
Abstract
Purpose
Abundant isolated remains of vertebrate fossils were recovered in 1980 in Middle Triassic strata (Muschelkalk Facies) of Canales de Molina (Guadalajara, Spain). However, none of them was figured or described in detail. The careful study of these fossils is performed here.
Methods
The eosauropterygian remains from Canales de Molina are described and figured in detail, and compared with the Iberian record of Eosauropterygia.
Results
The study of these elements allows us to identify abundant eosauropterygian remains, including vertebrae, teeth, femora, and a partial humerus. This humerus presents a combination of characters traditionally considered as exclusive affinity of the genus Nothosaurus. Other relatively large elements from this site are also compatible with the bones of this genus. Isolated teeth and a neural arch are identified as belonging to indeterminate nothosaurs. The other elements studied here are poorly informative, and can be only assigned to indeterminate members of Eosauropterygia of small to medium size.
Conclusions
The presence of Nothosaurus or a related form is confirmed in Canales de Molina. Other fossils from the site do not allow to assess if more taxa are present.
Resumen
Objetivo
Abundantes restos aislados de vertebrados fueron recuperados en 1980 en los niveles del Triásico Medio (Muschelkalk Facies) de Canales de Molina (Guadalajara, España). Sin embargo, ninguno de ellos se ha figurado o descrito detalladamente hasta ahora. El estudio detallado de estos fósiles es llevado a cabo aquí.
Métodos
Los restos de eosauropterigios de Canales de Molina son descritos y figurados en detalle, y comparados con el registro ibérico de Eosauropterygia.
Resultados
El estudio de estos elementos nos permite identificar abundantes restos de eosauropterigios, incluyendo vértebras, dientes, fragmentos de fémures y parte de un húmero. Este húmero presenta una combinación de caracteres tradicionalmente considerada exclusiva del género Nothosaurus. Otros elementos relativamente grandes de este yacimiento son también compatibles con este género. Dientes aislados y un arco neural se identifican como pertenecientes a notosaurios indeterminados. Los otros elementos estudiados aquí son poco informativos, y sólo pueden ser asignados a miembros indeterminados de Eosauropterygia de pequeño a mediano tamaño.
Conclusiones
La presencia de Nothosaurus o de una forma cercanamente emparentada es confirmada en Canales de Molina. Otros fósiles del yacimiento no permiten asegurar si hay otros taxones presentes.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Sauropterygia was a successful clade of diapsid aquatic reptiles, known from the Early Triassic (Olenekian) to the end of the Cretaceous (Storrs 1991; Bakker 1993; Jiang et al. 2014). This clade experienced a rapid diversification from the Middle to the early Late Triassic (mainly during the Anisian and Carnian), reaching a high disparity, present until the end of the Triassic (Stubbs and Benton 2016). The global fossil record of Triassic sauropterygians is relatively well-known. Abundant and complete specimens have been recovered mostly in Europe and China, but also in North America, the north of Africa and the Middle East (Rieppel 2000; Bardet et al. 2014).
Remains of several sauropterygian clades have been recovered in the Triassic strata of the Iberian Peninsula, including cyamodontoid and non-cyamodontoid placodonts, pachypleurosaurs, nothosauroids and pistosauroids (e.g., Fortuny et al. 2011; Reolid et al. 2013; Miguel Chaves et al. 2015). Most of these remains consist of isolated and fragmentary elements, which usually prevent a precise taxonomic attribution. These Triassic sauropterygian fossils have been found in Muschelkalk and Keuper Facies, in the provinces of Barcelona (Almela and Llopis Lladó 1947; Sanz 1983), Lleida (Fortuny et al. 2011), and Tarragona (Via Boada et al. 1977; Sanz 1983; Martínez-Delclòs 1995; Alafont and Sanz 1996; Rieppel and Hagdorn 1998; Quesada and Aguera González 2005), in Catalonia; Huesca (Ferrando 1912; Sanz 1976), and Teruel (Kuhn-Schnyder 1966; Lapparent 1966; Rubio et al. 2003; Miguel Chaves et al. 2015), in Aragón; Albacete (Lapparent 1966; Westphal 1975; Sanz 1991; Alafont, 1992), Ciudad Real (Pinna 1990), Cuenca (Lapparent 1966; Sanz 1983; López-Gómez 1985; Pinna 1990; Alafont 1999), and Guadalajara (Alafont 1999; Quesada et al. 2009), in Castilla-La Mancha; Soria (Miguel Chaves et al. 2016), in Castilla y León; Jaén (Sanz 1991; Alafont 1992; Niemeyer 2002; Reolid et al. 2013), in Andalusia; and Balearic Islands (Bauzá Rullán 1955).
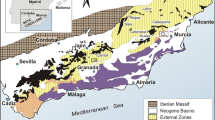
Vertebrate remains identified as fishes and reptiles were discovered in the summer of 1980 in the vicinity of the village of Canales de Molina (Guadalajara, Castilla-La Mancha; Castilian branch of the Iberian Ranges, Fig. 1), in Middle Triassic strata exposed in the margins of the road N-211. This finding was notified by Alférez et al. (1983), but none of these remains were described or figured so far. The fossiliferous levels from which these fossils come correspond to dolomitic limestones of the upper Muschelkalk Facies, belonging to the Royuela Dolostones, Marls and Limestones Formation (Alférez et al. 1983). These levels were deposited during the Ladinian (Middle Triassic), in a coastal environment (Márquez-Aliaga and García-Gil 1991; Vargas et al. 2009; Miguel Chaves et al. 2016; Sánchez-Moya et al. 2016).

