
Abstract
New radiolarian taxa are described from the Lower and Middle Famennian of the Polar Urals and Volga–Urals Basin: three new genera Natgorella gen. nov., Spinoalius gen. nov., Diversiacus gen. nov., and three new species N. elegans sp. nov., N. hirsuta sp. nov., and S. melekessensis sp. nov. New taxa belong to two orders, Entactiniata and Cancelliata, of two classes of spherical radiolarians: Sphaerellaria and Spumellaria. Morphotypes of the outer skeletons of Spinoalius gen. nov. and Diversiacus gen. nov are shown to be convergently similar. A new Middle Famennian Natgorella hirsuta–Spinoalius melekessensis radiolarian ecozone is established.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
HISTORY OF STUDY
In the mid-20th century, Bykova (1955) studied the so-called “primitive” small foraminifera from the Lower and Middle Famennian of the Volga–Ural Basin (Figs. 1a–1p). Bykova (1955, p. 110) for the first time commented on a certain similarity of some Archaesphaera and Parathurammina with radiolarians: “in the shell structure they are closer to ancient radiolarians than to extant or Mesozoic–Cenozoic planktonic foraminifers.” The only difference between these organisms and radiolarians, according to Bykova, was that their skeleton was calcareous, whereas the radiolarian skeleton is siliceous.

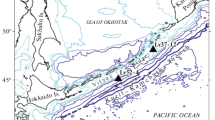
Distribution of Famennian radiolarians; explanations: (1) locality of the radiolarian ecozones: (I) Peri-Caspian Basin, (II) Polar Urals, Lemva Zone, (III) Timan–Pechora Basin, Saremboy–Lekkeyaga Borehole, (IV) western slope of the Middle Urals, (V) Pripyat Depression, (VI) Volga–Ural Basin (I, III Afanasieva, 2000; II–IV Afanasieva and Amon, 2011, 2012; V Kruchek and Nazarov, 1977; VI this paper); (2) “Uralian” assemblage (Bykova, 1955): (a–d) North Urals: (a) Petrunikha River, (b) Storozhevaya River, (c) Vishera River, (d) Yazva River; (e) South Urals, Sikaza River; (f–h) western Bashkortostan: (f) Elatminki (=Novokonstantinovka), (g) Kargaly River, (h) Tuimazy; (i–k) Tatarstan: (i) Shugurovo Village, (j) Kamskoe Ustie Village, (k) Verkhnii Uslon Village; (l–n) Samara Region: (l) Krasnovka Village, (m) Berezovka Village, (n) Syzran; (o, p) Saratov Region: (o) Pugachev, (p) Teplovka Village; (3) Karachaganak Fossil Reef; (4–10) paleobiogeographic scheme of the Famennian of the Russian Platform and the Greater Urals (Afanasieva, 2000; Afanasieva et al., 2011; Afanasieva and Amon, 2012): (4) low salinity sea; (5–10) normal salinity sea: (5) coastal region, (6) shallow inner shelf, (7) inner shelf, (8) outer shelf; (9) continental slope, (10) paleoocean seafloor; (11) active foldbelts; (11) land; (12a) margin of the Russian Platform, (12b) land.
At the same time, it is known that during lithification of the host rock, radiolarian skeletons can be recrystallized, resulting in the either replacement of the primary opal of the skeleton by other minerals (calcite, apatite, fluorite, garnets, pyrite, and gold), or only the spheres remain of the skeleton, and are often misidentified as small primitive foraminifers or algae. All the above suggests that some species from the genera Archaesphaera and Parathurammina can be considered as radiolarians (Braun and Amon, 1991; Afanasieva, 2000; Vishnevskaya and Sedaeva, 2002; Sedaeva and Vishnevskaya, 2002, 2008; Afanasieva and Amon, 2011, 2012). However, this opinion is not shared by all paleontologists.
Almost a quarter of a century later, undoubtedly Early Famennian radiolarians with a siliceous skeleton were found in the Eletsian Regional Stage of the Pripyat Depression (Figs. 1, V; 2, V; 3, V) and Nazarov studied them using an electron microscope (Kruchek and Nazarov, 1977; Nazarov, 1988). Later, Early Famennian radiolarians were studied (Afanasieva, 2000) on the Saremboy–Lekkeyaga Borehole in the Timan–Pechora Basin and in the north slope of the Peri-Caspian Basin (Figs. 1, I, III; 2, I, III; 3, I, III) and were found in the Polar Urals and in the Vilva River basin on the western slope of the Middle Urals (Figs. 1, II, IV; 2, II; 3, II, IV) (Afanasieva and Amon, 2011, 2012).
In the Volga–Ural Basin, no confirmed Famennian radiolarians have been recorded. At present, L.I. Kononova during the processing of conodont samples from the Middle Famennian Lebedyanian Regional Substage studied in the Melekess R-1 Borehole, found excellently preserved whole radiolarian shells (Figs. 1, VI; 2, VI; 3, VI).
This paper discusses the results of the study of the first-ever discovered true Middle Famennian radiolarians in the Volga–Ural Basin, and the description of the new Middle Famennian Natgorella hirsuta– Spinoalius melekessensis radiolarian ecozone.
