
Abstract
A new species of Sinemys (Testudines: Cryptodira: Sinemydidae), S. brevispinus n. sp. is described on the basis of three skeletons from the Early Cretaceous Luohandong Formation, Chabu Sumu, Ordos Basin, Inner Mongolia. The new taxon is characterised by an elongate carapace with a pair of distinct but short lateral spines, a sculptured shell surface, closed lateral carapacial fontanelles and an open incisura columellae auris. A nearly complete turtle skeleton from the Early Cretaceous of Hedaochuan, Huanxian, Gansu Province which was previously referred to S. lens is revised and assigned to S. cf. brevispinus. The systematics of Sinemydidae is discussed and the family is restricted to the genus Sinemys. S. brevispinus likely lived in a fluvial environment with the spines acting as a stabiliser to limit the rolling of the carapace, as in S. gamera, but the difference in the shape of the shell suggests that these two species were adapted to different hydrodynamic regimes. The distribution of Sinemys adds to the evidence for biogeographically distinct aquatic communities in eastern and western China during the Early Cretaceous.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Sinemys is a highly aquatic turtle with a flat shell and a seventh peripheral that is laterally expanded, resulting in a lateral process that varies in size from a small process to a large spine. Two species of Sinemys are recognised: S. lens Wiman, 1930 from the Late Jurassic or Early Cretaceous of the Mengyin Formation of Shandong and S. gamera Brinkman and Peng, 1993 from the Early Cretaceous Luohandong Formation of Inner Mongolia (Brinkman and Peng 1993a; Wiman 1930; Ye 1963). Here we report a new species of Sinemys from the Early Cretaceous of Inner Mongolia. The material was collected by the Sino-Canadian expedition from the vicinity of Chabu Sumu, west of Otog Qi, Ordos Basin, Inner Mongolia in 1990, together with several specimens of the champsosaur Ikechosaurus sunailinae, near a major footprint track site (Brinkman and Dong 1993; Dong 1993).
This description is based on three skeletons, one of which is fully prepared. This latter specimen was mentioned by Gaffney and Ye (1992) and used for comparisons. A nearly complete Sinemys skeleton reported by Ye (1963) from the Early Cretaceous of Hedaochuan, Huanxian, Gansu Province is revised. All specimens are housed in the Institute of Vertebrate Paleontology and Paleoanthropology (IVPP), Academia Sinica, Beijing, China.
Systematic palaeontology
Testudines Linnaeus, 1758
Cryptodira Cope, 1868
Eucryptodira Gaffney, 1975
Sinemydidae Ye, 1963
Sinemys Wiman, 1930
Sinemys brevispinus n. sp.

Sinemys brevispinus n. sp. (IVPP V9538-1) from the Early Cretaceous Luohandong Formation of Chabu Sumu, Ordos Basin, Inner Mongolia, China. Skull in dorsal (a, b), ventral (c, d) and right lateral (e) views. Scale bar: 2 cm bo basioccipital; bs basisphenoid; ex exoccpital; op opisthotic; pa parietal; pr prootic; qu quadrate; so supraoccipital; sq squamosal

Sinemys brevispinus n. sp. (IVPP V9538-1) from the Early Cretaceous Luohandong Formation of Chabu Sumu, Ordos Basin, Inner Mongolia, China. Carapace (a, b), plastron (c, d) and detail of ornamentation on the carapace (e). Scale bar: 5 cm (a–d); 1 cm (e)

Sinemys brevispinus n. sp. (IVPP V9538-1) from the Early Cretaceous Luohandong Formation of Chabu Sumu, Ordos Basin, Inner Mongolia, China. Humerus in dorsal (a) and ventral (b) views, scapula (c), cervical 4 (d), cervical 5 (e) and cervical 6 (f) in lateral view (the cervical 4 and 6 are mirror-image for convenience). Scale bar: 2 cm. Vertical scale bar for a–c and horizontal scale bar for e, f
Holotype: IVPP V9538-1, a nearly complete skeleton including the posterior part of the skull, a complete series of cervical vertebrae, a partial carapace articulated with a complete plastron, pectoral and pelvic girdles, some limb bones and caudal vertebrae.
Hypodigm: Two articulated skeletons (IVPP V9538-2 and IVPP V9538-3).
Locality and horizon: Chabu Sumu, 63 km west of Otog Qi (37°52′N, 107°35′E), Ordos Basin, Inner Mongolia, China; Early Cretaceous Luohandong Formation, Zhidan Group (Brinkman and Dong 1993).
Etymology: brevis, Latin: short; spinus, Latin: spine.
Diagnosis: A species of Sinemys. Differs from S. lens Wiman, 1930 in having an ornamented shell surface and a well-developed lateral spine in the adult. Differs from S. gamera Brinkman and Peng, 1993 in having an elongate carapace that is less abbreviated posteriorly, a shorter lateral spine, closed lateral carapacial fontanelles and an open incisura columellae auris.
Measurements: see Table 1 .
Description and comparisons
Preservation: The type specimen IVPP V9538-1 is well preserved, with clear sutures, although the shell is somewhat dorsoventrally crushed. The carapace is damaged, lacking the middle portion and the posterior part of the right side. IVPP V9538-2 and IVPP V9538-3 are preserved on one block, both visible in ventral view and dorsoventrally flattened.
Shell surface ornamentation (Fig. 2e): The whole shell surface is decorated by irregular bumps and winkles. The ornamentation of the plastron is similar to that of the carapace, but finer.
Skull (Fig. 1)
The skull is low and narrower than that of S. gamera. The temporal emargination, although incompletely preserved, is well-developed. It is comparable to that of S. gamera (IVPP V9532-11) (Brinkman and Peng 1993a), exposing the entire otic capsule in dorsal view. The crista supraoccipitalis is broken; its dorsal surface is flat and broader than that of S. gamera. The skull surface is worn, although some light ornamentation is present. In comparison, the skull surface of S. gamera is ornamented with irregular bumps and winkles that match those of the carapace.
