
Abstract
Sauropterygian vertebrate remains from the Ladinian (Middle Triassic) of the proximity of Calasparra (Murcia, Spain) are described here. They represent the first documented Triassic vertebrate remains found in the Province of Murcia, and the first in the Subbetic domain of the Betic Cordillera (Southeastern Spain). These new remains consist of incomplete vertebrae, some fragmented neural spine apophyses, fragments of transverse processes, a partial thoracic vertebrae mould and one isolated thoracic vertebral centrum, seemingly of a single specimen. Their features permit to refer them to the suborder Eusauropterygia. These fossils have been found in the middle part of the upper member of the Cehegín Formation. The biostratigraphical framework of the Cehegín Formation consists of a relatively abundant fossil content that allowed its attribution to the uppermost Fassanian (Lower Ladinian). The sedimentary features and the palaeontological content of the site are typical of an epicontinental platform.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Sauropterygia was a diverse group of diapsid marine reptiles that ranged from the late Early Triassic to the end of the Cretaceous. Their Triassic radiation was restricted to near-shore habitats of the Tethys Ocean, connecting epicontinental seas that primarily concerned shallow marine forms such as Placodontia, Pachypleurosauria, Nothosauria, and the open marine Pistosauria. The global fossil record of Triassic sauropterygians is relatively well known in Europe, China North America, the North of Africa and the Middle East (Rieppel 2000). In Spain, most of the marine vertebrate remains, documented to date from the Triassic, consist of isolated elements, attributed to indeterminate nothosaurs, pachypleurosaurs and placodonts (e.g., Kuhn-Schnyder 1966; Lapparent 1966; Westphal 1976; Vía Boada et al. 1977; Sanz 1980, 1983; Alafont 1992, 1999; Sanz and Moratalla 1993; Alafont and Sanz 1996; Rieppel and Hagdorn 1998; Rubio et al. 2003; Quesada and Aguera González 2005; Quesada et al. 2009; Fortuny et al. 2011; Miguel Chaves et al. 2014a, b, 2015, 2016, 2017; Berrocal-Casero and Castaninha 2015; Berrocal-Casero et al. 2018). These specimens were collected in several Triassic sites in central and Northern Spain, but scarce references about Triassic vertebrate remains are known from the South of Spain (Alafont 1992; Niemeyer 2002; Reolid et al. 2013), all of them from the Province of Jaén. Alafont (1992) and Reolid et al. (2013) described some isolated vertebrate remains of nothosaurs and placodonts in the Tabular Cover. Niemeyer (2002) studied the Ladinian Muschelkalk facies from the Hornos-Siles Formation in the Prebetic domain (External Zones of the Betic Cordillera) and identified disarticulated fossil remains of marine reptiles corresponding to prolacertiform and the Superorder Sauropterygia (Pachypleurosauridae, Nothosauridae and Cyamodontidae). The aim of the present paper is to report the first Triassic Sauropterygia remains from the Province of Murcia that also supposes the first record of vertebrate remains documented from the Triassic of the Subbetic domain of the External Zones of the Betic Cordillera. These remains were recovered and positioned with biostratigraphical control by the Pérez-Valera brothers in the 90 s during different fieldworks, carried out before the completion of their PhD theses.
Regional geology and stratigraphy
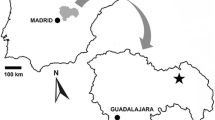
The Betic Cordillera constitutes the northern branch of the Betic-Rif orogen, the westernmost orogen of the Alpine, peri-Mediterranean folds belts, geographically situated in the southern part of the Iberian Peninsula (Fig. 1). Classically, the Betic Cordillera is divided in two domains: the Internal and the External Zones (e.g., Martín Algarra and Vera 2004). The Internal Zones are composed of three tectonic complexes, from base to top: Nevado-Filábride, Alpujárride and Maláguide, limited by tectonic contacts and with a wide variety of metamorphic features that show the complex, polyphasic history of their emplacement (García-Dueñas et al. 1992) during the westward migration of the Mesomediterranean (Alborán) terrain (Balanyá and García-Dueñas 1987; Platt et al. 1995). The External Zones are composed of Triassic to Miocene, non-metamorphic rocks resulting of folding and thrusting of the South-Iberian Palaeomargin sediments as the result of the collision with the Alborán domain, in Miocene times (Sanz de Galdeano 1990). The External Zones, in turn, can be subdivided in two important sectors (García-Hernández et al. 1980): the more proximal Prebetic sector, which, next to the Hercinian antepais foreland shows a predominance of coastal shallow marine facies; and the more distal Subbetic sector, more abundant in hemipelagic facies.

