
Abstract
The distinctive characteristic of the Monte Desert, the most arid rangeland of Argentina, is reflected in the varied composition of its small mammal fauna. The present study analyses the taphonomic features and the palaeoenvironmental implications of small mammal bone and tooth accumulations recovered in open-air archaeological sites from the Monte Desert plains of northern Mendoza (Central Western Argentina) in the South American arid diagonal. In order to identify the agents responsible for such accumulations relative abundance trends, bone breakage patterns and digestive corrosion were evaluated. The action of postdepositional processes, like weathering and trampling, also were analysed. Taphonomic results allowed the detection of avian raptors—owls in particular—as the principal accumulating agents of small mammal bone and tooth accumulations of open-air archaeological sites from the Monte Desert. In order to make palaeoenvironmental inferences, the taxonomic structure of archaeological and modern small mammal assemblages was analysed and compared, added to the evaluation of taphonomic dynamics in the archaeological sites. The taxonomic composition of archaeological assemblages differed from that of the current assemblages from the study area. Thus, an extinct species (Holochilus lagigliai) and a little-known species highly adapted to desert environments (Tympanoctomys barrerae) were detected in the archaeological record. Recent pellet samples showed major proportions of opportunistic and generalist species as well as the presence of an exotic species (Mus musculus). The general impoverishment of current small mammal assemblages in comparison with past ones (e.g. richness, diversity and evenness) could be attributed to the deep impact of the anthropic perturbations generated by economic activities since the arrival of European conquerors ca. 500 years BP. Small mammal archaeological assemblages could indicate a scenario of greater environmental heterogeneity during the Late Holocene than in the present, and thus, greater availability of patch resources in the landscape.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Small mammal (≤ 1 kg) bone and tooth remains recovered from archaeological sites have been overlooked in South American archaeological studies for various reasons. Among them are the difficulties associated with taxonomic determinations, deficiencies present in sample recovery and preservation in fieldwork and the lack of a coherent theoretical-methodological model applied to the study of such zooarchaeological remains (Stahl 1996; Pardiñas 1999; Fernández et al. 2017a). Small mammals, in general, have narrow ecological requirements, and the remains recovered in archaeological contexts offer the possibility of performing ecological inferences over past environments and their evolution over time, taking into account the current distribution of each taxon (Andrews 1995; Fernández-Jalvo et al. 2011).
In South America, mainly in Argentina, most research on archaeological small mammal bone assemblages has focused on the Pampean, Patagonian and Northwestern regions, mainly corresponding to Pampa, Patagonian steppe and Puna phytogeographic provinces. Most of them showed the potential of such remains to improve our comprehension of human-environmental systems in the past (e.g. Pardiñas 1999; Fernández et al. 2009, 2011a, 2015a, b; Fernández 2012; Ortíz et al. 2012; Scheifler et al. 2012; Teta et al. 2013; Quintana 2016). Palaeoenvironmental studies on historical contexts from the Monte Desert based on documentary evidence point out major environmental perturbations during the last 500 years. Such alterations are related with the economic activities developed since the arrival of European conquerors, such as mining, land clearing, intensive livestock and agriculture, felling and manipulation of watercourses, among others (e.g. Abraham and Prieto 1981; Prieto et al. 2003, 2008, 2012; Prieto and Chiavazza 2006; Prieto and Rojas 2012). At the same time, some regional ecological studies have pointed out the deep impact that recent economic and commercial activities have had on animal and plant communities, also strongly modifying the landscape (Ojeda and Mares 1982; Roig 1991; Tabeni and Ojeda 2003, 2005). However, there are still few studies that evaluate environmental fluctuations during Prehistoric times, particularly the Late Holocene, probably associated with the establishment of current climatic conditions in the region around 4000 years ago (Navarro et al. 2012 and references therein). The scarcity of environmental information for the Late Holocene in Monte Desert contexts precludes a systematic approach to human-environment relationships during Pre-Hispanic times.
Some studies conducted in Central Western Argentina in the phytogeographic units of Puna, Monte and Patagonia have utilised archaeological small mammal assemblages not only to make palaeoenvironmental inferences but also to interpret human behaviour in the past (e.g. Fernández et al. 2009; Fernández 2012; López et al. 2016; Fernández and Pardiñas 2018). However, these works have generally been conducted in rockshelters or caves sites, possibly due to the sedimentary composition of the hundreds of sites located in the plains of the Monte Desert, mainly on sandy and unconsolidated soils, often without defined stratigraphies in aeolian environments (e.g. Chiavazza 2009, 2012, 2014). Despite these characteristics, archaeological studies from the central Monte Desert sites demonstrated the potential of these sites to understand both human occupations in the past and the environmental fluctuations of the last two millennia when human occupation of the central Monte Desert plains was recorded (e.g. Chiavazza 2007, 2012, 2014, 2016; García Llorca and Cahiza 2007; Cahiza and Ots 2014; Ots et al. 2016). A recent study argued that regional archaeological preferences by researchers towards cave or rockshelter sites and stratified contexts shows an incomplete picture of prehistoric occupation and landscape use in Central Western Argentina. Thus, that study highlights the importance of sites located in the plains of the Monte Desert—which covers more than 50% of the territory of Central Western Argentina—has been underestimated due to aeolian action and fragmentation of the archaeological record (Garvey and Bettinger 2018).
