recent Pelobates, the more distal element of the prehallux is enlarged and elaborated to support the keratinized spade. Although prehallical elements are rarely preserved in fossils, Špinar found them in non-fossorial Eopelobates bayeri (Špinar 1972, text-fig. 92). In summary, existing fossil evidence does not allow us to estimate when burrowing evolved in pelobatids. It may not have occurred until post Pliocene times.
Notes
Some of these occurrences are mentioned, together with Eopelobates, in the discussion portion of this paper. Fossil occurrences listed here are limited to reports of indeterminate pelobatids (in the broader sense used by earlier workers) and may not be assignable to Pelobatidae in the sense that we view that family here; occurrences identified to genus or species are presented in the corresponding genus accounts.
Only records determined as Pelobates are included.
References
Antunes MT, Russell DE (1981) Le gisement de Silveirinha (Bas Mondego, Portugal): La plus ancienne faune de vertébrés éocènes connue en Europe. CR Acad Sci Paris II 293:1099–1102
Archibald SB, Johnson KR, Mathewes RW, Greenwood DR (2011) Intercontinental dispersal of giant thermophilic ants across the Arctic during early Eocene hyperthermals. Proc R Soc Lond B 278:3679–3686. doi:10.1098/rspb.2011.0729
Archibald SB, Makarkin VN (2006) Tertiary giant lacewings (Neuroptera: Polystoechotidae) revision and description of new taxa from western North America and Denmark. J Syst Palaeontol 4:119–155. doi:10.1017/S1477201906001817
Armstrong-Ziegler JG (1980) Amphibia and Reptilia from the Campanian of New Mexico. Fieldiana Geol N Ser 4:1–39
Bailon S, Bour R, Rage J-C (1988) Quand les especes de l’herpetofaune francaise sont-elles apparues? Bull Soc Herp Fr 45:1–8
Beard KC, Dawson MR (1999) Intercontinental dispersal of Holarctic land mammals near the Paleocene/Eocene boundary: paleogeographic, paleoclimatic and biostratigraphic implications. Bull Soc Géol Fr 170:697–706
Bellon H, Bůžek C, Gaudant J, Kvaček Z, Walther H (1998) The České Středohoří magmatic complex in Northern Bohemia 40K-40Ar ages for volcanism and biostratigraphy of the Cenozoic freshwater formations. Newsl Stratigr 36:77–103
Böhme M (2008) Ectothermic vertebrates (Teleostei, Allocaudata, Urodela, Anura, Testudines, Choristodera, Crocodylia, Squamata) from the Upper Oligocene of Oberleichtersbach (Northern Bavaria, Germany). Cour Forsch-Inst Senckenberg 260:161–183
Böhme M (2010) Ectothermic vertebrates (Actinopterygii, Allocaudata, Urodela, Anura, Crocodylia, Squamata) from the Miocene of Sandelzhausen (Germany, Bavaria) and their implications for environment reconstruction and palaeoclimate. Paläontol Z 84:3–41. doi:10.1007/s12542-010-0050-4
Böhme M, Vasilyan D (2014) Ectothermic vertebrates from the late Middle Miocene of Gratkorn (Austria, Styria). In: Böhme M, Gross M, Prieto J (eds) The Sarmatian vertebrate locality Gratkorn, Styrian Basin. Palaeobio Palaeoenv 94:21–40
Böhme W, Roček Z, Špinar ZV (1982) On Pelobates decheni Troschel, 1861, and Zaphrissa eurypelis Cope, 1966 (Amphibia: Salientia: Pelobatidae) from the Early Miocene of Rott near Bonn, West Germany. J Vertebr Paleontol 2:1–7
Bolkay SJ (1919) Osnove uporedne osteologije anurskih batrahija. Glas Zemaljskog Muz Bosni Hercegovini 31(3):277–356 (available online from http://www.scribd.com/doc/76034384/glasnik-zemaljskog-muzeja-1919-god-31-knj-3)
Bonaparte CLJL (1850) Conspectus systematum herpetologiae et amphibiologiae. Editio altera reformata. Brill, Leiden [Lugdini Batavorum]
Borsuk-Białynicka M (1978) Eopelobates leptocolaptus sp. n. – the first Upper Cretaceous pelobatid frog from Asia. Palaeontol Pol 38:57–63
Boulenger GA (1910) Les Batraciens, et principalement ceux d’Europe. Doin, Paris
Bradley WH (1931) Origin and microfossils of the oil shale of the Green River Formation of Colorado and Utah. US Geol Surv Prof Pap 168:1–58