RESULTS
The New Natgorella Hirsuta–Spinoalius Melekessensis Radiolarian Ecozone
A new association of the Middle Famennian radiolarians is established in the Melekess Depression of the Volga–Urals Basin. Radiolarians were found in the middle subformation of the Malaya Cheremshana Formation (interval 1940–1980 m) at two levels of the Melekess P-1 Borehole (Sample 7336, interval 1961.9–1963.9 m; Sample 7398, interval 1973.9–1974.7 m)Footnote 1 (Figs. 1, VI; 2, VI; 3, VI). The samples were disintegrated in 10% acetic acid solution using the standard procedure for isolating conodonts and radiolarians. Together with radiolarians in Sample 7398, Kononova discovered the conodonts Palmatolepis glabralepta Ziegler and Huddle, 1969. This subspecies ranges from the Late crepida Zone to the Late trachytera Zone. In Sample 7336, no conodonts were recorded. The age of the interval with radiolarians is emended based on a more diverse conodont assemblage identified in the lower part of the middle subformation (interval 1973.9–1976.5 m): Palmatolepis glabra lepta Ziegler and Hudd., P. glabra pectinata Ziegler, P. gracilis gracilis Branson and Mehl, P. utahensis Ziegler and Sandberg, P. perlobata helmsi Ziegler, P. perlobata sigmoidea Ziegler, P. perlobata schindewolfi Müller, P. inflexa Müller, P. rugosa grossi Ziegler, and P. marginifera marginifera Helms. Many of these species and subspecies have large stratigraphic ranges in the Famennian, but P. utahensis occurs only in the Late marginifera Zone, and P. inflexa ranges from the Late rhomboidea Zone to the Early marginifera Zone. This conodont association occurs almost 2 m below Sample 7398 with radiolarians, and is characteristic of the Early marginifera and Late marginifera zones, corresponding to the lower part of the Middle Famennian Lebedyanian regional stage (Rodionova et al., 1995).
Thus, the new Natgorella hirsuta–Spinoalius melekessensis ecozone characterizes part of the middle subformation of the Malaya Cheremshana Formation, which Kononova assigned based on conodonts to the Lebedyanian Regional Substage, which correlates with the Late marginifera and trachytera zones (Figs. 2, VI; 3, VI) (Fortunatova et al., 2016, 2018).

Famennian Sphaerellaria: (I–VI) ecozones of Famennian radiolarians: (I) Tetrentactinia barysphaera–Holoeciscus auceps, Peri-Caspian Basin, Karachaganak Fossil Reef; (II) Tetrentactinia barysphaera–Caspiaza spinifera, Polar Urals, Lemva Zone; (III) Tetrentactinia barysphaera–Retientactinosphaera magnifica, Timan–Pechora Basin, Saremboy–Lakkeyaga Borehole; (IV) Haplentactinia alekseevi–Haplentactinia vilvaensis, western slope of the Middle Urals, Vilva River; (V) Tetrentactinia barysphaera–Ceratoikiscum famennium, Pripyat Depression; (VI) Natgorella hirsuta–Spinoalius melekessensis, Volga–Ural Basin, Melekess Depression (I, III) Afanasieva, 2000; (II–IV) Afanasieva and Amon, 2011, 2012; (V) Kruchek and Nazarov, 1977; (VI) Afanasieva, this paper).
The taxonomic diversity of the new ecozone is characterized by the presence of six species of radiolarians (Figs. 2; 3): four species of three genera of latticed radiolarians of the class Spumellaria (Figs. 4a, 4b; Pl. 1) and two species of two genera of porous radiolarians of the class Sphaerellaria (Fig. 4c; Pl. 2).

Famennian Spumellaria: (I–VI) ecozones of Famennian radiolarians (see Fig. 2).

Radiolarians of the Melekess Depression: (a) Retientactinosphaera magnifica Afanasieva, 2011, specimen PIN, no. 5369/187-L1015 (scale bar = 131 μm); (b) Polyentactinia rudihispida Nazarov and Ormiston, 1993, specimen PIN, no. 5369/0132016-019 (scale bar = 71 μm); (c) Diversiacus zuraevi (Afanasieva and Amon, 2011), specimen PIN, no. 5369/329-L1024 (scale bar = 92 μm); Upper Devonian, Middle Famennian, Lebedyanian Regional Substage, Late marginifera and trachytera conodont zones; Melekess P-1 Borehole: (a, b) Sample 7336 (interval 1961.9–1963.9 m); (c) Sample 7398 (interval 1973.9–1974.7 m).
Typical species of the new assemblage include Natgorella elegans sp. nov. and N. hirsuta sp. nov. (pl. 1), as well as Spinoalius melekessensis sp. nov. (Pl. 2, figs. 1–4). The species Retientactinosphaera magnifica Afanasieva, 2011, Polyentactinia rudihispida Nazarov and Ormiston, 1993 and Diversiacus zuraevi (Afanasieva and Amon, 2011) are represented by a few specimens (Fig. 4).