Geographical situation of the Ladinian (Middle Triassic, upper Muschelkalk Facies) fossil site of Canales de Molina (Guadalajara Province, Spain)
The available information about the eosauropterygians remains from Canales de Molina was so far extremely limited. All the remains belonging to this clade have been analysed, the most relevant of them being presented and discussed here. Despite of the isolated nature of these elements, the collection of eosauropterygians presented here represent one of the largest known so far in Central Spain. Thus, teeth, appendicular elements and vertebral remains are studied and discussed. The presence of Nothosaurus or a related-form is recognized here.
Institutional abbreviations: MGM, Museo Geominero, Instituto Geológico y Minero de España, Madrid, Spain.
2 Systematic palaeontology
Sauropsida Huxley 1864
Sauropterygia Owen 1860
Eosauropterygia Rieppel 1994
Eosauropterygia indet.
(Figure 2a–s3)

Some of the eosauropterygian vertebral remains from the Muschelkalk of Canales de Molina (Guadalajara Province, Spain). Eosauropterygian centra in dorsal (a-d1, e1, f1, g, h1, i-l1, m1), ventral (d2, e2, m4), anterior (f2, h2, l2, m2) and lateral (m3) views (from a to m: MGM-468T, MGM-469T, MGM-470T, MGM-471T, MGM-472T, MGM-473T, MGM-371T, MGM-474T, MGM-475T, MGM-376T, MGM-374T, MGM-362T and MGM-293T). Partial eosauropterygian vertebrae, MGM-287T (n) and MGM-476T (o). Eosauropterygian neural arches in anterior (q MGM-290T; r MGM-477T) and posterior views (p MGM-289T; s MGM-478T). t MGM-324T, nothosaur neural arch in anterior (t1), posterior (t2) and lateral (t3 and t4) views. Anatomical abbreviations: ns neural spine, poz postzygapophysis, prz prezygapophysis, tp transverse process, zph zygosphene, zyg zygantrum. Scale bar equals 20 mm
Material: More than 130 isolated specimens, corresponding to abundant vertebral remains (most of them being centra, but isolated neural arches and some vertebrae preserving the fused centrum and arch being also recognized) (Fig. 2a–s), and some appendicular elements (Fig. 3).

Femoral eosauropterygian remains from the Muschelkalk of Canales de Molina (Guadalajara Province, Spain). a MGM-325T, a partial femur including the diaphysis and the distal region. b MGM-326T, diaphysis of a femur. c MGM-327T, a complete right femur in ventral (c1), dorsal (c3) and lateral (c2 and c4) views. d MGM-331T, proximal portion of femur in ventral (d1), dorsal (d3) and lateral (d2 and d4) views. Anatomical abbreviation: it internal trochanter. Scale bar equals 20 mm
Locality and horizon: Canales de Molina (Guadalajara, Castilla-La Mancha, Spain). Royuela Dolostones, Marls and Limestones Formation, Ladinian (Middle Triassic, upper Muschelkalk Facies) (Márquez-Aliaga and García-Gil 1991; Vargas et al. 2009; Miguel Chaves et al. 2016; Sánchez-Moya et al. 2016).
Description: More than a hundred isolated vertebral centra have been recovered, ranging from 0.3 to 2.5 cm in length (Fig. 2a–m). In dorsal view, all the centra present a butterfly-shaped articular surface (sensu Rieppel 1994) for the neural arch (Fig. 2a–d1, f1, g, h1, i–l1, m1). They are non-notochordal. Some of the centra are platicoelous (e.g. Fig. 2m, o), whereas others are slightly amphicoelous (e.g. Fig. 2n). In addition, some of them are strongly constrained dorsoventrally, resulting in relatively flattened elements (Fig. 2f–h). Lateral and ventral constriction are also present in some centra (e.g., Fig. 2k). Most of the centra belong to the dorsal region (e.g., Fig. 2l–o), being characterized by the absence of intercentra and articular heads for the ribs. However, some of them are cervical centra, being ventrally keeled and presenting an articulation for the ribs (Fig. 2e2). Finally, others belong to caudal vertebrae, showing a ventral keel posteriorly bifurcated, conforming the articular facets for the chevrons (Fig. 2d2).
Most of the complete isolated neural arches have a relatively small size, the maximum height corresponding to 2.4 cm, and the maximum width to 3.2 cm (Fig. 2n–s). Although most of them are wider than high (Fig. 2q, r), some incomplete arches are interpreted as being higher than wide (e.g., Fig. 2p). The preserved neural spines of the arches attributed here to Eosauropterygia indet. are pointed and very low (Fig. 2q, r). The preserved transverse processes are tall and slightly prominent. The prezygapophyses and postzygapophyses are recognized as being poorly conspicuous. A zygosphene-zygantrum articulation can be observed in the arches that preserve this area (Fig. 2p, s).
Several appendicular elements have also been recovered. Most of them are broken and poorly preserved fragments, so the systematic information provided by them is limited. Four elements (i.e. MGM-325T, MGM-326T, MGM-327T and MGM-331T) correspond to relatively well-preserved femoral remains (Fig. 3). MGM-325T preserves a part of its diaphysis and also the distal area, whereas MGM-326T is a diaphyseal fragment (Fig. 3a, b). The length of the preserved region of the first one is 5.5 cm, and that of the other is 8.1 cm. They are slender and slightly sigmoidal elements, showing a diaphyseal constriction. The articular facets for the tibia and fibula, observed in the distal head of MGM-325T, are not prominent. MGM-327T is a complete right femur (Fig. 3c). It is 6.5 cm in length. It shares with MGM-325T and MGM-326T the very slender and curved morphology, and the presence of a constrained diaphysis. The proximal region is more expanded and massive than the distal one. The proximal region presents an internal trochanter not separated from the femur because the intertrochanteric fossa is poorly developed. Thus, the femoral head shows a triangular section in transversal view. The distal area of the femur is less developed than the proximal. One of the articular facets is broken. MGM-331T is a proximal portion corresponding to a bigger femur than those previously described (Fig. 3d). The maximum length of the preserved region of this bone is 4 cm, the maximum diameter being 2.6 cm. The external surface of this bone is poorly preserved, being dorsally eroded. The internal trochanter is poorly developed. The intertrochanteric fossa is very shallow.