The Monte phytogeographic province, the most arid rangeland of Argentina, is located in western Argentina (24° 35′, 44° 20′ S; 62° 54′, 69° 5′ W). It is an arid-semiarid zone, where the ratio between annual rainfall and evapotranspiration is lower than 0.5 (between 0.05 and 0.5) (Abraham et al. 2009; Labraga and Villalba 2009). Physiognomically, the central Monte Desert is a mosaic of two vegetation types: shrubby steppes dominated by Larrea cuneifolia, Larrea divaricata, Larrea nítida, Acantholippia spp. and Junellia spp. and open mesquite forests of Prosopis—mainly between 29° and 33° S—in addition to large proportions of bare sandy soils (Rundel et al. 2007). The distinctive feature of the Monte is reflected in the varied composition of its mammal fauna, particularly small mammals including rodents and marsupials (Ojeda et al. 2002). Therefore, taphonomic and palaeoenvironmental studies based on small mammal bone accumulations recovered in archaeological sites from the central Monte Desert constitute a valuable tool to interpret human-environment relationships and past landscape features.
The present study analyses the taphonomic features of small mammal bone and tooth accumulations recovered in six open-air archaeological sites from the central Monte Desert (Central Western Argentina) in an area crossed by the South American arid diagonal (Bruniard 1982). In order to identify possible accumulating agents of such bone sets, relative abundances, breakage patterns, digestive corrosion, thermal alterations and cut marks were evaluated, as well as signals produced by postdepositional processes (e.g. weathering and trampling). Taphonomic results were interpreted and compared with those reported in regional archaeological research performed at Monte Desert sites and with information provided by actualistic studies of different predators that inhabit the study area. With the aim of making palaeoenvironmental inferences, the taxonomic compositions of archaeological assemblages and current regional ones were analysed and compared.
Materials and methods
The study was carried out in a large area of northeastern Mendoza Province, Argentina, (Figs. 1 and 2) including different areas of the Monte phytogeographic province in open-air archaeological sites from aeolian sedimentary contexts between 900 and 500 masl.

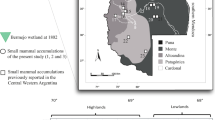
Archaeological and recent small mammal bone accumulations recovered in the Monte Desert in a phytogeographic context. Open-air archaeological sites: 1, Agua de los Pajaritos; 2, Punto Arqueológico 14.7 (PA14.7); 3, Punto Arqueológico 52 (PA52); 4, Punto Arqueológico 24.2 (PA24.2); 5, Punto Arqueológico 46.4 (PA46.4); 6, Punto Arqueológico 68.1 (PA68.1). Recent accumulations from owl pellets: 7, Athene cunicularia from Lunlunta (Maipú); 8, Tyto alba from San José (Lavalle); 9, Tyto alba from Paleocauce central (PA46.4)

Some representative environments from the Monte Desert of northern Mendoza and some open-air archaeological sites of the present study. a, b Punto Arqueológico 68 archaeological site in a scrubland (Larrea spp. mainly) with some proportions of bare sandy soils. c Mendoza river in the lowlands; d Agua de los Pajaritos archaeological site in the piedmont
The small mammal bone remains analysed in this study were recovered in archaeological excavations conducted between 1998 and 2014. Open-air archaeological samples were generally obtained from sandy contexts associated with different kinds of watercourses. Thus, three of them were recovered in paleoriverbeds corresponding to Mendoza river (PA24.2, PA46.4, PA52); one sample belongs to current lagoon contexts (Lagunas de Guanacache; PA14.7); one sample was obtained from the current riverbed of Desaguadero river (PA68.1); whereas, the last sample comes from the lower piedmont of the Andean precordillera (Agua de los Pajaritos). As determined through radiocarbon and relative dates, all sites correspond to human occupation contexts during the last 2000 years (Table 1). The general features of each archaeological site can be seen in the Electronic supplementary material (S1).
Considering the general lack of stratified contexts on sandy soils in these archaeological sites, the material was obtained through surface and subsurface (0–5 cm) excavations. However, an open-air stratified site from the piedmont (Agua de los Pajaritos) was also included in the present study. Although PA68.1 is a stratified site, all small mammal bone remains were recovered in surface and subsurface sectors (0–5 cm). A 2-mm screen mesh was used for the recovery of the remains.
Bone and tooth remains were examined using a stereomicroscope (variable magnification of × 8 to × 50). Taxonomic determinations were made at species level, whenever possible, based on cranial and dental elements, through comparison with current specimens from the Vertebrate Collection of IADIZA (CCT-Mendoza) as well as with specialised literature (e.g. De Santis et al. 1991; Fernández et al. 2011b). Some species are indistinguishable from cranial and dental remains, which in certain cases the determinations were made to genus level (e.g. Abrocoma sp., Ctenomys sp., Eligmodontia sp.).