Modified from Pérez-López and Pérez-Valera (2012)
Geographical location and geological setting of the South-Iberian Triassic outcrops within the tectonic context and structural units of the Betic Cordillera (Southeastern Spain). CL Calasparra site
The Triassic stratigraphy of the External Zones shows the typical features of the trilogy of Germanic facies, with detritical (siliciclastic) and evaporitic units (Buntsandstein and Keuper facies), below and above a shallow marine intercalation of Muschelkalk facies. The term South-Iberian Triassic is used to refer the Triassic units in the External Zones (e.g. Pérez-López and Pérez-Valera 2007) and shows diverse formal lithostratigraphic units: (1) the Arroyo Molinos Formation (Pérez-Valera 2005a, b) (Buntsandstein facies), (2) the Siles and Cehegín Formations (Pérez-Valera and Pérez-López 2008), that refer to the proximal (Prebetic) and distal (Subbetic) regions of the South-Iberian Paleomargin, respectively (Muschelkalk facies), and 3) the Jaén Keuper Group (Pérez-López 1998) (Keuper facies). The here-presented fossil bone remains are located in the Cehegín Formation (Muschelkalk facies), near the town of Calasparra, in the Province of Murcia (Southeastern Spain) (Fig. 1).
The Cehegín Formation is formed by a succession of dark limestones with clay, marly intervals and presence of bioclastic beds (tempestites) in the upper part (Pérez-López and Pérez-Valera 2007, 2012). Two members can be distinguished in this unit, representing a complete, third-order, transgressive–regressive cycle. The lower member is deposited above the siliciclastic, fluvial deposits of the Arroyo Molinos Formation. In it, three laminated, dark-gray limestone beds can be recognized. This member has been interpreted as a carbonate ramp developed under a transgressive stage (Pérez-López et al. 2005). The upper member is deposited over a hard-ground developed on the maximum flooding surface and is constituted by a succession of marly limestone, thin bedded limestone, bioclastic beds and green, ochre clays, with dolomites and gypsum layers in the upper transition to the Keuper facies, showing the progradation of coastal and shallow-water deposit over the ramp of the lower member (Pérez-López and Pérez-Valera 2012). All the sedimentary features are typical of an epicontinental platform (Pérez-Valera 2005a, b; Pérez-Valera and Pérez-López 2008). The vertebrate remains of the nothosaurids occur in the middle of the upper member of the Cehegín Formation together with ammonoids, nautiloids, bivalves and brachiopods, in the Calasparra section (Fig. 2). The bones appear in a centimetric thick alternation of marls and thin-bedded marly limestone situated over a 20-cm-thick, bioclastic key level that marks the end of a regressive parasequence. Therefore, the fossiliferous interval can represent the beginning of the next parasequence, as interpreted by Pérez-Valera and Pérez-López (2008) and Baeza-Calatarrá et al. (2018).

The relatively abundant fossil content of the Cehegín Formation has allowed a consistent biostratigraphical framework in which three ammonite biochronozones have been distinguished (Fig. 2), all belonging to the Ladinian stage (Pérez-Valera 2005a, b, 2015; Pérez-Valera et al. 2011). This age agrees with the previous data provided by bivalves, conodonts, and nautiloids, being these fossils also characteristic of the Ladinian (Márquez-Aliaga et al. 2001; Plasencia et al. 2007; Pérez-Valera et al. 2017). Specifically, the levels containing the vertebrate remains are situated in the Gevanites epigonus Zone (Fig. 2), and they can be attributed to the uppermost Fassanian (Lower Ladinian).
Material
The vertebrate remains (Sauropterygia: Eosauropterygia: Eusauropterygia) studied here consist in some fragments of spine apophyses (UCM-CLV25-26), some fragments of transverse processes (UCM-CLV27-28), two incomplete dorsal vertebrae with their incomplete neural arches (UCM-CLV29-32), the mould of one of them (UCM-CLV33), and one isolated vertebral centrum (UCM-CLV34). Some incomplete vertebrae preserve the centrum and the neural arch, but they are not articulated. The spine apophysis (UCM-CL2V25) seems to belong to the fragmented vertebra UCM-CLV29-30, because the fragments were recovered together, although the material is very incomplete. At the same time, the neural spine UCM-CL2V26 seem to belong to the remain UCM-CLV31-32. These fragmented vertebrae with their corresponding neural arches and broken neural spines have been recovered, in pieces, but together (Figs. 3, 4). The vertebrae described here correspond to thoracic vertebrae because of the absence of the articular facets for the ribs in the vertebral centra.