Parietal: Only the posterior part of the parietals is preserved, showing long tapering posterolateral edges on the skull surface. The processus inferior parietalis ends well medial to the processus trochlearis oticum as in other Sinemys species. It contacts the prootic ventrally and the supraoccipital posteroventrally.
Pterygoid: Bothe pterygoids are incomplete anteriorly, lacking the processus pterygoideus externus. The pterygoid contacts the basisphenoid medially and the quadrate laterally, and forms the ventral margin of the fenestra postotica. There is also a posteromedial contact with the opisthotic, preventing a contact between the pterygoid and basioccipital as in S. gamera. The canalis caroticus internus and canalis caroticus lateralis are open as a groove that follows the basisphenoid/pterygoid suture; the groove is mainly formed by the basisphenoid. This differs from S. gamera (IVPP V9532-11), which has a partly closed canal. The canal ends near the posterior end of the pterygoid, so that the foramen posterius canalis carotici interni is located lateral to the ventral exposure of the opisthotic, instead of anterior to the opisthotic as in S. gamera. Inside the canal, the foramen posterius canalis caroticus cerebralis (Brinkman et al. 2012) is well preserved on both sides. This structure is located along the medial margin of the carotid canal, where the basisphenoid narrows. The carotid artery branches into the cerebral and palatine arteries at this point; the cerebral artery enters the skull through the foramen posterius caroticus cerebralis while the palatine artery continues forward in a well-defined groove. The anterior end of this groove is missing; the foramen posterius canalis caroticus lateralis is not preserved.
Basisphenoid: the basisphenoid is nearly complete. As in S. gamera, the basisphenoid is an elongate element, with a narrow anterior part and a wide posterior portion. Since its anterior end is damaged, it is uncertain whether or not the basisphenoid completely separates the pterygoids. The basisphenoid and the basioccipital are separated by a distinct suture, rather than fused as in S. gamera (Brinkman and Peng 1993a). The ventral surface of the basisphenoid is slightly convex, without a distinct pair of pits, in contrast to S. gamera and other sinemydids/‘macrobaenids’. The basisphenoid has a pair of lateral processes (basipterygoid processes) which extends lateral to the canalis caroticus lateralis. Both processes are damaged, but the right one is better preserved. Such processes are also present in S. gamera, but differ in that the processes are located in a large depression. The basisphenoid contacts the pterygoid laterally, the basioccipital posteriorly and the opisthotic posterolaterally.
Basioccipital: the basioccipital is complete. The shape is similar to that of S. gamera, but with less strongly developed tubercula basioccipitalis and a slightly concave ventral surface. The posterior end of the basioccipital is covered by matrix and hidden by the attached cervical vertebrae, so the condylus occipitalis is not visible. The basioccipital contacts the basisphenoid anteriorly, the opisthotic anterolaterally and the exoccipital laterally.
Exoccipital: the exoccipitals are mostly hidden by the attached cervicals; the shape and contacts seem to be similar to those of S. gamera. The ventral exposure of the exoccipital lateral to the basioccipital is large and triangular as in S. gamera. It forms the medial margin of the foramen jugulare posterius.
Supraoccipital: the supraoccipital is nearly complete, lacking the posterior end of the crista supraoccipitalis. The crista supraoccipitalis has a flat dorsal surface, so the cross section is rectangular. This is different from the triangular cross section of the crista supraoccipitalis in S. gamera. The supraoccipital contacts the parietal anteriorly, the prootic anterolaterally and the opisthotic posterolaterally. The contact with the exoccipital is not visible because of the attached cervicals.
Quadrate: Both quadrates are damaged. The cavum tympani are incomplete on both sides. The incisura columellae auris is not fully closed by bone, but connected to the Eustachian tube by a narrow gap. This is different from the fully closed incisura of S. gamera. A large precolumellar fossa is present anterior to the incisura columellae auris as in S. gamera, although this fossa is not as deep as in S. gamera. Ventrally, both condyli mandibularis of the quadrate are damaged on their lateral end. The articulation surfaces are flat. Anteriorly, the quadrate forms the lateral portion of the processus trochlearis oticum as in S. gamera. The quadrate contacts the prootic anteromedially, the opisthotic posteromedially and the squamosal posteriorly on the dorsal surface; it also contacts the pterygoid medially on the ventral surface.
Prootic: Both prootics are complete. Dorsally, the prootic is an elongate element between the parietal and quadrate, which appears to be narrower than that of S. gamera. The prootic forms medial part of the processus trochlearis oticum and the dorsal rim of the foramen nervi trigemini. The processus trochlearis oticum is moderately developed as in S. gamera. The foramen stapedio-temporale is located on the prootic/quadrate suture and extended by a posteromedially directed groove in the prootic as in S. gamera. The diameter of the foramen is larger than the foramen posterius canalis caroticus cerebralis.
Opisthotic: Both opisthotics are complete and in contact with the prootic anteriorly, the supraoccipital medially, the quadrate anterolaterally and the squamosal laterally. The processus interfenestralis of the opisthotic is well developed and more exposed posteroventrally than in S. gamera, completely separating the foramen jugulare posterius and the fenestra postotica. On the ventral surface of the skull, the opisthotic sends a short process between the basioccipital and the pterygoid as in S. gamera.