The number of identified specimens by taxon (NISP), minimal number of individuals (MNI) and minimal number of elements (MNE) was calculated (Lyman 2008). Shannon diversity index (H′) was calculated on the basis of MNI, taking into account the logarithm of richness as the highest value (greatest diversity) that each sample could reach. An evenness index also was calculated based on H′ (J = H′/Hmax), where Hmax is the logarithm of specific richness. Values vary from 0 to 1. The closer to number 1, the more heterogeneous and/or equally the species are represented, respectively (Krebs 1989).
To increase the information on modern small mammal assemblages from the study region, and thus, strengthen the potential of our palaeoenvironmental inferences, owl pellet samples were recovered both near archaeological sites (e.g. PA46.4), whenever possible, and in other meaningful areas associated with intensive wine and olive agriculture (Lunlunta). Only cranial and dental elements were analysed from the current samples with the aim to assess the taxonomic composition of these communities. Data previously reported on modern small mammal assemblages were also considered (e.g. Rodríguez and Ojeda 2013; Castillo et al. 2018; López et al. 2018).
The taphonomic analysis followed the methodology proposed by Andrews (1990), Fernández-Jalvo and Andrews (1992) and Fernández et al. (2017b). This methodology also examines the possible natural predators of small mammals from the study area. Modification categories consider alteration, frequency and intensity degrees of affected elements based on three main variables: relative abundances, breakage and digestion. Relative abundances of skeletal elements were evaluated taking into account the representation of each element in the whole sample (MNEi) on the basis of the expected number of each element in one individual (Ei) and on the basis of MNI by using the following formula: MNEi / (Ei × MNI) × 100. To evaluate the relationships between cranial and postcranial elements, two indexes were calculated: postcrania/crania (pc/c) = ((femora + tibiae + humeri + radii + ulnae) × (8) / (mandibles + maxillae + molars) × (5)) and f + h/md + mx = ((femora + humeri / mandibles + maxillae)). To evaluate the relationship between the representation of distal and proximal elements of the skeleton, one index was used: t + u/f + h = ((tibiae + ulnae / femora + humeri)), whereas two calculations were used for measuring tooth loss: alv/m = ((empty mandibular molar alveoli + empty maxillary molar alveoli/molars)) and alv/i = ((empty mandibular incisor alveoli + empty premaxillary incisor alveoli/incisors)). Didelphid mammals were not included in the last two calculations, as their dental formula was considerably different from that of rodents.
Breakage patterns on cranial (skull, mandible, teeth) and postcranial bones (femora, humeri, tibiae, ulnae) were analysed taking into account the degree of completeness of each element, considering complete and fractured elements separately and calculating the percentage of breakage. Four categories were used for skull breakage: complete, maxillary with zygomatic, maxillary without zygomatic and minor fragments; four degrees were used for mandible breakage: complete, ascendant ramus broken, without ascendant ramus and without ascendant ramus and inferior edge broken. Breakage of diagnostic long bones (femora, humeri, tibiae and ulnae) was recorded as complete, shaft, proximal and distal parts.
Modifications by digestion were evaluated on diagnostic elements (molars, incisors, femora and humeri) distinguishing four degrees of digestive corrosion (light, moderate, heavy and extreme), except when digestion was absent. Digestion is the most reliable taphonomic variable to identify the possible accumulating agent of small mammal bone remains from archaeological and palaeontological sites, owing to its clear taphonomic signals, which can be differentiated from other taphonomic alterations, and whose traces are not modified by postdepositional processes (Andrews 1990; Fernández-Jalvo et al. 2014; Fernández et al. 2017b).
In order to strengthen the interpretation of possible accumulating agents, size and predictability indexes were calculated (Pardiñas 1999). The first one allows distinguishing small mammal bone accumulations generated by nocturnal raptors (mainly strigiforms) from those accumulated by other predators, and is calculated using the following formula: (MNI small sized -≤ 50 g- + MNI medium sized –50 to 150 g-) / (MNI large sized -≥ 150 g-). The index considers that strigiforms feed on smaller species than accipitriforms, carnivorous mammals and humans. The second is calculated to evaluate relationships between grouped and non-grouped species of small mammals, using the following formula: (MNI colonials + MNI gregarious + MNI grouped) / (MNI cryptic + MNI solitaries). Nocturnal raptors generally prey on cryptic and solitaries small mammal species, whereas the proportions of grouped species in assemblages generated by accipitriforms and humans are higher than that observed for strigiforms.
The action of postdepositional processes was also analysed, including weathering, trampling, root etching and abrasion (Andrews 1990; Fernández-Jalvo et al. 2014). Principal component analysis (PCA) was conducted against previously reported archaeological and modern samples (López et al. 2016; López 2018; Andrews 1990; Gómez 2005; Montalvo and Tejerina 2009; Montalvo et al. 2016; López et al. 2017a, b, 2018, respectively) in order to explore sample ordination in a multivariate space. PCA worked over a data matrix composed by percentage of digestion on incisors, molars and postcranial elements; percentage of breakage on cranial and postcranial elements; calculated indexes of relative abundances; and average of relative abundance of skeletal elements. PCA was performed using the software Paleontological Statistics (PAST) version 3.12 (Hammer et al. 2001).