Incomplete vertebrae remains from the Middle Triassic of Calasparra (Murcia, Southeastern Spain). a Neural apophyses UCM-CLV25 in: Cranial (a1), lateral (a2) and caudal (a3) view. b Neural apophyses UCM-CLV26 in: Cranial (b1) and lateral (b2) views. c, d Fragments of transverse processes UCM-CLV27 (c), UCM-CLV28 (d). e Part of the neural arch (UCM-CLV29) with its corresponding vertebral centrum (UCM-CLV30) in: anterior (e1), posterior (e2), lateral (e3) view. Vertebral centrum (UCM-CLV30) in: dorsal (e4), and ventral (e5) view. f Part of the neural arch (UCM-CLV31) with its corresponding vertebral centrum (UCM-CLV32) in: cranial (f1), caudal (f2) and lateral (f3) view. Vertebral centrum (UCM-CLV32) in: dorsal (f4), and ventral (f5) view. gr groove, pl prespinal lamina

Vertebrae remains from the Middle Triassic of Calasparra (Murcia, Southeastern Spain). a Mould UCM-CLV33 of the vertebral centrum and part of the neural arch corresponding to the bone remains UCM-CLV31-CLV32. b Isolated vertebral centrum UCM-CLV34 in: cranial (b1), dorsal (b2), ventral (b3), right lateral (b4) and left lateral (b5) views. gr groove, fo foramina
The neural spine UCM-CL2V25 is subrectangular in outline, very high, narrow and its apex is ornamented by longitudinal grooves (Fig. 3a1–a2). It is 8.20 cm in high, 1.8 cm in length (high/length ratio: 4.55) and its maximum width is 1.05 cm (in the apical zone) being slightly mediolaterally inflated in its apical region (Fig. 3a1–a3). The edge is sharp in cranial view (Fig. 3a) and shows a depression in caudal view (Fig. 3a3). The spine apophysis CL2V26 is long and flattened (Fig. 3b), but differs from UCM-CL2V25 because it is shorter and it is not inflated or wider in the apical zone. It is 6 cm in high, 1.8 cm in length (high/length ratio: 3.33) and its width corresponds to 1.25 cm. This spine shows sharp edges. A fragment of a broken prespinal lamina of 1 mm width is observed in the cranial area of the neuropophysis (Fig. 3b). Other fragments correspond to the transverse processes (UCM-CL2V27-28) (Fig. 3c, d).
The vertebrae UCM-CL2V29-30 preserve the vertebral centrum (UCM-CL2V29) and part of the neural arch (UCM-CL2V30) (Fig. 3e). The vertebral centrum is platicoelus. The centra of the vertebra present a characteristic cruciform-suture facet (“kreuzform”) in dorsal view (sensu Rieppel 1994) (Fig. 3e4, e5). This centrum is ventrally constricted in lateral view. It is 2.3 cm in high, 1.73 cm in with and 1.34 cm in length. The centrum of dorsal vertebrae displays a distinct, shallow arcuate groove crossing the right lateral site diagonally from the posterior dorsal core to the ventral margin (Fig. 3e3). The neural arch is broken and incomplete, and only a part around the neural canal is preserved, with part of prezygapophysis (Fig. 3e1). Dorsally, the neural canal is very big, ovoid, taller than wider and narrower at the upper part, with the convex part up.
Another incomplete dorsal vertebra UCM-CL2V31-32 shows similar features than previously mentioned (Fig. 3f). The vertebral centrum UCM-CL2V32 (2.37 cm in length and 2.4 cm width), is platicoelus to slightly amphicoelous and laterally constricted (Fig. 3f4, f5). The shallow arcuate groove crossing the right lateral site diagonally from the posterior dorsal core to the ventral margin (Fig. 3f3) and the cruciform-suture facet also can be observed in this centrum (Fig. 3f4). The height of the incomplete neural arch is around 2.6 cm. One transverse process is preserved (Fig. 3f1, f2). It is moderately elongated and projecting. The zygantrium area is not preserved. The pre- and postzygapophyses are broad. The articulate facets on the pre- and postzygapophyses are almost horizontally oriented and show considerable erosion (Fig. 3f1, f2). Dorsally, the neural canal is big, ovoid, taller than wider and narrower at the upper part. The mould of this vertebra (UCM-CLV33) shows the position of the vertebral centrum and the neural arch and part of the neural spine (Fig. 4a).
Other isolated vertebral centrum was recovered with the remains previously mentioned (Fig. 4b). The vertebral centrum UCM-CLV34 is badly preserved, it is 2.32 cm in length 2.62 cm in width and, it is constricted laterally. The kreuzform suture facet can be observed in dorsal view (Fig. 4b2). In this centrum, on the groove crossing the right lateral site diagonally from the posterior dorsal core to the ventral margin, two very small foramina can be observed (Fig. 4b4).