Shell (Fig. 2)
Carapace: The carapace is oval and elongate and is narrower than that of S. lens and S. gamera. The posterior part of the carapace is not as shortened as in S. gamera. The complete cervical notch is distinct and wide. The carapace is very low as other Sinemys spp. There is a clear midline longitudinal depression, extending from the posterior margin of the nuchal posteriorly to the suprapygal. The neurals are on the bottom of the groove. Such a depression is also present in S. lens and S. gamera (Brinkman and Peng 1993a). The lateral borders of the carapace diverge posteriorly to the seventh peripheral where a lateral spine is formed, then converge to the eleventh peripheral. The lateral border of the carapace from the nuchal to the fifth peripheral is blunt and that of the third to fifth peripherals turned downwards, while the posterolateral border from the seventh through eleventh peripherals is horizontal and sharp. From the third peripheral posteriorly, each peripheral bears a light process, making the carapace margin slightly serrated. A series of small isolated openings is present between the costals and peripherals, exposing the free rib ends on the dorsal surface. On the inner surface, the first thoracic rib is long and sutured to the first costal, reaching the second peripheral and almost contacting the axillary buttress. IVPP V9538-3 and IVPP V9538-2 have a slightly wider shell than in IVPP V9538-1.
The nuchal is complete. It is very wide, and its width is threefold its length. The first neural is complete and is rectangular and narrow. The second neural is nearly complete and is six-sided with short anterolateral sides and a concave lateral border. The remaining neurals are not preserved. Two suprapygals are preserved. The first suprapygal is incomplete, lacking its right portion. It is wider than long and slightly narrower than the second suprapygal. In comparison, the first suprapygal is much smaller than the second in S. lens and Manchurochelys manchoukuoensis. The second suprapygal is almost complete. It is much wider than long and contributes to the posterior border of the carapace. A pygal is absent, as in other Sinemys spp. Eight costal plates are present. The first costal is roughly as long as the second costal. All costals have a distinct free rib end inserting on the socket of the peripheral, visible on the dorsal surface of the carapace. The complete peripheral series are preserved on the left side and the first to fourth peripherals and anterior portion of the fifth peripheral are preserved on the right side. The first to fifth peripherals are narrow. The sixth and seventh peripherals are fused dorsally and bear the lateral spine, while the suture between the sixth and seventh peripherals is visible on the ventral surface in both IVPP V9538-1 and IVPP V9538-2. The seventh to tenth peripherals are wide. The eleventh peripheral is a small triangle and separated from its counterpart by the second suprapygal. The free rib end of the first costal inserts onto the anterior part of the third peripheral; that of the second and third costals inserts onto the middle of the fourth and fifth peripherals respectively; that of the fourth costal inserts onto the middle of fused sixth and seventh peripherals; that of the sixth to eighth costals inserts onto the limit of the peripherals.
There is no cervical scute and the first vertebral scute is exposed on the anterior carapace margin as in Sinemys lens and Dracochelys bicuspis (Brinkman and Peng 1993a; Brinkman 2001). The first vertebral reaches the anterior margin of the carapace; it is much wider than the nuchal, reaching the lateral end of the first peripheral. The second vertebral is almost as long as wide. The incomplete fourth vertebral is as wide as the second vertebral. The fifth vertebral is exposed on the posterior margin of the carapace, separating the pair of twelfth marginals. All marginals are narrow and restricted to the peripherals except the second one. The third to fifth marginals are slightly narrower than the corresponding peripherals while the widths of the seventh to tenth marginals are less than half width of the corresponding peripherals. The first and the twelfth marginals are triangular.
Plastron: The plastron is as long as the carapace and cruciform, with the anterior border extending slightly beyond the anterior carapace margin. The hyoplastron is loosely connected to the second and third peripherals. The hypoplastron contacts the carapace by means of a series of pegs inserting into one large pocket on the sixth and seventh peripherals. The anterior lobe is triangular. The bridge is long, being as long as the posterior lobe. The posterior lobe is very long and narrow, with parallel lateral borders and extends posteriorly nearly to the posterior end of the carapace. There is a large oval central fontanelle between the hyoplastron and hypoplastron and a small and narrow fontanelle between the hypoplastron and the xiphiplastron. A large slit-like lateral fontanelle is present between the hyoplastron, hypoplastron and the peripherals as in S. lens (Fig. 2c, d).
The epiplastron and entoplastron are slightly displaced. The epiplastron is a small triangule and contacts its counterpart along its full length. The entoplastron is broad and separates the epiplastron from the hyoplastron as in S. lens. The bridge part of the hyoplastron is similar in length to the bridge part of the hypoplastron, while the midline length of the latter is greater than that of the hyoplastron. The xiphiplastron is very narrow and elongate.
The humeropectoral sulcus is located on the base of the anterior lobe and convex posteriorly. The pectoroabdominal sulcus crosses the anterior part of the central fontanelle with the lateral end turned posteriorly. The abdominofemoral sulcus originates from the inguinal notch and crosses the posterior end of the central fontanelle. The femoroanal sulcus is greatly convex anteriorly and extends onto the hypoplastron. The inframarginals are not visible.
Vertebral column
Cervical vertebrae (Figs. 1a, d, 3d–f): The complete cervical series is preserved; the first to the third cervicals are articulated and attached to the skull, the fourth to sixth cervicals are preserved as separate elements and the seventh and eighth cervicals are attached to the shell. The centrum of the atlas is disarticulated from the occipital condyle. The atlas arches is a pair of roughly triangular-shaped plates in dorsal view, with rounded anterior end and narrow posterior end (Fig. 1a, b). The centra, from the axis to the seventh cervicals, are long and their length remains unchanged. The centrum of the atlas has a very low and broad convex anterior articulation surface, which is displaced ventrally. The second to seventh cervicals have opisthocoelous centra which bear a low ventral ridge and a single transverse process. The transverse process is located close to the anterior end of the centrum on the axis. Its position shifts to the middle of the centrum posteriorly along the column, being close to the middle of the centrum on the seventh cervical. The eighth cervical is only partially exposed. The posterior articulation surface, from the axis to the fifth cervical, is slightly higher than wide, while the posterior articulation surface of the sixth cervical is slightly wider than high. All articulations are single. These cervical vertebrae differ from those of Dracochelys and Ordosemys in having a longer centrum and a single transverse process (Brinkman 2001; Brinkman and Peng 1993b).