Sauropsida Huxley 1864
Sauropterygia Owen 1860
Eosauropterygia Rieppel 1994
Nothosauroidea Baur 1889
Nothosauria Baur 1889
Nothosauria indet.
(Figure 4)

Nothosaur isolated teeth from the Muschelkalk of Canales de Molina (Guadalajara Province, Spain). From left to right: MGM-425T (a), MGM-424T (b), MGM-394T (c), MGM-395T (d) and MGM-396T (e). Scale bar equals 20 mm
Material: Five isolated teeth belonging to indeterminate members of the clade Nothosauria (Fig. 4).
Locality and horizon: Canales de Molina (Guadalajara, Castilla-La Mancha, Spain). Royuela Dolostones, Marls and Limestones Formation, Ladinian (Middle Triassic, upper Muschelkalk Facies) (Márquez-Aliaga and García-Gil 1991; Vargas et al. 2009; Miguel Chaves et al. 2016; Sánchez-Moya et al. 2016).
Description: Some teeth belonging to indeterminate nothosaurs have also been recovered in Canales de Molina (Fig. 4). The maximum length of the preserved region of these teeth ranges from 0.6 to 1.7 cm. These teeth are conical, pointed and slender (Fig. 4c), ranging from almost straight elements (Fig. 4c, d) to others with a distinct curvature in the basal half of the crown (Fig. 4a, b, e). They are circular in section. Some of the teeth preserve the partial root, lacking a constriction between the crown and this area (Fig. 4a, c, d). The teeth present ornamented enamel along the entire crown, composed of numerous longitudinal ridges that are equidistantly located from each other. The depressions between the ridges are very narrow when compared to the width of the ridges. Although most of the ridges are simple, some of them are bifurcated close to the root of the tooth.
Sauropsida Huxley 1864
Sauropterygia Owen 1860
Eosauropterygia Rieppel 1994
Nothosauroidea Baur 1889
Nothosauria Baur 1889
Nothosauridae Baur 1889
Nothosauridae indet.
(Figure 2t)
Material: MGM-324T, an almost complete isolated neural arch, lacking the transverse processes (Fig. 2t).
Locality and horizon: Canales de Molina (Guadalajara, Castilla-La Mancha, Spain). Royuela Dolostones, Marls and Limestones Formation, Ladinian (Middle Triassic, upper Muschelkalk Facies) (Márquez-Aliaga and García-Gil 1991; Vargas et al. 2009; Miguel Chaves et al. 2016; Sánchez-Moya et al. 2016).
Description: This neural arch is 3.4 cm in height, 2.8 in width, and 1.2 in length. The neural canal is slightly ovoid, taller than wide. The neural spine is straight. It is taller than the rest of the arch, its height being 1.9 cm. Its apical region is slightly ornamented by small pits (Fig. 2t5). This spine is lateromedially expanded toward the tip, being apically inflated. MGM-324T presents a well-developed bipartite zygosphene in the anterior surface (Fig. 2t5, t6), and a zygantrum consisting of two depressions separated by a thin lamina of bone in the posterior surface of the neural arch (Fig. 2t2). The prezygapophyses and postzygapophyses are also preserved, being recognized as poorly developed.
Sauropsida Huxley 1864
Sauropterygia Owen 1860
Eosauropterygia Rieppel 1994
Nothosauroidea Baur 1889
Nothosauria Baur 1889
Nothosauridae Baur 1889
cf. Nothosaurus Münster 1834
cf. Nothosaurus
(Figure 5)

MGM-285T, a partial humerus of cf. Nothosaurus from the Muschelkalk of Canales de Molina (Guadalajara Province, Spain). a MGM-285Ta, proximal head in ventral (a1) and dorsal (a2) views. b MGM-285Tb, distal head in ventral (b1) and dorsal (b2) views. c Schematic reconstruction of the humerus with the grey areas representing the recovered fragments. Anatomical abbreviation: dpc deltopectoral crest, ect. gr. ectepicondylar groove, ldi latissimus dorsi insertion (sensu Bickelmann and Sander 2008). Scale bar equals 20 mm
Material: MGM-285T, the proximal (MGM-285Ta) and distal (MGM-285Tb) portions of a right humerus (Fig. 5).
Locality and horizon: Canales de Molina (Guadalajara, Castilla-La Mancha, Spain). Royuela Dolostones, Marls and Limestones Formation, Ladinian (Middle Triassic, upper Muschelkalk Facies) (Márquez-Aliaga and García-Gil 1991; Vargas et al. 2009; Miguel Chaves et al. 2016; Sánchez-Moya et al. 2016).
Description: MGM-285T is a partial and eroded right humerus that only preserves the proximal and distal regions (MGM-285Ta and MGM-285Tb, respectively), lacking the diaphysis (Fig. 5). Both fragments corresponds to dorsoventrally compressed elements (Fig. 5a3, b3). The preserved region of the proximal region has 6.8 cm in height and 4.6 cm in width. Its ventral surface is flat (Fig. 5a1). The dorsal surface presents a concavity in the postaxial half, and also an elevated area that corresponds to a well-developed region for the latissimus dorsi insertion (sensu Bickelmann and Sander 2008) (Fig. 5a2). This humerus also presents a well-developed and relatively flat deltopectoral crest (sensu Bickelmann and Sander 2008). The deltopectoral crest and the area for the latissimus dorsi insertion give the proximal portion an angulated aspect. The distal portion of the humerus is relatively flat element, with a fan-like morphology (Fig. 5b). It is 8.4 cm in height and 4.5 cm in width. Its external surface is eroded, being partially lost, especially in the ventral surface. The epicondyles are weakly developed. The ectepicondylar groove is located in the preaxial margin. The entepicondylar foramen cannot be recognized, due to the poor preservation of the bone.