Buffeteaut E (1997) New remains of the giant bird Gastornis from the Upper Paleocene of the eastern Paris Basin and the relationships between Gastornis and Diatryma. N Jb Geol Paläont, Mh 1997:179–190
Chkhikvadze VM (1981) [Survey of the evidence on the fossil remains of the Amphibia and Reptilia from the Neogene deposits of the north Black Sea shore region]. In: Problems in Herpetology. Proceedings of the 5th All-Union Herpetological Conference, pp. 151–152 Nauka, Leningrad. (In Russian)
Chkhikvadze VM (1984) Survey of the fossil urodelan and anuran amphibians in the USSR. Proc Acad Sci Georgian SSR Ser Biol 10:5–13 (In Russian, with English summary)
Chkhikvadze VM (1985) Preliminary results of studies on Tertiary amphibians and squamate reptiles of the Zaisan Basin. In: Darevsky IS (ed) Problems in Herpetology. Proceedings of the 6th All-Union Herpetological Conference. Leningrad, Nauka, pp 234–235 (In Russian)
Cope ED (1865) Sketch of the primary groups of Batrachia Salientia. Nat Hist Rev 1865:97–120
Cope ED (1866) On the structure and distribution of the genera of the arciferous Anura. J Acad Nat Sci Phila 6:67–112
Crochet JY (1972) Les vertébrés de l’Oligocene supérieur du Pech du Fraysse, poche a phosphate du Quercy (commune de Saint-Projet, Tarn-et-Garonne). CR Somm Seanc Soc Geol Fr 6:316–317
Crochet JY, Hartenberger J-L, Rage J-C, Remy JA, Sige B, Sudre J, Vianey-Liaud M (1981) Les nouvelles faunes de vertebres anterieurs a la “Grande Coupure” decouverts dans les phosphorites du Quercy. Bull Mus Natl Hist Nat Paris, ser.4, 3, sect.C(3):245–266
Cuvier G (1829) Le Regne Animal distribue d’apres son Organisation, pour servir de Base a l’Histoire naturelle des Animaux et d’Instruction a l’Anatomie comparee. Nouvelle Edition, revue et augmentee par P.A. Latreille. Vol.2. Deterville, Paris
de Bonis L, Crochet JY, Rage J-C, Sige B, Vianey-Liaud M (1973) Nouvelles faunes de vertébrés oligocenes des phosphorites du Quercy. Bull Mus Natl Hist Nat Paris 174(3):105–113
Delwig W (1928) Eine neue Art der Gattung Pelobates Wagl. aus dem zentralen Transkaukasus. Zool Anz 75:24–31
DeMar DG Jr, Breithaupt BH (2008) Terrestrial and aquatic vertebrate paleocommunities of the Mesaverde Formation (Upper Cretaceous, Campanian) of the Wind River and Bighorn basins, Wyoming. In: Sankey JT, Baszio S (eds) Vertebrate microfossil assemblages: Their role in paleoecology and paleobiogeography. Indiana University Press, Bloomington, pp 78–103
Denton RK, O’Neill RC (1998) Parrisia neocesariensis, a new batrachosauroidid salamander and other amphibians from the Campanian of eastern North America. J Vertebr Paleontol 18:484–494
Duellman WE, Trueb L (1994) Biology of the amphibians. Johns Hopkins University Press, Baltimore
Duffaud S (2000) Les faunes d’amphibiens du Crétacé supérieur à l’Oligocène inférieur en Europe: paléodiversité, évolution, mise en place. PhD thesis, Muséum national d’Histoire Naturelle, Paris
Eberle JJ, Greenwood DR (2012) Life at the top of the greenhouse Eocene world – A review of the Eocene flora and vertebrate fauna from Canada’s High Arctic. Geol Soc Am Bull 124:3–23
Estes R (1964) Fossil vertebrates from the Late Cretaceous Lance Formation, eastern Wyoming. Univ Calif Publ Geol Sci 49:1–180 + 5 plates
Estes R (1969) A new fossil discoglossid frog from Montana and Wyoming. Breviora 328:1–7
Estes R (1970) New fossil pelobatid frogs and a review of the genus Eopelobates. Bull Mus Comp Zool 139:293–339
Estes R, Hutchison JH (1980) Eocene lower vertebrates from Ellesmere Island, Canadian Arctic Archipelago. Palaeogeogr Palaeoclimatol Palaeoecol 30:325–347
Estes R, Sanchíz B (1982) New discoglossid and palaeobatrachid frogs from the Late Cretaceous of Wyoming and Montana, and a review of other frogs from the Lance and Hell Creek Formations. J Vertebr Paleontol 2:9–20