Dorsal vertebrae: the dorsal vertebrae are preserved in articulation to the carapace, but only part of them are exposed through the plastral fontanelles. The dorsal vertebrae are long with a blunt ventral surface.
Caudal vertebrae (Fig. 2c, d): Five and one-half articulated caudal vertebrae are exposed in ventral view. The first three caudals are procoelous, the fourth one is amphicoelous and the two more posterior ones are opisthocoelous. These caudals have a pair of long transverse processes that are as long as their centra and located at the midlength of the centrum. There is a pair of ventral processes on the posterior end of the centrum, similar to those of Ordosemys leios, visible on the second to fifth caudals.
Appendicular skeleton
Pectoral girdle (Figs. 2c, d, 3c): Both scapulae are preserved but incomplete, with the extremities damaged. The scapular process forms an angle of 60° with the acromion process, which is smaller than in Dracochelys bicuspis and Ordosemys leios (Brinkman 2001; Brinkman and Peng 1993b) and reflects the lower carapace in Sinemys spp. A distinct neck is present on the scapular at the base of the glenoid as in D. bicuspis and O. leios. The coracoid is preserved on the left side, with the distal end hidden by the plastron.
Humerus (Fig. 3a, b): A nearly complete right humerus and a proximal portion of the left humerus are preserved. The humerus is more slender with less expanded proximal and distal ends than in Dracochelys bicuspis and Ordosemys leios. The oval-shaped articular head is upturned from the shaft. The medial process is slightly lower than the head. The fossa between the medial and lateral processes is ill-defined. The ectepicondyle foramen is closed. The humerus of S. brevispinus differs from that of O. leios in the absence of the sharp ridge on dorsal surface of the shaft along the base of the lateral process.
Pelvis girdle (Fig. 2c, d): The pelvic girdle is presented by both pubes and ischia, and the left ilium. The pubes and ischia are preserved in their anatomical position, but are somewhat crushed. The position of the pelvic girdle relative to the posterior end of the plastron is comparable to Ordosemys leios (Brinkman and Peng 1993b). The ilium is incomplete and has a blade-like main body. The pubis has a blade-like ventral plate and the pectineal process has a rugosity at its anterolateral end. The ischium contributes to the formation of the acetabulum. The metischial process is directed posteriorly. The obturator fenestrae are large and at least the posterior portion is divided by the ischium.
Femur (Fig. 2c, d): Both femora are preserved in articulation to the acetabulum and visible in anterior view in V9538-1, but the distal ends are missing. The femur is like the humerus in being long, slender and having a slightly S-curved shaft. The shaft has a nearly round cross section.
Sinemys cf. brevispinus (Fig. 4)
Referred material: IVPP V1034; Early Cretaceous, Hedaochuan, Huanxian, Gansu Province.

Sinemys cf. brevispinus (IVPP V1034) from the Early Cretaceous of Hedaochuan, Huanxian, Gansu Province; in dorsal view (a, b) and detail of ornamentation on the carapace (c). Scale bar : 5 cm (a, b); 1 cm (c)
Preservation: Almost complete skeleton, including skull, neck, carapace, tail and part of front and hind limbs.
Description and comparisons: The skull is visible in ventral view but crushed, and therefore provides few details; although the general morphology is comparable to that of IVPP V9538-1. The complete series of cervical vertebrae is mostly visible in ventral view.
The carapace is almost complete. It is smaller than IVPP V9538-1 (for measurements see Table 1). The ornament on the surface of the carapace is similar to IVPP V9538-1, but appears to be sharper. The carapace shape, and the pattern of bones and scutes forming the carapace is similar to IVPP V9538-1. The lateral spine is damaged on both sides. The imprint on the right side indicates that it is short, even shorter than in IVPP V9538-1, but more developed than in adult Sinemys lens (Brinkman and Peng 1993a). There are six neurals. In addition, a prenueral is likely present. IVPP V1034 differs from IVPP V9538-1 by the presence of a hexagonal first neural with short posterolateral sides, the rectangular second neural and the triangular first peripheral with a very short contact with the first costal.
IVPP V1034 is preserved together with several specimens of Sinamia and was originally assigned to S. lens (Ye 1963). Based on the carapace morphology, this specimen is referred here to S. cf. brevispinus.
Discussion
Sinemys is a highly derived aquatic turtle. The genus is characterised by a series of autapomorphic features: (1) the cervical scute is absent and the first vertebral scute enters the anterior edge of the carapace; (2) the pygal is lost, and the second suprapygal forms the posterior margin of the carapace; (3) the fifth vertebral scute enters posterior edge of the carapace; (4) the entoplastron is a transverse element that completely separates the epiplastra from hyoplastra (Brinkman and Peng 1993a). In addition, the seventh peripheral is laterally expanded, resulting in a convex to angled lateral edge or a long spine. All of these characters are present in the specimens described above.
Two species of Sinemys have been hitherto recognised: S. lens Wiman, 1930 from the Mengyin Formation, Ninjiagou, Xintai, Shandong Province (Wiman 1930) and S. gamera Brinkman and Peng, 1993 from the Luohandong Formation, Laolonghezi, west of Hanjinqi, Inner Mongolia (Brinkman and Peng 1993a). S. lens differs from S. gamera by its smooth shell surface, absence of the costal-peripheral fontanelles, slitlike lateral fontanelle on the plastron and a seventh peripheral with a convex lateral edge or a weak lateral process, except in the juvenile, in which a spine is present. Ye (1963) reported a skeleton of Sinemys lens from the Early Cretaceous of Hedaochuan, Huanxian, Gansu Province; this specimen is assigned here to S. cf. brevispinus.