3 Discussion
Although the reptile remains from the Middle Triassic levels of Canales de Molina studied here are isolated and, in some cases, fragmentary elements, they provide enough information to identify them as Eosauropterygia.
The vertebral centra of the members of Eosauropterygia have a butterfly-shaped articular surface with the neural arch, recognized as a synapomorphy for this clade (Rieppel 1994). This morphology is present in all the centra from Canales de Molina studied here, allowing us its attribution to Eosauropterygia. Some of the studied centra are ventrally keeled and present an articulation for the ribs (Fig. 2e2), characters that allow us to recognize them as cervical centra; others show a ventral keel posteriorly bifurcated, conforming the articular facets for the chevrons (Fig. 2d2), indicating a caudal position (Rieppel 1994, 2000). Most of the centra, however, lack these structures, being identified as belonging to the dorsal region. The centra identified as dorsal show several morphologies and sizes. Some centra are wider than larger (Fig. 2f), the opposite condition being recognized in others (e.g., Fig. 2b, e). Some of them are strongly constrained dorsoventrally, resulting in flattened elements (Fig. 2f–h), but this condition is not present in others (e.g., Fig. 2i–k, m). This flattening is not caused by taphonomical causes. Finally, whereas some of vertebral centra are slightly amphicoelus (e.g., Fig. 2n), others are completely platycoelous (e.g., Fig. 2m). The basal sauropterygians (i.e., those members of the clade Sauropterygia that do not include the plesiosaurs) present a paedomorphic postcranial skeleton, including the lack of fusion of the neurocentral suture (Rieppel 1993, 2000). Therefore, the finding of isolated centra and neural arches of eosauropterygians cannot been explained by ontogenetic causes. This, together with the poorly informative nature of the centra of this group, only allow the attribution of these elements from Canales de Molina to indeterminate members of Eosauropterygia.
The neural arches from the Muschelkalk of Canales de Molina can be also attributed to the clade Eosauropterygia. They present an additional articulation in the neural arch, the zygosphene-zygantrum articulation (Fig. 2p, s, t1, t2), which is an exclusive character for the clade (Rieppel 1994). Some of the best preserved arches are of small size, lacking prominent transverse processes and characterized by the presence of a very low spine (Fig. 2q, r). This morphology is present in some clades of Eosauropterygia of small size, like pachypleurosaurs and most of the lariosaurs, but also in the cervical vertebrae of eosauropterygians with high neural spines, like some species of Nothosaurus (considered here as a monophyletic genus following Lin et al. 2015, although some studies, like Liu et al. 2014 and Klein et al. 2016, indicate that it could be paraphyletic) (see Figs. 5 and 7 in Diedrich 2013a). Thus, most of the neural arches from Canales de Molina are identified as belonging to indeterminate members of Eosauropterygia. However, the availability of more characters which provide systematic information in other vertebral elements from this site allows a more precise taxonomic attribution, being assigned to Nothosauridae. This is the case of MGM-324T (Fig. 2t), a well preserved neural arch that shows a well-developed bipartite zygosphene (Fig. 2t1, t5, t6). The neural arches of the nothosaurs present this structure, whereas its development is smaller in other eosauropterygians (Rieppel and Wild 1996; Rieppel et al. 1997).
The femoral remains studied here can also be identified as belonging to Eosauropterygia (Fig. 3). The femur in this group is usually a slender and sigmoidally curved element, with the proximal region bigger than the distal one, and presenting confluent articular facets for the tibia and fibula not projected from the distal end of the femur (Sanz 1984; Rieppel 2000; Klein 2010). MGM-325T, MGM-326T, MGM-327T and MGM-331T share these characters (Fig. 3). In addition, the intertrochanteric fossa is very shallow in this clade, the internal trochanter being weakly developed, resulting in a triangular section of the proximal area (Rieppel 1994, 2000). The characteristic eosauropterygian proximal head, with reduced internal trochanter and intertrochanteric fossa, can be observed in MGM-327T and MGM-331T (Fig. 3c, d). Although the appendicular skeleton in basal sauropterygians is paedomorphic for several characters, including the presence of reduced epicondyles, these structures are already ossified at early stages of development (Rieppel 1993, 2000; Lin and Rieppel 1998). Thus, other aspects, as the humerus/femur ratio of the same specimen (not known here) are used as indicators of maturity (Lin and Rieppel 1998). In addition, the femur in Eoauropterygia is a conservative element, so it is commonly not possible to distinguish those of different members (Sanz 1984; Rieppel 2000; Klein 2010). Considering all these ideas, and also the fact that the femoral remains from Canales de Molina are broken and isolated bones, there is no possibility to assess whether the differences in size and morphology described in this paper correspond to interspecific or intraspecific variability.