P. rudihispida is found in all studied Lower and Middle Famennian radiolarian localities (Fig. 1, I–VI). However, this species first appeared in the Timan–Pechora Basin in the Frasnian (Fig. 3, I–VI) (Afanasieva, 2000). The species D. zuraevi (Fig. 2, II, VI) and R. magnifica (Fig. 3, III, VI) are characteristic only of the Famennian (Afanasieva, 2011; Afanasieva and Amon, 2011). At the same time these species are very important, as they are shared with the new radiolarian association of the Volga–Ural Basin (Middle Famennian, Lebedyanian regional substage) and older Lower Famennian ecozonesFootnote 2 (Afanasieva and Amon, 2011, 2012) (Figs. 2, 3):
Tetrentactinia barysphaera–Holoeciscus auceps, Peri-Caspian Basin of Kazakhstan, Karachaganak Fossil Reef, lower Famennian (Afanasieva, 1987, 2000);
Tetrentactinia barysphaera–Caspiaza spinifera, Polar Urals, Lemva Zone, Palnik-Yu River, triangularis and crepida conodont zones (Afanasieva, 2000; Afanasieva and Amon, 2011, 2012);
Tetrentactinia barysphaera–Retientactinosphaera magnifica, Timan–Pechora Basin, Saremboy–Lekkeyaga Borehole, Zapadno-Lekkeyaginskaya Borehole-65 (interval 2460–2467 m), triangularis and crepida conodont zones (Afanasieva, 2000; Afanasieva and Amon, 2011, 2012);
Haplentactinia alekseevi–Haplentactinia vilvaensis, western slope of the Middle Urals, Vilva River, triangularis and crepida conodont zones (Afanasieva and Amon, 2011, 2012);
Tetrentactinia barysphaera–Ceratoikiscum famennium, Pripyat Depression of Belarus, Eletsian Regional Substage; conodonts collected alongside radiolarians include: Palmatolepisglabra Ulrich and Bassler, P. minuta Branson and Mehl., P. aff. quadrantinodoslobata Sannemann, Prioniodina smithi Stauffer, Scutula venusta Sannemann (Kruchek and Nazarov, 1977; Nazarov, 1988), characterizing the rhomboidea Zone;
Natgorella hirsuta–Spinoalius melekessensis, Volga–Ural Basin, Melekess Depression, Lebedyanian Regional Substage, Late marginifera and trachytera conodont zones.
The radiolarian association of the new ecozone is surprisingly similar to that of the Haplentactinia alekseevi–Haplentactinia vilvaensis ecozone of the Middle Urals (Afanasieva and Amon, 2011, 2012). The similarity is observed in the presence of the same radiolarian morphotypes, in both cases represented mainly by lattice skeletons, and in the presence of the shared species Polyentactinia rudihispida, but the taxonomic composition of the ecozones is different.
Hereby I consider the new Middle Famennian radiolarian association as the Natgorella hirsuta–Spinoalius melekessensis ecozone, corresponding to the Middle Famennian Lebedyanian Regional Substage and correlating with the Late marginifera and trachytera conodont zones. The new radiolarian ecozone is recognized in the territory of the Volga–Ural Basin. The substantiation of a new ecozone of radiolarians is important for the Devonian stratigraphy of the region for a number of reasons. For instance, this is the first locality for true Middle Famennian radiolarians, the geochronology of which is supported by conodonts. In addition, the stratigraphic range of radiolarians is expanded from the triangularis to trachytera conodont zones (Figs. 2, 3). Knowledge on the taxonomic and morphological diversity of radiolarians in the Lower–Middle Famennian ecozones is enhanced. The assemblages differ in taxonomic composition, but at the same time are dominated by lattice representatives of the class Spumellaria and have the shared species Retientactinosphaera magnifica and Polyentactinia rudihispida.
Convergent Evolution
E.O. Schmidt introduced the term ‘convergent evolution’ in biology to indicate convergent or parallel emergence of isolated morphological characters or sets of characters, for taxa of various ranks (Mayr, 1970). Convergent evolution is “a process of irreversible acquisition of characters by members of two or more phyletic lineages in connection with their lifestyle, but not connected to their ancestry” (Paleontologiya…, 1995, p. 165).
All convergent characters are secondary developments that emerged independently in the evolutionary process in order to adapt to a specific environment. The convergent similarity of morphotypes is never deep, and involves only superficial morphological characters. Therefore in cases of convergent evolution even within the same taxonomic group, convergent characters are never exact copies of characters of other organisms. The resemblance of the superficial morphology of the compared taxa is usually partial and relative, rather than the morphology being entirely identical.
The study of the skeletons of the new radiolarian association revealed convergent morphological similarity of morphotypes of the outer appearance of skeletons of the new genera Spinoalius gen. nov. and Diversiacus gen. nov. (Fig. 5). The convergent similarity of these genera is based on a set of shared characters: (1) outer porous sphere; (2) six main spines, of which five spines are beam-like, and one is four-bladed or three-bladed (pl. 2), (3) rimming of the base of the beam-like spines by four petal-like pores (Pl. 2, figs. 1a, 1b; 7).

Convergent similarity of morphotypes of radiolarian outer skeletons: (a) Spinoalius, (b) Diversiacus.
Differences are observed in the inner structure of the skeletons, which are characters of the subfamily level (Afanasieva, 2000; Afanasieva et al., 2005; Afanasieva and Amon, 2006): the genus Spinoalius is characterized by the presence of only one outer sphere (subfamily Entactiniinae) (Fig. 5a, Pl. 2, fig. 2a), whereas the genus Diversiacus differs in having two spheres, outer and inner (subfamily Bientactinosphaerinae) (Fig. 5b; Pl. 2, figs. 5, 6, 8).