Results
Taphonomic analysis
A total of 734 specimens was recovered and analysed. With the exception of isolated incisors, tibiae, calcanea and astragali in one site (PA46.4) and femora, astragalus and calcaneus in another site (PA24.2), the relative abundance of each skeletal element was low (Fig. 3). Thus, except in the aforementioned cases, the relative abundance of each skeletal element did not exceed 50%. Most elements had relative abundances of less than 20%. Relative abundance averages ranged between 6.05% (Agua de los Pajaritos) and 27.15% (PA46.4).

Top, relative abundances of skeletal elements of small mammal bone accumulations recovered in open-air archaeological sites from the Monte Desert. Down, relative abundance of skeletal elements in modern small mammal bone accumulations: Strix chacoensis (López et al. 2018), average Tyto alba, average of Strigiformes (after Andrews 1990); Average Falconiformes-Accipitriformes (after Andrews 1990); Athene cunicularia (Montalvo and Tejerina 2009); small felids (López et al. 2017a)
The results of the indices used to evaluate relationships between cranial and postcranial elements (pc/c and f + h/md + mx) revealed three accumulations that indicated a better representation of postcranial elements (PA14.7, PA24.2, PA52), whereas in all others, cranial bones were better represented than postcranial bones (Tables 1 and 2). In the three sites with higher postcranial representation, proximal elements were better represented than distal ones, whereas distal elements were better represented in the remaining sites. The relationship between the number of preserved alveolar spaces and incisors showed a loss of maxillae and mandibles in all subsurface sites, whereas in the stratified site, a loss of molars and incisors was detected. Regarding the alv md + alv mx / m index, in four sites tooth loss was observed (PA14.7, PA24.2, PA68.1 and Agua de los Pajaritos), whereas in PA46.4 and PA52, a loss of mandibles and maxillae was identified (Table 1).
Breakage of cranial and postcranial bones was considerably high (Tables 1 and 3). In the subsurface open-air sites, all cranial bones were recovered broken, whereas in four out of five sites, all postcranial bones were fractured. Although in the stratified site, breakage was lower than in the subsurface sites, the proportion of broken bones continued to be high. The great majority of broken bones (≥ 90%) showed rough and/or angled broken edges.
Digestive corrosion (almost always light and exceptionally moderate in PA14.7, PA24.2, PA46.4 and Agua de los Pajaritos—≤ 5% by site or heavy in Agua de los Pajaritos—≤ 5%) was detected in the diagnostic elements in all sites, although in different proportions (Tables 1 and 4; Fig. 4). Regarding incisors, proportions of digestion ranged from 0% in Agua de los Pajaritos and PA68.1 to 50% in PA24.2 (n = 1). The proportions of molars modified by digestion ranged from 0% in PA24.2 to 34% in PA46.4 (n = 18). Postcranial elements were affected by digestion from 0% in PA68.1 to 80% in Agua de los Pajaritos (n = 4).

Examples of weathering and digestion in open-air archaeological sites from the Monte Desert of Mendoza (a–f). Holochilus lagigliai (g–h). a Humerus with category 1 of weathering and breakage with rough and angled edges, b slight cracking and light digestion on femur, c phalanx with two modification category of weathering, d weathering (category 2) on femur, e light weathering (category 1) on humerus shaft, f weathering (category 1) and digestion (light) on Caviidae molar, g left hemimandible of H. lagigliai recovered in Punto Arqueológico 14.7 (Lagunas de Guanacache, Mendoza) in labial view, h left lower molar series of such hemimandible of H. lagigliai in occlusal view, with m1 and m2
A high proportion of elements, primarily identified as category 1, were weathered. In most sites, more than 75% of the elements were affected by this postdepositional process, except PA14.7 with 45% of weathered elements. No significant amounts of skeletal elements were detected with thermal alterations (≤ 11% with the exception of PA14.7–20%—in each set, Table 5). Generally, these elements were calcined or burned completely (in reference to the amount of surface area of the bone modified) not showing thermal alterations associated with anthropic activity, such as partial, differential or heterogeneously burned bones (e.g. Medina et al. 2012; Fernández et al. 2017a). Cut marks were not detected. Root traces were detected at very low frequencies (≤ 2.4% in each set), and abrasion from fluvial transport was not observed.
Taxonomic composition of archaeological small mammal bone assemblages
Small mammal assemblages recovered in open-air archaeological sites of the present study are mainly composed by typical species from the Monte Desert (Table 6). Large-sized small mammals (≥ 150 g) dominated in all sites. The detected species were the didelphid Thylamys pallidior (pallid fat-tailed opposum) (PA14.7 and Agua de los Pajaritos) and the caviomorph rodents Abrocoma sp. (chinchilla rat) (Agua de los Pajaritos), Galea leucoblephara (lowland yellow-toothed cavy) (PA24.2, PA46.4, PA68.1 and Agua de los Pajaritos), Microcavia australis (southern mountain cavy) (PA14.7, 24.2, PA46.4, PA68.1 and Agua de los Pajaritos), Ctenomys sp. (tuco-tuco) (PA14.7, PA46.4, PA52, PA68.1 and Agua de los Pajaritos) and Tympanoctomys barrerae (viscacha rat) (PA14.7, PA46.4, PA52); and the cricetid rodents Eligmodontia sp. (gerbil mouse) (PA24.2, PA46.4), Graomys griseoflavus (gray leaf-eared mouse) (PA24.2) and Holochilus lagigliai (marsh rat) (PA14.7) (Fig. 3).
