
Abstract
The abundant Late Miocene proboscidean remains of Greece have never been studied in detail and compared with those of Eurasia in order to determine their taxonomy and their biostratigraphical and palaeoecological significance. The first results of such study are given in this article. During the past decades, several new proboscidean specimens have been added to the old collections, significantly enriching the available material. The Axios Valley (Macedonia, Greece) proboscidean fossils belong mainly to two species of Choerolophodon: C. anatolicus of early Vallesian age and C. pentelici of late Vallesian–Turolian age. Deinotherium giganteum is rare and recognized only in the late Vallesian locality Ravin de la Pluie of Axios Valley. A zygodont form has also been identified in the Turolian of Axios Valley, attributed to “Mammut” sp. The Late Miocene localities of Nikiti (Macedonia, Greece) revealed several remains of C. pentelici, which are similar to the Turolian ones of Axios Valley. The Samos proboscidean collection includes C. pentelici, “Tetralophodon” atticus, “Mammut” sp. and Deinotherium gigantissimum. The taxonomy of the Late Miocene peri-Mediterranean Choerolophodon is given, and the biostratigraphy and palaeoecology of the Greek Late Miocene proboscideans are discussed.
Kurzfassung
Die obermiozänen Proboscidier aus Griechenland, obwohl sie ziemlich verbreitet sind, wurden niemals im Detail untersucht und mit anderen aus Eurasien verglichen, um ihre Taxonomie, Biostratigraphie und Paläoökologie zu bestimmen. Die ersten Ergebnisse dieser Studie sind in diesem Artikel präsentiert. In den letzten Jahrzehnten kamen viele neue Proboscidier-Funde hinzu und bereichern erheblich das Material der alten Sammlungen. Der Großteil der Proboscidea-Fauna aus dem Axios Tal (Mazedonien, Griechenland) wurde der Gattung Choerolophodon mit zwei Arten zugewiesen: C. anatolicus aus dem unteren Vallesium und C. pentelici aus dem oberen Vallesium-Turolium. Die Art Deinotherium giganteum ist selten und wurde nur in der oberen Vallesium Fundstelle Ravin de la Pluie des Axios Tals nachgewiesen. Eine zygodonte Form ist aus dem Turolium des Axios Tals bekannt und wurde der Gattung “Mammut” zugewiesen. Aus den obermiozänen Fundstellen Nikiti (Mazedonien, Griechenland) stammen zahlreiche Reste von C. pentelici, die denjenigen aus dem Turolium des Axios Tals ähneln. Die Proboscidier-Fauna aus Samos enthält die Arten C. pentelici, “Tetralophodon” atticus, “Mammut” sp. und Deinotherium gigantissimum. Die Taxonomie der obermiozänen Arten von Choerolophodon um den Mittelmeerraum wird vorgestellt und die Biostratigraphie und Paläoökologie der griechischen Proboscidier aus dem Oberen Miozän werden besprochen.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Proboscidean remains are common in the Late Miocene faunas, including several taxa; they are present in all Late Miocene mammal fossiliferous sites of Greece, with relatively abundant material. Although they are common in the Greek fossil record, they have never been thoroughly studied. Since a lot of new material has been recently unearthed, the need has arisen for a more complete study of their taxonomy and for an effort to interpret their relationships and their biostratigraphical and palaeoecological significance. In the present article the proboscidean remains from the Late Miocene localities of Macedonia and Samos Island (Greece) are studied. The material comes from both old collections and new ones, revealed during recent excavations.
The proboscideans of Macedonia studied in the present work come from two areas: the lower Axios Valley and Chalkidiki Peninsula (Fig. 1). A set of fossil mammal localities are known from Axios Valley (Fig. 1), belonging to three different formations, correlated to the whole Late Miocene. The Axios Valley proboscidean collection is quite rich, including all discoveries of the last ~40 years of field expeditions in the area, and is housed at LGPUT. Additionally, the proboscideans of the old Arambourg’s collection, housed at MNHN and described by Arambourg and Piveteau (1929), are included in this study.

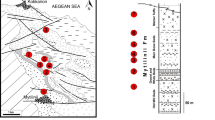
Map of Greece indicating the location of the fossiliferous sites studied in the text. 1 Pikermi, 2 Samos Island, 3 Nikiti, 4 Pentalophos, 5 Xirochori, 6 Ravin de la Pluie and Ravin des Zouaves 1, 7 Ravin des Zouaves 5, 8 Vathylakkos, 9 Prochoma, 10 Dytiko. Inset enlarged area of Axios Valley (localities 4–10). The map was taken from www.shaded-relief.com
The Nikiti localities are situated on Chalkidiki Peninsula (Fig. 1) and have provided a quite rich fossil mammal fauna (Koufos 2006a and refs. cited). Two fossiliferous sites are located in the Nikiti area, belonging to two different formations correlated to the late Vallesian and the early Turolian, respectively (Koufos et al. 1991; Koufos in press and refs. cited). The Nikiti collection includes several remains of proboscideans, housed at LGPUT.
The fossil mammal localities of Samos Island (Fig. 1) are well known since the 19th century. Several old collections from Samos are housed at various museums and institutes and include many interesting proboscidean remains; the main ones (NHMW, MGL, HGI) have been included in the present study. Recent excavations in the Samos fossiliferous sites have provided a new rich mammal collection, housed at NHMA; the proboscidean remains of this new collection are few and have already been described (Konidaris and Koufos 2009). Detailed information on the stratigraphy, the old and new localities, as well as the age of the Samos faunas is given in Kostopoulos et al. (2009) and Koufos et al. (2009).
The majority of the studied proboscidean remains belong to the genus Choerolophodon. Study of this material allows us to follow and recognize the evolution of Choerolophodon during the whole Late Miocene in the peri-Mediterranean region. Therefore, it seems necessary to begin with the systematics of the Late Miocene choerolophodons of this area. For dental measurements of the studied choerolophodons see “Appendix”.
Taxonomy of the Late Miocene peri-Mediterranean choerolophodons
Taxonomical study of the choerolophodont remains from the Late Miocene of the peri-Mediterranean region has a long history. They were originally found in Pikermi (Attica, Greece) and referred to as Mastodon pentelicus, without further description (Gaudry and Lartet 1856). A few years later, the Pikermi material was described in detail under the name Mastodon pentelici (Gaudry 1862, 1867). Material belonging to “Tetralophodon” atticus was also included in this species (Gaudry 1867: pl. 23, figs. 2–4) (Tassy 1985). Subsequently, Schlesinger (1917, 1922) described juvenile choerolophodont material from Samos Island (Greece) and Maragheh (Iran) under the name Mastodon (Choerolophodon) pentelici. Since then, all the Late Miocene choerolophodont material has been referred to as C. pentelici without any distinction. The first recognition of different evolutionary characters in the dental morphology of the Late Miocene choerolophodons was given by Gaziry (1976), who studied Turkish material. Later, Tassy et al. (1989) studied a mandible from the Vallesian locality Eşme Akçaköy (Turkey) and recognized differences from the typical C. pentelici. The authors distinguished two subspecies, based on mandibular and dental morphology: C. p. lydiensis with deflected mandibular symphysis and less expressed choerolophodonty on the molars and C. p. pentelici with straight symphysis and more expressed choerolophodonty. A few years later, the study of the choerolophodons from Kemiklitepe localities (Turkey) indicated two different evolutionary forms of C. pentelici: a primitive one in Kemiklitepe-D (KTD) correlated with the early Turolian, MN 11, and a more advanced one from Kemiklitepe A-B (KTA-B) correlated with the middle Turolian, MN 12 (Tassy 1994a).
Meanwhile, a new choerolophodont species, C. anatolicus, had been previously erected by Ozansoy (1965) based on material from Yassiören (Middle Sinap, Turkey). This species has been regarded as synonymous with C. pentelici for a long time (Gaziry 1976; Tassy 1994a; Tassy et al. 1989). Its validity was recognized quite later, when new material from Sinap (Turkey) was described (Sanders 2003).