Thus, in the two subfamilies Entactiniinae and Bientactinosphaerinae, independently similar, geochronologically parallel outer skeletons arose, which may indicate a similar habitat, and similar direction of natural selection. This difference in the morphology of skeletons justifies the recognition of two new genera: Spinoalius gen. nov. and Diversiacus gen. nov.
MATERIAL
Collections of Famennian radiolarians, nos. 5312 and 5369, are housed in the A.A. Borissiak Paleontological Institute (PIN). Radiolarian taxonomy follows Afanasieva and Amon (2003, 2006) and Afanasieva et al. (2005). Measurements of the radiolarian skeleton and the morphometric analysis of the absolute and relative values of the Paleozoic radiolarian shells followed a standard method (Fig. 6) (Afanasieva, 2000; Afanasieva et al., 2005; Afanasieva and Amon, 2006).

Major absolute and relative morphometric parameters of skeletons of Paleozoic spherical radiolarians (Afanasieva, 2000; Afanasieva et al., 2005; Afanasieva and Amon, 2006): (D) outer sphere diameter, (Ds) primary sphere diameter, (dp) diameter of pores of the outer sphere, (ds) diameter of pore of the primary sphere, (wp) inter-pore width of the outer sphere, (ws) inter-pore width of the primary sphere, (tw) wall thickness of the outer sphere, (ts) wall thickness of the primary sphere, (L) length of main spines, (wL) width of the base of the main spines, (ht) height of spinules.
-
SYSTEMATIC PALEONTOLOGY
-
PHYLUM RADIOLARIA
-
SUPERCLASS POLYCYSTINA
-
CLASS SPUMELLARIA
-
Order Cancelliata
-
Family Haplentactiniidae Nazarov in Nazarov and Popov, 1980
-
Subfamily Pseudorotasphaerinae Noble, 1994
-
Genus Natgorella Afanasieva gen. nov.
Etymology. In honor of the paleontologist and stratigrapher Nataliya V. Goreva; feminine.
Type species. Natgorella hirsuta sp. nov.
Diagnosis. Shell spherical, with two lattice shells, formed by entangled rounded bars. Rays of inner six-ray spicule continue as six main rod-like spines with rounded cross-section. Surface of bars of outer spheres with two types of spinules: conical and thin rod-like, bifurcating, in approximately upper third of length.
Species composition. Type species and N. elegans sp. nov.
Comparison. This genus differs from the genera Cancellosphaera Afanasieva, 2000, Russirad Afanasieva, 2000 and Arcoclathrata Afanasieva and Amon, 2016, which have a latticed outer shell, a porous inner shell and six main 3-bladed spines, in the two lattice spheres, and six main rod-like spines. The new genus is distinguished from the genus Retientactinosphaera Afanasieva, 2011, which has only one three-bladed spine along with six main rod-like spines.
-
Natgorella hirsuta Afanasieva sp. nov.
-
Plate 1, figs. 4–6
Etymology. From the Latin hirsutus (hirsute).

Explanation of Plate 1
Upper Devonian, Middle Famennian, Lebedyanian Regional Substage, Late marginifera and trachytera conodont zones; Volga–Ural Basin, Melekess Depression, Melekess P-1 Borehole: Sample 7336, interval 1961.9–1963.9 m (1–3, 5, 6) and Sample 7398, interval 1973.9–1974.7 m (4).
Figs. 1–3.Natgorella elegans sp. nov.: (1) holotype PIN, no. 5369/328-M1014: (1a) fragment (scale bar = 42 μm), (1b) general view (scale bar = 83 μm), (1c) fragment (scale bar = 41 μm); (2) specimen PIN, no. 5369/328-M1020 (scale bar = 87 μm); (3) specimen PIN, no. 5369/328-M1021: (3a) general view (scale bar = 86 μm), (3b) fragment (scale bar = 29 μm).
Figs. 4–6.N. hirsuta sp. nov.: (4) specimen PIN, no. 5369/327-M1023: (4a) general view (scale bar = 98 μm), (4b) fragment (scale bar = 49 μm); (5) holotype PIN, no. 5369/327-M1016: (5a) general view (scale bar = 136 μm), (5b) fragment (scale bar = 59 μm); (6) specimen PIN, no. 5369/327-M1017: (6a) fragment (scale bar = 44 μm), (6b) general view (scale bar = 90 μm), (6c) fragment (scale bar = 33 μm).
Holotype. PIN, no. 5369/327-M; Volga–Ural Basin, Melekess Depression, Melekess P-1 Borehole (interval 1973.9–1974.7 m); Upper Devonian, Middle Famennian, Lebedyanian Regional Substage, Late marginifera and trachytera conodont zones.
Description. The shell is spherical, very large (D = 231–248 μm), composed of two lattice shells with six main rod-like spines and numerous spinules. The outer sphere is composed of thin entangled bars (D/tw = 34.4–42.8), forming very large (D/dp = 8.3–10.2) polygonal pores of approximately the same size. The inner sphere is large (D/Ds = 3.1–3.3), relatively thin-walled (Ds/ts = 23.3–25.7) and is 30.3–31.9% of the diameter of the outer spheres. It is formed by entangled bars surrounding very large (Ds/ds = 5.9) polygonal pores. Rays of the six-ray spicules are represented by six relatively short (L/D = 0.7–1.0) and thin (L/wL = 7.8–13.0) main rod-like spines. The bars of the outer sphere possess small spinules of two kinds: (1) few, but relatively long (D/ht1 = 15.4–20.1) conical spinules; (2) very numerous and very long (D/ht2 = 2.3–3.4) rod-like thin spinules, double-bifurcated at approximately mid-length. The spinules are 29.1–44.4% of the diameter of the outer sphere, and the double bifurcation of numerous thin spinules (Pl. 1, fig. 5b) gives the species a shaggy, scruffy appearance, which is reflected in the species name.
