
Abstract
Senghor Seamount is an important fishing ground around the Cape Verde archipelago in the Eastern Central Atlantic. On an experimental field survey in October 2009 and December 2011, a total of 115 deep-sea fishes of 26 species belonging to 18 families were caught on the seamount summit, along the slopes and on the adjacent abyssal plane, using longlines, fish traps, beam trawl and otter trawl. Here we report seven new records for the Cape Verde deep-sea fish fauna. Most species are known from other areas of the Atlantic Ocean, but our findings are an important contribution to our understanding of the distribution of deep-water fish species. The co-occurrence of northern and southern Atlantic ichthyofauna components at Senghor Seamount, and the Cape Verde area in general, can be attributed to the large-scale hydrographic regime with two water masses merging at the Cape Verde Frontal Zone, the North Atlantic Central Water and the South Atlantic Central Water.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Seamounts are underwater mountains, globally distributed in the ocean basins (Wessel et al. 2010), often showing higher diversity than the surrounding abyssal plain (Shank 2010; Stocks et al. 2012). Seamounts are also known to provide habitat and foraging ground for fishes, including commercial species. Several studies have characterized the seamount ichthyofauna in regions of the temperate NE Atlantic (e.g., Pakhorukov 2008; Christiansen et al. 2009; Menezes et al. 2009, 2012; Christiansen et al. 2014). Knowledge of the distribution patterns of bathyal and abyssal fish species, however, is generally poor. In the eastern Atlantic, most studies have focused on the boreal and subtropical regions (e.g., Gordon and Duncan 1987; Merrett 1987; Haedrich and Merrett 1988; Merrett et al. 1991; Merrett 1992; Gordon et al. 1996; Martin and Christiansen 1997), and only scattered information is available from the tropics (e.g., Merrett and Marshall 1981; Merrett 1992; Vakily et al. 2002; Vieira et al. 2013).
Oceanographic research off the Cape Verde islands began in the mid-nineteenth century, and studies of the archipelago’s wildlife are still continuing. Some of the original work on marine fishes from Cape Verde was conducted by Franca and Vasconcelos (1962) and Lloris et al. (1991). Reiner (1996) and Reiner (2005) reviewed the ichthyofauna of the Cape Verde Islands, while Brito et al. (2006), Monteiro et al. (2008), Almeida et al. (2010) and González et al. (2010, 2014) added new information on the marine fish communities of the archipelago. A compilation of data on coastal fish species, complementing the knowledge on the ichthyofauna from the Cape Verde archipelago with zoogeographic remarks, was provided by Wirtz et al. (2013) and Freitas (2014), and Hanel and John (2015) compiled data specifically on mesopelagic fishes. However, information on deep-sea species remains scarce.
Benthopelagic fishing surveys off the Cape Verde archipelago were conducted during the 1980s and 1990s to investigate stock sizes of commercially important species along the islands’ shelves and the adjacent seamounts (e.g., Magnússon and Magnússon 1985, 1987; Thorsteisson et al. 1995). More recently, Menezes et al. (2004) investigated the demersal fish communities from several locations in the Cape Verde archipelago, including Senghor Seamount, but provided limited data on deeper-reaching species. Hanel et al. (2010) contributed valuable insight on the larval fish communities from the seamount, which were dominated by lanternfishes (Myctophidae).
The Senghor Seamount, known locally as Nova Holanda Bank, is an important fishing area for the regional longline fishery and also supports a small-scale crab and lobster fishery (MAAP/GEP 2004). Recent ROV surveys have revealed that the seamount features various habitats for benthopelagic and benthic fishes, including soft bottom and hard substrate with rich coral cover (Christiansen et al. 2011). Hanel et al. (2010) reported evidence of a seamount effect related to the topographic features and oceanographic conditions in the region. Denda and Christiansen (2014), however, detected no current topography-related effect on the distribution and concentration of mesozooplankton as potential prey for seamount-associated fishes at Senghor Seamount. No evidence was found for enhanced food availability at the seamount, such as higher concentrations of zooplankton biomass, compared to the surrounding ocean. Nevertheless, Senghor Seamount provides special habitats for benthic organisms, and hosts diverse and abundant communities of sponges, corals, hydroids, polychaetes, crustaceans and echinoderms (Christiansen et al. 2011; Chivers et al. 2013), and may be an attractive location in the open ocean for commonly dispersed oceanic pelagic fishes as well as for typical shelf species.
Official data about the type and effort of fishing exerted on the Cape Verde offshore seamounts is scarce. The artisanal fleets contribute to most captures in the region (D’Oliveira Fonseca 2000; Benchimol et al. 2009), but are limited to the near-shore areas. However, the extent of the impact caused by industrial fisheries, particularly if international fishing vessels are included, is likely underestimated (MAAP/GEP 2004; Benchimol et al. 2009). Taking this into account, it is important to evaluate the deep-sea resources in order to characterize the role and conservation status of the seamounts around the Cape Verde archipelago.