The Axios Valley choerolophodont material spans between biozones MN 9 to MN 13 and supports the separation into two distinct species.
Order Proboscidea Illiger, 1811
Superfamily Elephantoidea Gray, 1821
Genus Choerolophodon Schlesinger, 1917
Diagnosis (based on Tassy 1986):
Narrow cranium with zygomatic arches slightly remote from the skull; strong development of the facial region, elongated in comparison with the cerebral region; palate extends behind the last functional molar; nasal fossa and orbits redressed above the posterior half (at least) of the last functional molar; crista orbitotemporalis subvertical or oblique to the base and in front; large and deep symphyseal gutter; upper tusks curve upwards, lack enamel (tusks belonging to juvenile individuals bear an enamel cap), and the cross section is almost circular; lack of lower tusks; lack of dp2, p2, p3/P3, p4/P4; DP2 and dp3/DP3 bilophodont and intermediate molars trilophodont; in the dp3 the first posttrite halflophid posteriorly offset and connected with second pretrite halflophid; in the DP3 the second pretrite halflophid anteriorly offset; choerolophodont teeth: choerodonty, ptychodonty, cementodonty and additionally the intermediate molars and m3/M3 with chevroning of the loph(id)s except for the first one.
Type species:
C. pentelici (Gaudry and Lartet, 1856) (Late Miocene, Southeastern Europe–West Asia)
Other species included:
C. kisumuensis (MacInnes, 1942) (Early to Middle Miocene, North and East Africa)
C. palaeindicus (Lydekker, 1884) (?Early Miocene, Bugti Hills, Pakistan, South Asia)
C. chioticus Tobien, 1980 (Middle Miocene, Chios Island, Greece, Southeastern Europe)
C. ngorora (Maglio, 1974) (Middle to Late Miocene, Kenya, East Africa)
C. zaltaniensis Gaziry, 1987 (Middle Miocene, Libya, North Africa)
C. anatolicus (Ozansoy, 1965) (Late Miocene, Southeastern Europe–West Asia)
C. corrugatus (Pilgrim, 1913) (Late Miocene, Pakistan, South Asia)
Choerolophodon pentelici (Gaudry and Lartet, 1856)
Synonyms
-
1856 Mastodon pentelicus Gaudry and Lartet: p. 273
-
1862 Mastodon pentelici Gaudry: p. 142
-
1867 Mastodon pentelici Gaudry: pl. 22, figs. 1–3; pl. 23, fig. 1
-
1911 Mastodon angustidens Bakalow: p. 37; pl. 1, figs. 1, 2; pl. 2, figs. 1–3; pl. 3, figs. 1, 2; pl. 4, figs. 1–3
-
1913 Mastodon pentelici Pavlow: p. 36; pl. 7, fig. 4
-
1914 Mastodon pentelici Pavlow: p. 184; pl. 8, fig. 16
-
1917 Mastodon (Choerolophodon) pentelici Schlesinger: p. 181; pl. 23, fig. 1; pl. 24, figs. 1, 2; pl. 25, figs. 1–4; pl. 26, figs. 1, 2; pl. 27, figs. 1–3; pl. 28, figs. 1–3; pl. 29, figs. 1–6; pl. 30, figs. 1–4; pl. 31, figs. 1, 2; pl. 32, figs. 1, 2; pl. 33, fig. 1
-
1922 Mastodon (Choerolophodon) pentelici Schlesinger: p.142; pl. 20, figs. 1, 2; pl. 21, fig. 1
-
1924 Mastodon pentelici Mecquenem: p. 137; pl. 1, figs. 1–5, 7, 8, 10–14
-
1929 Mastodon pentelici Arambourg and Piveteau: p. 74; pl. 2, fig. 1
-
1936 Trilophodon (Choerolophodon) pentelicus Osborn: p. 263; fig. 205–208
-
1950 Mastodon pentelici Lehmann: p. 221; pl. 22, fig. 109
-
1952 Mastodon pentelici Senyürek: p. 451; figs. 6, 7
-
1957 Mastodon pentelici Ćirić: p. 13; pl. 9, 10
-
1962 Trilophodon angustidens Bakalov and Nikolov: p. 98; pl. 48; pl. 49, figs. 1–3; pl. 50, figs. 1–3; pl. 51, figs. 1, 2
-
1962 Trilophodon (=Choerolophodon) pentelicus Bakalov and Nikolov: p.105; pl. 52
-
1976 Choerolopodon pentelici Gaziry, p.76; pl. 12, fig. 7
-
1977 Mastodon (Choerolophodon) pentelici Koufos: p. 98; pl. 1, figs. a, b
-
1980 Choerolophodon cf. pentelici Thomas et al.: p. 272
-
1980 Gomphotherium pentelici Koufos: p. 93; figs. 34, 35; pl. 11, fig. 1
-
1980 Tetralophodon longirostris Koufos: p. 106; fig. 37; pl. 11, fig. 2
-
1985 Choerolophodon (Synconolophus) serridentinoides Apostolov and Nikolov: p. 126; pl. 3, figs. 1, 2; pl. 4, figs. 1–3
-
1994 Choerolophodon pentelici Tassy: p. 144; p. 146; pl.1, figs. a, b; pl. 3, figs. a, b
-
2003 Choerolophodon sp. Theodorou et al.: p. 523; fig. 4f
-
2005 Choerolophodon pentelici Tassy: p. 708; figs. 1, 2
-
2009 Choerolophodon pentelici Konidaris and Koufos: p. 140; pl. 1, figs. 1, 2
Type locality:
Pikermi, Attica, Greece.
Age:
Middle Turolian, MN 12, Late Miocene.
Lectotype:
Juvenile skull with DP2-DP4 dex and DP4 sin and associated mandible with dp3-dp4 dex and dp4 sin, MNHN-PIK-3665 (Fig. 2), described by Gaudry (1862: p. 142–152) and figured by Gaudry (1867: pl. 22, figs. 1–3, pl. 23, fig. 1).

Choerolophodon pentelici, Pikermi (PIK), Attica, Greece, middle Turolian, MN 12. Skull and associated mandible, LECTOTYPE, MNHN-PIK-3665; a, c lateral, and b, d occlusal view
Diagnosis:
Large choerolophodont species; elongated skull with moderately inclined facial region and low cerebral region; orbits situated at the top of the skull and far behind the last molar in function, the anterior zygomatic processus is situated behind the dentition, at the level of the anterior border of the choane, and the whole perinasal area is redressed; distant temporal lines and wide parietal region; the symphysis is situated ventrally at the extension of the horizontal ramus, in both juvenile and adult specimens; anterior border of ascending and horizontal ramus form obtuse angle; in mandibles bearing the m3 there is a large retromolar gap; upper tusks emerge sub-horizontally and outwards and then curve upwards; deciduous teeth are large; the posterior cingulum of the DP3 is well developed, and the metacone is separated from the posttrite cusplet of the posterior cingulum; the posterior cingulum of the dp3 is prominent and isolated from the second lophid in the more advanced morphs, forming a third lophid; five lophids or four lophids with an isolated posterior “heel” in the m3; the M3 bears four lophs; well-expressed choerodonty, ptychodonty and cementodonty.
Important localities:
Pikermi, Samos Island, Xirochori, Ravin de la Pluie, Ravin des Zouaves 5, Dytiko (Greece), Maragheh (Iran), Kemiklitepe (Turkey), Burgas (Bulgaria).
Choerolophodon anatolicus (Ozansoy, 1965)
-
1965 Trilophodon (Choerolophodon) anatolicus Ozansoy: p. 47; pl. 9, figs. 1, 2
-
1976 Choerolophodon pentelici Gaziry: p. 46, 49, 52; pl. 5, figs. 5–7; pl. 6, figs. 1–5; pl. 7, figs. 1–4; pl. 8, figs. 1–3; pl. 9, figs. 1–4, 6–9; pl. 10, figs. 1–6; pl. 11, figs. 1–3; pl. 12, figs. 1, 2
-
1989 Choerolophodon pentelici lydiensis Tassy et al.: p. 2146; figs. a, b
-
2003 Choerolophodon anatolicus Sanders: p. 208; figs. 10.5, 10.6a-c, 10.8a-b
Type locality:
Yassiören (Middle Sinap), Turkey.