Dimensions in µm and ratios (Fig. 6):
Specimen no. | D | dp | tw | Ds | ds | ts | L | wL | ht1 | ht2 |
|---|---|---|---|---|---|---|---|---|---|---|
5369/327-M1016 (holotype) | 248 | 21 | 5 | – | – | – | 245 | 20 | 14 | 110 |
5369/327-M1017 | 241 | 29 | 6 | 77 | 13 | 3 | 221 | 17 | 12 | 70 |
5369/327-M1023 | 231 | 23 | 6 | 70 | – | 3 | 155 | 20 | 15 | 73 |
Specimen no. | D/tw | D/dp | L/D | L/wL | D/ht1 | D/ht2 | D/Ds | Ds/ds | Ds/ts |
|---|---|---|---|---|---|---|---|---|---|
5369/327-M1016 (holotype) | 42.8 | 10.2 | 1.0 | 12.3 | 17.7 | 2.3 | – | – | – |
5369/327-M1017 | 34.4 | 8.3 | 0.9 | 13.0 | 20.1 | 3.4 | 3.1 | 5.9 | 25.7 |
5369/327-M1023 | 38.5 | 10.0 | 0.7 | 7.8 | 15.4 | 3.2 | 3.3 | – | 23.3 |
Comparison. This species is distinguished from N. elegans sp. nov., characterized by few thin rod-like spinules only occasionally bifurcating in the upper third of their length and numerous, very small conical spines on the surface of the inner sphere, by the presence of very numerous thin rod-like spinules that are frequently double bifurcated.
Material. Three specimens from the type locality.
-
Natgorella elegans Afanasieva sp. nov.
-
Plate 1, figs. 1–3
Etymology. From the Latin elegans (elegant).
Holotype. PIN, no. 5369/328-M1014; Volga–Ural Basin, Melekess Depression, Melekess P-1 Borehole (interval 1961.9–1963.9 m); Upper Devonian, Middle Famennian, Lebedyanian Regional Substage, Late marginifera and trachytera conodont zones.
Description. The shell is spherical, very large (D = 206–219 μm), consisting of two lattice spheres with six main rod-like spines and numerous spinules. The outer sphere is composed of thin entangled bars (D/tw =36.5–42.0), forming very large (D/dp = 7.5–10.3) polygonal pores of approximately the same size. The inner sphere is large (D/Ds = 2.8–3.2), relatively thick-walled (Ds/ts = 22.7–25.0), and is 31.1–35.9% of the outer sphere diameter. It was formed by entangled bars surrounding very large (Ds/ds = 5.7–6.8) polygonal pores. The surface of the bars of the inner sphere show numerous small conical spinules from 2 to 5 μm long (Pl. 1, fig. 3b). Six main rod-like spines are continuations of the inner six-ray spicule: the spines are relatively short (L/D = 0.5–0.9) and thin (L/wL = 10.2–13.4). The surface of the bars of the outer sphere possesses two kinds of spinules: (1) very numerous long (D/ht1 = 12.2–13.7) conical spinules; (2) few, but very long (D/ht2 = 3.8–4.3) rod-like thin spinules, sometimes bifurcating in approximately the upper third of their length.
Dimensions in µm and ratios (Fig. 6):
Specimen no. | D | dp | tw | Ds | ds | ts | L | wL | ht1 | ht2 |
|---|---|---|---|---|---|---|---|---|---|---|
5369/328-M1014 (holotype) | 210 | 28 | 5 | 75 | 11 | 3 | 201 | 15 | 17 | 50 |
5369/328-M1020 | 219 | 23 | 5 | 68 | 12 | 3 | – | 11 | 18 | 58 |
5369/328-M1021 | 206 | 20 | 5 | 74 | 13 | 3 | 102 | 10 | 15 | 48 |
Specimen no. | D/tw | D/dp | L/D | L/wL | D/ht1 | D/ht2 | D/Ds | Ds/ds | Ds/ts |
|---|---|---|---|---|---|---|---|---|---|
5369/328-M1014 (holotype) | 42.0 | 7.5 | 0.9 | 13.4 | 12.4 | 4.2 | 2.8 | 6.8 | 25.0 |
5369/328-M1020 | 36.5 | 9.5 | – | – | 12.2 | 3.8 | 3.2 | 5.7 | 22.7 |
5369/327-M1021 | 41.2 | 10.3 | 0.5 | 10.2 | 13.7 | 4.3 | 2.8 | 5.7 | 24.7 |
Material. Three specimens from the type locality.
-
CLASS SPHAERELLARIA
-
Order Entactiniata
-
Family Entactiniidae Riedel, 1967
-
Subfamily Entactiniinae Riedel, 1967
-
Genus Spinoalius Afanasieva gen. nov.