Discussion
The dorsal neural arches still associated with their respective unfused centra and the close association of the fragments of transverse processes suggest that the vertebrae were not totally disarticulated. Basal sauropterygians present a paedomorphic postcranial skeleton, including the lack of fusion of the neurocentral suture (Rieppel 1994, 2000), being the centrum and the neural arch usually separated. This dissociation reveals a broadened facet formed by the dorsal surface of the centrum to receive the pedicles of the neural arch with a cruciform or butterfly-shaped platform. This structure is apomorphic for Eosauropterygia (Rieppel 1994). Thus, the features of the here studied remains allow to assign them to Eosauropterygia.
Rieppel (1994; Rieppel and Hagdorn 1998) included Pachypleurosauroidea and Eusauropterygia (Nothosauroidea + Pistosauroidea) in the order Eosauropterygia. The genus Corosaurus was firstly remitted to Eosauropterygia (Rieppel 1994), and subsequently reconsidered in Pistosauridea (Rieppel and Hagdorn 1998). Pachypleurosauroidea vertebrae use to present dimensions of around 4–15 mm according to Alafont (1992, 1999), being not compatible with the remains here studied from Calasparra (Fig. 3). The vertebrae of Corosaurus alcovensis Case, 1936 are amphicoelous (Storrs 1991; Rieppel and Hagdorn 1998), being not consistent with the platicoelus vertebral centra from Calasparra. In some Eusauropterygia as in Simosaurus, the dorsal surface of the centra is copiously widened providing a characteristic butterfly-shaped platform for the articulation with the neural arch (Rieppel 1994). Posterolaterally, the articular platform carries triangular areas on both sides with a conspicuous pattern of pitting, and similar pitted areas, but much smaller and less well defined, are located in the anterolateral corner of its articular surface (Rieppel 1994). This butterfly platform is different to the kreuzform-shaped platform of the vertebrae here studied (Figs. 3e4, f4; 4b2), so Simosaurus can be also discarded. Moreover, in Simosaurus, the neural arches of the dorsal region are characteristic by the development of infrapre- and infrapostzygapophyses (von Huene 1952: 167) as accessory articulations in addition to the zygosphene–zygantrum articulation. These structures cannot be identified in the arches from the proximity of Calasparra due to their preservation (Fig. 3f1, f2).
It is characteristic the height of the neural spines of the vertebrae here studied (Fig. 3a, b). The presence of high neural spines is indicated by Schmidt (1986) as a parallelism between the Plesiosaurs, Pistosaurus, the Nothosaurus species of the Upper Muschelkalk and Ceresiosaurus; while in other nothosaurids such as Nothosaurus giganteus and Lariosaurus, these processes are short. In Nothosaurus, high dorsal neural spines were considered a synapomorphy of a clade composed of Nothosaurus mirabilis Münster, 1834, N. haasi Rieppel and Mazin, 1997 and N. tchernovi Haas, 1980 (Rieppel 2000). The neural arches of the nothosaurs present well-developed bipartite zygosphene, whereas its development is smaller in other eusauropterygians (Rieppel and Wild 1996; Rieppel 1997). This structure cannot be recognized in the neural arches from Calasparra, but the remains are too broken to confirm their absence. The dorsal neural spines of the pistosaur Bobosaurus forojuliensis Dalla Vecchia, 2006 from the Alpine Basin of Italy are higher than those of N. mirabilis figured by Rieppel (2000: 12). The remains here studied show some affinities with the Pistosaur B. forojuliensis from the Carnian of the Alpine Basin (Dalla Vecchia 2006, 2017): (1) the neural spine is high and shows a thin prespinal lamina (feature also characteristic in some plesiosaurs). This broken prespinal lamina can be observed in UCM-CLV26 (Fig. 3b1). (2) In Bobosaurus, the apex shows grooves and ridges at the top of the lateral face, giving striated aspect, as can be observed also in UCM-CLV25 (Fig. 3a2). (3) The centrum is spool shaped with constricted lateral sides, as the centra here studied (Figs. 3e5, f5; 4b). (4) The centra of dorsal vertebrae display a distinct, shallow arcuate groove crossing the right lateral site diagonally from the posterior dorsal core to the mid ventral margin (Figs. 3e3, f3; 4b4). (5) The foramina subcentralia are very small, being present only in some dorsal vertebral centra (Fig. 4b4).
In pistosaurs, the ventral facets of the centra bear a low median ridge separating a pair of oval nutritive foramina (foramina subcentralia) at about midcentrum (Sues 1987). According to some authors, the subcentral foramina is also present in some nothosaurs (Sanz 1983), but the presence of subcentral foramina, is a diagnostic feature of the Pistosauria (Pistosauridae and Plesiosauria) for Rieppel (2000), and is absent in those of Nothosaurus according to Diedrich (2013).