The main objective of this study, therefore, was to extend the knowledge of the deep-water fish fauna of Cape Verde and Senghor Seamount, at different depths, including the summit plateau, the slopes and the adjacent abyssal plain.
Material and methods
Study site
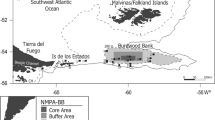
Senghor Seamount is an isolated seamount located at 17°12′N, 021°57′W ca. 60 nmi northeast of the island of Sal, Cape Verde (Fig. 1a), within the sphere of the NE Atlantic tropical gyre (e.g., Mittelstaedt 1991; Lathuilière et al. 2008). The hydrographic regime is characterized by the North Equatorial Current system and Mauritanian upwelling filaments (Mason et al. 2011), but also by northward-flowing South Atlantic Central Water (SACW), merging in the Cape Verde Frontal Zone (CVFZ, e.g., Zenk et al. 1991). Senghor Seamount is nearly circular in shape, with a base diameter of ca. 40 km, steep slopes and a small elliptical summit plateau, which is about 2.5 km wide and 5 km long. Water depth at the base of the seamount is about 3300 m; the minimum summit depth is 90 m. Video footage from the summit and upper slopes showed that the substrate structure varies between sediment-covered areas and rocky outcrops (Christiansen et al. 2011). The sediment-dominated reference station (RS) on the abyssal plain was located ca. 60 nm north of Senghor Seamount, at 18°05′N, 022°00′W, at a water depth of about 3300 m, which corresponds to the depth of the base of the seamount.

a Cape Verde archipelago and the location of the Senghor Seamount (triangle) and abyssal sampling stations (RS reference station). b Central area of Senghor Seamount with locations of beam trawl tracks and longline deployments during cruises M79/3 and P423
Sampling
During the cruises M79/3 of the RV Meteor in September/October 2009 and P423 of the RV Poseidon in December 2011, fishes were sampled at distinct depths on the summit plateau and along the slopes of Senghor Seamount and the adjacent bathyal plain (Fig. 1b, Table 1). Depending on the area, various types of fishing gear were used, including demersal longline, fish traps, beam trawl and otter trawl, aiming primarily at demersal fishes, but pelagic species were collected as well during slacking and heaving of the non-closing gears (Table 1).
The longline sets were used on the summit and on the upper slopes of the seamount at several depths ranging from 101 to 950 m. A total of 100 hooks of different shapes and sizes were attached to the main line by gangions approximately 60 cm in length and separated by approximately 2.5 m. The hooks were baited with squid. On seven of the longline sets, a fish trap baited with squid was also deployed at the end of the longline where the main anchor rope was attached.
The beam trawl (2 m beam length) was used to sample demersal fishes on the seamount summit at a depth of 103 m and on the upper slope at a depth of 260 m. The trawl was equipped with a 6-mm mesh net. The otter trawl (headrope length 15 m, mesh size 44 mm in the front part and 37 mm in the intermediate part, with 13-mm inner liner in the codend) was used in deep waters at the reference station, at ca. 3300 m.
Immediately after recovery of the catch, all fishes were identified to the lowest possible taxonomic level using Whitehead et al. (1984–1986), and were measured to total and standard length, respectively, and wet weight (TW). Stomachs and gonads of selected species were dissected, and tissue samples for DNA and isotopic ratios were taken for further analysis. Fishes from cruise P423 were discarded at sea due to a lack of freezing facilities during transport, whereas all fishes from M79/3 were frozen at −20 °C and transferred to the laboratory at the University of Hamburg to verify onboard identification. Here, the onboard identification was checked as far as possible (see Table 2). Rare specimens will be included in the collections of the Zoological Museum Hamburg.
Results
We provide a list of 115 demersal fishes belonging to 26 species (excluding unidentified) in 18 families (Table 2). Two species in two families were Chondrichthyes, all others were Teleostei, Actinopterygii. Additionally, several pelagic fishes (44 specimens) were caught in the otter trawl during its descent or ascent through the water column, including members of families Nemichthyidae (1 species), Sternopthychidae (5 species and 2 unidentified), Phosichthyidae (1 species), Stomiidae (5 species and several unidentified specimens), Myctophidae (3 species), Melanocetidae (1 species), Melamphaidae (2 species). These small mesopelagic fishes are not included in the list presented here.
Most of the bathyal fishes belonged to the family Macrouridae, making up 46 % of all fishes caught in the otter trawl, followed by Alepocephalidae (16 %), Ophidiidae (16 %) and Ipnopidae (11 %). At the slopes and summit of the seamount, Synaphobranchidae and Sebastidae were the most frequently caught fishes. The most species-rich families were the Ophidiidae (four species) and Ipnopidae (three species), but since among the macrourids many specimens could not be identified to species level the number of three species representing the family is likely to be underestimated. The most common identified species at bathyal depths were the abyssal grenadier Coryphaenoides armatus (Family Macrouridae) and the slimehead Bathytroctes macrolepis (Family Alepocephalidae), with 15 and 13 individuals, respectively. It is particularly worth mentioning that 9 individuals of the grenadier Coryphaenoides armatus were adult females, with large ovaries containing ripening oocytes.