The new specimens described here as S. brevispinus are more similar to S. gamera in the sculptured shell surface and development of a lateral spine on the carapace. S. brevispinus has an elongate carapace with a pair of short spine on the seventh peripheral. The continuous growth series of S. lens shows considerable change in carapace shape and spine length. The smallest juvenile specimen of S. lens has a short and wide carapace with greatly abbreviated posterior end and a quite long spine as in large specimen of S. gamera; while the carapace of large adult individual of S. lens is elongate, with longer posterior part and the seventh peripheral is only laterally expanded, without a distinct spine. In S. gamera, the spine is variable in length (Brinkman and Peng 1993a; fig. 7). More than 20 isolated seventh peripherals of S. gamera show the spine length along its anterior edge to range from 36 to 90 mm. In S. brevispinus, the spine length of all three specimens is about 10 mm.
S. gamera is considered to be neotenic because its carapace morphology and the presence of costal-peripheral fontanelles are similar to that of a juvenile of S. lens (Brinkman and Peng 1993a). IVPP V9538-1 is an adult or sub-adult, and its size is comparable to that of S. gamera and adult S. lens; its carapace shape resembles that of adult S. lens. The costal-peripheral fontanelles in IVPP V9538-1 are greatly reduced as a series of isolated openings, exposing the rib end on the carapace surface.
The skull of IVPP V9538-1 has open canalis caroticus internus and canalis caroticus lateralis. The unfloored internal carotid canal is known in the xinjiangchelyid Annemys levensis and is considered to be a plesiomorphic feature (Sukhanov 2000). In Sinemys lens, S. gamera, Dracochelys bicuspis and the ‘macrobaenids’ Ordosemys leios, O. liaoxiensis, Kirgizemys dmitrievi and K. hoburensis, the internal carotid canal is covered ventrally at least partially by a thin layer of bone (Brinkman and Peng 1993a, b; Danilov et al. 2006; Gaffney and Ye 1992; Li and Liu 1999; Sukhanov 2000; Tong et al. 2004; ). In addition, S. brevispinus differs from S. gamera in having an open incisura columellae auris. The open incisura columellae auris is widespread in sinemydids/‘macrobaenids’, such as Dracochelys bicuspis, Ordosemys sp. and Kirgizemys dmitrievi (Brinkman and Wu 1999; Danilov et al. 2006; Gaffney and Ye 1992; ), and is considered to be a primitive feature. An unusual feature on the S. brevispinus skull is the absence of a pair of pits on the ventral surface of the basisphenoid. According to Brinkman and Peng (1993a), these pits likely mark the position of insertion of major ligaments from the neck and are widely spread in sinemydids/‘macrobaenids’ such as Sinemys lens and S. gamera, Kirgizemys dmitrievi, K. hoburensis, Ordosemys liaoxiensis and O. sp. and also the basal trionychoid Basilochelys macrobios (Brinkman and Peng 1993a, b; Danilov et al. 2006; Li and Liu 1999; Sukhanov 2000; Tong et al. 2004; Tong et al. 2009).
The family Sinemydidae
The family of Sinemydidae was created by Ye in 1963 to include two monotypic genera: Sinemys Wiman, 1930 and Manchurochelys Endo and Shikama 1942. Ye (1963) defined the family by the following characters: shell oval or sub-rounded covered by scutes; carapace low; eight neurals; two suprapygals where the second one is very wide; nuchal large and wide; vertebrals mostly hexagonal; costoperipheral sutures usually above the pleuromarginal sulci; plastron reduced and not fully ossified, cruciform, with narrow and elongate posterior lobe; medial and lateral plastral fontanelles present; femoral and anal scutes very narrow.
Brinkman and Peng (1993a) retained Ye’s (1963) concept of the Sinemydidae, characterising the family by two synapomorphies: the elongate posterior lobe with a narrow base and the first suprapygal being smaller than the second. An additional taxon, Dracochelys bicuspis Gaffney and Ye, 1992 was later included in the Sinemydidae (Brinkman 2001). The most recent phylogenetic analyses recognise the Sinemydidae as a monophyletic group consisting of Sinemys, Manchurochelys and Dracochelys, united by three characters: postorbital/squamosal contact absent, biconvex cervical 8 and low spine on cervical 8 (Zhou 2010a, b).
Chkhikvadze (1976) expanded the Sinemydidae to also include Macrobaena Tatarinov, 1959, Kirgizemys Nessov and Khosatzky, 1973, Hangaiemys Sukhanov and Narmandakh 1974 and Yaxartemys Riabinin, 1948. These genera share with Sinemys and Manchurochelys the plastron with reduced axillary and inguinal buttresses, well-developed plastral fenestra and a short, triangular anterior lobe. Gaffney (1996) redefined the Sinemydidae by including Sinemys, Dracochelys/Hangaiemys, Ordosemys and TMP 87.2.1 (which was later named as Judithemys sukhanovi; Parham and Hutchison 2003) based on two unambiguous synapomorphies: a thin floor of the canalis caroticus internus and biconvex cervical 8. Following the concept of Gaffney (1996), Brinkman et al. (2008) included all genera from the Early Cretaceous of China that were previously placed in Sinemydidae/‘Macrobaenidae’ in the Sinemydidae. These include Sinemys, Wuguia Matzke, Maisch, Pfretzschner, Sun et Stöhr, 2004, Dracochelys Gaffney and Ye, 1992, ‘S.’ wuerhoensis Ye, 1973, Honkongchelys Ye, 1999, Manchurochelys, Ordosemys Brinkman et Peng, 1993 and Yumenemys Bohlin, 1953 that are united by the biconvex cervical 8. Two distinct groups are recognised among these genera: (1) a primitive group typified by Dracochelys, which has a complete row of marginal scutes meeting posteriorly on the pygal; (2) a derived group, including Sinemys, in which the marginal series is reduced and the vertebral 5 enters the posterior margin of the carapace (Brinkman et al. 2008). Almost all previous phylogenetic analyses agree that Sinemydidae/‘Macrobaenidae’ form a paraphyletic group (Anquetin 2011; Brinkman and Wu 1999; Gaffney 1996; Gaffney et al. 2007; Joyce 2007; Parham and Hutchison 2003; Vandermark et al. 2009; Zhou 2010a, b).