Evans SE, Milner AR (1993) Frogs and salamanders from the Upper Jurassic Morrison Formation (Quarry Nine, Como Bluff) of North America. J Vertebr Paleontol 13:24–30
Fabrezi M, Alberch P (1996) The carpal elements of anurans. Herpetologica 52:188–204
Folie A, Rana RR, Rose KD, Sahni A, Kumar K, Singh L, Smith T (2013) Early Eocene frogs from Vastan Lignite Mine, Gujarat, India. Acta Palaeontol Pol 58:511–524
Frost DR (2014) Amphibian Species of the World: an Online Reference. Version 6.0 (30 January 2014). Electronic Database accessible at http://research.amnh.org/herpetology/amphibia/index.html. American Museum of Natural History, New York, USA
Frost DR, Grant T, Faivovich J, Bain RH, Haas A, Haddad CFB, De Sá RO, Channing A, Wilkinson M, Donnellan SC, Raxworthy CJ, Campbell JA, Blotto BL, Moler P, Drewes RC, Nussbaum RA, Lynch JD, Green DM, Wheeler WC (2006) The amphibian tree of life. Am Mus Nat Hist Bull 297:1–370
Gamble T, Bauer AM, Colli GR, Greenbaum E, Jackman TR, Vitt LJ, Simons AM (2011) Coming to America: multiple origins of New World geckos. J Evol Biol 24:231–244. doi:10.1111/j.1420-9101.2010.02184.x
García-París M, Buchholz DR, Parra-Olea G (2003) Phylogenetic relationships of Pelobatoidea re-examined using mtDNA. Mol Phylog Evol 28:12–23
Gardner JD (1999) Comments on amphibians from the Green River Formation, with a description of a fossil tadpole. In: Gillette DD (ed) Vertebrate paleontology in Utah. Utah Geol Surv Misc Publ 99–1:455–461
Gardner JD (2008) New information on frogs (Lissamphibia: Anura) from the Lance Formation (late Maastrichtian) and Bug Creek Anthills (late Maastrichtian and early Paleocene), Hell Creek Formation, USA. In: Sankey JT, Baszio S (eds) Vertebrate microfossil assemblages: their role in paleoecology and paleobiogeography. Indiana University Press, Bloomington, pp 219–249
Gardner JD, DeMar DG Jr. (2013) Mesozoic and Palaeocene lissamphibian assemblages of North America: a comprehensive review. In: Gardner JD, Nydam RL (eds) Mesozoic and Cenozoic lissamphibian and squamate assemblages of Laurasia. Palaeobio Palaeoenv 93:459–516
Gaudant J (1985) Mise au point sur les Vertébrés inférieurs de l’Oligocene de Sieblos (Hesse, Allemagne). CR Acad Sci Paris 300(2):185–188
Gayet H, Rage J-C, Rana RS (1984) Nouvelles ichthyofaune et herpetofaune de Gitti Khadan, le plus ancien gisement connu du Deccan (Cretace, Paleocene) a microvertebres. Implications paleogeographiques. Mem Soc Geol Fr NS 147:55–65
Godinot M, de Lapparent de Broin F (2003) Arguments for a mammalian and reptilian dispersal from Asia to Europe during the Paleocene-Eocene boundary interval. In: Reumer JWF, Wessels W (eds) Distribution and migration of Tertiary mammals in Eurasia. A volume in honour of Hans de Bruijn. Deinsea, 10:255–275
Golz DJ, Lillegraven J (1977) Summary of known occurrences of terrestrial vertebrates from Eocene strata of southern California. Univ Wyo Contrib Geol 15:43–65
Gosner KL (1960) A simplified table for staging anuran embryos and larvae with notes on identification. Herpetologica 16:183–190
Grande L (1984) Paleontology of the Green River Formation with a review of the fish fauna. Geol Surv Wyo Bull 63:1–333
Grande L (2013) The lost world of Fossil Lake: Snapshots from deep time. University of Chicago Press, Chicago
Gubin YM (1996) The first find of pelobatids (Anura) in the Paleogene of Mongolia. Paleont Zh 1996:73–76 (in Russian, with English summary)
Haas A (2003) Phylogeny of frogs as inferred from primarily larval characters (Amphibia: Anura). Cladistics 19:23–90
Hecht MK (1959) Amphibian and reptiles. In: McGrew PO, Berman JE, Hecht MK, Hummel JM, Simpson GG, Wood AE (authors) The geology and paleontology of the Elk Mountain and Tabernacle Butte area, Wyoming. Am Mus Nat Hist Bull 117:130–146