Comparing to other Spanish Sauropterygia remains, Alafont (1992: 62–63) mentioned the presence of diagonal grooves with small foramina in some vertebral centra from Bienservida-Villarodrigo (Jaén, South of Spain) and classified these remains as Nothosauria indet. Some spine apophyses studied by the same author are very high, but no evidences of prespinal lamina were mentioned by this author. In the same region, Niemeyer (2002) described smaller vertebrae assigned to Pachypleosauria and Nothosauridae, but without constriction or grooves in the centra. The vertebral centra here studied differs from the material form the Ladinian of the Riba de Santiuste (Guadalajara, Central Spain) (Berrocal-Casero et al. 2018) because these vertebral centra do not show spool shape, lateral constriction, any groove crossing the centra or any subcentral foramina on the vertebrae centra. Comparing to other Spanish nothosaur remains identified in Central Spain, Nothosaurus cf. giganteus has a lower neural spine that is a characteristic feature of this species (Rieppel and Wild 1996; de Miguel Chaves et al. 2016). The neural arches from Canales de Molina (Guadalajara) classified as Eosauropterygia indet. are characterized by the presence of a very low spine and they do not show prespinal lamina. de Miguel Chaves et al. (2016) do not mention grooves or foramina in the vertebral centra.
The fragmentary material here studied is not enough to allow, for the moment, a more precise systematic identification, but these remains are compatible with Eusauropterygia, a suborder including the Nothosauroidae and Pistosauroidae (Rieppel 2000), and they seem to show more affinity with the latter family. Thought the results presented here are still preliminary, a detailed taxonomic study of these new remains, and a deeper exploration of the sites can provide new information about these unknown Triassic vertebrates from the Betic Cordillera.
Conclusion
The Lower Ladinian (Middle Triassic) site of Calasparra (Murcia, Spain), located in Muschelkalk Facies belonging to the Cehegín Formation, has provided the first record of Triassic sauropterygian of the Murcia region. These are also the first vertebrate remains in the Subbetic domain of the External Zones of the Betic Cordillera (Southeastern Spain), adding new data to the palaeontological record from the Triassic of this area. The presently studied vertebral remains from Calasparra here studied are very different comparing to the Middle Triassic vertebrae remains known from the Iberian record, being characterized by the presence of very high neural apophyses with prespinal lamina, the presence of an arcuate groove crossing the right lateral site diagonally in the vertebral centra, and the presence of small subcentral foramina in some of these centra. These remains are compatible with Eusauropterygia: a suborder including the Nothosauroidae and Pistosauroidae (Rieppel 2000), showing more affinity to Pistosauridae. This discovery adds new information about the diversity and geographical distribution of Sauropterygia in the Middle Triassic, indicating that it was greater than previously believed until the moment.
References
Alafont, L.S. 1992. Notosaurios y Placodontos (Reptilia) del Triásico Medio de Bienservida Villarrodrigo. Instituto de Estudios albacetenses de la Excma. Diputación de Albacete, Albacete.
Alafont, L.S. 1999. Reptiles del Triásico de Castilla-La Mancha. In La huella del pasado. Fósiles de Castilla-La Mancha, eds. E. Aguirre and I. Rábano, 143–159. Toledo: Serie Patrimonio Histórico Arqueología.
Alafont, L.S., and J.L. Sanz. 1996. Un nuevo Sauropterigio (Reptilia) en el Triásico de la Sierra de Prades (Tarragona). Cuadernos de Geología Ibérica 20: 313–329.
Baeza-Calatarrá, J.F., F. Pérez-Valera, and J.A. Pérez-Valera. 2018. The oldest post-Paleozoic (Ladinian, Triassic) brachiopods record from the Betic Range (SE Spain). Acta Palaeontologica Polonica 63: 71–85.
Balanyá, J.C., and V. García-Dueñas. 1987. Grandes fallas de contracción y de extensión implicadas en el contacto entre los dominios de Alborán y Sudibérico en el arco de Gibraltar. Geogaceta 1: 19–21.
Berrocal-Casero, M., and R. Castaninha. 2015. Nuevos hallazgos de vertebrados triásicos en el anticlinal de Riba de Santiuste (Guadalajara, España). In Current Trends in Paleontology and Evolution (XIII EJIP Conference Proceedings), eds. L. Domingo, S. Domingo, O. Fesharaki, B. García Yelo, A.R. Gómez Cano, V. Hernández-Ballarín, D. Hontecillas, J.L. Cantalapiedra, P. López Guerrero, A. Oliver, J. Pelegrín, M. Pérez de los Ríos, M. Ríos, O. Sanisidro, and A. Valenciano, pp. 1–58. Madrid.