Some of the isolated teeth recovered from Canales de Molina present a root, which indicates a thecodont implantation (Fig. 4a, c, d). The members of Sauropterygia, like many other groups of reptiles, present thecodont implantation (Rieppel 2001a). The teeth of Canales de Molina show an ornamented crown consisting of long and longitudinal ridges (Fig. 4). This is consequence of the folding of the enamel, a character shared by most of the Triassic eosauropterygians (Sanz 1980; Alafont 1992). Although the ornamentation of the enamel has taxonomic utility in plesiosaurs, this is not the case in basal eosauropterygians due to their huge intraspecific and, even, intraindividual degree of variability (Sanz 1980, 1984; Alafont 1992). The teeth from Canales de Molina are slender, conical, pointed and recurved. Within basal eosauropterygians, this dental morphology has been traditionally associated with nothosaurs (Sanz 1980; Alafont 1992), but some non-nothosaurid eosauropterygians present similar dentition to that of the nothosaurs (e.g., Corosaurus alcovensis from the United States, or Chinchenia sungi and Dawazisaurus brevis, from China. See Storrs 1991; Rieppel 1999; Cheng et al. 2016). However, the only known eosauropterygians with this morphology from the Triassic record of Europe are the nothosaurs, which are relatively common in the Spanish Middle Triassic record (e.g., Kuhn-Schnyder 1966; Sanz 1976, 1983, 1991; Alafont 1992, 1999; Niemeyer 2002; Reolid et al. 2013). Thus, the pachypleurosaurs present a straight crown, only curved in the apex; Simosaurus has blunt and conical teeth, constrained in the base; and the pistosaurs lack ornamentation (Sanz 1980; Rieppel 1994; Diedrich 2013b). The different degree of curvature in the teeth from this site can be explained by the strong heterodonty present in the nothosaurs (Sanz 1980). Therefore, we attributed the teeth from Canales de Molina to indeterminate members of Nothosauria.
MGM-285T is a partial humerus which only preserves the proximal and distal regions (Fig. 5). Although the distal portion is poorly preserved, the epicondyles, weakly developed, can be identified (Fig. 5b). The basal members of Sauropterygia present poorly developed epicondyles in the long bones, which is a consequence of the paedomorphosis that characterizes the postcranial skeleton of the group (Rieppel 1993, 2000; Bickelmann and Sander 2008). The proximal head is angulated. It presents a distinct deltopectoral crest and a well-developed latissimus dorsi insertion (Fig. 5a). This character combination has been identified as being exclusive of the eosauropterygian Nothosaurus (Bickelmann and Sander 2008). However, the humerus from Canales de Molina is rather flat, whereas the humerus in Nothosaurus, as in other eosauropterygians, usually presents a massive proximal region (Rieppel 2000; Bickelmann and Sander 2008; Klein 2010). The flattening of the proximal portion of the humerus from Canales de Molina does not seem to be caused by taphonomical causes, so we classify this element as cf. Nothosaurus.
Remains attributed to the sauropterygian genus Nothosaurus are relatively common in Europe, having been found in Middle and lower Upper Triassic levels of several countries (e.g., France, Germany, the Netherlands, Spain), but also in the north of Africa (Tunisia), Middle East (Israel) and Asia (China) (Rieppel and Wild 1996; Rieppel et al. 1997, 1999; Rieppel 2000, 2001b; Albers and Rieppel 2003; Li and Rieppel 2004; Albers 2005; Jiang et al. 2006; Shang 2006; Bickelmann and Sander 2008; Klein and Albers 2009; Ji et al. 2014; Liu et al. 2014; Klein et al. 2015). This taxon have been recognized in several sites of the Iberian Peninsula: in the Spanish provinces of Teruel (Aragón), Lleida (Catalonia), Jaén (Andalusia) and Soria (Castilla y León) (Kuhn-Schnyder 1966; Sanz 1983; Niemeyer 2002; Reolid et al. 2013; Miguel Chaves et al. 2016). The first of the confirmed references to Nothosaurus in the Iberian Peninsula comes from the Muschelkalk of the locality of Royuela (Teruel), where a sacral vertebrae, and several teeth and ribs were assigned to this genus (Kuhn-Schnyder 1966). A new species of Nothosaurus, N. cymatosauroides, was described based on the cast of a skull, vertebrae, ribs and gastral elements an individual from the Muschelkalk of Alcover (Lleida) (Sanz 1983). However, the validity of this species was subsequently discussed by Rieppel (2000), due to the poorly informative nature of the specimen. Some isolated bones (including vertebrae, teeth, ribs, a possible fragment of jaw, a clavicle, a scapula, a coracoid, two humeri, a femur, a tibia and an ilion) were recovered from the Muschelkalk of Siles (Jaén), and attributed to Nothosaurus (Niemeyer 2002). Several vertebral remains from the Muschelkalk of Puente Génave-Villarodrigo, also in Jaén, were identified as belonging to Nothosaurus (Reolid et al. 2013). The last identified reference to this genus in the Iberian Peninsula corresponds to four partial vertebrae of a single specimen from the Muschelkalk of Fuencaliente de Medinaceli (Soria), attributed to Nothosaurus cf. giganteus (Miguel Chaves et al. 2016).
4 Conclusions
The Ladinian (Middle Triassic) site of Canales de Molina (Guadalajara, Spain), located in upper Muschelkalk Facies belonging to the Royuela Dolostones, Marls and Limestones Formation, has provided an abundant sample of isolated eosauropterygian fossil remains. These bones include vertebrae, teeth and appendicular elements. Most of them only allow the identification of small to medium sized indeterminate members of Eosauropterygia. Other elements allow more precise taxonomical attributions. Thus, the isolated teeth and the neural arch MGM-324T allow us to recognize the presence of nothosaurs in Canales de Molina. The relatively large fragmentary humerus MGM-285T shows a combination of characters traditionally considered exclusive of the eosauropterygian Nothosaurus. In addition, other indeterminate and relatively large eosauropterygian elements are also compatible with this taxon, also corresponding to relatively large individuals. This is the case of some of the vertebral elements (the centrum MGM-293T or the neural arch MGM-324T), the teeth or the proximal head of the femur MGM-331T. However, considering the limited availability of characters, they are attributed here to indeterminate nothosaurs (the teeth and the neural arch MGM-324T) and to indeterminate members of Eosauropterygia (the vertebral centrum MGM-293T and the partial femur MGM-331T). Considering the relatively limited availability of characters, and the paedomorphic nature of the postcranial skeleton of this group, the attribution of the small elements to other small forms identified in the Iberian Triassic record (i.e. lariosaurs or pachypleurosaurs) or to juvenile individuals of Nothosaurus or to a taxon closely related to it, cannot be performed. Thus, at least a relatively large member of Eosauropterygia, closely related to Nothosaurus, is identified in this site.