Age:
Vallesian, Late Miocene; MN 9 according to Mein (1990), de Bruijn et al. (1992)
Lectotype:
Maxilla with DP2-DP4 dex and DP3-DP4 sin (Figs. 3a, 4a). We designate the lectotype of C. anatolicus, since Ozansoy (1965: p. 47–48, pl. 9, figs. 1, 2) did not determine the holotype of this species, and the included material originated from different fossiliferous layers and did not thus belong to the same individual. The maxilla presents diagnostic morphological features which differentiate it from the type species C. pentelici. The maxilla is housed at MNHN (MNHN-TRQ-1000).

Choerolophodon anatolicus, Yassiören (Middle Sinap), Turkey, Vallesian, MN 9–10. a maxilla, LECTOTYPE, MNHN-TRQ-1000, occlusal view; b, c left mandibular fragment (cast housed at MNHN-YAS-49, 50), b lateral, and c occlusal view

a DP3 dex, C. anatolicus, Yassiören (MNHN-TRQ-1000); b DP3 sin (reversed), C. pentelici, Pikermi (MNHN-PIK-1705)
Diagnosis:
Small choerolophodont species; elongated skull with moderately inclined facial region and low cerebral region; orbits situated at the top of the skull and just behind the last functional teeth; anterior zygomatic processus situated anterior of the choanae at the level of the last molar; moderate redressed perinasal area; distant temporal lines and wide parietal region; in juvenile specimens the mandibular symphysis is situated ventrally horizontal, while in adult ones it is deflected downwards; ascending and horizontal ramus form obtuse angle, and retromolar gap is present; upper tusks emerge sub-horizontally and outwards and then curve upwards; deciduous teeth are small; in DP3 moderate development of the posterior cingulum and metacone connected to the posttrite cusplet of the posterior cingulum; in dp3 weak development of the posterior cingulum, which is connected to the second pretrite lophid; m3 with four or five lophids; in some tetralophodont m3 the posterior cingulum is connected to the fourth lophid; M3 with four lophs; moderate to well-expressed choerodonty, ptychodonty and cementodonty.
Differential diagnosis:
C. anatolicus differs from the type species in the ventrally deflected adult mandibular symphysis, the more anteriorly situated orbit and anterior zygomatic processus and the generally less redressed perinasal area, the weaker posterior cingulum in the dp3/DP3 connected to the second loph(id) (Fig. 5), the smaller deciduous teeth and the variable presence of tetralophodont m3’s with posterior cingulum connected to the fourth lophid (primitive character observed in earlier choerolophodons and absent in C. pentelici); the molars tend to be smaller in size.

DP3 and dp3 of Choerolophodon from various localities of Greece. a C. anatolicus, LGPUT-PNT-93, DP3 sin (reversed); b C. pentelici, LGPUT-XIR-21, DP3 sin (reversed); c C. pentelici, LGPUT-NIK-1605, DP3 dex; d C. anatolicus, LGPUT-PNT-158, dp3 sin (reversed); e C. pentelici, LGPUT-RPI-259, dp3 dex; f C. pentelici, LGPUT-NIK-1590, dp3 dex
In agreement with Tassy (1983) and Tassy et al. (1989), the ventral margin of the mandibular symphysis must be considered as a distinctive character among choerolophodont species. C. anatolicus presents a mandibular symphysis that is horizontal in juvenile individuals (Fig. 3b) and deflected in adult ones (Tassy et al. 1989: fig. b). According to Tassy et al. (1989), in elephantiformes the rostrum of juvenile specimens is horizontal, regardless of the deflection in adult ones; for example, Tassy (1994b) mentions the straight ventrally mandibular symphysis in juvenile specimens of Gomphotherium angustidens (Cuvier, 1817) and the deflection in adult ones. Tassy et al. (1989) proposed that the absence of inclination in adult C. pentelici could be attributed to paedomorphosis and cannot be interpreted as retention of a primitive character.
Important localities:
Sinap-Middle Member, Eşme Akçaköy, Akin, Kayadibi (Turkey), Pentalophos (Greece).
Axios Valley localities
The fossiliferous sites of Axios Valley were discovered in the beginning of the 20th century; the first collection is housed at MNHN and is labelled as “Salonique” (Arambourg and Piveteau 1929). This collection originated from several stratigraphic horizons, and its mixing hampers comparisons. The new series of excavations in Axios Valley started in 1972 (de Bonis et al. 1973) and provided a large number of fossils, including numerous proboscidean ones. The study of the Late Miocene deposits of Axios Valley suggests three different Formations (Koufos in press and refs. cited):
-
1.
Nea Mesimvria Formation, which includes the localities Pentalophos 1 (PNT), Xirochori 1 (XIR), Ravin de la Pluie (RPl) and Ravin des Zouaves 1 (RZ1)
-
2.
Vathylakkos Formation, which includes the localities Ravin des Zouaves 5 (RZO), Prochoma 1 (PXM), Vathylakkos 1, 2, 3 (VLO, VTK, VAT)
-
3.
Dytiko Formation, which includes the localities Dytiko 1, 2, 3 (DTK, DIT, DKO).
More information on the stratigraphy, fauna and age of the Axios Valley Late Miocene mammal localities is given in Koufos (2006a) and Koufos (in press and refs. cited).
Locality Pentalophos 1 (PNT):
The locality Pentalophos has revealed a rich proboscidean material, including juvenile and adult specimens. All of them show typical choerolophodont features and can be attributed to Choerolophodon. Previous references on the presence of Tetralophodon longirostris (Kaup, 1832) (de Bonis and Koufos 1999; Koufos 2006a) were in fact based on molars belonging to adult individuals of Choerolophodon. The dp3/DP3, which are used by several authors (e.g. Tassy 1994a) for the definition of evolutionary stages among Late Miocene Eurasian choerolophodons, show the most primitive characters among the Axios Valley material (Fig. 5a, d). The morphology of these teeth fits with that of C. anatolicus from Yassiören (Middle Sinap, Turkey) described by Ozansoy (1965) (Figs. 3, 4a) and with that of unpublished material from this locality housed at MNHN (MNHN-YAS-24, 44, 46). Moreover, the dimensions of all the PNT juvenile teeth (dp3/DP2-DP4) are close to those of C. anatolicus (Ozansoy 1965; Sanders 2003) (Fig. 6) and confirm their specific attribution. Concerning the adult material, an unworn m3 (PNT-154) with four lophids resembles in morphology and size those from Eşme Akçaköy, Turkey (GPIH-T.K.-Nr. 1792 and cast MNHN-EA-1) described by Gaziry (1976) and Tassy et al. (1989). Unfortunately, an available senile mandible (PNT-156), bearing an almost worn m3 (possibly with five lophids), lacks the symphysis. Based on:

Scatter diagrams (length–width) of the lower and upper third deciduous premolars (dp3/DP3) of Choerolophodon from various localities
-
1.
The more primitive morphology of the juvenile material from Yassiören [MN 9, according to Mein (1990), de Bruijn et al. (1992) and Sanders (2003)]
-
2.
The teeth and mandibular characters of the material from Eşme Akçaköy [early Vallesian (Sickenberg et al. 1975; Mein 1990; de Bruijn et al. 1992; Fortelius 2011)]
-
3.
The more primitive morphology of the adult skull from Akin (Turkey) [Vallesian (Gaziry 1976; Tassy 1985; Sickenberg et al. 1975)]
-
4.