Shannon diversity index (H′) showed values between 0.50 and 0.70 in all subsurface open-air sites, whereas in the stratified site, H′ reached 0.54. The evenness index was high in all subsurface open-air sites (≥ 0.89), whereas in the stratified site the value was moderate, close to 0.5. The predictability index showed values higher than 1 in all sites, whereas the size index resulted in values lower than 1, except in PA52. Frequency and index values can be observed in Table 6.
Taxonomic composition of modern small mammal bone assemblages
Small mammal assemblages recovered from modern pellet samples are composed by the didelphid T. pallidior (San José), the caviomorph rodents M. australis (Paleocauce central) and Ctenomys sp. (San José and Paleocauce central); the cricetid rodents Akodon dolores (dolorous grass mouse) (Paleocauce central), Calomys musculinus (drylands vesper mouse) (San José, Paleocauce central and Lunlunta), Eligmodontia sp. (San José and Paleocauce central), G. griseoflavus (San José and Paleocauce central); and the exotic muridae Mus musculus (house mouse) (Paleocauce central and Lunlunta). Species frequency and index values at each site can be observed in Table 7.
The H′ index resulted in values between 0.18 (Lunlunta) and 0.74 (Paleocauce central), whereas the evenness index indicated values between 0.61 (Lunlunta) and 0.88 (Paleocauce central). The predictability index was considerably lower than 1 in all current pellet samples, whereas the size index was higher than 1 in all samples.
Discussion
Accumulating agents and taphonomic issues
The results obtained from the taphonomic analysis of small mammal bone and tooth accumulations from open-air archaeological sites in the Monte Desert suggest the action of avian raptors, particularly owls. The absence of anthropic activity signals (e.g. kind and frequency of thermal alterations or cut marks) discards the participation of human beings in the generation of the aforementioned small mammal bone and tooth accumulations. The high level of breakage and the low relative abundance of skeletal elements (even total averages) suggest a considerable loss of bones.
Digestion is the most reliable variable to infer agents responsible for small mammal bone accumulations recovered in archaeological and palaeontological sites. This is due to the fact that digestive damage has a unique morphology and therefore can be clearly be distinguished from any other modification on bone and tooth surfaces (Andrews 1990; Fernández-Jalvo et al. 2014). On the contrary, other taphonomic variables (e.g. breakage, relative abundances and proportions of a kind of species) can be influenced both by predator behaviours and by postdepositional processes. This accounts for the difficulty in differentiating both processes on bone surface, thus generating equifinality problems (Andrews 1990; Pardiñas 1999).
Digestive corrosion, mainly light, was detected in all small mammal bone accumulations analysed in the present study. Although in some cases proportions of this effect were high, this could be the consequence of small sample size or of the high loss of skeletal elements. These features, added to the action of weathering and trampling on sandy soils, could bias the representation of each sample. Thus, high proportions of all types of diagnostic elements in only one assemblage were never detected. For example, where a high frequency of incisors digested was found (PA24.2), neither molars nor postcranial elements were strongly modified by digestion. These “contradictory” values (even in the stratified site) could also be influenced both by site characteristics (subsurface and/or open-air sites) and by the effect of postdepositional processes, like trampling and weathering. These processes are strong modifiers of “original” bone accumulations, altering site integrity and resolution as well as presence, abundance, completeness and proportion of skeletal elements (Andrews 1990). For example, calculated indexes to evaluate relationships between skeletal parts, however, did not show incoherencies inside each archaeological sample. On the other hand, such contradictory values could also be the effect of mixing accumulations from different predator groups or the selective transport of particular elements (e.g. digested incisors) in the sites (open-air sites in particular). However, the presence of only one type of digestion (almost always light) in all archaeological sites, the considerable loss of bones, the high level of breakage by trampling, the intense action of weathering, the absence of signals of fluvial transport, added to the sandy and surface features of the studied open-air sites, allow thinking about the strong influence of postdepositional processes in the generation of these confusing and scarcely coherent values. Thus, the material was accumulated by owls and later trampled and moved postdepositionally (through wind and weathering) occurring bone loss. Small molars may also have been lost as they move through screens easier than longer incisors. At this point, the role of digestion is reinforced as a proxy to approach the possible accumulating agents of small mammal bone and tooth accumulations recovered in open-air archaeological sites from the Monte Desert. Without stratigraphy, we can be sure these are not intrusive species burrowing taking into account the lack of complete articulated specimens, the discouloring of the bones, which does not look modern, and the type and frequencies of modifications by digestion.