Berrocal-Casero, M., J. Audije-Gil, R. Castaninha, J.A. Pérez-Valera, V.F. Santos, and M. Segura. 2018. New discoveries of vertebrate remains from the Triassic of Riba de Santiuste, Guadalajara (Spain). Proceedings of the Geologists’ Association 129: 526–541.
Case, E.C. 1936. A nothosaur from the Triassic of Wyoming. University of Michigan. Contribution from the Museum of Paleontology 5: 1–36.
Dalla Vecchia, F.M. 2006. A new sauropterygian reptile with plesiosaurian affinity from the Late Triassic of Italy. Rivista Italiana di Paleontologia e Stratigrafia 112: 207–225.
Dalla Vecchia, F.M. 2017. Comments on the skeletal anatomy of the Triassic reptile Bobosaurus forojuliensis (Sauropterygia, Pistosauroidea). Gortania (Geologia, Paleontologia, Paletnologia) 38: 39–45.
Diedrich, C.G. 2013. The oldest “Subaquatic Flying” reptile in the World - Pistosaurus longaevus Meyer, 1839 (Sauropterygia) from the Middle Triassic of Europe. New Mexico Museum of Natural History and Science 61: 169–215.
Fortuny, J., A. Bolet, A.G. Sellés, J. Cartanyà, and À. Galobart. 2011. New insights on the Permian and Triassic vertebrates from the Iberian Peninsula with emphasis on the Pyrenean and Catalonian basins. Journal of Iberian Geology 37: 65–86.
García-Dueñas, V., J.C. Balanyá, and J.M. Martínez-Martínez. 1992. Miocene extensional detachments in the outcropping basement of the northern Alboran Basin (Betics) and their tectonic implications. Geo-Marine Letters 12: 88–95.
García-Hernández, M., A.C. López-Garrido, P. Rivas, C. Sanz de Galdeano, and J.A. Vera. 1980. Mesozoic palaeogeographic evolution of the External Zones of the Betic Cordillera. Geologie in Mijnbouw 59: 155–168.
Haas, G. 1980. Ein Nothosaurier-Schädel aus dem Muschelkalk des Wadi Ramon (Negev, Israel). Annalen des Naturhistorischen Museums in Wien 83: 119–125.
Huene, F. von. 1952. Skelett und Verwandschaft von Simosaurus. Palaeontographica (A: Paläozoologie, Stratigraphie) 102: 163–182.
Kuhn-Schnyder, E. 1966. Über Wirbeltier-Reste aus dem Muschelkalk der Provinz Teruel (Spanien). Oberrheinische geologische Abhandlungen 15: 99–106.
Lapparent, A.F. 1966. Nouveaux gisements de reptiles mésozoiques en Espagne. Notas y Comunicaciones del Instituto Geológico y Minero de España 15: 99–106.
Márquez-Aliaga, A., A. Goy, and J.J. Gómez. 2001. Bivalvos del Ladiniense en la región de Riópar (Albacete, España). Publicaciones del seminario de Paleontología de Zaragoza 5: 144–152.
Martín Algarra, A., and J.A. Vera. 2004. La Cordillera Bética y las Baleares en el contexto del Mediterráneo Occidental. In Geología de España, ed. J.A. Vera, pp. 352–354. Madrid: SGE, IGME.
Miguel Chaves, C. de, Pérez-García, A., Ortega, F., Sánchez-Chillón, B., Quesada, J., and J.L., Sanz. 2014a. Systematic implications of a sauropterygian skull from the Upper Triassic of Guadalajara (Spain). In Program and Abstracts, 74th Annual Meeting of the Society of Vertebrate Paleontology, Berlin 2014, eds. E. Maxwell, and J. Miller-Camp, p. 118.
Miguel Chaves, C. de, Pérez-García, A., Quesada, J., and F. Ortega. 2014b. Dientes atribuidos a sauropterigios del Triásico Superior de El Atance (Guadalajara, España). Libro de resúmenes de las XXX Jornadas de Paleontología de la Sociedad Española de Paleontología. ¡Fundamental! 24: 143–144.
Miguel Chaves, C. de, A.P. García, A. Cobos, R. Royo-Torres, F. Ortega, and L. Alcalá. 2015. A diverse Late Triassic tetrapod fauna from Manzanera (Teruel, Spain). Geobios 48: 479–490.
Miguel Chaves, C. de, S. García-Gil, F. Ortega, J.L. Sanz, and A. Pérez-García. 2016. First Triassic tetrapod (Sauropterygia, Nothosauridae) from Castilla y León: evidence of an unknown taxon for the Spanish record. Journal of Iberian Geology 42: 29–38.