Hecht M, Hoffstetter R (1962) Note preliminaire sur les amphibiens et les squamates du Landenien superieur et du Tongrien de Belgique. Bull Inst R Sci Nat Belg 38:1–30
Henrici A (1994) Tephrodytes brassicarvalis, new genus and species (Anura: Pelodytidae), from the Arikareean Cabbage Patch beds of Montana, USA, and pelodytid-pelobatid relationships. Ann Carnegie Mus 63:155–183
Henrici A (2000) Reassessment of the North American pelobatid anuran Eopelobates guthriei. Ann Carnegie Mus 69:145–156
Henrici A (2002) Redescription of Eopelobates grandis, a late Eocene anuran from the Chadron Formation of South Dakota. Ann Carnegie Mus 71:241–259
Henrici AC, Haynes SR (2006) Elkobatrachus brocki, a new pelobatid (Amphibia: Anura) from the Eocene Elko Formation of Nevada. Ann Carnegie Mus 75:11–35. doi:10.2992/0097-4463(2006)75[11:EBANPA]2.0.CO;2
Henrici AC, Báez AM, Grande L (2013) Aerugoamnis paulus, new genus and species (Anura: Anomocoela): first reported anuran from the early Eocene (Wasatchian) Fossil Butte Member of the Green River Formation, Wyoming. Ann Carnegie Mus 81:295–309
Hodrová M (1981) Plio-Pleistocene frog fauna from Hajnáčka and Ivanovce, Czechoslovakia. Věst ÚÚG 56:215–224
Hodrová M (1985) Amphibia of Pliocene and Pleistocene Včeláre localities (Slovakia). Čas Min Geol 30:145–161
Hodrová M (1987) Lower Miocene frogs from the Dolnice locality in the Cheb Basin (Czechoslovakia). Acta Univ Carol Geol 1987: 97–115
Hodrová M (1988) Miocene frog fauna from the locality Devínska Nová Ves – Bonanza. Věst ÚÚG 305–310
Holman JA (2003) Fossil frogs and toads of North America. Indiana University Press, Bloomington
Hooker JJ, Dashzeveg D (2003) Evidence for direct mammalian faunal interchange between Europe and Asia near the Paleocene-Eocene boundary. In: Wing SL, Gingerich PD, Schmitz B and Thomas E (eds) Causes and consequences of globally warm climates in the early Paleogene. Geol Soc Am Spec Pap 369:479–500
Ivanov M (2008) Early Miocene Amphibians (Caudata, Salientia) from the Mokrá-Western Quarry (Czech Republic) with comments on the evolution of Early Miocene amphibian assemblages in Central Europe. Geobios 41:465–492
Khosatzky LI (1985) [A new species of spade-foot toads from the Pliocene of Moldavia]. In: Fauna and flora of the Late Cenozoic of Moldavia, pp. 59–72. Shtiintsa, Kishinev (In Russian)
von Koenigswald W, Martin T, Mörs T, Pfretzschner HU (1992) Die oberoligozäne Wirbeltierfauna von Rott bei Hennef am Siebengebirge. Synonymien und Literatur 1828-1991. Decheniana 145:312–340
Krishtalka L, West RM, Black CC, Dawson MR, Flynn JJ, Turnbull WD, Sucky RK, McKenna MC, Bown TM, Golz DJ, Lillegraven JA (1987) Eocene (Wasatchian through Duchesnean) biochronology of North America. In: Woodburne MO (ed) Cenozoic mammals of North America: Geochronology and biostratigraphy. University of California Press, Berkeley, pp 77–117
Kuhn O (1941) Die eozänen Anura aus dem Geiseltale nebst einer Übersicht über die fossile Gattungen. Nov Acta Leopold NF 10:345–376
Laurenti JN (1768) Specimen medicum, exhibens synopsin reptilium emendatam cum experimentis circa Venena et antidota reptilium Austriacorum. Thom, Vienna
Lebedkina NS (2004) Evolution of amphibian skull. Pensoft, Sofia-Moscow
Maglia AM (1998) Phylogenetic relationships of extant pelobatoid frogs (Anura: Pelobatoidea): Evidence from adult morphology. Univ Kansas Nat Hist Mus Sci Pap 10:1–19
Mason GM (2011) Fossil insect larvae provide clues for interpretation of the depositional environment of the Green River Formation in early summer … give or take a few million years. Proceedings of 31st Oil Shale Symposium, Colorado School of Mines, Golden, Colorado, USA, 17–19 October 2011. Available at: http://www.costar-mines.org/oss/31/F-paper-sec/18-09_Mason_Glenn_Paper_Final.pdf