Top predator species collected in the longline sets included the elasmobranchs smooth lanternshark Etmopterus pusillus, the sharpnose sevengill shark Heptranchias perlo and bony fishes such as the escolar Promethichthys prometheus. Some commercially important species were captured on the summit and slopes with the longlines, including the splendid alfonsino Beryx splendens and the blackbelly rosefish Helicolenus dactylopterus. The H. dactylopterus collected on the summit with the beam trawl were much smaller in size (8.4–13.2 cm TL and 10–25 g TW; N = 3) than those caught on the slopes with longlines (35.5–44.0 cm TL and 660–1390 g TW; N = 6).
Discussion
All species encountered in our surveys are known from other areas of the Atlantic Ocean, but several are new for the Cape Verde region, thus contributing to a better understanding of the distribution of deep-water fish species.
Senghor Seamount features an ichthyofauna that includes both northern and southern elements. In the depth range influenced by SACW (down to 700 m), we captured a total of eight species. Two of these were caught for the first time at this seamount: the red eel Myroconger compressus and the deep-body boarfish Antigonia capros. Both species have been reported in the coastal waters of the Cape Verde archipelago but not in the offshore area (Strømme et al. 1982; Menezes et al. 2004), supporting the importance of seamounts for the oceanic dispersal of typical shelf species between continental slopes or archipelagos. Commercially important species in the catches included Helicolenus dactylopterus and Beryx splendens. The bottom-living Helicolenus dactylopterus was found over a wide depth range from the summit to the mid-slope (100–734 m) of Senghor Seamount. Individuals were larger at the deep stations, suggesting a size segregation of this species around the seamount, as was also observed by Morales-Nin et al. (2003) and Colloca et al. (2010). However, the number of specimens caught in our samples is too small for final conclusions, and gear selection may also play a role. Beryx splendens is frequently found at seamounts and on the continental slopes in the NE Atlantic (Paxton 1999; Pakhorukov 2008; Menezes et al. 2009) in layers of the North Atlantic Central Water (NACW), but also deeper in the saltier waters of the Mediterranean Outflow (600–1300 m, see Christiansen et al. 2014).
Most of the demersal fishes caught at Senghor are typical of the NE Atlantic seamounts and shelfs, for example, Etmopterus pusillus, Helicolenus dactylopterus, Beryx splendens and Antigonia capros. Others, such as Myroconger compressus and Coloconger cadenati, have not been recorded at the more northern seamounts (see Christiansen et al. 2014) and can be regarded as southern species. Similarly, Promethichthys prometheus has not been reported from the NE Atlantic Seine and Horseshoe seamounts, but has been caught at seamounts south of the Azores (Great Meteor, Ewing, Jer; Shcherbachev et al. 1985). It is known from landings in the Azores (Santos et al. 1997) and Morocco (Iglésias 2013), and spawning has been observed in waters around Madeira (Nakamura and Parin 1993). Other southern species have been reported by Menezes et al. (2004) for Senghor Seamount. The mixture of temperate- and warm-water species can be attributed to the large-scale hydrographic regime with two water masses merging at the CVFZ, NACW and SACW (Pérez-Rodríguez et al. 2001). The water mass distribution indicates that Senghor Seamount and the reference site lie within the so-called shadow zone, south of the CVFZ (see Tomczak 1981; Zenk et al. 1991; Pierre et al. 1994), but the CVFZ is a strongly meandering feature and may reach near Senghor Seamount at times (Lázaro et al. 2005). At both sampling sites, temperature and salinity were characteristic for SACW (9.7–15.2 °C; 35.2–35.7) between 150 and 600 m, lying below a warm mixed surface layer of about 30 m depth, followed by a thermocline (Fig. 2, see Denda and Christiansen 2014). Between 700 and 1200 m, an intermediate layer of minimum salinity (34.92–34.98) corresponds to the Antarctic Intermediate Water (AAIW); underneath, North Atlantic Deep Water (NADW) lies between 1200 and 3300 m (6.1–2.5 °C; 34.9–35.0). In the lower bathyal and abyssal water layers, a temperature of about 2.0 °C would be the common indicator for the Antarctic Bottom Water (AABW, Pierre et al. 1994).