References
Alafont, L. S. (1992). Notosaurios y Placodontos (Reptilia) del Triásico Medio de Bienservida-Villarrodrigo. Diputación de Albacete, Spain: Instituto de Estudios albacetenses de la Excma. Diputación de Albacete, Spain.
Alafont, L. S. (1999). Reptiles del Triásico. In A. Aguirre & I. Rábano (Eds.), La huella del pasado: Fósiles de Castilla–La Mancha (pp. 143–159). Junta de Comunidades de Castilla–La Mancha.
Alafont, L. S., & Sanz, J. L. (1996). Un nuevo sauropterigio (Reptilia) en el Triásico de la Sierra de Prades (Tarragona). Cuadernos de Geología Ibérica, 20, 313–329.
Albers, P. C. H. (2005). A new specimen of Nothosaurus marchicus with features that relate the taxon to Nothosaurus winterswijkensis. PalArch’s Journal of Vertebrate Palaeontology, 3(1), 1–7.
Albers, P. C. H., & Rieppel, O. (2003). A new species of the sauropterygian genus Nothosaurus from the lower Muschelkalk of Winterswijk, The Netherlands. Journal of Paleontology, 77(4), 738–744.
Alférez, F., Molero, G., & Maldonado, E. (1983). Un nuevo yacimiento con vertebrados fósiles (Peces y Reptiles) en el Trías de la Cordillera Ibérica (Canales de Molina (p. 44). Guadalajara): Real Sociedad Española de Historia Natural. VI Reunión Bienal, Santiago.
Almela, A., & Llopis Lladó, N. (1947). Hoja geológica num. 392 (Sabadell). Mapa Geológico de España E. 1:50.000. Segunda serie, I.G.M.E., Madrid.
Bakker, R. T. (1993). Plesiosaur extinction cycles—Events that mark the beginning, middle and end of the Cretaceous. In W. G. E. Caldwell & E. G. Kauffman (Eds.), Evolution of the Western Interior Basin (pp. 641–664). Geological Association of Canada.
Bardet, N., Falconnet, J., Fischer, V., Houssaye, A., Jouve, S., Pereda Suberbiola, X., et al. (2014). Mesozoic marine reptile palaeobiogeography in response to drifting plates. Gondwana Research, 26, 869–887.
Bauzá Rullán, J. (1955). Notas paleontológicas de Mallorca: sobre el hallazgo del Nothosaurus en el Trias. Boletín de la Sociedad de Historia Natural de Baleares, 1, 87.
Bickelmann, C., & Sander, P. M. (2008). A partial skeleton and isolated humeri of Nothosaurus (Reptilia: Eosauropterygia) from Winterswijk, The Netherlands. Journal of Vertebrate Paleontology, 28(2), 326–338.
Cheng, Y., Wu, X., Sato, T., & Shan, H. (2016). Dawazisaurus brevis, a new eosauropterygian from the Middle Triassic of Yunnan. China. Acta Geologica Sinica (English Edition), 90(2), 401–424.
Diedrich, C. G. (2013a). Shallow marine sauropterygian reptile biodiversity and change in the Bad Sulza Formation (Illyrian, Middle Triassic) of central Germany, and a contribution to the evolution of Nothosaurus in the Germanic Basin. In L. H. Tanner, J. A. Siepelmann & S. G. Lucas (Eds.), The Triassic System: New Developments in Stratigraphy and Paleontology. New Mexico Museum of Natural History and Science, Bulletin (Vol. 61, pp. 132–158).
Diedrich, C. G. (2013b). The oldest “subaquatic flying” reptile in the world—Pistosaurus longaevus MEYER, 1839 (Sauropterygia) from the Middle Triassic of Europe. In L. H. Tanner, J. A. Siepelmann & S. G. Lucas (Eds.), The Triassic System: New Developments in Stratigraphy and Paleontology. New Mexico Museum of Natural History and Science, Bulletin (Vol. 61, pp. 169–215).
Ferrando, P. (1912). Sobre el reptil fósil sauropterigido notosaurido, encontrado en Estada (Huesca). Boletín de la Real Sociedad Española de Historia Natural, 12, 188.
Fortuny, J., Bolet, A., Sellés, A. G., Cartanyà, J., & Galobart, À. (2011). New insights on the Permian and Triassic vertebrates from the Iberian Peninsula with emphasis on the Pyrenean and Catalonian basins. Journal of Iberian Geology, 37, 65–86.
Ji, C., Jiang, D.-Y., Rieppel, O., Motani, R., Tintori, A., & Sun, Z.-Y. (2014). A new specimen of Nothosaurus youngi from the Middle Triassic of Guizhou. China. Journal of Vertebrate Paleontology, 34(2), 465–470.
Jiang, D.-Y., Maisch, M. W., Hao, W.-C., Sun, Y.-L., & Sun, Z.-Y. (2006). Nothosaurus yangjuanensis n. sp. (Reptilia, Sauropterygia, Nothosauridae) from the middle Anisian (Middle Triassic) of Guizhou, southwestern China. Neues Jahrbuch für Geologie und Paläontologie Monatshefte, 5, 257–276.