Maus M, Wuttke M (2002) Comparative anatomical and taphonomical examination of the larvae of Pelobates decheni Troschel 1861 and Eopelobates anthracinus Parker 1929 (Anura: Pelobatidae) found at the Upper Oligocene sites at Enspel (Westerwald/Germany) and Rott (Seibengebirge/Germany). Cour Forsch-Inst Senckenberg 237:129–138
Maus M, Wuttke M (2004) The ontogenetic development of Pelobates cf. decheni tadpoles from the Upper Oligocene of Enspel (Westerwald/Germany). N Jb Geol Paläont, Abh 232:215–230
Mertens R (1923) Beiträge zur Kenntnis der Gattung Pelobates Wagler. Senck biol 5:118–128
Michahelles C (1830) Neue südeuropäische Amphibien. Isis (Oken) 1830:806–809
Milner AC, Milner AR, Estes R (1982) Amphibians and Squamates from the Upper Eocene of Hordle Cliff, Hampshire - a preliminary report. Tertiary Res 4:149–154
Milner AR (1983) The biogeography of salamanders in the Mesozoic and Early Caenozoic: a cladistic-vicariance model. In: Sims RW, Price JH, Whalley PES (eds.), Evolution, time and space: the emergence of the biosphere. Syst Assoc Spec Vol 23. Academic Press, London, pp. 431–468
Młynarski M (1961) Plazy (Amphibia) z pliocenu Polski. Acta Paleontol Pol 6:261–282
Młynarski M (1962) Notes on the amphibian and reptilian fauna of the Polish Pliocene and early Pleistocene. Acta Zool Cracov 7:177–194
Młynarski M (1977) New notes on the amphibian and reptilian fauna of the Polish Pliocene and Pleistocene. Acta Zool Cracov 22:13–36
Młynarski M, Szyndlar Z (1989) Plazy i gady – Amphibia et reptilia. Folia Quaternaria Krakow 59–60:69–88
Młynarski M, Szyndlar Z, Estes R, Sanchíz B (1984) Amphibians and reptiles from the Pliocene locality of Weze II near Działoszyn (Poland). Acta Paleontol Pol 29:209–226
Müller J (1832) Über die natürliche Einteilung der Amphibien. Isis (Oken) 5:504–539
Naylor BG (1981) Cryptobranchid salamanders from the Paleocene and Miocene of Saskatchewan. Copeia 1981(1):76–86
Nel A, de Plöeg G, Dejax J, Dutheil D, de Frasceschi D, Gheerbrant E, Godinot M, Hervet S, Menier J-J, Augé M, Bignot G, Cavagnetto C, Duffaud S, Gaudant J, Hua S, Jossang A, de Lapparent de Broin F, Pozzi J-P, Paicheler J-C, Beuchet F, Rage J-C (1999) Un gisement sparnacien exceptionnel à plantes, arthropodes et vertébrés (Éocène basal, MP7): Le Quesnoy (Oise, France). CR Acad Sci Paris Sci Terre Planèt 329:65–72
Nessov LA (1981) [Cretaceous salamanders and frogs of Kizylkum Desert]. Trudy Zoologicheskogo Instituta. Akad Nauk SSSR 101:57–88 (In Russian)
Nieuwkoop PD, Faber J (1967) Normal table of Xenopus laevis (Daudin). North Holland, Amsterdam
Noble GK (1924) A new spadefoot toad from the Oligocene of Mongolia with a summary of the evolution of the Pelobatidae. Am Mus Novit 132:1–15
Noble GK (1931) The biology of the Amphibia. McGraw Hill, New York
Paicheler J-C, de Broin F, Gaudant J, Mourier-Chauvre C, Rage J-C, Vergnaud-Grazzini C (1978) Le basin lacustre miocene de Bes-Konak (Anatolie-Turquie): geologie et introduction a la paleontologie des vertébrés. Geobios 11:43–65
Parker HW (1929) Two fossil frogs of the lower Miocene of Europe. Ann Mag Nat Hist 10:270–281
Pasteur G (1958) Sur les tendences evolutives et la phylogenie des Pelobates (Batraciens anoures) actuels. CR Acad Sci Paris 247:1037–1039
Paunović M (1984) Fische, Amphibien und Reptilien aus oberpleistozänen Ablagerungen von Šandalja bei Pula (Istrien, Kroatien). Palaeont Jugosl 31:5–44
Petrulevičius JF, Nel A, Rust J, Bechly G, Kohls D (2007) New Paleogene Epallagidae (Insecta: Odonata) recorded in North America and Europe. Biogeographic implications. Alavesia 1:15–25
Prasad GVR, Sahni A (1987) Coastal-plain microvertebrate assemblage from the terminal Cretaceous of Asifabad, peninsular India. J Paleo Soc India 32:5–19
Prasad GVR, Sahni A (2009) Late Cretaceous continental vertebrate fossil record from India: Palaeobiogeographical insights. Bull Soc Geol Fr 180:369–381
Přikryl T, Aerts P, Havelková P, Herrel A, Roček Z (2009) Pelvic and thigh musculature in frogs (Anura) and origin of anuran jumping locomotion. J Anat 214:100–139