The PNT material and its age,
we recognize C. anatolicus as a valid species and we agree with Sanders (2003) in including in this species C. p. lydiensis. The available data suggest that the biostratigraphic range of this species is restricted to the early Vallesian (MN 9) and not extended to the early Turolian as suggested by Sanders (2003).
As mentioned by Koufos (in press), the Pentalophos fauna is the most peculiar Late Miocene one. Since the PNT choerolophodont material shows more primitive characters than the XIR one, a locality which corresponds to the lower part of late Vallesian, ~9.6 Ma (see below), an older age (early Vallesian) is possible for Pentalophos. This age is in agreement with data from the rest of the fauna; for instance, the PNT giraffids and hipparions are more primitive than the XIR and RPl ones (Geraads 1989; Koufos 2000; de Bonis and Bouvrain 2003), and the PNT aardvark Orycteropus pottieri Ozansoy, 1965 is similar to the Vallesian one from Middle Sinap (de Bonis et al. 1994). The choerolophodont material from Pentalophos gives thus additional evidence for an early Vallesian age for the locality.
Locality Xirochori 1 (XIR):
The locality XIR is also placed into the Nea Mesimvria Fm, and as mentioned above, its choerolophodons bear more evolved characters than those from the PNT material, which places them closer to the typical species C. pentelici. The dp3/DP3 (Fig. 5b) clearly have a more advanced morphology and stoutness in the posterior cingulum, and the dimensions of the juvenile teeth have on average an intermediate position between Pentalophos and the Turolian samples (Fig. 6). The only available m3 (XIR-6) bears five lophids and is morphologically and metrically similar to the m3 from Sinap 83, Turkey [described by Sanders (2003: fig. 10.7); estimated age ~9.45 Ma according to Kappelman et al. (2003)], being larger than the teeth from Yulafli (Turkey, MN 10) described by Geraads et al. (2005). A juvenile skull (XIR-23) presents slightly more primitive morphology in comparison with the type material of C. pentelici from Pikermi in the less redressed perinasal area, whereas a juvenile mandible (XIR-20) shows similar morphology to mandibles of C. pentelici. Based on the dp3/DP3 morphology and size, the XIR choerolophodont material can be determined as C. pentelici. The XIR fauna is correlated with the late Vallesian (MN 10), and the magnetostratigraphic record suggests an estimated age of ~9.6 Ma (Koufos in press and refs. cited). The dental morphology of the XIR Choerolophodon and the age of the locality suggest that it corresponds to the most primitive form of C. pentelici.
Localities Ravin de la Pluie (RPl) and Ravin des Zouaves 1 (RZ1):
Both fossiliferous sites are located in the upper part of Nea Mesimvria Fm and belong to the same horizon (Koufos in press and refs. cited). The RPl choerolophodons (Fig. 5e) are similar to the XIR ones, corresponding to the same evolutionary stage and suggesting a late Vallesian (MN 10) age for the former locality. The RZ1 material includes a juvenile skull and the associated mandible (both bearing the complete juvenile dentition). The dental dimensions of these specimens are rather small (Fig. 6), but unfortunately both dp3/DP3 are totally worn and do thus not provide any morphological data. However, the morphology of the skull and the mandible are comparable to the already known material of C. pentelici from Pikermi and Samos. Apart from the choerolophodont fossils, an isolated deinothere p4 is also known from RPl. Its morphology and size (74.7 × 51.1+ mm) are similar to those of the Montredon specimens (France, late Vallesian) and the large specimens from Dinotheriensande (Germany, Vallesian) [described by Tobien (1988) and Gräf (1957) respectively], permitting its attribution to Deinotherium giganteum Kaup, 1829. The previously mentioned presence of Tetralophodon in RPl (Koufos 1980; de Bonis and Koufos 1999; Koufos 2006a) is not confirmed. The rest of the fauna is diverse in taxa and indicates a late Vallesian (MN 10) age; the magnetostratigraphic record suggests an estimated age of ~9.3 Ma (Koufos in press and refs. cited).
Locality Ravin des Zouaves 5 (RZO):
This locality is situated in the lower part of the Vathylakkos Fm and includes several proboscidean remains. The choerolophodont material is quite rich, containing mostly juvenile specimens. The dp3/DP3 show advanced morphology, similar to that of the Pikermi and Samos teeth, and all deciduous teeth are large, showing dimensions similar to the Turolian samples from Pikermi, Samos, Akkaşdaği, Kemiklitepe A, B (Fig. 6). Therefore, both morphology and dimensions of the RZO juvenile material support a Turolian age. A sub-adult mandible with the symphysis situated ventrally at the extension of the horizontal ramus confirms the attribution of the material to C. pentelici.
Apart from Choerolophodon, a mammutid species has already been identified by Koufos (1980) as “Mammut tapiroides”. It concerns a juvenile skull with clearly developed zygodont characters in the deciduous dentition, thus permitting its attribution to “Mammut” [for the use of quotation marks for the generic name see Markov (2004)]. The particular skull and its teeth are similar to those of a juvenile individual from Pikermi (NHMUK-M 10104), described by Tassy (1985) and identified as Zygolophodon cf. borsoni (Hays, 1834). The RZO skull is also similar (comparison only based on the provided photo) with a juvenile skull from Belka (Ukraine, MN 12) identified as Turicius turicensis (Schinz, 1824) by Korotkevich (1988: pl. 25, fig. b). Furthermore, the dental elements of the RZO skull are similar to those of a juvenile maxilla from Halmyropotamos (Greece, MN 12) described by Melentis (1967: pl. 6, figs. 1–3) as “Mastodon (Zygolophodon) tapiroides”. As has been demonstrated by Markov (2008), the Turolian “Mammut” differs from the Pliocene “M.” borsoni and belongs to “M.” obliquelophus (Mucha, 1980). This species was originally erected for a mandible with a long symphysis and “borsoni” teeth from Romanovka (Ukraine), of uncertain age. A mandible from the Turolian locality of Ahmatovo (Bulgaria) (Nikolov and Kovačev 1966: pl. 2, fig. 1), which is similar to that of “M.” obliquelophus, reinforces the validity of the taxon. The distinctive character between “M.” obliquelophus and “M.” borsoni is the length of the mandibular symphysis. Since no mandible is known from RZO, the studied skull is referred to as “Mammut” sp. The RZO fauna is older than the Pikermi one, and it is correlated to the early Turolian (MN 11); magnetostratigraphic study suggests an estimated age of ~8.2 Ma (Koufos in press and ref. cited).
Locality Prochoma (PXM):
The locality PXM is placed in the Vathylakkos Fm. Its proboscidean remains include some specimens of Choerolophodon. The large size of the lower deciduous teeth (Fig. 6), which are metrically similar to the type material of C. pentelici, as well as the size of two m1’s, which is similar to that of the C. pentelici m1’s from Maragheh (Iran, Mecquenem’s collection, MNHN-MAR-4006, 4143), suggest the attribution of the PXM species to C. pentelici. The PXM fauna is correlated to the middle Turolian (MN 12), and the magnetostratigraphic record suggests an estimated age of ~7.4 Ma (Koufos in press and refs. cited).
Locality Vathylakkos 2 (VTK):
This locality is also situated in the Vathylakkos Fm. The proboscideans are scanty, including only two juvenile maxillary fragments. The morphology of the deciduous teeth, especially of the DP3, which is the largest among the studied ones (Fig. 6), confirms the presence of C. pentelici in the locality. The fauna of VTK is also dated to the middle Turolian (MN 12) (Koufos in press and refs. cited).
Localities Dytiko 2 (DIT) and Dytiko 3 (DKO):
These localities are placed in the Dytiko Fm and provided some proboscidean remains. Choerolophodons are represented by very juvenile specimens. The morphology and size of their dp3/DP3 (Fig. 6) are similar to those of C. pentelici from Samos and Kemiklitepe A, B, allowing their attribution to this species. The sole specimen from DKO is a hemimandible preserving only the dp3. The morphology of the mandible, as well as the morphology and dimensions of the dp3 (Fig. 6), indicate C. pentelici. The complete Dytiko fauna is younger than the Pikermi one and is correlated to the late Turolian (MN 13) (Koufos in press and refs. cited).