On the other hand, Khosatzky and Nessov (1979) argue that the Sinemydidae and ‘Macrobaenidae’ should not be united because derived features of Sinemys are not present in the ‘macrobaenids’. Brinkman and Peng (1993a) point out that the presence of a reduced, fenestrated plastron should be considered as a primitive character state since it is widespread in other groups of cryptodires, including trionychoids, chelydrids and chelonioids, and the short and subtriangular-shaped posterior lobe seen in ‘Macrobaenidae’ (Hangaiemys, Kirgizemys, Anatolemys and Macrobaena) approaches the condition of the primitives Jurassic cryptodire Xinjiangchelys. Thus, this character is of limited use in establishing suprageneric groups, and it is difficult to define the group on the basis of derived characters. Contrary to the expanded concept of the family, Sukhanov (2000) restricted the Sinemydidae to the genus Sinemys, which is in turn composed of three species: S. lens Wiman, 1930, S. gamera Brinkman et Peng, 1993 and ? S. efremovi Khosatzky, 1996. Manchurochelys was excluded from the Sinemydidae because its shell is strikingly different. The revision of S. efremovi led to the placement of this species in the ‘macrobaenid’ Wuguia (Danilov and Sukhanov 2006). This reduced concept of the Sinemydidae matches with the derived group of sinemydids of Brinkman et al. (2008).
In order to construct a monophyletic group, the concept of the Sinemydidae restricted to Sinemys is adopted here. The monogeneric family includes three species: S. lens Wiman, 1930; S. gamera Brinkman and Peng, 1993 and S. brevispinus n. sp, and is diagnosed as follows: (1) shell length of about 20 cm; (2) carapace flat; the cervical scute absent, and the first vertebral scute enters the anterior edge of the carapace; (3) pygal lost, and the second suprapygal forms the posterior margin of the carapace; (4) fifth vertebral scute enters posterior edge of the carapace; (5) seventh peripheral is laterally expanded, resulting in a convex to angled lateral edge or a long spine; (6) epiplastron is a tiny triangle; (7) entoplastron as exposed ventrally is a transverse element separating completely the epiplastra from hyoplastra; (8) posterior end of the plastron extending nearly to the posterior margin of the carapace; (9) skull low; (10) prefrontals do not meeting along the midline; (11) postorbital short; (12) basisphenoid extending far forward between pterygoids; (13) opisthotic exposed ventrally between basisphenoid/basioccipital, pterygoid and exoccipital.
We exclude Manchurochelys from the Sinemydidae. The new material of Manchurochelys manchoukuoensis, including skull and complete carapace, described recently provides additional information on this taxon (Zhou 2010b). The carapace of Manchurochelys does not present the derived features of Sinemys (absence of cervical and pygal, vertebral 5 enters to the posterior margin and laterally expanded peripheral 7). Concerning the plastron, the anterior lobe remains unknown; the posterior lobe shares with Sinemys the narrow base, but differs in its shortness, since its posterior end is far anterior to the posterior margin of the carapace. In addition to the shell, which is very distinct from that of Sinemys, the long postorbital, the relatively short basisphenoid, the long midline contact of pterygoids and the absence of a ventral exposure of the opisthotic between the basisphenoid/basioccipital, pterygoid and exoccipital in Manchurochelys are more similar to ‘macrobaenids’ than to Sinemys.
Dracochelys is considered here as the sister taxon of Sinemydidae. The shell of Dracochelys shares with that of Sinemys the absence of the cervical scute and the entry of vertebral 1 onto the anterior margin of the carapace. However, Dracochelys retains the pygal, and the marginals meet on the midline at the posterior margin of the carapace (Brinkman 2001). The plastron of Dracochelys shows the generalised shape seen in ‘macrobaenids’, and the skull has a relatively extensive roof with a long postorbital as in ‘macrobaenids’ such as Kirgizemys hoburensis, K. dmitrievi, Ordosemys liaosiensis, O. sp., Liaochelys jianchangensis (Brinkman and Wu 1999; Danilov et al. 2006; Sukhanov 2000; Tong et al. 2004; Zhou 2010a).
Palaeoecology and palaeobiogeography
Turtles are an important component of Early Cretaceous palaeocommunities in China, sometimes, as in the case of Sinemys lens, occurring in great numbers in low diversity assemblages. Sinemys is of particular palaeoecological interest because it is one of the few Early Cretaceous turtles from Asia that typically occurs in fluvial environments. Based on the sedimentology of the beds in which it is found and the associated fauna, Brinkman and Peng (1993a) suggested that S. gamera lived in fast-moving streams. The occurrence of S. brevispinus in a sandstone indicates that this species also lived in a fluvial environment. The environment of deposition of S. lens is unknown, but the absence of Sinemys in any of the well-documented lacustrine or paludal assemblages, such as the Jehol fauna in Liaoning, is consistent with the idea that species of Sinemys were primarily adapted to a fluvial environment. Brinkman and Peng (1993a) suggested that the development of spines in S. gamera was an adaptation for life in fast-flowing streams, with the spines acting as stabilisers to limit the rolling of the carapace around the midline. Since S. brevispinus likely also lived in a fluvial environment, the spines in that species may have served a comparable function, although the more streamlined shell suggests that this turtle was more efficient at swimming than S. gamera. Thus, although it is likely that both these turtles lived in a fluvial environment, the differences in shell shape indicate that they were adapted to different hydrodynamic regimes.