Rage J-C (1972) Les Amphibiens et les Reptiles des couches paleochretiennes de la grotte de l’Hortus. Etudes Quatern Mem 1:299–301
Rage J-C (1988) Le gisement du Bretou (phosphorites du Quercy, Tarn-et-Garonne, France) et sa faune de vertébrés de l’Eocene superieur. I. Amphibiens et reptiles. Palaeontographica Abt A 205:3–27
Rage JC, Augé M (2003) Amphibians and squamate reptiles from the lower Eocene of Silveirinha (Portugal). Ciências Terra (UNL) 15:103–116, http://hdl.handle.net/10362/4731
Rage J-C, Ford RLE (1980) Amphibians and squamates from the upper Eocene of the Isle of Wight. Tertiary Res 3:47–60
Rage J-C, Hossini S (2000) Les Amphibiens du Miocène moyen de Sansan. In: Ginsburg L (ed) La faune miocène de Sansan et son environment. Mém Mus Nat. Hist Nat 183:177–217
Rage J-C, Roček Z (2003) Evolution of anuran assemblages in the Tertiary and Quaternary of Europe, in the context of palaeoclimate and palaeogeography. Amphibia-Reptilia 24:133–167
Rage J-C, Sen S (1976) Les amphibiens et les reptiles du Pliocene superieur de Çalta (Turquie). Geol Mediter 3:127–134
Rage J-C, Vergnaud-Grazzini C (1978) La poche a phosphate de Ste-Neboule (Lot) et sa faune de vertebres du Ludien supérieur. 2. Amphibiens. Note preliminaire. Palaeovertebrata Montpellier 8(2–4):175–179
Rage JC, Folie A, Rana RS, Singh H, Rose KD, Smith T (2008) A diverse snake fauna from the early Eocene of Vastan Lignite Mine, Gujarat, India. Acta Palaeontol Pol 53:391–403
Rana RS, Kumar K, Escarguel G, Sahni A, Rose KD, Smith T, Singh H, Singh L (2008) An ailuravine rodent from the lower Eocene Cambay Formation at Vastan, western India, and its palaeobiogeographic implications. Acta Palaeontol Pol 53:1–14
Ratnikov VY (1988) Upper Quaternary herpetofaunas of the Belgorodskaya Region. Paleontol Zh 1988:119–122
Roček Z (1981) Cranial anatomy of frogs of the family Pelobatidae Stannius, 1856, with outlines of their phylogeny and systematics. Acta Univ Carol Biol 1980:1–164
Roček Z (1982) Macropelobates osborni Noble, 1924—Redescription and reassignment. Acta Univ Carol Geol 1982:421–438
Roček Z (1995) A new specimen of Eopelobates (Anura: Pelobatidae) from the Tertiary near Bonn (Germany) and the problem of E. anthracinus-E. bayeri relations. Paläontol Z 69:283–287
Roček Z (2005) Late Miocene Amphibia from Rudabánya. Palaeont Ital 90(2004):11–29
Roček Z (2013) Mesozoic and Tertiary Anura of Laurasia. Palaeobio Palaeoenv 93:397–439. doi:10.1007/s12549-013-0131-y
Roček Z, Nessov LA (1993) Cretaceous anurans from Central Asia. Palaeontographica Abt A 266:1–54
Roček Z, Rage JC (2000) Tertiary anura of Europe, Africa, Asia, North America, and Australia. In: Heatwole H, Carroll RL (eds) Amphibian Biology, vol 4, Paleontology. Surrey Beatty, Chipping Norton, pp 1332–1387
Roček Z, Wuttke M (2010) Amphibia of Enspel (Late Oligocene, Germany). In: Wuttke M, Uhl D, Schindler T (eds) Fossil-lagerstätte Enspel - exceptional preservation in an Upper Oligocene maar. Palaeobio Palaeoenv 90:321–340
Roček Z, Eaton JG, Gardner J, Přikryl T (2010) Evolution of anuran assemblages in the Late Cretaceous of Utah, USA. Palaeobio Palaeoenv 90:341–393
Roček Z, Dong L, Přikryl T, Sun C, Tan J, Wang Y (2011) Fossil frogs (Anura) from Shanwang (Middle Miocene; Shandong Province, China). Geobios 44:499–518
Ročková H, Roček Z (2005) Development of the pelvis and posterior part of the vertebral column in the Anura. J Anat 206:17–35
Roelants K, Bossuyt F (2005) Archaeobatrachian paraphyly and Pangaean diversification of crown-group frogs. Syst Biol 54:111–126
Sahni A (1972) The vertebrate fauna of the Judith River Formation, Montana. Am Mus Nat Hist Bull 147:323–412