Miguel Chaves, C. de, F. Ortega, and A. Pérez-García. 2017. The eosauropterygian fossils from the Middle Triassic of Canales de Molina (Guadalajara, Spain). Journal of Iberian Geology 43: 129–138.
Münster, G. 1834. Vorläufige Nachricht über einige neue Reptilien im Muschelkalke von Baiern. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde 1834: 521–527.
Niemeyer, J. 2002. Invertebraten und Vertebraten aus dem Muschelkalk von Siles (Jaén), Spanien. Münstersche Forschungen zur Geologie und Paläontologie 94: 99.
Pérez-López, A. 1998. Epicontinental Triassic of the Southern Iberian Continental Margin (Betic Cordillera, Spain). In Epicontinental Triassic, eds. G.H. Bachmann and I. Lerche. Zentralblatt für Geologie und Paläontologie (I: Allgemeine, angewandte, regionale und historische Geologie) 1998(9–10): 1009–1031.
Pérez-López, A., L. Márquez, and F. Pérez-Valera. 2005. A foraminiferal assemblage as a bioevent marker of the main Ladinian transgressive stage in the Betic Cordillera, Southern Spain. Palaeogeography, Palaeoclimatology, Palaeoecology 224: 217–231.
Pérez-López, A., and F. Pérez-Valera. 2007. Palaeogeography, facies and nomenclature of the Triassic units in the different domains of the Betic Cordillera (S Spain). Palaeogeography, Palaeoclimatology, Palaeoecology 254: 606–626.
Pérez-López, A., and F. Pérez-Valera. 2012. Tempestite facies models for the epicontinental Triassic carbonates of the Betic Cordillera (southern Spain). Sedimentology 59(2): 646–678.
Pérez-Valera, F. 2005a. Estratigrafía y tectónica del Triásico Sudibérico en el sector oriental de la Cordillera Bética. Unpublished PhD Thesis. Granada: University of Granada.
Pérez-Valera, J.A. 2005b. Ammonoideos y bioestratigrafía del Triásico Medio (Anisiense superior-Ladiniense) en la sección de Calasparra (sector oriental de la Cordillera Bética, Murcia, España). Coloquios de Paleontología 55: 125–161.
Pérez-Valera, F., and A. Pérez-López. 2008. Stratigraphy and sedimentology of Muschelkalk carbonates of the Southern Iberian Continental Palaeomargin (Siles and Cehegín Formations, Southern Spain). Facies 54: 61–87.
Pérez-Valera, J.A. 2015. Ammonoideos y bioestratigrafía del Triásico medio (Ladiniense) del sector oriental de la Cordillera Bética. Unpublished PhD Thesis. Madrid: Complutense University.
Pérez-Valera, J.A., Goy, A., Pérez-Valera, F., and A., Pérez-López. 2011. Ammonoid biostratigraphy of Muschelkalk carbonates (Ladinian) of the South-Iberian Triassic (Betic External Zones, S. Spain). In Livret des Résumés du Sixième Réunion du Groupe Marocain du Permien et du Trias, Tétouan, ed. A. Maate, 23–24. Tétouan: University of Tétouan.
Pérez-Valera, J.A., F. Barroso-Barcenilla, A. Goy, and F. Pérez-Valera. 2017. Nautiloids from the Muschekalk facies of the Southiberian Triassic (southern Spain). Journal of Systematic Palaeontology 15: 171–191.
Plasencia, P., A. Márquez-Aliaga, F. Pérez-Valera, J.A. Pérez-Valera, A. Goy, J. López-Gómez, L. Márquez, and A. Pérez-López. 2007. Registro paleontológico del Triásico Medio de Calasparra (Murcia). In Libro de Resúmenes de las XXIII Jornadas Sociedad Española de Paleontología, eds. J.C. Braga, A. Checa, and M. Company, 187–188. Murcia: Caravaca de la Cruz.
Platt, J., S. Allerton, A. Kirker, and E. Platzman. 1995. Origin of the western Subbetic arc (South Spain): palaeomagnetic and structural evidence. Journal of Structural Geology 17: 765–775.
Quesada, J.M., and S. Aguera González. 2005. Descripción del primer ejemplar de Ceresiosaurus (Reptilia: Sauropterygia), hallado en la Península Ibérica en el Ladiniense (Triásico Medio) de Mont–ral–Alcover (Tarragona). Estudios Geológicos 61: 247–269.
Quesada, J., A. Pérez-García, F. Ortega, F. Escaso, and B. Sánchez-Chillón. 2009. Simosaurus (Nothosauria) fossil site: a new Spanish window to the Upper Triassic. Journal of Vertebrate Paleontology 29: 168.