Profiles of temperature (°C), salinity (PSU) and oxygen concentration (ml l−1) at the reference site of Senghor Seamount (RS reference station)
The fishes collected on the bathyal plain adjacent to Senghor Seamount are typical deep-sea species, but to the best of the authors’ knowledge, most have never before been reported from Cape Verde waters. First registered records in the Cape Verde region include Aldrovandia rostrata, Ipnops murrayi, Acanthonus armatus, Bassozetus levistomatus, Bassozetus taenia, Barathronus cf. parfaiti (Fig. 3). These species are generally rare, but all are known in the subtropical and tropical eastern Atlantic, usually north of 20°N (Merrett and Marshall 1981; Merrett 1992; Nielsen 1990; Nielsen and Merrett 2000), and most of the species belong to one or more of the three latitudinally distinct deep-sea fish assemblages of the eastern North Atlantic described by Merrett (1992). Among the three assemblages, the composition of the dominant species changes, and species richness increases from north to south (Merrett and Fasham 1998), with increasing importance of Ophidiidae and Ipnopidae, and decreasing importance of Macrouridae. Coryphaenoides armatus, for example, is the dominant species in the northerly assemblage, becoming less important south of 38°N in the waters off Morocco and NW Africa (Haedrich and Merrett 1988; Merrett 1992). It was also the dominant species in trawl catches from the Iceland Basin at 59°N, 020°W, accounting for ∼60 % of the total catch, whereas at 47°N, 020°W, the species had a lower share of ∼30 % (Martin and Christiansen 1997). In the southerly assemblage described by Merrett (1992), C. armatus made up only 4 %; however, in our samples on the bathyal plain adjacent to Senghor Seamount, it was the most abundant species, at 46 %, which is consistent with baited camera observations by Henriques et al. (2002) on the Cape Verde terrace (∼120 nm SE of our abyssal station) and the Cape Verde abyssal plain (∼210 nm SE of our abyssal station), which showed C. armatus as the main species attracted to bait. Ophidiidae, Alepocephalidae and Ipnopidae, on the other hand, represented a substantial share of our samples, making up 43 % of all deep-water fishes. Christiansen and Martin (2000) concluded that high relative abundance of Ophidiidae and Ipnopidae might be typical for low-latitude deep-sea fish assemblages.

Rare fishes collected on the abyssal plain adjacent to Senghor Seamount: a Ipnops murrayi (16.6 cm TL); b Paracaristius sp. (20.3 cm TL); c Barathronus cf. parfaiti (10.5 cm TL); d Acanthonus armatus (31.6 cm TL); e Bassozetus levistomatus (68.0 cm TL); f Bassozetus taenia (26.0 cm TL); g Aldrovandia rostrata (59.0 cm TL)
Merrett (1992) associated the faunal change between the southerly and northern assemblages with the influence of upwelling and increased surface production along the continental margin of NW Africa. The less intense productivity at our bathyal station (18°N) south of the CVFZ than in the upwelling zone (20–22°N) to the north or within the CVFZ may be responsible for the observed differences in species composition between the deep-sea fish assemblages, due to differences in food supply and feeding preferences. However, the patterns involving distribution and connectivity among the different deep-sea fish assemblages of the eastern Atlantic seamount populations are still poorly known or are limited to commercially valuable species (White et al. 2009; Carlsson et al. 2011; Varela et al. 2013).
Since there is still little knowledge about the reproduction of the common deep-water species C. armatus, the collection of the nine adult C. armatus females with large ovaries containing ripening oocytes in our samples is remarkable. The only record of C. armatus with ripe or close to ripe eggs was reported by Stein (1985) for individuals larger than 74 cm TL. The female with the ripest eggs was 96 cm long. The author concluded that 1) only very large females become sexually mature, 2) reproduction may not be simultaneous within one population, because not all females with ripening eggs were at the same stage of ovarian development, and 3) the reproduction of the species may be semelparous. In our collection, C. armatus females with ripening eggs were smaller in size, ranging from 54.5 to 83.0 cm TL (corresponding to 610–3280 g TW), which may challenge the hypotheses made by Stein (1985). Our findings, rather, support the assumption that C. armatus females can mature at medium size (probably with simultaneous reproduction), making semelparous reproduction very unlikely.
Conclusions
In our study, the area around Senghor Seamount is shown to be a transition between northern and southern Atlantic fauna (Fig. 4), underscoring the role of the Cape Verde archipelago in a broader biogeographic context. The similarity between the demersal fauna of Senghor Seamount and that of subtropical NE Atlantic seamounts supports the role of seamounts as "stepping stones" for oceanic dispersal of benthopelagic fishes, particularly typical shelf and slope species, across large distances—for example, between continental margins and mid-ocean ridges. The relatively high number of new records for the area at the reference station, including rarely caught species, just indicates that abyssal and bathyal depths are undersampled in the tropical eastern Atlantic.