Trampling has been pointed out as one of the main causes of bone loss (cranial bones and teeth mainly) (Andrews 1990) observed in the analysed archaeological samples. The great abundance of large species could be the consequence of these destructive processes, and it should not be attributed to the dietary preferences of certain predators. Thus, differential preservation has been found to be strongly linked with structural bone density in vertebrates of all sizes (e.g. Denys et al. 1997; Andrews and Armour-Chelu 1998). Atmospheric processes, like weathering, can considerably affect bone accumulations. For this reason, small mammal bones corresponding to larger species could be better preserved than bones from smaller species (Denys et al. 1997), although also small species could preserve better due to faster sediment cover speeds. The high abundance of autopodium bones (more dense than other skeletal elements) can also reflect differential preservation and destruction by postdepositional processes. Even small mammal bone accumulation from the stratified site showed high proportions of weathered bones and breakage by trampling.
Small mammal bone and tooth accumulations recovered in open-air archaeological sites from the Monte Desert showed some differences with those accumulations reported for rockshelter sites from the same study area located in highlands (refer to Table 1 for examples). Thus, loss of skeletal elements, breakage and weathering were higher in open-air sites than in rockshelter sites. In general, small mammal bone accumulations recovered in rockshelter sites, including regional ones, show better preservation conditions than those small mammal bone accumulations recovered in open-air sites (Andrews 1990; Fernández 2012; López 2018).
When taphonomic values were incorporated in a data matrix to explore ordination in a multivariate space through PCA, the distribution of open-air archaeological samples was much more similar among them than with any of the current predator samples from the Monte Desert (Fig. 5 top; components 1 and 2 account for 82.89% of the variation; eigenvalues PC1 = 3460.32, PC2 = 1923.46). This is interesting, considering that archaeological bone samples were “modern” bone samples in the past. The grouping of open-air archaeological samples and the discrimination of current small mammal bone samples generated by several predators reflects that the common features detected in small mammal bone samples from open-air archaeological sites are even stronger than the taphonomic trends generated by each predator. These common features are probably related with the sedimentary characteristics of open-air sites—all of them on sandy soils and almost always from surface and subsurface record—with the strong action of weathering and trampling, and with the participation of the same type of predators in all small mammal bone and tooth accumulations.

Principal component analysis based on taphonomic variables obtained from small mammal bone assemblages from de Monte Desert. Top, comparison between taphonomic values of open-air samples of the present study and taphonomic values reported for bone accumulations generated by several predators. Down, comparison between taphonomic values of open-air samples of the present study; taphonomic values reported for bone accumulations recovered in rockshelter sites from the study area (López et al. 2016; López 2018); and taphonomic values reported for bone accumulations generated by several predators Small felids (López et al. 2017a); Geranoaetus melanoleucus and Geranoaetus polyosoma (López et al. 2017b); Strix chacoensis (López et al. 2018); Tyto alba (Andrews 1990); Bubo virginianus nacurutu (Gómez 2005); Bubo virginianus magellanicus (Montalvo et al. 2016); Athene cunicularia (Montalvo and Tejerina 2009)
When small mammal bone samples from rockshelter archaeological sites were included in another PCA analysis, bone accumulations from open-air archaeological sites of the Monte Desert were also grouped and discriminated both from current predator bone samples and from small mammal bone accumulations recovered in rockshelter archaeological sites (Fig. 5 down; components 1 and 2 account for 83.04% of the variation; eigenvalues PC1 = 3048.98, PC2 = 1594.99). In addition, archaeological samples recovered for the present study on sandy soils were probably never incorporated into the sedimentary context of each site or did so very slowly.
Despite the high level of destruction by sedimentary features and by postdepositional processes, degrees and frequencies of digestion allowed to infer avian raptors (particularly owls) as accumulating agents of small mammal bone and teeth accumulations from open-air archaeological sites of the Monte Desert.
If the evaluation of digestion traces had not been considered, the high level of destruction and bone loss could lead to misinterpretations. Thus, these highly modified bone assemblages could be erroneously associated with carnivorous mammals, diurnal raptors or humans, which strongly modify their prey remains, both in frequency and intensity, and produce assemblages of primarily large-sized small mammal prey (e.g. López et al. 2017a, b; Fernández et al. 2017a, respectively).
Palaeoenvironmental considerations
Small mammal assemblages recovered in open-air archaeological sites for the present study are mainly composed by typical species connected to xeric environments from the Monte Desert. They differ among them, however, in their habitat use patterns, ecological preferences and the sites in which each species appears (Gonnet and Ojeda 1998; Braun and Mares 2002; Tabeni and Ojeda 2003; Corbalán 2004, 2006; Corbalán and Debandi 2005; Creighton and Gardner 2007; Ojeda et al. 2007; Albanese 2010; Díaz et al. 2015; Dunnum 2015; Patton et al. 2015). The didelphid T. pallidior is mostly present in open, low-complexity areas, although they can also be found both in shrubby areas and in more humid sites with dense vegetation along watercourses. Gerbil mouse Eligmodontia sp. frequents open areas with stripped sandy soils and disperse shrubs. Gray leaf-eared mouse G. griseoflavus prefers closed and complex habitats with trees and high shrub cover. Marsh rat H. lagigliai is only known for the type locality (El Nihuil, Mendoza) from one specimen collected in the 1950s (Fernández et al. 2017b). Taking into account that the species of genus Holochilus is adapted to a semiaquatic life, in riparian and swampy environments of the tropical lowlands (Gonçalves et al. 2015), the presence of H. lagigliai in arid and semi-arid lands could correspond to apparently sub-optimal areas related with the existence of permanent watercourses. The wetland where H. lagigliai was found (PA14.7; Lagunas de Guanacache) could serve during the late Holocene like a refuge to relictual populations of some subtropical mammals adapted to mesic conditions within the arid context of the Monte Desert (Roig 1991). This has also been suggested for other mammalian species, such as Chrysocyon brachyurus (Roig 1991; Fernández et al. 2017c).