Jiang, D.-Y., Motani, R., Tintori, A., Rieppel, O., Chen, G.-B., Huang, J.-D., et al. (2014). The Early Triassic eosauropterygian Majiashanosaurus discocoracoidis, gen. et sp. nov. (Reptilia, Sauropterygia), from Chaohu, Anhui Province, People’s Republic of China. Journal of Vertebrate Paleontology, 34(5), 1044–1052.
Klein, N. (2010). Long bone histology of Sauropterygia from the lower Muschelkalk of the Germanic Basin provides unexpected implications for phylogeny. PLoS One, 5(7), e11613.
Klein, N., & Albers, P. C. H. (2009). A new species of the sauropsid reptile Nothosaurus from the Lower Muschelkalk of the western Germanic Basin, Winterswijk, The Netherlands. Acta Palaeontologica Polonica, 54(4), 589–598.
Klein, N., Voeten, D. F. A. E., Haarhuis, A., & Bleeker, R. (2016). The earliest record of the genus Lariosaurus from the early middle Anisian (Middle Triassic) of the Germanic Basin. Journal of Vertebrate Paleontology. doi:10.1080/02724634.2016.1163712
Klein, N., Voeten, D. F. A. E., Lankamp, J., Bleeker, R., Sichelschmidt, O. J., Liebrand, M., et al. (2015). Postcranial material of Nothosaurus marchicus from the Lower Muschelkalk (Anisian) of Winterswijk, The Netherlands, with remarks on swimming styles and taphonomy. Paläontologische Zeitschrift, 89, 961–981.
Kuhn-Schnyder, E. (1966). Über Wirbeltier-Reste ausdem Muschelkalk der Provinz Teruel (Spanien). Oberrheinische Geologische Abhandlungen, 15, 99–106.
Lapparent, A. F. (1966). Noveaux gisements de reptiles mesozoiques en Espagne. Notas y Comunicaciones del Instituto Geológico y Minero de España, 84, 103–110.
Li, J., & Rieppel, O. (2004). A new nothosaur from Middle Triassic of Guizhou, China. Vertebrata PalAsiatica, 42(1), 1–12.
Lin, W.-B., Ji, C., Jiang, D., Rieppel, O., Motani, R., Tintori, A., et al. (2015). Postcranial differences between Nothosaurus and Lariosaurus (Sauropterygia: Nothosauridae) revealed by the new specimens from the Middle Triassic of Southwestern China. Journal of Vertebrate Paleontology, Program and Abstracts, 2015, 164.
Lin, K., & Rieppel, O. (1998). Functional morphology and ontogeny of Keichousaurus hui (Reptilia, Sauropterygia). Fieldiana: Geology, 39, 1–35.
Liu, J., Hu, S.-X., Rieppel, O., Jiang, D.-Y., Benton, M. J., Kelley, N. P., et al. (2014). A gigantic nothosaur (Reptilia: Sauropterygia) from the Middle Triassic of SW China and its implication for the Triassic biotic recovery. Scientific Reports, 4(7142), 1–9.
López-Gómez, J. (1985). Sedimentología y estratigrafía de los materiales pérmicos y triásicos del sector SE de la Rama Castellana de la Cordillera Ibérica entre Cueva de Hierro y Chelva (provincias de Cuenca y Valencia). Seminarios de Estratigrafía, Serie Monografías, 11, 1–344.
Márquez-Aliaga, A., & García-Gil, S. (1991). Paleontología y ambientes del Triásico Medio en el sector noroccidental de la Cordillera Ibérica (Provincias de Soria y Guadalajara, España). Estudios Geológicos, 47, 85–95.
Martínez-Delclòs, X. (1995). Plants. Invertebrates. Systematic Paleontology. In X. Martínez-Delclòs (Ed.), II International Symposium on Lithographic Limestones. Montsec & Montral-Alcover. Two Konservat-Lagerstätten, Field trip guide book (pp. 81–87). Catalonia, Spain.
Miguel Chaves, C. de, García-Gil, S., Ortega, F., Sanz, J. L., & Pérez-García, A. (2016). First Triassic tetrapod (Sauropterygia, Nothosauridae) from Castilla y León: evidence of an unknown taxon for the Spanish record. Journal of Iberian Geology, 42(1), 29–38.
Miguel Chaves, C. de, Pérez-García, A., Cobos, A., Royo-Torres, R., Ortega, F., & Alcalá, L. (2015). A diverse Late Triassic tetrapod fauna from Manzanera (Teruel, Spain). Geobios, 48, 479–490.
Niemeyer, J. (2002). Invertebraten und Vertebraten aus dem Muschelkalk von Siles (Jaén), Spanien. Münstersche Forschungen zur Geologie und Paläontologie, 94, 1–98.
Pinna, G. (1990). I rettili placodonti dei terreni triassici di Spagna. Atti della Societa Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano, 131, 137–143.
Quesada, J. M., & Aguera González, S. (2005). Descripción del primer ejemplar de Ceresiosaurus (Reptilia: Sauropterygia), hallado en la Península Ibérica en el Ladiniense (Triásico Medio) de Mont–ral–Alcover (Tarragona). Estudios Geológicos, 61, 247–269.
Quesada, J., Pérez-García, A., Ortega, F., Escaso, F., & Sánchez-Chillón, B. (2009). Simosaurus (Nothosauria) fossil site: a new Spanish window to the Upper Triassic. Journal of Vertebrate Paleontology, 29 (3)-ABSTRACTS, 168A.
Reolid, M., Pérez-Valera, F., Benton, M. J., & Reolid, J. (2013). Marine flooding event in continental Triassic facies identified by a nothosaur and placodont bonebed (South Iberian Paleomargin). Facies, 60, 277–293.
Rieppel, O. (1993). Middle Triassic reptiles from Monte San Giorgio: recent results and future potential of analysis. Paleontologia Lombarda, 2, 131–144.