Sahni A, Kumar K, Hartenberger J-L, Jaeger J-J, Rage J-C, Sudre J, Vianey-Liaud M (1982) Microvertébrés nouveaux des Trapps du Deccan (Inde): Mise en evidence d’une voie de communication terrestre probable entre la Laurasie et l’Inde à la limite Crétacé-Tertiaire. Bull Soc Géol Fr 24:1093–1099
Sanchíz FB (1977) Catalogo de los anfibios fosiles de Espana (Noviembre de 1977). Acta Geol Hisp 12:103–107
Sanchíz B (1983) The fossil record of living European amphibians. Abstracts 2nd Ord. Gen. Meet. SEH, Leon, pp. 16–17
Sanchíz B (1984) Algunas faunas pleistocenicas de islas del Mediterraneo oriental. In: Castroviejo (Ed.) Actas II Reunion Iberoamericana Conservacion Zoologia Vertebrados, pp. 59–69. Estacion Biol. Doñana, Sevilla
Sanchiz B (1998) Salientia. In: Encyclopedia of Paleoherpetology, Part 4 (ed. Wellnhofer P). Verlag Dr. Friedrich Pfeil, München, pp. 1–275
Sanchíz FB, Mlynarski M (1979) Remarks on the fossil anurans from the Polish Neogene. Acta Zool Cracov 24:153–174
San Mauro D, Vences M, Alcobendas M, Zardoya R, Meyer A (2005) Initial diversification of living amphibians predated the breakup of Pangaea. Am Nat 165:590–599
Schmalhausen JJ (1907) Die Entwickelung des Skelettes der vorderen Extremität der anuren Amphibien. Anat Anzeiger 31:177–187
Skutschas PP, Bannikov AF (2009) The first find of a spadefoot toad (Anura, Pelobatidae) in the Miocene of Moldova. Paleont J 43:433–437
Smith ME, Carroll AR, Singer BS (2008) Synoptic reconstruction of a major ancient lake system: Eocene Green River Formation, western United States. Geol Soc Am Bull 120:54–84
Smith ME, Chamberlain KR, Singer BS, Carroll AR (2010) Eocene clocks agree: coeval 40Ar/39Ar, U-Pb, and astronomical ages from the Green River Formation. Geology 38:527–530
Smith R (2003) Les vertébrés terrestres de l’Oligocène inférieur de Belgique (Formation de Borgloon, MP 21): inventaire et interprétation des données actuelles. Coloquios Paleontol 1:647–657
Smith T, Rose KD, Gingerich PD (2006) Rapid Asia–Europe–North America geographic dispersal of earliest Eocene primate Teilhardina during the Paleocene–Eocene Thermal Maximum. Proc Natl Acad Sci USA 103:11223–11227. doi:10.1073/pnas.0511296103
Smith T, Rana RS, Missiaen P, Rose KD, Sahni A, Singh H, Singh L (2007) High bat (Chiroptera) diversity in the early Eocene of India. Naturwissenschaften 94:1003–1009
Solé F, Smith T (2013) Dispersals of placental carnivorous mammals (Carnivoramorpha, Oxyaenodonta & Hyaenodontida) near the Paleocene-Eocene boundary: a climatic and almost worldwide story. Geol Belg 16:254–261
Špinar ZV (1952) Eopelobates bayeri—a new frog from the Tertiary of Bohemia. Sbor ÚÚG Paleontol 19:457–488
Špinar ZV (1972) Tertiary frogs from Central Europe. Academia, Prague
Špinar ZV, Roček Z (1984) The discovery of the impression of the ventral side of Eopelobates anthracinus Parker, 1929 holotype. Amphibia-Reptilia 5:87–95
Špinar ZV, Tatarinov LP (1986) A new genus and species of discoglossid frog from the Upper Cretaceous of the Gobi Desert. J Vertebr Paleontol 6:113–122
Sullivan RM, Augé M, Wille E, Smith R (2012) A new glyptosaurine lizard from the earliest Eocene of Dormaal, Belgium. Bull Soc Géol Fr 183:627–633
Troschel FH (1861) Uebersicht aller bisher aus der Braunkohle des Siebengebirges beschriebenen fossilen Tiere etc. Sitzungsber Niederrheinischen Ges Bonn 1861:55–56
Trueb L, Hanken L (1992) Skeletal development in Xenopus laevis (Anura: Pipidae). J Morphol 214:1–41
Venczel M (2001) Anurans and squamates from the Lower Pliocene (MN 14) Osztramos 1 locality (Northern Hungary). Fragm Paläontol Hung 19:79–90
Venczel M (2004) Middle Miocene anurans from the Carpathian Basin. Palaeontographica Abt A 271:151–174
Venczel M, Hír J (2013) Amphibians and squamates from the Miocene of Felsötárkány Basin, N-Hungary. Palaeontogr A 300:117–158
Vergnaud-Grazzini C (1970) Les Amphibiens fossiles du gisement d’Arondelli. Palaeontogr Ital 66:45–65