Systematic trap works and an extensive analysis of recent owl pellets, both in the present study and in other ecological works, suggest the extinction of H. lagigliai in recent times (Contreras and Rosi 1980; Rodríguez and Ojeda 2013; Fernández et al. 2017c; Castillo et al. 2018). Viscacha rat T. barrerae is an endemic octodontid rodent from Central Western Argentina, distributed in the phytogeographic provinces of Monte and Patagonia, between 300 and 1400 masl. It presents a patchy distribution associated with salty (Ojeda et al. 2007), sandy and open shrubland environments and has been described as one of the rodent species best adapted to desert environments (Verzi et al. 2002; Díaz et al. 2015). Southern mountain cavy M. australis prefers shrubby and forested environments from the plains and piedmont, mainly (Sassi et al. 2011; Taraborelli et al. 2011). Lowland yellow-toothed cavy G. leucoblephara inhabits grasslands and shrubby environments with good vegetation cover (Ojeda 2006; Dunnum 2015). The genus Abrocoma is endemic from South America. Species of this genus inhabit mountain slopes from the piedmont and plains connected with rocky outcrops, scattered low shrubs and hard grasses with low cover and cacti (Braun and Mares 2002; Taraborelli et al. 2011). The Ctenomys species presents a wide distribution in all vegetation patches of the phytogeographic units from the study area (Rosi et al. 2002, 2005).
On the other hand, some species were not detected in the archaeological record, although they were identified in recent pellet samples. Dolorous grass mouse A. dolores is associated with shrubby steppes and thick grasslands (Corbalán 2004, 2006). Drylands verper mouse C. musculinus is connected with grasslands and scrublands of high cover, both in piedmont and plains. It is a widely distributed opportunistic species also abundant in areas strongly affected by intensive agriculture (e.g. Contreras and Rosi 1980; Fernández 2012, 2014; Teta et al. 2014). House mouse M. musculus is an exotic species distributed worldwide, accidentally introduced in America after the arrival of the European conquistadors. In South America, M. musculus has been reported since the middle of the sixteenth century (Novillo and Ojeda 2008). This species adapts quickly to new and varied environments, competing with native rodents (Novillo and Ojeda 2008; Ballari et al. 2016). It is a commensal species, which prefers habitats related with human activities mainly in urban and suburban areas, although it has also been detected in areas strongly affected by intensive agriculture from the Monte Desert of Mendoza (e.g. Contreras and Rosi 1980; Lunlunta and Paleocauce central in the present study).
In a recent study conducted in the study area, Oligoryzomys flavescens (yellow pygmy rice rat) was detected from pellets generated by Athene cunicularia in a lagoon context (Castillo et al. 2018; López 2018). That rodent species, of which neither archaeological nor current samples were detected in the present study, prefers open areas with grasslands near water bodies. It also inhabits the edges of cultivated lands (Weksler and Bonvicino 2015) being a typical component of Pampean agroecosystems (Cirignoli et al. 2006). In Mendoza, it has been detected in the central northern plains of Monte Desert, with a preference for herbaceous and shrubby vegetation associated with relatively humid sites or close to water bodies (Contreras and Rosi 1980; Fernández 2012).
The greater richness, evenness and diversity detected in small mammal archaeological assemblages compared with present-day communities reflect environmental conditions during the late Holocene different than during modern times (Figs. 6, 7 and 8). At the same time, the co-occurrence of species in the archaeological record with current allopatric distributions (non-analogue communities) also supports this idea. These changes in the taxonomic structure of archaeological small mammal assemblages might reflect minor or moderate environmental fluctuations capable of producing modifications in vegetation structure, water availability and the consequent availability of optimal habitats and microhabitats for each species.