Etymology. From the Latin spina (spine) and alius (other); masculine.
Type species. Spinoalius melekessensis sp. nov.
Diagnosis. Shell spherical, porous. Rays of the inner six-ray spicules continue into six main spines: one dominant four-bladed spine with a  -like cross-section of the base and five rod-liked spines with rounded sections of the base. Six petaloids are present near the base of beam-like spines (Pl. 2, figs. 1a, 1b).
-like cross-section of the base and five rod-liked spines with rounded sections of the base. Six petaloids are present near the base of beam-like spines (Pl. 2, figs. 1a, 1b).
Species composition. Type species.
Comparison. This genus differs from the most similar genus Borisella Afanasieva, 2000, with a porous skeleton possessing six main beam-like spines, in the presence of a single dominant four-bladed spine with a  -like cross-section and five rod-like spines.
-like cross-section and five rod-like spines.
-
Spinoalius melekessensis Afanasieva sp. nov.
-
Plate 2, figs. 1–4
Etymology. After the locality in the Melekess Depression in the Volga–Ural Basin.
Holotype. PIN, no. 5369/329-M1027; Volga–Ural Basin, Melekess Depression, Melekess P-1 Borehole, interval 1973.9 –1974.7 m; Upper Devonian, Middle Famennian, Lebedyanian Regional Substage, Late marginifera and trachytera conodont zones.

Explanation of Plate 2
Upper Devonian: (1–4, 7) Middle Famennian, Lebedyanian Regional Substage, Late marginifera and trachytera conodont zones; Volga–Ural Basin, Melekess Depression, Melekess P-1 Borehole, Sample 7398 (interval 1973.9–1974.7 m); (5, 6, 8) Lower Famennian Substage, Palmatolepis triangularis and P. crepida conodont zones; Polar Urals, Lemva Zone, Palnik-Yu River: Sample 101/599 (5), Sample 101/594 (6, 8).
Figs. 1–4.Spinoalius melekessensis sp. nov.: (1) specimen PIN, no. 5369/329-M1025: (1a) fragment (scale bar = 40 μm), (1b) general view (scale bar = 98 μm), (1c) fragment (scale bar = 40 μm); (2) specimen PIN, no. 5369/329-M1022: (2a) fragment (scale bar = 38 μm), (2b) general view (scale bar = 96 μm); (3) holotype PIN, no. 5369/329-M1027, general view (scale bar = 88 μm); (4) specimen PIN, no. 5369/329-M1026 (scale bar = 88 μm).
Figs. 5–8 .Diversiacus zuraevi (Afanasieva and Amon, 2011): (5) specimen PIN, no. 5312/160-15958 (scale bar = 79 μm); (6) holotype PIN, no. 5312/160-15718 (scale bar = 98 μm); (7) specimen PIN, no. 5369/160-M1024 (scale bar = 100 μm): (7a) fragment (scale bar = 27 μm), (7b) general view (scale bar = 92 μm); (8) specimen PIN, no. 5312/160-15732 (scale bar = 67 μm).
Description. The shell is spherical, very large (D = 214–252 μm), composed of one porous sphere. The outer sphere is thick-walled (D/tw = 19.4–25.6) with large, rounded pores approximately of the same diameter (D/dp = 12.2–14.4). The inter-pore wall are acute and relatively narrow (dp/wp = 4.5–5.7). The rays of the inner six-ray spicule continue as six main spines: (1) one dominant four-bladed spine with a  ‑like cross-section, relatively long (L1/D = 0.7–1.1) and thin (L1/wL1 = 7.3–11.7); (2) five rod-like spines with a rounded cross-section, the spines are short (L2/D = 0.4–0.6) and relatively thick (L2/wL2 = 3.9–6.3). Six petal-like pores are present near the base of the beam-like spines (Pl. 2, figs. 1a, 1b). The inter-pore wall possess ornamentation in the shape of very high acute spinules, sometimes bifurcating distally (Pl. 2, figs. 3, 4).
‑like cross-section, relatively long (L1/D = 0.7–1.1) and thin (L1/wL1 = 7.3–11.7); (2) five rod-like spines with a rounded cross-section, the spines are short (L2/D = 0.4–0.6) and relatively thick (L2/wL2 = 3.9–6.3). Six petal-like pores are present near the base of the beam-like spines (Pl. 2, figs. 1a, 1b). The inter-pore wall possess ornamentation in the shape of very high acute spinules, sometimes bifurcating distally (Pl. 2, figs. 3, 4).
Dimensions in mm and ratios (Fig. 6):
Specimen no. | D | dp | wp | tw | L1 | wL1 | L2 | wL2 | ht |
|---|---|---|---|---|---|---|---|---|---|
5369/329-M1027 (holotype) | 230 | 16 | 3 | 9 | 363 | 31 | 86 | 22 | 54 |
5369/329-M1022 | 231 | 19 | 4 | 11 | 220 | 30 | 136 | 28 | 52 |
5369/329-M1025 | 252 | 18 | 4 | 13 | – | 32 | 125 | 20 | 30 |
5369/329-M1026 | 214 | 17 | 3 | 9 | – | 27 | 102 | 18 | 40 |
Specimen no. | D/dp | D/tw | dp/wp | L1/D | L1/wL1 | L2/D | L2/wL2 | D/ht |
|---|---|---|---|---|---|---|---|---|
5369/329-M1027 (holotype) | 14.4 | 25.6 | 5.3 | 1.6 | 11.7 | 0.4 | 3.9 | 4.3 |
5369/329-M1022 | 12.2 | 21.0 | 4.8 | 1.0 | 7.3 | 0.6 | 4.9 | 4.4 |
5369/329-M1025 | 14.0 | 19.4 | 4.5 | – | – | 0.5 | 6.3 | 8.4 |
5369/329-M1026 | 12.6 | 23.8 | 5.7 | – | – | 0.5 | 5.7 | 5.4 |
Remarks. This species differs from species of the most similar genus Borisella Afanasieva, 2000, which have six rod-like spines, in the presence of one dominant four-bladed spine with a  -like cross-section and five rod-like spines.