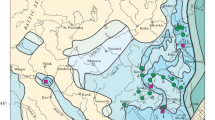
The distribution of Sinemys is of biogeographic interest with respect to evidence presented by Chang and Miao (2004) for the presence of three distinct faunal zones in the Early Cretaceous of Asia. Based on the fish assemblages, Chang and Miao (2004) recognised a western zone, a north-eastern zone, and a south-eastern zone (Fig. 5). The western zone is present in Xinjiang and adjacent portions of Mongolia and is characterised by the presence of abundant archaeomaenids or archaeomaenid-like fishes. Teleosts are absent in fish assemblages from this region. The north-eastern zone is present in eastern Siberia, Mongolia, northern China and northern Korea. It contains several forms endemic to this area, including members of the Peipiaosteidae and the Polyodontidae (both members of the Asipenceriformes), and the basal osteoglossomorph Lycoptera. The south-eastern zone is present in southern China, southern Korea, and northern Kyushu, Japan. This zone is characterised by the presence of ichthyodectiformes and clupeomorphs, particularly Paraclupea. The border between the Lycoptera assemblage and the Paraclupea assemblage is imprecise, since the taxa Paralycoptera and Huashia appear to overlap between the two assemblages. As well, the south-eastern and north-eastern zones are similar in that they share the presence of the amiiformes Sinamia and lkechaoamia and the semionotid Neolepidotes. None of these taxa are present in the western zone.

Map of China showing the distribution of Sinemys and the three biogeographic regions recognised by Chang and Miao (2004). 1 Sinemys lens, Xintai, Shandong, 2 Sinemys gamera, Laolonghuozi, Inner Mongolia, 3 S. brevispinus, Chabu Sumu, Inner Mongolia, 4 Sinemys cf. brevispinus, Huanxian, Gansu. A Western region, B north-eastern region, C south-eastern region. Broken lines indicate possible extensions of the western and south-eastern regions
Since turtles are widely distributed in northern China and show a high diversity, they provide an independent test of the extent to which aquatic vertebrates generally show distinct faunal provinces within Asia during the Early Cretaceous. This is particularly so with Sinemys, since Ye (1963) noted an association of Sinemys with the fish Sinamia in Hedaoshuan, Huanxian, Gansu and Ninjiagou, Xintai, Shandong. Since Sinemys is present in Gansu, Inner Mongolia, and Shandong and absent in Xinjiang (Fig. 5) this lack of association adds to the evidence of distinct east–west biogeographic zones. Further, within the eastern region, Sinemys occurs only in the north-eastern zone; however, this provides only weak support for distinct north-eastern and south-eastern zones since little information is available on the diversity of Early Cretaceous turtles from the south-eastern zone. The distribution of the turtle Sinochelys (including Peishanemys) is similar to that of Sinemys since it is also present in Shandong and Gansu, but not Xinjiang or Liaoning. Thus, the distribution of Sinochelys also adds to the evidence for distinct eastern and western zones. Although the absence of Sinemys and Sinochelys in Liaoning may have regional biogeographic significance, their restricted distribution within the north-eastern zone is more likely a result of differences in the ecological conditions being sampled since the Liaoning turtles were recovered from lacustrine deposits while, as discussed above, Sinemys tends to occur in fluvial deposits.
Thus, although our knowledge of the Early Cretaceous turtle fauna in China is still very limited, the distribution of Sinemys and Sinochelys help confirm the presence of biogeographic differentiation of the Early Cretaceous aquatic communities in China by adding to the evidence for differences in the aquatic communities of the eastern and western zones. The distribution of these turtles is also consistent with the presence of distinct north-eastern and south-eastern zones, although support for this is much weaker since there is little information on Early Cretaceous turtles from the south-eastern zone.