Locations of Senghor Seamount and the reference site (RS) in the eastern Atlantic with schematic surface circulation, NW African upwelling zones and fish assemblages. Representative fish species caught at Senghor Seamount and the abyssal plain are grouped according to their common distribution in the eastern Atlantic (see text for literature) and arranged according to depth of occurrence within one assemblage. AzC Azores Current, CC Canary Current, CVFZ Cape Verde Frontal Zone, MO Mediterranean Outflow, NEC North Equatorial Current, RCG Recirculation Gyre
References
Abecasis D, Cardigos F, Almada F, Gonçalves JMS (2009) New records on the ichthyofauna of the Gorringe seamount (Northeastern Atlantic). Mar. Biol Res 5:605–611. doi:10.1080/17451000902729696
Almeida AJ, Biscoito M, Santana JI, González JA (2010) New records of grey cutthroat, Synaphobranchus affinis (Actinopterygii: Anguilliformes: Synaphobranchidae) from the eastern-central Atlantic Ocean. Acta Ichthyol Piscat 40:66–70
Benchimol C, Francour P, Lesourd M (2009) The preservation of marine biodiversity in West Africa, the Case of Cape Verde Islands: proposal of a new biodiversity policy management. In: 1st Cape Verde Congress of Regional Development, Praia, Santiago Island, Cape Verde. APDR, pp 297—318
Brito A, Falcón JM, Herrera R (2006) Características geográficas de la ictiofauna litoral de las islas de Cabo Verde y comparación con los archipiélagos macaronésicos. Rev Acad Canaria Cienc 18:93–109
Carlsson J, Shephard S, Coughlan J, Trueman CN, Rogan E, Cross TF (2011) Fine-scale population structure in a deep-sea teleost (orange roughy, Hoplostethus atlanticus). Deep-Sea Res I 58:627–636. doi:10.1016/j.dsr.2011.03.009
Chivers AJ, Narayanaswamy BE, Lamont PA, Dale A, Turnewitsch R (2013) Changes in polychaete standing stock and diversity on the northern side of Senghor Seamount (NE Atlantic). Biogeosciences 10:3535–3546
Christiansen B, Martin B (2000) Deep-sea benthopelagic nekton observations at two stations in the northern Arabian Sea: links to organic matter supply? Deep-Sea Res II 47:3027–3038
Christiansen B, Martin B, Hirch S (2009) The benthopelagic fish fauna of Seine Seamount, NE Atlantic: composition, population structure and diets. Deep-Sea Res II 56:2705–2712. doi:10.1016/j.dsr2.2008.12.032
Christiansen B, Brand T, Büntzow M et al (2011) Structure and function of Seamount ecosystems in the Cape Verde Region, Northeast Atlantic – Cruise No. M79/3 – September 24 – October 23, 2009 – Las Palmas (Spain) – Mindelo (Cape Verde). METEOR-Berichte, M79/3. DFG-Senatskommission für Ozeanographie, Bremen. doi:10.2312/cr_m79_3
Christiansen B, Vieira RP, Christiansen S, Denda A, Oliveira F, Gonçalves JMS (2014) The fish fauna of Ampère Seamount (NE Atlantic) and the adjacent abyssal plain. Helgoland Mar Res 69:13–23. doi:10.1007/s10152-014-0413-4
Christiansen B, Türkay M, Emeis K-C (2015) Biology and biogeochemistry of the Eastern Mediterranean Sea – Cruise No. M71 – December 11, 2006 – February 4, 2007 – Heraklion (Greece) – Istanbul (Turkey). METEOR-Berichte, M71. DFG-Senatskommission für Ozeanographie, Bremen. doi:10.2312/cr_m71
Cohen DM, Inada T, Iwamoto T, Scialabba N (1990) FAO Species Catalogue. Vol. 10. Gadiform fishes of the world (Order Gadiformes). An annotated and illustrated catalogue of cods, hakes, grenadiers and other gadiform fishes known to date. FAO Fish Synop 125:442
Colloca F, Carpentieri P, Balestri E, Ardizzone G (2010) Food resource partitioning in a Mediterranean demersal fish assemblage: the effect of body size and niche width. Mar Biol 157:565–574
D’Oliveira Fonseca B (2000) Expansion of pelagic fishereies in Cape Verde. A feasibility study. The United Nations University - Fisheries Training Programme, Reykjavik
Denda A, Christiansen B (2014) Zooplankton distribution patterns at two seamounts in the subtropical and tropical NE Atlantic. Mar Ecol 35:159–179. doi:10.1111/maec.12065
Ehrich S (1977) Die Fischfauna der Großen Meteorbank. Meteor Forschungsergeb Reihe D 25:1–23
Franca MLP, Vasconcelos MS (1962) Peixes do Arquipélago de Cabo Verde. Notas Mimeog Centro Biol Piscat 28:1–86
Franco MAL, Braga AC, Nunan GWA, Costa PAS (2009) Fishes of the family Ipnopidae (Teleostei: Aulopiformes) collected on the Brazilian continental slope between 11° and 23° S. J Fish Biol 75:797–815. doi:10.1111/j.1095-8649.2009.02324.x
Freitas R (2014) The coastal ichthyofauna of the Cape Verde Islands: a summary and remarks on endemism. Zool Caboverdiana 5:1–13