Arambourg’s collection (MNHN):
Arambourg and Piveteau (1929) list Mastodon pentelici from the localities Ravin X (R. X), Ravin des Zouaves, Ravin R. Ar., Ravin C, Ravin de Vatilük ou Ravin G (=Vathylakkos 3, VAT), Ravin de Konikovo (=DTK). Unfortunately, the authors do not provide details for the exact localities of the specimens. There is only one illustration (Arambourg and Piveteau 1929: pl. II, fig. 1) of a juvenile mandible bearing the dp4’s (MNHN-SLQ-2), which in the MNHN collection is labelled as originating from R. X. The symphysis is situated at the extension of the horizontal ramus, and the dimensions of the dp4’s are similar to the known Turolian ones. Additionally, a DP4 (MNHN-SLQ-3) labelled as coming from Ravin X is close metrically to the type material of C. pentelici from Pikermi, indicating a Turolian age. The rest of the R. X fauna also suggests a Turolian age for the locality (Koufos 2006a). Other proboscidean remains of the Arambourg’s collection lack any locality indication; however, they can be attributed to C. pentelici due to the dimensions of the teeth (Fig. 6) and the morphology of the DP3 (e.g. MNHN-SLQ-1122).
Nikiti localities
The fossiliferous localities of Nikiti (Chalkidiki Peninsula, Macedonia) were discovered in 1990 (Koufos et al. 1991). There are two localities, Nikiti 1 (NKT) and Nikiti 2 (NIK), which are located in the Nikiti Formation (Koufos in press and refs. cited). The NKT represents the oldest one, dated to the late Vallesian, MN 10 (Kostopoulos and Koufos 1999; Koufos 2000; in press), whereas NIK, situated about 20 m above the latter, is dated to the early Turolian (Kostopoulos and Koufos 1999; Vlachou and Koufos 2002). The locality NIK provided a rich mammalian fauna, the overall study of which is in progress. No proboscidean material is known from NKT, apart from some postcranial elements.
Locality Nikiti 2 (NIK):
The sole known proboscidean species from NIK is C. pentelici, represented by numerous juvenile dental remains. The morphology (Fig. 5c, f) and the size of the dp3/DP3 (Fig. 6) are typical for the Turolian choerolophodons. Moreover, several juvenile mandibles have the same morphological features as those of the type material of C. pentelici from Pikermi. Apart from these remains, an isolated tusk belonging to an adult individual was found about 100 m away from NIK at the same level. The tusk is long, strongly curved, lacks enamel and has circular cross section. The strong curvature clearly separates it from the straighter tusks of “Mammut” and Tetralophodon and suggests its attribution to Choerolophodon.
Samos localities
The mammal localities of Samos Island are among the richest Late Miocene ones, and several scientists have unearthed important material (e.g. E. Forsyth Major, B. Brown). Most specimens of the old collections lack stratigraphic indications, and their comparison and dating are thus difficult. During a recent field campaign, which started in 1993, numerous fossils have been unearthed, and extensive stratigraphic work has been carried out aiming at correlation of the various old and new fossiliferous sites (Koufos and Nagel 2009). The most recent stratigraphic interpretations for Samos are given by Kostopoulos et al. (2009). The fossiliferous localities of Samos are situated in the Mytilinii Formation, which is dated to Turolian. The proboscidean remains from the recent excavations come from two sites, Mytilinii-1A (MTLA) and Mytilinii-1B (MTLB), dated to the middle Turolian, MN 12 (Koufos et al. 2009).
Forsyth Major (1894), in his catalogue for the MGL collection of Samos, referred to the taxa Mastodon pentelici, Mastodon turicensis and Deinotherium sp., originating from “Adriano” site. A detailed study of the Samos choerolophodons was given by Schlesinger (1917, 1922), who invented the subgenus Choerolophodon and ascribed several juvenile skulls and mandibles to Mastodon (Choerolophodon) pentelici, which are housed at NHMW and HGI. Osborn (1936) described a partial skull belonging to C. pentelici, housed at AMNH. Later, Lehmann (1950) described some more material stored at BSPG, including two juvenile maxillary fragments of C. pentelici, as well as a skull and a mandible that were attributed to Mastodon longirostris [all the BSPG proboscidean material from Samos was destroyed during World War II (pers. communication Heissig 2011)]. Bernor et al. (1996) reported D. giganteum, M. borsoni, C. pentelici and Stegotetrabelodon grandincisivus (Schlesinger, 1917) from the Main Bone Beds of the Mytilinii Formation. Tassy (2005) mentioned that the latter authors listed S. grandincisivus instead of “T.” atticus (Wagner, 1857) for Pikermi, which might be the case also for Samos. Recently, Konidaris and Koufos (2009) described C. pentelici from MTLB and Zygolophodon turicensis possibly from MTLA.
Apart from the certain presence of C. pentelici in Samos, the more detailed studies during the past few years provided additional results. The collection housed at MGL includes the species C. pentelici and D. gigantissimum Stefanescu, 1892. The latter species is represented by a partial right mandible with the dp3 (MGL-S 1048) and the erupting dp4 (MGL-S 380), associated to each other and forming the horizontal ramus, and a left mandibular fragment with the unerupted dp4 (MGL-S 381). Possibly both specimens belong to the same individual, but this cannot be confirmed. The presence of Zygolophodon turicensis or any other mammutid in the MGL collection has not been confirmed. The size of the deinothere dp3 (57.6 × 42.3 mm) places it among the largest known so far from Europe, surpassing the Late Miocene dp3’s from Montredon (France, FSL-210393), Prottes (Austria, Huttunen 2002), Kayadibi (Turkey, Gaziry 1976) and Nessebar (Bulgaria, SU-190, 191). The only larger dp3 is known from the Turolian locality of Baltavár (Hungary, HGI-Ob-3150). The deinothere material from Nessebar was originally ascribed to Prodeinotherium by Bakalow (1914) and Bakalov and Nikolov (1962), but Markov (2004, 2008) records it as juvenile specimens of D. gigantissimum. Accepting the validity of D. gigantissimum, as demonstrated by Tarabukin (1974) and Markov (2008), and keeping in mind the large dimensions of the Samos specimen, we allocate it to this species.
As far as the tetralophodons from Samos housed at BSPG are concerned, their direct observation is impossible as they have been destroyed. Based on the description and illustration of Lehmann (1950: pl. 22, fig. 108), their attribution to “T.” atticus seems plausible [for the use of quotation marks for the generic name see Tassy (2005)]. It should be mentioned, however, that the morphology of the juvenile teeth of “Mastodon” grandincisivus [an amebelodont, which reached tetralophodonty in parallel to tetralophodont gomphotheres (Tassy 2005)] remains unclear. Moreover, this species shares with “T.” atticus the same geographic and biostratigraphic distribution, coexisting with it in several localities (Markov 2008). Therefore, a more detailed work is necessary for this taxonomic problem. Apart from the BSPG material, an unpublished juvenile left mandible is housed at NHMW (NHMW-V 49), labelled as C. pentelici. Unfortunately the juvenile dentition is inside the mandibular bone, preventing any observation of its morphology, but the mandible has a strong symphysis, which bears a lower juvenile tusk. The morphology of the mandible excludes any affinities with other elephantoids present at Samos (Choerolophodon, “Mammut”) and indicates similarities with “T.” atticus.