Wagler JG (1830) Natürliches System der Amphibien, mit vorangehender Classification der Saugethiere und Vögel. Gotta, Munich
Wassersug R, Wake D (1995) Fossil tadpoles from the Miocene of Turkey. Alytes 12:145–157
Weitzel K (1938) Propelodytes wagneri n. g. n. sp., ein Frosch aus dem Mittel-eozän von Messel. Notizbl Hess Geol Landesanst 19:42–46
West RM, Dawson MR (1978) Vertebrate paleontology and the Cenozoic history of the north Atlantic region. Polarforschung 48:103–119
Wuttke M (1988) Untersuchungen zur Morphologie, Paläobiologie und Biostratonomie der mitteleozänen Anuren von Messel. Mit einem Beitrag zur Aktuopaläontologie von Anuren und zur Weichteildiagenese der Wirbeltiere von Messel. PhD thesis, University of Mainz, Mainz
Wuttke M (2012a) Redescription of the Middle Eocene frog Lutetiobatrachus gracilis Wuttke in Sanchiz, 1998 (Lower Geiseltalian, “Grube Messel”, near Darmstadt, southern Hesse, Germany). Kaupia—Darmstädter Beitr Naturgesch 18:29–41
Wuttke M (2012b) The genus Eopelobates (Anura, Pelobatidae) from Messel, Geiseltal, and Eckfeld (middle Eocene, Germany). Part I: Redescription of Eopelobates wagneri (Weitzel, 1938) from Messel (lower Geiseltalium, Germany). Kaupia—Darmstädter Beitr Naturgesch 18:43–71
Wuttke M, Přikryl T, Ratnikov VY, Dvořák Z, Roček Z (2012) Generic diversity and distributional dynamics of the Palaeobatrachidae (Amphibia: Anura). Palaeobio Palaeoenv 92:367–395. doi:10.1007/s12549-012-0071-y
Yang J (1977) On some Salientia and Chiroptera from Shanwang, Linqu, Shandong. Vertebr Pal Asiat 15:76–80
Zerova GA (1985) Preliminary results of investigation of Middle Sarmatian herpetofauna of Ukraine. Problems of herpetology – Abstracts of 6th All-Union Herpetological Conference, Tashkent, p. 78. Nauka, Leningrad
Zweifel RG (1956) Two pelobatid frogs from the Tertiary of North America and their relationships to fossil and recent forms. Am Mus Novit 1762:1–45
Acknowledgements
We thank Sandra Chapman (Natural History Museum, London), Boris Ekrt (National Museum, Prague), Yuri Gubin (Paleontological Institute, Moscow), Robert Farrar and Neal Larson (Black Hills Institute, Hill City), and Daniel Brinkman and Jacques Gauthier (Yale Peabody Museum, New Haven) for allowing us to study specimens under their care; Larry Hutson and Ashley Waldorf (Black Hills Institute) for help with procuring a replica of the holotype of Eopelobates deani; and Graeme Housego for accessioning that replica into the Royal Tyrrell Museum of Palaeontology collections. Robert Farrar deserves additional thanks for having canvassed collectors and dealers on our behalf about locality information for the paratype of E. deani and for hosting J.D.G.’s visit to the Black Hills Institute in October 2013. Meinolf Hellmund (Geiseltalmuseum, Halle) provided information on the lithostratigraphy of the Cecilie III and IV localities. Amy Henrici (Carnegie Museum of Natural History, Pittsburgh) and Jean-Claude Rage (Muséum national d’Histoire naturelle, Paris) reviewed the manuscript and provided helpful comments and suggestions. Expenses for museum visits were covered by the Geological Institute, Academy of Sciences of the Czech Republic, Prague for Z.R., by the General Department of Cultural Heritage Rhineland Palatinate for M.W., and by the Royal Tyrrell Museum Cooperating Society for J.D.G. The photograph in Fig. 2a is courtesy of the Natural History Museum (London) and the photographs in Fig. 7a, b are from the archive of Zdeněk V. Špinar currently housed in the Department of Paleontology, National Museum, Prague.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Roček, Z., Wuttke, M., Gardner, J.D. et al. The Euro-American genus Eopelobates, and a re-definition of the family Pelobatidae (Amphibia, Anura). Palaeobio Palaeoenv 94, 529–567 (2014). https://doi.org/10.1007/s12549-014-0169-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-014-0169-5