Rieppel, O. (1994). Osteology of Simosaurus gaillardoti and the relationships of stem–group Sauropterygia. Fieldiana: Geology, 28, 1–85.
Rieppel, O. (1999). The sauropterygian genera Chinchenia, Kwangsisaurus, and Sanchiaosaurus from the Lower and Middle Triassic of China. Journal of Vertebrate Paleontology, 19(2), 321–337.
Rieppel, O. (2000). Part 12A. Sauropterygia I. In P. Wellnhofer (Ed.), Handbuch der Paläoherpetologie (pp. 1–134). Verlag Friedrich Pfeil, München.
Rieppel, O. (2001a). Tooth implantation and replacement in Sauropterygia. Paläontologische Zeitschrift, 75(2), 207–217.
Rieppel, O. (2001b). A new species of Nothosaurus (Reptilia: Sauropterygia) from the Upper Muschelkalk (Lower Ladinian) of southwestern Germany. Paleontographica A, 263, 137–161.
Rieppel, O., & Hagdorn, H. (1998). Fossil reptiles from the Spanish Muschelkalk (Mont–ral and Alcover, Province Tarragona). Historical Biology, 13, 77–97.
Rieppel, O., Mazin, J. M., & Tchernov, E. (1997). Speciation along rifting continental margins: a new nothosaur from the Negev (Israel). Earth and Planetary Sciences, 325, 991–997.
Rieppel, O., Mazin, J. M., & Tchernov, E. (1999). Sauropterygia from the Middle Triassic of Makhtesh Ramon, Negev, Israel. Fieldiana: Geology, 40, 1–85.
Rieppel, O., & Wild, R. (1996). A revision of the genus Nothosaurus (Reptilia: Sauropterygia) from the Germanic Triassic, with comments on the status of Conchiosaurus clavatus. Fieldiana: Geology, 34, 1–82.
Rubio, C., Díez Ferrer, J. B., Ferrer, J., Gámez Vintaned, J. A., Sender, L. M., & Zianni, H. (2003). La Comarca de Gúdar—Javalambre da a conocer su rico patrimonio geológico y paleontológico. Naturaleza Aragonesa, 11, 21–34.
Sánchez-Moya, Y., Herrero, M. J., & Sopeña, A. (2016). Strontium isotopes and sedimentology of a marine Triassic succession (upper Ladinian) of the westernmost Tethys, Spain. Journal of Iberian Geology, 42(2), 171–186.
Sanz, J. L. (1976). Lariosaurus balsami (Sauropterygia, Reptilia) de Estada (Huesca). Estudios Geológicos, 32, 547–567.
Sanz, J. L. (1980). Los notosaurios españoles. Análisis filogenético y sistemático de los sauropterigios triásicos. Ph.D. thesis, Universidad Complutense de Madrid, Madrid.
Sanz, J. L. (1983). Los notosaurios (Reptilia, Sauropterygia) españoles. Estudios Geológicos, 39, 193–215.
Sanz, J. L. (1984). Osteología comparada de las familias Nothosauridae y Simosauridae (Reptilia, Sauropterygia). Estudios Geológicos, 40, 81–105.
Sanz, J. L. (1991). Notosaurios y placodontos (Reptilia). Su registro en el yacimiento de Bienservida–Villarodrigo. Actas de las Jornadas sobre el Medio Natural Albacetense (pp. 17–32). Albacete.
Shang, Q. (2006). A new species of Nothosaurus from the early Middle Triassic of Guizhou, China. Vertebrata PalAsiatica, 4(3), 237–249.
Storrs, G. W. (1991). Anatomy and relationships of Corosaurus alcovensis (Diapsida: Sauropterygia) and the Triassic Alcova Limestone of Wyoming. Bulletin of the Peabody Museum of Natural History Yale University, 44, 1–151.
Stubbs, T. L., & Benton, M. J. (2016). Ecomorphological diversifications of Mesozoic marine reptiles: the roles of ecological opportunity and extinction. Paleobiology, 42(4), 547–573.
Vargas, H., Gaspar-Escribano, J. G., López-Gómez, J., Van Wees, J.-D., Cloetingh, S., de La Horra, R., et al. (2009). A comparison of the Iberian and Ebro Basins during the Permian and Triassic, eastern Spain: A quantitative subsidence modelling approach. Tectonophysics, 474(1–2), 160–183.
Via Boada, L., Villalta, J. F., & Esteban Cerdá, M. (1977). Paleontología y paleoecología de los yacimientos fosilíferos del Muschelkalk superior entre Alcover y Mont-ral (Montañas de Prades, provincia de Tarragona). Cuadernos Geología Ibérica, 4, 247–256.
Westphal, F. (1975). Bauprinzipien im Panzer der Placodonten (Reptilia triadica). Paläontologische Zeitschrift, 49, 97–125.
Acknowledgements
This research is funded by a FPI UNED Grant (Ref. 0271864713 Y0SC001170) and received support from the SYNTHESYS Project (http://www.synthesys.info/) which is financed by European Community Research Infrastructure Action under the FP7 “Capacities” Program (FR-TAF-5710 to C. de Miguel Chaves). We thank Ana Bravo, Silvia Menéndez and the Instituto Geológico y Minero de España for the access to the collection, and to Eleuterio Baeza for the preparation of the material. We also thank the editors Fidel Torcida Fernández-Baldor and José Ignacio Canudo, and the reviewers Torsten Scheyer and Jun Liu for their kind comments, which have helped improve this paper.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
de Miguel Chaves, C., Ortega, F. & Pérez-García, A. The eosauropterygian fossils from the Middle Triassic of Canales de Molina (Guadalajara, Spain). J Iber Geol 43, 129–138 (2017). https://doi.org/10.1007/s41513-017-0011-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s41513-017-0011-9