The other issue concerning the Samos proboscideans is the mammutid material. The only known published material is an isolated dp4 described by Konidaris and Koufos (2009). Direct studies of comparative juvenile material, including specimens of Z. turicensis from Simorre (France, MNHN-Si 11) and Innersdorf (Germany, BSPG-1916 I 1), the Pikermi material (MNHN-PIK-3613, NHMUK-M 10104) and “M.” borsoni from Valčedram (Bulgaria, SU) [this specimen was originally identified as Z. tapiroides by Bakalov and Nikolov (1962) but later as “M.” borsoni by Markov (2004)], permit the attribution of the Samos dp4 to “Mammut”, based on the clearly developed zygodont characters, the height of the crown in the interlophids on the lingual side [for this character see Tassy (1985)] as well as the similar dimensions to the dp4 from Pikermi (NHMUK). Therefore, the dp4 of Samos should be referred to as “Mammut” sp.
Biostratigraphic and palaeoecological remarks
The current knowledge on the Late Miocene proboscideans from Macedonia and Samos can provide useful biostratigraphic information. The species C. anatolicus, present in the locality Pentalophos, characterizes the early Vallesian (Fig. 7). No other locality of this age with large mammals is known from Greece (Koufos 2006a). During the late Vallesian, two proboscideans were present, C. pentelici and D. giganteum, associated at Ravin de la Pluie (Fig. 7; Table 1). The absence of a tetralophodont elephantoid can be attributed either to the scarcity of localities of this age or to limited sampling (which is not the case, since RPl has brought to light a rich collection) or to ecological reasons. The coexistence of D. giganteum and T. longirostris was common during the Vallesian of Central and Western Europe [Choerolophodon did not migrate to this area (Koufos 2003)], but very scarce in Southeastern Europe (where Choerolophodon is the dominant proboscidean species); for example, the Middle Sinap collection does also not include any tetralophodont elephantoid. However, in Yulafli (Turkey), Geraads et al. (2005) mark the coexistence of Choerolophodon, Tetralophodon and Deinotherium. The authors suggest a wet and probably forested landscape for the area, whereas Koufos (2006b) considers more open conditions during the whole Vallesian for the more southern part of Europe. It appears possible that different ecological conditions during this period prohibited the migration of tetralophodons to South and of choerolophodons to Central and Western Europe.

Biostratigraphic distribution of the Late Miocene proboscidean taxa of Greece mentioned in the text. The “?” in “T.” atticus and D. gigantissimum is placed because both species are known from the old collections of Samos, thus indicating a general Turolian age. However, “T.” atticus is known from Pikermi (type locality) dated to MN 12
During the Turolian, the typical form of C. pentelici is present at all Greek localities (Fig. 7; Table 1) and is by far the most abundant proboscidean species, indicating that it was well adapted to this region. The dental microwear of Choerolophodon from Axios Valley is under study and is expected to provide more data on the dietary preferences of this proboscidean. The situation is more “balanced” in the Pikermi proboscidean assemblage, where more material and taxa are present (the material housed at AMPG is under study). In RZO, C. pentelici coexisted with “Mammut” (Table 1). At Samos, C. pentelici, “Mammut” sp., “T.” atticus and D. gigantissimum were present. Noticeable is the absence of “T.” atticus and Deinotherium at the Turolian localities of the Axios Valley. “T.” atticus is known from the middle Turolian localities of Pikermi (Wagner 1857, Gaudry 1862) and Akkaşdaği (Tassy 2005). Furthermore, the coexistence of Choerolophodon, “T.” atticus and Deinotherium is known from several Turolian localities, including the close geographically localities of Bulgaria (e.g. Hadjidimovo) (Markov 2008 and refs. cited), indicating that these species occupied the wider region of Southeastern Europe. Therefore, most probably, the absence of “T.” atticus and Deinotherium from the Turolian localities of the Axios Valley can be attributed to the limited sampling, since the proboscidean specimens of the middle Turolian localities Prochoma and Vathylakkos are rather few, particularly when compared with the rich samples of Pikermi, Samos and Hadjidimovo. In any case, although proboscideans seem to be a significant palaeoecological indicator, their remains are usually scarce, and the absence of a fossil taxon does not necessarily mean that it was not included in the fauna.
All these Turolian species did not survive until the Pliocene. Schmidt-Kittler et al. (1995) list C. pentelici from Maramena (Serres Basin, Greece), which is dated, according to these authors, at the Turolian/Ruscinian boundary (MN 13/MN 14). This is probably one of the last occurrences of Choerolophodon pentelici.
Results and conclusion
The preliminary study of the proboscideans from the Late Miocene of Macedonia and Samos Island provided the following results:
-
The study of choerolophodons from the Axios Valley localities (which cover the whole Late Miocene) permitted the recognition of different evolutionary stages for the genus Choerolophodon. Moreover, the distinction at specific level was confirmed with the most primitive form belonging to C. anatolicus and the advanced one to C. pentelici. This allowed the taxonomic revision of the Late Miocene peri-Mediterranean choerolophodons.
-
At all studied localities, the dominant proboscidean is Choerolophodon. C. anatolicus was recognized at Pentalophos and characterizes the early Vallesian (MN 9). The most primitive form of C. pentelici is recognized at Xirochori, Ravin de la Pluie and Ravin des Zouaves 1 (late Vallesian, MN 10). The typical and advanced form of C. pentelici is traced at the localities Ravin des Zouaves 5, Vathylakkos 2, Prochoma, Dytiko 2, 3, Samos (old collections), Mytilinii-1B and Nikiti 2 (Turolian). Consequently, C. pentelici characterizes the late Vallesian-Turolian (MN 10-MN 13).
-
The mammutid material from Ravin des Zouaves 5 (early Turolian, MN 11) and Mytilinii-1A (middle Turolian, MN 12) is attributed to “Mammut” sp.
-
“T.” atticus is rare and recognized only in Samos (old collections), thus only a general Turolian age can be assumed for the species. The type locality of the species, Pikermi, is dated to middle Turolian.
-
The deinotheres are represented by two species. Deinotherium giganteum was traced at the late Vallesian (MN 10) locality Ravin de la Pluie. Deinotherium gigantissimum is present only at Samos (old collections), thus only a general Turolian age can be assumed for the species.
-
During the Vallesian, three proboscidean species were present: C. anatolicus during the early Vallesian, and C. pentelici and D. giganteum during the late Vallesian. The absence of Tetralophodon at the Vallesian localities of Greece can probably be attributed to ecological reasons. The open environments of Southeastern Europe prohibited the migration of Tetralophodon in the area, whereas the more closed ecological conditions of Western and Central Europe were favourable for this elephantoid.
-
During the Turolian, four proboscidean species were recognized: C. pentelici, “Mammut” sp., “T.” atticus and D. gigantissimum. The absence of the two latter species from the Turolian localities of Axios Valley is possibly attributed to limited sampling, as they are present in the neighbouring areas (e.g. Pikermi, Samos, Hadjidimovo).
New material from the broader region is needed to clarify the taxonomy, evolution and relationships, especially of the rarer proboscidean taxa. The study of the Pikermi material (AMPG) will probably shed more light on these issues. Moreover, several palaeoecological methods should be applied (e.g. dental microwear, isotope analysis) to understand the ecological niche of these proboscideans.
Abbreviations
- AMNH:
-
American Museum of Natural History (New York, USA)
- AMPG:
-
Athens Museum of Palaeontology and Geology, National and Kapodistrian University of Athens (Greece)
- BSPG:
-
Bayerische Staatssammlung für Paläontologie und Geologie, München (Germany)
- FSL:
-
Faculté des Sciences de Lyon (France)
- GPIH:
-
Geologisch-Paläontologisches Institut, Hamburg (Germany)
- HGI:
-
Hungarian Geological Institute (Budapest)
- LGPUT:
-
Laboratory of Geology and Palaeontology, Aristotle University of Thessaloniki (Greece)
- MGL:
-
Musée Cantonal de Géologie, Lausanne (Switzerland)
- NHMA:
-
Natural History Museum of the Aegean, Mytilinii (Samos Island, Greece)
- NHMUK:
-
Natural History Museum United Kingdom (London)
- NHMW:
-
Naturhistorisches Museum Wien (Austria)
- MNHN:
-
Muséum National d’Histoire Naturelle (Paris, France)
- SU:
-
Palaeontology Museum of the Sofia University (Bulgaria)
References
Arambourg, C., and J. Piveteau. 1929. Les Vertébrés du Pontien de Salonique. Annales de Paléontologie 18: 59–138.