References
Anquetin J (2011) Reassessement of the phylogenetic interrelationships of basal turtles (testudinata). J Syst Palaeontol 1:3–45. doi:10.1080/14772019.2011.558928
Brinkman D (2001) New material of Dracochelys (Eucryptodira: Sinemydidae) from the Junggar Basin, Xinjiang, People’s Republic of China. Can J Earth Sci 38:1645–1651
Brinkman D, Dong Z-M (1993) New material of Ikechosaurus sunailinae (Reptilia: Choristodira) from the Early Cretaceous Laohongdong Formation, Ordos Basin, Inner Mongolia, and the interrelationships of the genus. Can J Earth Sci 30:2153–2162
Brinkman D, Peng J-H (1993a) New material of Sinemys (Testudines, Sinemydidae) from the Early Cretaceous of China. Can J Earth Sci 30:2139–2152
Brinkman D, Peng J-H (1993b) Ordosemys leios n. gen., n. sp., a new turtle from the Early Cretaceous of the Ordos Basin, Inner Mongolia. Can J Earth Sci 30:2128–2138
Brinkman D, Wu X-C (1999) The skull of Ordosemys, an Early Cretaceous turtle from the Inner Mongolia, People’s Republic of China, and the interrelationships of Eucryptodira (Chelonia, Cryptodira). Paludicola 2:134–147
Brinkman D, Li J-L, Ye X-K (2008) Order Testudines. In: Li J-L, Wu X-C, Zhang F-C (eds) The Chinese fossil reptiles and their kin. Science Press, Beijing, pp 35–102
Brinkman D, Eberth DA, Xu X, Wu X-C (2012) Turtles from the Jurassic Shishugou Formation of the Junggar Basin, People’s Republic of China, with comments on the basicranial region of basal Eucryptodires. In: Brinkman D, Holroyd PA, Gardner JD (eds) Morphology and evolution of turtles. Springer, Heidelberg Berlin, pp 147-172
Chang MM, Miao DS (2004) An overview of Mesozoic fishes in Asia. In: Arratia G, Tintori A (eds) Meozoic fishes 3—systematics, paleoenvironments, and biodiversity. Verlag Dr. Friedrich Pfeil, Munich, pp 535–563
Chkhikvadze VM (1976) The fossil turtles of the family Sinemydidae. Akad Nauk Gruz SSR, Izv, Ser Biol 3:265–270
Danilov IG, Sukhanov VB (2006) A basal eucryptodiran turtle “Sinemys” efremovi (=Wuguia efremovi) from the Early Cretaceous of China. Acta Palaeontol Pol 51:105–110
Danilov IG, Averianov AO, Skutchas PP, Rezvyi AS (2006) Kirgizemys (Testudines, ‘Macrobaenidae’): new material from the Lower Cretaceous of Buryatia (Russia) and taxonomic revision. Fossil Turtles Res, vol. 1. Russ J Herpetol 13 [Suppl]:46–62
Dong Z-M (1993) The field activities of the Sino-Canadian dinosaur project in China, 1987–1990. Can J Earth Sci 30:1997–2001
Gaffney ES (1996) The postcranial morphology of Meiolania platyceps and a review of the Meiolaniidae. Bull Am Mus Nat Hist 229:1–166
Gaffney ES, Ye X (1992) Dracochelys, a new Cryptodiran turtle from the Early Cretaceous of China. Am Mus Novit 3048:1–13
Gaffney ES, Rich TH, Vickers-Rich P, Constantine A, Vacca P, Kool L (2007) Chubutemys, a new Eucryptodiran turtle from the Early Cretaceous of Argentina, and the relationships of the Meiolaniidae. Am Mus Novit 3599:1–35
Joyce WG (2007) Phylogenetic relationships of Mesozoic turtles. Bull Peabody Mus Nat Hist 48:3–102
Khosatzky LI, Nessov LA (1979) Large turtles from the Late Cretaceous of Soviet Middle Asia. Akad Nauk SSSR, Zool Inst 89:98–107
Li J-L, Liu J (1999) The skull of Manchurochelys laoxiensis (Testudines: Sinemydidae) from the Yixian Formation of Beipiao, Liaoning and phylogenetic position of this taxon. In: Chen P-J, Jin F (eds) Palaeoworld II, Jehol Biota. Press of University of Sciences and Technology of China, Hefei, pp 281–295
Parham JF, Hutchison JH (2003) A new eucryptodiran turtle from the Late Cretaceous of North America (Dinosaur Provincial Park, Alberta, Canada). J Vertebr Paleontol 23:783–798
Sukhanov VB (2000) Mesozoic turtles of Middle and Central Aisa. In: Benton MJ, Shishkin MA, Unwin DM, Kurochkin EN (eds) The Age of Dinosaurs in Russia and Mongolia. Cambridge University Press, Cambridge, pp 309–367
Tong H, Ji S-A, Ji Q (2004) Ordosemys (Testudines: Cryptodira) from the Yixian Formation of Liaoning Province, Northeastern China: New specimens and systematic revision. Am Mus Novit 3438:1–20
Tong H, Claude J, Naksri W, Suteethorn V, Buffetaut E, Khansubha S, Wongko K, Yuangdetkla P (2009) Basilochelys macrobios n. gen. and n. sp., a large cryptodiran turtle from the Phu Kradung Formation (terminal Jurassic-basal Cretaceous) of the Khorat Plateau, NE Thailand. In: Buffetaut E, Cuny G, Le Loeuff J, Suteethorn V (eds) Late Palaeozoic and Mesozoic continental ecosystems of Southeast Asia. Geological Society, London, pp 153–174
Vandermark D, Tarduno JA, Brinkman DB, Cottrell RD, Mason S (2009) New Late Cretaceous macrobaenid turtle with Asian affinities from the High Canadian Arctic. Geology 37:183–186
Wiman C (1930) Fossile Schildkröten aus China. Palaeontol Sinica Ser C 6:5–53
Ye X-K (1963) Fossil turtles of China. Palaeontol Sin 150, New Ser C 18:1–112
Zhou C-F (2010a) A new eucryptodiran turtle from the Early Cretaceous Jiufotang Formation of western Liaoning, China. Zootaxa 2676:45–56
Zhou C-F (2010b) A second specimen of Manchurochelys manchoukuoensis Ento and Shikama, 1942 (Testudines: Eucryptodira) from the Early Cretaceous Yixian Formation of western Liaoning, China. Zootaxa 2534:57–66
Acknowledgements
The authors thank Fang Zheng (IVPP, Beijing) for access the studied specimens and comparative material in the IVPP collection in her care, Z.-M. Dong (IVPP, Beijing) and other colleagues who took part in the field work, Walter Joyce (Tübingen) and Ren Hirayama (Tokyo) for reviewing the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
This article is registered in Zoobank under urn:lsid:zoobank.org:pub:28836E5A-75F9-42C2-9B3B-DCB6E311A152
Rights and permissions
About this article
Cite this article
Tong, H., Brinkman, D. A new species of Sinemys (Testudines: Cryptodira: Sinemydidae) from the Early Cretaceous of Inner Mongolia, China. Palaeobio Palaeoenv 93, 355–366 (2013). https://doi.org/10.1007/s12549-012-0110-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-012-0110-8