Froese R, Pauly D (2016) FishBase. World Wide Web electronic publication. Version (03/2016)
González JA, Tariche O, Santana JI, García-Mederos AM, Tuset VM, Jimenez S, Biscoito M (2010) The family Moridae (Gadiformes) from the Cape Verde Islands (eastern central Atlantic Ocean), with first record of Physiculus cyanostrophus. Cybium 34:217–221
González JA, Martins A, Santana JI et al (2014) New and rare records of teleost fishes from the Cape Verde Islands (eastern-central Atlantic Ocean). Cybium 38:289–300
Gordon JDM, Duncan JAR (1987) Deep-Sea bottom-living fishes at two repeat stations at 2200 m and 2900 m in the Rockall Trough, northeastern Atlantic Ocean. Mar Biol 96:309–325
Gordon DM, Merrett NR, Bergstad OA, Swan SC (1996) A comparison of the deep-water demersal fish assemblages of the Rockall Trough and Porcupine Seabight, eastern North Atlantic: continental slope to rise. J Fish Biol 49 (Suppl. A):217—238
Haedrich RL, Merrett NR (1988) Summary atlas of deep-living demersal fishes in the North Atlantic Basin. J Nat Hist 22:1325–1362
Hanel R, John H-C (2015) A revised checklist of Cape Verde Islands sea fishes. J Appl Ichthyol 31:135–169. doi:10.1111/jai.12621
Hanel R, John H-C, Meyer-Klaeden O, Piatkowski U (2010) Larval fish abundance, composition and distribution at Senghor Seamount (Cape Verde Islands). J Plankton Res 32:1541–1556. doi:10.1093/plankt/fbq076
Henriques C, Priede IG, Bagley PM (2002) Baited camera observations of deep-sea demersal fishes of the northeast Atlantic Ocean at 15–28°N off West Africa. Mar Biol 141:307–314
Iglésias SP (2013) Actinopterygians from the North-eastern Atlantic and the Mediterranean (A natural classification based on collection specimens, with DNA barcodes and standardized photographs), Vol. I (plates), version 09, 01 April 2013
Kukuev EI, Parin NV, Trunov IA (2012) Materials for the revision of the family Caristiidae (Perciformes). 2. Manefishes from the East Atlantic (redescription of Platyberyx opalescens Zugmayer and description of two new species Platyberyx mauli sp. n. and Caristius andriashevi sp. n.). J Ichthyol 52:185–199
Lathuilière C, Echevin V, Lévy M (2008) Seasonal and intraseasonal surface chlorophyll-a variability along the northwest African coast. J Geophys Res 113:C05007. doi:10.1029/2007jc004433
Lázaro C, Fernandes MJ, Santos AMP, Oliveira P (2005) Seasonal and interannual variability of surface circulation in the Cape Verde region from 8 years of merged T/P and ERS-2 altimeter data. Remote Sens Environ 98:45–62. doi:10.1016/j.rse.2005.06.005
Lloris D, Rucabado J, Figueroa H (1991) Biogeography of the Macaronesian ichthyofauna (The Azores, Madeira, The Canary Islands, Cape Verde and the African enclave). Bol Mus Mun Funchal 43:191–241
MAAP/GEP (2004) Plano de Gestão dos Recursos da Pesca. MAAP/GEP, Praia
Magnússon J, Magnússon JVV (1985) Survey of demersal fish resources in the waters off Cape Verde Islands. II Report: the survey in May/June 1984. ICEIDA/Cape Verde Islands Fisheries Project. Icelandic International Development Agency, Reykjavík
Magnússon J, Magnússon JVV (1987) Survey of demersal fish resources in the waters off Cape Verde Islands. III Report: the survey in June/July 1985. ICEIDA/Cape Verde Islands Fisheries Project. Icelandic International Development Agency, Reykjavík
Marshall NB, Staiger JC (1975) Aspects of the structure, relationships and biology of the deep-sea fish Ipnops murrayi (Family Bathypteroidae). B Mar Sci 25:101–111
Martin B, Christiansen B (1997) Diets and standing stocks of benthopelagic fishes at two bathymetrically different midoceanic localities in the northeast Atlantic. Deep-Sea Res I 44:541–558
Mason E, Colas F, Molemaker J, Shchepetkin AF, Troupin C, McWilliams JC, Sangrà P (2011) Seasonal variability of the Canary Current: a numerical study. J Geophys Res 116. doi:10.1029/2010JC006665
Menezes GM, Tariche O, Pinho MR, Duarte PN, Fernandes A, Aboim MA (2004) Annotated list of fishes caught by the R/V Arquipélago off the Cape Verde archipelago. Arquipel Life Mar Sci 21A:57–71
Menezes GM, Rosa A, Melo O, Pinho MR (2009) Demersal fish assemblages off the Seine and Sedlo Seamounts (Northeast Atlantic). Deep-Sea Res II 56:2683–2704. doi:10.1016/j.dsr2.2008.12.028
Menezes GM, Rosa A, Melo O, Porteiro F (2012) Annotated list of demersal fishes occurring at Sedlo Seamount, Azores north-east central Atlantic Ocean. J Fish Biol 81:1003–1018. doi:10.1111/j.1095-8649.2012.03355.x