Reolid, M., F. Pérez-Valera, M.J. Benton, and J. Reolid. 2013. Marine flooding event in continental Triassic facies identified by a nothosaur and placodont bonebed (South Iberian Paleomargin). Facies 60: 277–293.
Rieppel, O. 1994. Osteology of Simosaurus gaillardoti and the relationships of stem-group Sauropterygia. Fieldiana: Geology 28: 1–85.
Rieppel, O. 1997. Part II: Sauropterygia. In Ancient marine reptiles, eds. J.M. Callaway and E.L. Nicholls, 107–119. San Diego: Academic Press.
Rieppel, O. 2000. Handbook of Paleoherpetology. Part 12A. Sauropterygia I. Placodontia, Pachypleurosauria, Nothosauria, Pistosauria. München: F. Pfeil.
Rieppel, O., and R. Wild. 1996. A revision of the genus Nothosaurus (Reptilia: Sauropterygia) from the Germanic Triassic, with comments on the status of Conchiosaurus clavatus. Fieldiana (Geology) 34: 1–82.
Rieppel, O., and J.M. Mazin. 1997. Speciation along rifting continental margins: a new nothosaur from the Negev (Israel). Comptes rendus de l’Académie des Sciences Série IIA, Sciences de la Terre et des planètes 326: 991–997.
Rieppel, O., and H. Hagdorn. 1998. Fossil reptiles from the Spanish Muschelkalk (Mont-ral and Alcover, Province Tarragona). Historical Biology 13: 77–97.
Rubio, C., J.B. Díez Ferrer, J. Ferrer, J.A. Gámez Vintaned, L.M. Sender, and H. Zianni. 2003. La Comarca de Gúdar—Javalambre da a conocer su rico patrimonio geológico y paleontológico. Naturaleza Aragonesa 11: 21–34.
Sanz, J.L. 1980. Los notosaurios españoles. Análisis filogenético y sistemático de los sauropterigios triásicos. PhD Thesis. Madrid: Universidad Complutense.
Sanz, J.L. 1983. Los Notosaurios (Reptilia, Sauropterygia) españoles. Estudios Geológicos 39: 193–215.
Sanz, J.L., and J.J. Moratalla. 1993. Triassic reptile faunas from Spain. In Evolution, Ecology and Biogeography of the Triassic Reptiles, eds. J.M. Mazin and G. Pinna, 153–164. Paleontologia Lombarda.
Sanz de Galdeano, C. 1990. Geologic evolution of the Betic Cordillera in the Western Mediterranean, Miocene to the present. Tectonophysics 172: 107–119.
Schmidt, S., 1986. Lokomotion und Lebenweise der Nothosaurier (Diapsida, Sauropterygia). Unpublished PhD Thesis. Tübingen: Universität Tübingen.
Storrs, W. 1991. Anatomy and relationships of Corosaurus alcovensis (Diapsida: Sauropterygia) and the Triassic Alcova Limestone of Wyoming. Bulletin of the Peabody Museum of Natural History 44: 1–151.
Sues, H.D. 1987. Postcranial skeleton of Pistosaururs and interrelationships of the Sauropterygia (Diapsida). Zoological Journal of the Linnean Society 90: 109–131.
Vía Boada, L., J.F. Villalta, and M. Esteban Cerdá. 1977. Paleontología y paleoecología de los yacimientos fosilíferos del Muschelkalk superior entre Alcover y Montral (Montañas de Prades, provincia de Tarragona). Cuadernos Geología Ibérica 4: 247–256.
Westphal, F. 1976. The dermal armor of some Triassic placodont reptiles. In Morphology and biology of reptiles, eds. A. Bellairs and C.B. Cox, 31–41. London: Academic Press.
Acknowledgements
We are very grateful to Dr. Fernando Barroso-Barcenilla from the Universidad de Alcalá (Spain) for his help and support. To Dr. Francis Hirsch (Naruto University of Education, Japan) for reviewing the English of the text. We gratefully acknowledge the comments from Dr. Silvio Renesto Claudio from the Università Degli Studi Dell’Insubria (Italy) and the PalZ editors (Mike Reich and Hans-Dieter Sues) that have helped to improve the manuscript. To the Research Project CGL2015-66604 from the Spanish Ministerio de Economía y Competitividad (Economy and Competiveness Ministry).
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Hans-Dieter Sues.
Rights and permissions
About this article

Cite this article
Pérez-Valera, J.A., Berrocal-Casero, M. & Pérez-Valera, F. First Triassic tetrapod (Eusauropterygia) in the Triassic of the Subbetic domain of the Betic Cordillera (Southeastern Spain). PalZ 94, 343–352 (2020). https://doi.org/10.1007/s12542-019-00500-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12542-019-00500-y