Bakalov, P., and I. Nikolov. 1962. Les fossiles de Bulgarie. X. Mammifères tertiaires. Sofia: Académie des Sciences de Bulgarie.
Bakalow, P. 1914. Beiträge zur Paläontologie Bulgariens. II. Dinotheriumreste aus Bulgarien. Annuaire de l’Université de Sofia 8–9: 1–29.
Bernor, R.L., N. Solounias, C.C. Swisher, and J.A. van Couvering. 1996. The correlation of three classical “Pikermian” mammal faunas—Maragheh, Samos and Pikermi—with the European MN unit system. In The evolution of Western Eurasian Neogene mammal faunas, ed. R.L. Bernor, V. Fahlbusch, and H.W. Mittmann, 137–154. New York: Columbia University Press.
de Bonis, L., G. Bouvrain, B. Keraudren, and J. Melentis. 1973. Premiers résultats des fouilles récentes en Grèce septentrionale (Macédoine). Comptes Rendus de l’Académie des Sciences Paris 277(D): 1431–1434.
de Bonis, L., G. Bouvrain, D. Geraads, G.D. Koufos, and S. Sen. 1994. The first aardwarks from the Miocene of Macedonia (Greece). Neues Jahrbuch für Geologie und Paläontologie Monatshefte 194: 343–360.
de Bonis, L., and G.D. Koufos. 1999. The Miocene large mammal succession in Greece. In Hominoid evolution and climatic change in Europe, vol 1, The evolution of Neogene terrestrial ecosystems in Europe, ed. J. Agusti, L. Rook, and P. Andrews, 205–237. New York: Cambridge University Press.
de Bonis, L., and G. Bouvrain. 2003. Nouveaux Giraffidae du Miocène supérieur de Macédoine (Grèce). In Advances in Vertebrate Paleontology “Hen to Panta”, ed. A. Petrulescu, and E. Stiucă, 5–16. Bucharest: Romanian Academy-“Emil Racoviţă” Institute of Speleology.
de Bruijn, H., R. Daams, G. Daxner-Höck, V. Fahlbusch, L. Ginsburg, P. Mein, J. Morales, et al. 1992. Report of the RCMNS working group on fossil mammals, Reisensburg 1990. Newsletters on Stratigraphy 26(2/3): 65–118.
Forsyth Major, C.J. 1894. Le gisement ossifère de Mytilini et catalogue d’ossements fossiles. Lausanne: Georges Bridel & Cie Editeurs.
Fortelius, M. (coordinator) 2011. Neogene of the Old World Database of Fossil Mammals (NOW), http://www.helsinki.fi/science/now/. University of Helsinki.
Gaudry, A. 1862. Animaux fossiles et géologie de l’Attique. Paris: Savy.
Gaudry, A. 1867. Animaux fossiles et géologie de l’Attique. Atlas. Paris: Savy.
Gaudry, A., and E. Lartet. 1856. Résultats des recherches paléontologiques entreprises dans l’Attique sous les auspices de l’Académie. Comptes Rendus de l’Académie des Sciences, Paris 43: 271–274.
Gaziry, A.W. 1976. Jungtertiäre Mastodonten aus Anatolien (Türkei). Geologisches Jahrbuch 22: 3–143.
Geraads, D. 1989. Un nouveau Giraffidé du Miocène supérieur de Macédoine (Grèce). Bulletin du Muséum national d’Histoire naturelle, 4ème sér 11(C, 4): 189–199.
Geraads, D., T. Kaya, and S. Mayda. 2005. Late Miocene large mammals from Yulafli, Thrace region, Turkey, and their biogeographic implications. Acta Palaeontologica Polonica 50(3): 523–544.
Gräf, I.E. 1957. Die Prinzipien der Artbestimmung bei Dinotherium. Palaeontographica 108(5–6): 131–185.
Huttunen, K. 2002. Deinotheriidae (Proboscidea, Mammalia) dental remains from the Miocene of Lower Austria and Burgenland. Annalen des Naturhistorischen Museums in Wien 103 A: 251–285.
Kappelman, J., A. Duncan, M. Feseha, J.P. Lunkka, D. Ekart, F. McDowell, T.M. Ryan, and C.C. Swisher III. 2003. Chronology. In Geology and Paleontology of the Miocene Sinap Formation, Turkey, ed. M. Fortelius, J. Kappelman, S. Sen, and R.L. Bernor, 41–66. New York: Columbia University Press.
Konidaris, G.E., and G.D. Koufos. 2009. The Late Miocene Mammal Faunas of the Mytilinii Basin, Samos Island, Greece: New Collection. 8. Proboscidea. In The late Miocene mammal faunas of Samos, ed. G.D. Koufos, and D. Nagel, 139–155. Wien: Beiträge zur Paläontologie.
Korotkevich, E.L. 1988. History of the formation of the Hipparion fauna of Eastern Europe. Kiev: Naukova Dumka.
Kostopoulos, D.S., and G.D. Koufos. 1999. The Bovidae (Mammalia, Artiodactyla) of the “Nikiti-2” [NIK] faunal assemblage (Chalkidiki peninsula, N. Greece). Annales de Paléontologie 85(3): 193–218.
Kostopoulos, D.S., G.D. Koufos, I.A. Sylvestrou, G.E. Syrides, and E. Tsombachidou. 2009. The late Miocene mammal faunas of the Mytilinii Basin, Samos Island, Greece: New Collection. 2. Lithostratigraphy and fossiliferous sites. In The late Miocene mammal faunas of Samos, ed. G.D. Koufos, and D. Nagel, 13–26. Wien: Beiträge zur Paläontologie.
Koufos, G.D. 1980. Palaeontological and stratigraphical study of the continental Neogene deposits of Axios Basin. PhD, University of Thessaloniki, Thessaloniki.
Koufos, G.D. 2000. New material of Vallesian hipparions (Mammalia, Perissodactyla) from the lower Axios valley, Macedonia, Greece. Senckenbergiana Lethaea 80: 231–255.
Koufos, G.D. 2003. Late Miocene mammal events and biostratigraphy in the Eastern Mediterranean. In Distribution and migration of Tertiary Mammals in Eurasia. A volume in honour of Hans de Bruijn, ed. J.W.F. Reumer, and W. Wessels, 343–371. Rotterdam: Deinsea.
Koufos, G.D. 2006a. The Neogene mammal localities of Greece: Faunas, chronology and biostratigraphy. Hellenic Journal of Geosciences 41: 183–214.
Koufos, G.D. 2006b. Palaeoecology and chronology of the Vallesian (late Miocene) in the Eastern Mediterranean region. Palaeogeography, Palaeoclimatology, Palaeoecology 234: 127–145.
Koufos, G.D. (in press). Neogene mammal biostratigraphy and chronology of Greece. In Asian neogene mammal biostratigraphy and chronology, ed. W. Xiaoming, M. Fortelius, and L. Flynn. New York: Columbia University Press.
Koufos, G.D., G.E. Syrides, K.K. Koliadimou, and D.S. Kostopoulos. 1991. Un nouveau gisement de vertébrés avec hominoïde dans le Miocene supérieur de Macédoine (Gréce). Compte Rendus de l’Académie des Sciences Paris 313(II): 691–696.
Koufos, G.D., and D. Nagel. 2009. The late Miocene mammal faunas of Samos. Wien: Beiträge zur Paläontologie.
Koufos, G.D., D.S. Kostopoulos, and T.D. Vlachou. 2009. The late Miocene mammal faunas of the Mytilinii Basin, Samos Island, Greece: New Collection. 16. Biochronology. In The late Miocene mammal faunas of Samos, ed. G.D. Koufos, and D. Nagel, 397–408. Wien: Beiträge zur Paläontologie.