Merrett NR (1987) A zone of faunal change in assemblages of abyssal Demersal fish in the Eastern North Atlantic: a response to seasonality in production? Biol Oceanogr 5:137–151
Merrett NR (1992) Demersal ichthyofaunal distribution in the abyssal eastern North Atlantic, with special reference to Coryphaenoides (Nematonurus) armatus (Macrouridae). J Mar Biol Assoc UK 72:5–24
Merrett NR, Fasham MJR (1998) Demersal ichthyofaunal distribution in the abyssal North Atlantic revisited: the effect of sample size on ordination. Mar Ecol Prog Ser 173:267–274
Merrett NR, Marshall NB (1981) Observations on the ecology of deep-sea bottom-living fishes collected off northwest Africa (08°–27°N). Prog Oceanogr 9:185–244
Merrett NR, Haedrich RL, Gordon JDM, Stehmann M (1991) Deep demersal fish assemblage structure in the Porcupine Seabight (eastern North Atlantic): results of single warp trawling at lower slope to abyssal soundings. J Mar Biol Assoc UK 71:359–373
Mittelstaedt E (1991) The ocean boundary along the northwest African coast: circulation and oceanographic properties at the sea surface. Prog Oceanogr 26:307–355
Monteiro P, Ribeiro D, Silva JA, Bispo J, Gonçalves JMS (2008) Ichthyofauna assemblages from two unexplored Atlantic seamounts: Northwest Bank and João Valente Bank (Cape Verde archipelago). Sci Mar 72:133–143
Morales-Nin B, Maynou F, Sardà et al (2003) Size influence in zonation patterns in fishes and crustaceans from deep-water communities of the western Mediterranean. J Northw Atl Fish Sci 31:413–430
Nakamura I, Parin NV (1993) FAO Species Catalogue. Vol. 15. Snake mackerels and cutlassfishes of the world (families Gempylidae and Trichiuridae). An annotated and illustrated catalogue of the snake mackerels, snoeks, escolars, gemfishes, sackfishes, domine, oilfish, cutlassfishes, scabbardfishes, hairtails, and frostfishes known to date. FAO Fish Synop 125:136
Nielsen JG (1966) Synopsis of the Ipnopidae (Pisces, Iniomi) with descriptions of two new abyssal species. In: Wolff T (ed) Galathea 2 Report, vol 08. The Galathea Committee, Copenhagen, pp 49–76
Nielsen JG (1969) Systematics and biology of the Aphyonidae (Pisces, Ophidioidea). In: Wolff T (ed) Galathea 2 Report, vol 10. The Galathea Committee, Copenhagen, pp 7–90
Nielsen JG (1990) Ophidiidae. In: Quero JC, Hureau JC, Karrer C, Post A, Saldanha L (eds) Check-list of the fishes of the eastern tropical Atlantic (CLOFETA), vol 2. JNICT, Lisbon, pp 564–573
Nielsen JG, Merrett NR (2000) Revision of the cosmopolitan deep-sea genus Bassozetus (Pisces : Ophidiidae) with two new species. In: Wolff T (ed) Galathea 2 Report, vol 18. The Galathea Committee, Copenhagen, pp 7–56
Nielsen JG, Cohen DM, Markle DF, Robins CR (1999) FAO Species Catalogue. Vol. 18. Ophidiiform fishes of the world (Order Ophidiiformes). An annotated and illustrated catalogue of pearlfishes, cusk-eels, brotulas and other ophidiiform fishes known to date. FAO Fish Synop 125:178
Pakhorukov NP (2008) Visual observations of fish from seamounts of the Southern Azores region (the Atlantic Ocean). J Ichthyol 48:114–123. doi:10.1134/S0032945208010104
Paxton JR (1999) Berycidae. Alfonsinos. In: Carpenter KE, Niem VH (eds) FAO species identification guide for fishery purposes. The living marine resources of the WCP. Vol. 4. Bony fishes part 2 (Mugilidae to Carangidae). FAO, Rome, pp 2218–2220
Pérez-Rodríguez P, Pelegrí JL, Marrero-Díaz A (2001) Dynamical characteristics of the Cape Verde frontal zone. Sci Mar 65:241–250
Pierre C, Vangriesheim A, Laube-Lenfant E (1994) Variability of water masses and of organic production regeneration systems as related to eutrophic, mesotrophic and oligotrophic conditions in the northeast Atlantic ocean. J Mar Syst 5:159–170
Reiner F (1996) Catálogo dos peixes do arquipélago de Cabo Verde. Publicações Avulsas do IPIMAR, Lisboa
Reiner F (2005) Peixes do Arquipélago de Cabo Verde. Instituto Nacional de Desenvolvimento das Pescas, Mindelo
Santos RS, Porteiro FM, Barreiros JP (1997) Marine fishes of the azores. Annotated checklist and bibliography. A catalogue of the Azorean marine ichthyodiversity. Arquipel Life Mar Sci Suppl 1:1–242
Shank TM (2010) Seamounts: deep-ocean laboratories of faunal connectivity, evolution, and endemism. Oceanography 23:108–122