-like cross-section and five rod-like spines.
Material. Four specimens from the type locality.
-
Subfamily Bientactinosphaerinae Afanasieva, 1999
-
Genus Diversiacus Afanasieva gen. nov.
Etymology. From the Latin diversus (diverse) and acus (needle); masculine.
Type species. Bientactinosphaera zuraevi Afanasieva and Amon, 2011 from the Upper Devonian, Lower Famennian, triangularis and crepida conodont zones of the Polar Urals, Lebedyanian Regional Substage, Late marginifera and trachytera conodont zones; Volga–Ural Basin.
Diagnosis. Shell spherical with two porous spheres. Inner frame in the shape of a six-rayed spicule (Pl. 2, fig. 8), the rays of which continue into six main spines. One main spine is three-faceted, with a  -like cross section and lobe-like cross-section of each blade. Five main spines rod-like with rounded cross-section. Six petal-shaped pores near base of rod-like spines (pl. 2, fig. 7).
-like cross section and lobe-like cross-section of each blade. Five main spines rod-like with rounded cross-section. Six petal-shaped pores near base of rod-like spines (pl. 2, fig. 7).
Species composition. Type species.
Comparison. This genus is distinguished from Bientactinosphaera Afanasieva, 2000, which has six main three-bladed spines, by the presence of one main three-bladed spine and five main rod-like spines.
Remarks. Originally, the species D. zuraevi was described in the genus Bientactinosphaera, based on the presence of two porous spheres (Afanasieva and Amon, 2011). However, skeleton morphology allowed the recognition of a new genus Diversiacus gen. nov., with D. zuraevi as the type species. In addition, a convergent similarity is established for the external morphology of the skeletons of the new genera Diversiacus and Spinoalius (Fig. 5). The skeletons of these genera have an outer porous sphere with five main rod-like spines and a one faceted main spine (four-bladed or three-bladed) (pl. 2). Note that the skeletons of the genus Spinoalius have (Fig. 5a; Pl. 2, fig. 2a) only one outer sphere (subfamily Entactiniinae), whereas Diversiacus differs (Fig. 5b; Pl. 2, figs. 5, 6, 8) in having two spheres: outer and inner (subfamily Bientactinosphaerinae).
Notes
For a detailed lithological and stratigraphic description of the Melekess P-1 Borehole and paleontological substantiation see Fortunatova et al. (2018).
“Ecozone is a complex zone, beds in which contains a fossil assemblage reflecting an ecological association during lifetime or the taphonomic features of their oryctocenosis” (Stratigraphic …, 2006, Article VII, 4g).
REFERENCES
Afanasieva, M.S., Late Paleozoic radiolarians of the Karachaganak Field and their facies affinity, in Stratigrafiya i paleontologiya Prikaspiiskoi vpadiny (Stratigraphy and Paleontology of the Caspian Basin), Il’in, V.D and Zamilatskaya, T.K, Eds., Moscow: Vses. Nauchno-Issled. Geol. Neft. Inst., 1987, pp. 26–47.
Afanasieva, M.S., Atlas radiolyarii paleozoya Russkoi platformy (Atlas of Paleozoic Radiolaria of the Russian Platform), Moscow: Nauchnyi Mir, 2000.
Afanasieva, M.S., Revision of the Genus Entactinosphaera Foreman, 1963 and the New Genus Retientactinosphaera gen. nov. (Paleozoic Radiolaria), Paleontol. J., 2011, vol. 45, no. 2, pp. 117–129.
Afanasieva, M.S. and Amon, E.O., A New Classification of the Radiolaria, Paleontol. J., 2003, vol. 37, no. 6, pp. 630–645.
Afanasieva, M.S. and Amon, E.O., Radiolyarii (Radiolaria), Moscow: Paleontol. Inst. Ross. Akad. Nauk, 2006.
Afanasieva, M.S. and Amon, E.O., Devonian radiolarians of Russia, Paleontol. J., 2011, vol. 45, no. 11, pp. 1313–1532.
Afanasieva, M.S. and Amon E.O., Stratigrafiya i paleobiogeografiya devonskikh radiiolarii Rossii (Stratigraphy and Paleobiogeography of Devonian Radiolarians of Russia), Moscow: Paleontol. Inst. Ross. Akad. Nauk, 2012.
Afanasieva, M.S., Amon, E.O., Agarkov, Yu.V., and Boltovskoy, D.S., Radiolarians in the geological record, Paleontol. J., 2005, vol. 39, no. Suppl. 3, pp. 135–392.