Lehmann, U. 1950. Über Mastodonreste in der Bayerischen Staatssammlung in München. Palaeontographica 99(A): 121–228.
Markov, G.N. 2004. The fossil proboscideans of Bulgaria and the importance of some Bulgarian finds—A brief review. Historia naturalis bulgarica 16: 139–150.
Markov, G.N. 2008. The Turolian proboscideans (Mammalia) of Europe: Preliminary observations. Historia naturalis bulgarica 19: 153–178.
Mein, P. 1990. Updating MN zones. In European neogene mammal chronology, ed. E.H. Lindsay, V. Falhbusch, and P. Mein, 73–90. New York: Plenum.
Melentis, J.K. 1967. Studien über fossile Vertebraten Griechenlands. 19. Die Pikermi-Fauna von Halmyropotamos (Euboä, Griechenland), 1. Teil: Odontologie und Kraniologie. Annales Géologiques des Pays Helléniques 19: 283–411.
Nikolov, I., and D. Kovačev. 1966. Pliozäne Säugetierfauna aus Assenovgrad. Travaux sur la géologie de Bulgarie - série paléontologie 8: 131–142.
Osborn, H.F. 1936. Proboscidea. A monograph of the discovery, evolution, migration and extinction of the mastodonts and elephants of the world, vol I: Moeritherioidea, Deinotherioidea, Mastodontoidea. New York: American Museum Press.
Ozansoy, F. 1965. Étude des gisements continentaux et des mammifères du Cénozoïque de Turquie. Mémoires de la Société Géologique de France 102: 1–92.
Sanders, W.J. 2003. Proboscidea. In Geology and Paleontology of the Miocene Sinap Formation, Turkey, ed. M. Fortelius, J. Kappelman, S. Sen, and R.L. Bernor, 202–219. New York: Columbia University Press.
Schlesinger, G. 1917. Die Mastodonten des K.K. Naturhistorischen Hofmuseums. Denkschrift des Naturhistorischen Hofmuseums, Geologisch – Paläontologische Reihe 1: 1–230.
Schlesinger, G. 1922. Die Mastodonten der Budapester Sammlungen. Geologica Hungarica 2(1): 1–284.
Schmidt-Kittler, N., H. de Bruijn, and C. Doukas. 1995. General introduction. In The vertebrate locality of Maramena (Macedonia, Greece) at the Turolian-Ruscinian boundary (Neogene), ed. N. Schmidt-Kittler, 9–18. München: Münchner Geowissenschaftliche Abhandlungen.
Şenyürek, M.S. 1952. A study of the Pontian fauna of Gökdere (Elmadaği), south-east of Ankara. Türk Tarih Kurumu Belleten 16: 449–492.
Sickenberg, O., J.D. Becker-Platen, L. Benda, D. Berg, B. Engesser, W. Gaziry, K. Heissig, et al. 1975. Die Gliederung des höheren Jungtertiärs und Altquartärs in der Türkei nach Vertebraten und ihre Bedeutung für die internationale Neogen-Stratigraphie. Geologisches Jahrbuch 15: 1–167.
Tarabukin, B.A. 1974. New data on the systematics, phylogeny and ecology of Suborder Deinotherioidea Osborn (1921). In Mammals of the late Cenozoic from Southwestern USSR, ed. A.I. David, 77–90. Shtiinca: Chişinău.
Tassy, P. 1983. Les Elephantoidea Miocènes du Plateau du Potwar, Groupe de Siwalik, Pakistan. IIe Partie: Choerolophodontes et Gomphothères. Annales de Paleontologie 69(3): 235–297.
Tassy, P. 1985. La place des mastodontes Miocènes de l’Ancien Monde dans la phylogénie des Proboscidea (Mammalia): Hypothèses et conjectures. PhD, Université Pierre et Marie Curie, Paris.
Tassy, P. 1986. Nouveaux Elephantoidea (Mammalia) dans le Miocène du Kenya. Paris: CNRS.
Tassy, P. 1994a. Les gisements de mammifères du Miocène supérieur de Kemiklitepe, Turquie: 7. Proboscidea (Mammalia). Bulletin du Muséum National d’Histoire Naturelle 16(1): 143–157.
Tassy, P. 1994b. Gaps, parsimony, and early Miocene elephantoids (Mammalia), with a re-evaluation of Gomphotherium annectens (Matsumoto, 1925). Zoological Journal of the Linnean Society 112: 101–117.
Tassy, P. 2005. Proboscideans (Mammalia) from the late Miocene of Akkaşdaği, Turkey. In Geology, mammals, environments at Akkaşdaği, late Miocene of Central Anatolia, vol. 4, ed. S. Sen, 707–714. Paris: Geodiversitas.
Tassy, P., S. Sen, J.J. Jaeger, J.M. Mazin, and N. Dalfes. 1989. Une sous-espèce nouvelle de Choerolophodon pentelici (Proboscidea, Mammalia) à Esme Akcaköy, Miocène supérieur d’Anatolie occidentale. Comptes Rendus de l’Académie des Sciences 309: 2143–2146.
Tobien, H. 1988. Contributions a l’etude du gisement miocene superieur de Montredon (Herault). Les grands mammiferes. 7—les proboscidiens Deinotheriidae. Palaeovertebrata Mémoire extraordinaire 1988: 135–175.
Vlachou, T., and G.D. Koufos. 2002. The hipparions (Mammalia, Perissodactyla) from the Turolian locality “Nikiti-2” (NIK), Macedonia, N. Greece. Annales de Paléontologie 88(4): 215–263.
Wagner, A. 1857. Neue Beiträge zur Kenntnis der fossilen Säugerthier-Ueberreste von Pikermi. Abhandlungen der königlich bayerischen Akademie der Wissenschafen 8: 109–158.
Acknowledgments
G. Konidaris would like to thank the following persons for allowing access to collections and offering help during his stay at the various museums and institutions: Prof. P. Tassy (MNHN), Dr. U. Göhlich (NHMW), Dr. A. Currant and Dr. C. Delmer (NHMUK), Dr. G. Rössner (BSPG), Dr. U. Kotthoff (GPIH), Prof. L. Kordos (HGI), Dr. M. Gasparik (HNHM Budapest), Dr. R. Marchant (MGL), Dr. M. Ivanov (SU) and A. Prieur (Université Lyon 1). Always valuable are the conversations with Assist. Prof. D. Kostopoulos (LGPUT), Lecturer S. Roussiakis (Univ. of Athens), Dr. A. Athanassiou (Hellenic Ministry of Culture), Prof. P. Tassy (MNHN) and Dr. G. Markov (NMNH Sofia). Special thanks go to Prof. G. Theodorou (Univ. of Athens) for the opportunity to study the Pikermi collection and to participate in the new Pikermi excavation series, and also to Dr. S. Sen for the wholehearted hospitality during his stay in Paris. Financial support was given by SYNTHESYS (European Union-funded Integrated Activities grant) for visits of the first author to Vienna (AT-TAF-3825), London (GB-TAF-864) and Budapest (HU-TAF-1683) and by the international collaborative grant PICS CNRS 5182 (CNRS & Aristotle University of Thessaloniki; PIs: G. Merceron & D. Kostopoulos) for visits to Lyon and Lausanne. We would like to thank Prof. P. Tassy for the comments on a premature version of this article, Dr. K. Vasileiadou (Natural History Museum of the Lesvos Petrified Forest) for the linguistic improvement of the text and all colleagues who participated in the excavations and prepared fossils all these years. Many thanks go to Dr. U. Göhlich and one anonymous reviewer for their useful comments on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Appendix
Appendix
See Table 2.
Rights and permissions
About this article
Cite this article
Konidaris, G.E., Koufos, G.D. Late Miocene Proboscidea (Mammalia) from Macedonia and Samos Island, Greece: preliminary results. Paläontol Z 87, 121–140 (2013). https://doi.org/10.1007/s12542-012-0147-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12542-012-0147-z