Shcherbachev YN, Kukuev EI, Shlibanov VI (1985) Composition of the benthic and demersal ichthyocenoses of the submarine mountains in the southern part of the North Atlantic range. J Ichthyol 25:110–125
Stein DL (1985) Towing large nets by single warp at abyssal depths: methods and biological results. Deep-Sea Res 32:183–200
Stevenson DE, Kenaley CP (2011) Revision of the manefish genus Paracaristius (Teleostei: Percomorpha: Caristiidae), with descriptions of a new genus and three new species. Copeia 2011:385–399. doi:10.1643/ci-10-129
Stocks KI, Clark MR, Rowden AA, Consalvey M, Schlacher TA (2012) CenSeam, an international program on seamounts within the Census of Marine Life: achievements and lessons learned. PLoS One 7:e32031. doi:10.1371/journal.pone.0032031
Strømme T, Sundby S, Sætersdal G (1982) A survey of the fish resources in the coastal waters of the Republic of Cape Verde, November 1981. Institute of Marine Research, Bergen
Sulak KJ (1977) Aldrovandia oleosa, a new species of the Halosauridae, with observations on several other species of the family. Copeia 1977:11–20
Thorsteisson V, Montero VMS, Almada EO (1995) Ground fish survey in the waters off Cape Verde 1994. Icelandic International Development Agency, Reykjavík
Tomczak MJ (1981) An analysis of mixing in the frontal zone of South and North Atlantic Central Water off North-West Africa. Prog Oceanogr 10:173–192
Trunov IA, Kukuev EI, Parin NV (2006) Materials for the revision of the family Caristiidae (Perciformes): 1. Description of Paracaristius heemstrai gen. et sp. nov. J Ichthyol 46:441–446
Vakily JM, Camara SB, Mendy AN et al (2002) Poissons marins de la sous-région nord-ouest africaine. EUR 20379 FR. Commission Européenne, Bruxelles
Varela AI, Ritchie PA, Smith PJ (2013) Global genetic population structure in the commercially exploited deep-sea teleost orange roughy (Hoplostethus atlanticus) based on microsatellite DNA analyses. Fish Res 140:83–90
Vieira RP, Thiel R, Christiansen B, Coelho R, Denda A, Gonçalves J (2013) The deep-sea fish Kali macrodon (Norman, 1929), a new record for the tropical eastern Atlantic off Cape Verde. Mar Biodivers Rec 6:1–3. doi:10.1017/S1755267212001248
Wessel P, Sandwell DT, Kim S-S (2010) The global seamount census. Oceanography 23:24–33
White TA, Stefanni S, Stamford J, Hoelzel AR (2009) Unexpected panmixia in a long-lived, deep-sea fish with well-defined spawning habitat and relatively low fecundity. Mol Ecol 18:2563–2573
Whitehead PJP, Bauchot M-L, Hureau J-C, Nielsen J, Tortonese E (eds) (1984–1986) Fishes of the North-eastern Atlantic and the Mediterranean. Vols. 1–3. UNESCO Press, Paris
Wirtz P et al (2013) The coastal fishes of the Cape Verde Islands – new records and an annotated check-list (Pisces). Spixiana 36:113–142
Zenk W, Klein B, Schröder M (1991) Cape Verde frontal zone. Deep-Sea Res 38:S505–S530
Acknowledgments
The authors would like to thank the crew of the RV Meteor and RV Poseidon for their professional support on the cruises, and Ralf Thiel (Zoological Museum, Universität Hamburg) for his valuable advice. An anonymous reviewer made important comments, which helped to improve the manuscript considerably.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Financial support
Rui Pedro Vieira received financial support from CCMAR and funds from DAAD - Deutscher Akademischer Austauschdienst (German Academic Exchange Service) for a short-term scholarship at the IHF – Universität Hamburg (Germany). Rui Pedro Vieira is currently supported by a Portuguese doctoral grant (FCT/SFRH/BD/84030/2012) and partially by European Funds through COMPETE and by National Funds through the Portuguese Science Foundation (FCT) within project PEst-C/MAR/LA0017/2013. Rui Coelho is supported by an Investigador-FCT contract from the Portuguese Science Foundation (FCT, Fundação para a Ciência e Tecnologia) supported by the EU European Social Fund and the Programa Operacional Potencial Humano (Ref: IF/00253/2014). The Deutsche Forschungsgemeinschaft (DFG) provided the ship time and financial support for the Meteor cruise M79/3, and the Universität Hamburg for the Poseidon cruise P423.
Additional information
Communicated by R. Thiel
Rights and permissions
About this article

Cite this article
Vieira, R.P., Coelho, R., Denda, A. et al. Deep-sea fishes from Senghor Seamount and the adjacent abyssal plain (Eastern Central Atlantic). Mar Biodiv 48, 963–975 (2018). https://doi.org/10.1007/s12526-016-0548-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12526-016-0548-4