Braun, A. and Amon, E.O., Fluoritisierte Radiolarien aus Kieselkalk-Bänken des Mittel-Viseums (Unterkarbon) des Rheinischen Schiefergebirges (Deutschland), Paläontol. Z., 1991, vol. 65, nos. 1–2, pp. 25–33.
Bykova, E.V., Devonian Foraminifera and Radiolaria of the Volga–Ural Region and the Central Devonian Field and their Stratigraphic Significance, in Foraminifery, radiolyarii i ostrakody devona Volgo-Ural’skoi oblasti (Foraminifers, Radiolarians, and Ostracods of the Devonian of the Volga–Ural Region), Tr. Vses. Neft. Geol.-Razved. Inst., Nov. Ser., no. 87, Leningrad: Gostoptekhizdat, 1955, pp. 5–190.
Fortunatova, N.K., Zaitseva, E.L., Bushueva, M.A., Shvets-Teneta-Guriy, A.G., Baranova, A.V., Kononova, L.I., Rakhimova, Ye.V., Mikheeva, A.I., Oleneva, N.V., and Mushin, I.A., Stratigrafiya verkhnego devona Volgo-Ural’skogo subregiona (po materialam aktualizatsii stratigraficheskikh skhem (Upper Devonian Stratigraphy of the Volga–Ural Subregion: Materials on the Actualization of Stratigraphic Schemes), Moscow: Vseross. Nauchno-Issled. Geol. Neft. Inst., 2016.
Fortunatova, N.K., Zaitseva, E.L., Kononova, L.I., Baranova, A.V., Bushuyeva, M.A., Mikheeva, A.I., Afanasieva, M.S., and Obukhovskaya, T.G., Lithology-Facies and biostratigraphic characterization of the Upper Devonian Beds in the Reference Melekess R-1 Borehole (Melekess Depression, Volga–Urals Subregion), Byull. Mosk. Obshch. Ispyt. Prir., Otd. Geol., 2018, vol. 93, nos. 5–6, pp. 3–49.
Kruchek, S.A. and Nazarov, B.B., Upper Devonian Radiolaria from the Russian Platform, Dokl. Akad. Nauk SSSR, 1977, vol. 237, no. 6, pp. 1445–1448.
Mayr, E. Populations, Species, and Evolution, Cambridge, Mass.: Belknap Press of Harvard University Press, 1970.
Nazarov, B.B., Radiolyarii paleozoya. Prakticheskoe rukovodstvo po mikrofaune SSSR (Paleozoic Radiolaria: Practical Manual on the Microfauna of the USSR), Leningrad: Nedra, 1988, vol. 2.
Paleontologiya i paleoekologiya: Slovar’-spravochnik (Paleontology and Paleoecology: Dictionary-Handbook), Makridin, V.P. and Barskov, I.S., Eds., Moscow: Nedra, 1995.
Rodionova, G.D., Umnova, V.T., Kononova, L.I., Ovnatanova, N.S., Rzhonsnitskaya, M.A., and Fedorova, T.I., Devon Voronezhskoi anteklizy i Moskovskoi sineklizy (Devonian of the Voronezh Anteclise and Moscow Syneclise), Moscow: Tsentr. Reg. Geol. Tsentr, 1995.
Sedaeva, G.M. and Vishnevskaya, V.S., Revision of the Superfamily Parathuramminoidae: foraminifers or radiolarians, Vestn. Mosk. Gos. Univ., Ser. 4, Geol.. 2002, no. 5, pp. 15–19.
Sedaeva, K.M. and Vishnevskaya, V.S., On the nature of problematic microfossils from the Upper Devonian and Lower Carboniferous, Nov. Paleontol. Stratigr., Prilozh. Geol. Geofiz., 2008, vol. 49, nos. 10–11, p. 152.
Stratigraficheskii kodeks Rossii. Izd. 3 (Stratigraphic Code of Russia, Third Edition), St. Petersburg: VSEGEI, 2006.
Vishnevskaya, V.S. and Sedaeva, K.M., A revision of some foraminiferal taxa of the order Parathuramminida and discussion of foraminiferal and radiolarian evolution, Paleontol. J., 2002, vol. 36, no. 6, pp. 581–590.
ACKNOWLEDGMENTS
I am sincerely grateful to L.I. Kononova for providing material of radiolarians and for consultation on the age of the host rocks, to A.S. Alekseev and V.S. Vishnevskaya for fruitful discussion of the paper and constructive recommendations, to A.F. Bannikov and N.M. Kuzmina for valuable advice and comments when preparing the manuscript for publication, and to S.V. Nikolaeva for translating this paper into English.
Funding
The work was done in accordance with the government assignment to the A.A. Borissiak Paleontological Institute of the Russian Academy of Sciences and supported by the Research Program of the Presidium of the Russian Academy of Sciences “The Evolution of the Organic World. The Role and Influence of Planetary Processes.”
Author information
Authors and Affiliations
Corresponding author
Additional information
Translated by S. Nikolaeva
Rights and permissions
About this article

Cite this article
Afanasieva, M.S. New Taxa and New Middle Famennian Natgorella hirsuta–Spinoalius melekessensis Radiolarian Ecozone of the Volga–Ural Basin. Paleontol. J. 53, 435–446 (2019). https://doi.org/10.1134/S0031030119050022
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S0031030119050022