Comparison of trends of diversity H′ and evenness between small mammal bone accumulations from open-air archaeological sites and from current pellet samples generated by several predators. Laguna de Soria—Athene cunicularia (Castillo et al. 2018; López 2018); San José—Tyto alba, Paleocauce central PA 46—Tyto alba and Lunlunta—Athene cunicularia (present study); Telteca—Strix chacoensis (López et al. 2018)


Boxplot of taxonomic composition of archaeological and recent small mammal bone accumulations from the Monte Desert. Top, diversity H′; down, evenness
Some palaeoenvironmental studies have highlighted the deep impact of human activities (mainly land clearing, overgrazing, intensive agriculture, urban development and trading) on the native landscape in the last 500 years with the arrival of European conquistadors to the study region. The economic activities mentioned above, were intensified during the last 150 years, generating drastic reductions, the disappearance and/or de-structuring of the vegetation communities (e.g. Prieto and Abraham 2000; Prieto et al. 2003; Villagra et al. 2009; Mafferra 2017; Castillo et al. 2018), strongly affecting soils and water availability. Such strong alterations in the landscape also negatively affected the existence and distribution of native mammals from the arid lands of South America and the Monte Desert in particular (Roig 1991; Tabeni and Ojeda 2003, 2005; Fernández 2014; Teta et al. 2014; López et al. 2016; Fernández et al. 2017b). In several cases, those impacts even influenced biological invasions of exotic species (Contreras and Rosi 1980; Castillo et al. 2018) such as M. musculus.
The effect of overgrazing and the consequent desertification in the Patagonian region, which would have strongly compacted the soils, have been identified as causes of the current extirpation or retraction of some subterranean species, like the Tympanoctomys genus, that would have had a wider and more continuous distribution in the past (Fernández 2014; Teta et al. 2014). The current patchy distribution of T. barrerae—not detected in current samples of the present study—could be generated by the impact of human activities and not by the original distribution of this species. Although C. musculinus is a native species connected with thick scrublands and grasslands, it has also been detected in areas strongly modified by intensive agriculture (Fernández 2014; Teta et al. 2014; Castillo et al. 2018; López 2018). The absence of this rodent in the archaeological record, and its presence and abundance in the recent small mammal assemblages, could also be related to the deep impact on native landscapes generated by intensive anthropic activities (e.g. intensive agriculture), added to the presence of the exotic M. musculus. Eligmodontia sp. increased its frequencies in recent pellet samples in comparison with those frequencies detected in the archaeological record. The increase of this species in the current samples has been associated with the impact on soils and vegetation generated by overgrazing and land clearing (Fernández 2014; Teta et al. 2014). The previously mentioned extinction of H. lagigliai has been associated with the intentional disappearance or the deep retraction of water bodies and wetlands due to the development of cities and the implementation of an intensive agriculture economic system after the arrival of the European conquerors since the middle of the sixteenth century. The handling of watercourses for purely productive and agricultural purposes (in a semi-arid area) negatively impacted the landscape and the availability of water (Fernández et al. 2017c; López 2018) also generating a major habitat uniformity and a simplified landscape.
The differences between past and present small mammal assemblages from the Monte Desert support the idea of a scenario of greater environmental heterogeneity during the late Holocene than in the present (López et al. 2016). This scenario may imply greater vegetation density and more availability of patch resources than in the present. Habitats with greater resource availability and diversity can sustain more species and higher diversities, whereas more available microhabitats offer more refuge against predators and provide more richness and diversity of food resources. Thus, as the number of microhabitats that can be occupied by small mammal species with different environmental requirements increases, this greater habitat complexity (vertical variation of vegetation) and heterogeneity (horizontal variation of vegetation) increase the coexistence of small mammal species (Cramer and Willig 2002; Corbalán 2004; Novillo et al. 2017).
The taxonomic trends detected in small mammal bone accumulations recovered in open-air archaeological sites in the present study are consistent with tendencies reported for small mammal bone accumulations in regional archaeological research at Central Western Argentina, for sets recovered in other types of sites, such as rockshelter sites (Fernández 2012; López et al. 2016; López 2018). Thus, the general impoverishment of current small mammal assemblages (lower richness, evenness and diversity than in the past; an increase in the representation of exotic and/or invasive species) in relation with past ones, has been detected both in open-air and rockshelter sites from Central Western Argentina. Despite the strong modifications generated by postdepositional processes and low site integrity and resolution, archaeological small mammal bone accumulation reflects original native Monte Desert environments during the late Holocene.
Conclusions
The results of the analysis of taphonomic variables of anatomical representation, breakage and digestion on small mammal bone and teeth remains recovered in open-air archaeological sites from the central Monte Desert (northeastern Mendoza) include identification of the accumulating agents as belonging to Andrews’ (1990) modification categories 1 and 2 (light and intermediate modifiers), such as owls. Despite the high abundance of medium- and large-sized small mammals, anthropic signals in such bone accumulations were not detected. The better preservation of the large-sized small mammals could be attributed to the differential preservation of bones and not to the hunting habits of predators. Small mammal bone and tooth accumulations recovered in open-air archaeological sites from the central Monte Desert showed high levels of destruction and bone loss. Thus, considerable proportions of elements affected by trampling and weathering were detected. Although these postdepositional processes modified the original features of bone assemblages, taphonomic traces (mainly digestion) were detected, providing the opportunity to infer the possible accumulating agents. Is it possible that another predator is responsible, someone we would not consider in cave contexts but which may be active in accumulating open-air assemblages? There are several predators in the study area, including strigiforms, which feed on small mammals and commonly accumulate small mammal bones in open-air contexts (e.g. López et al. 2017a, b, 2018). However, we need more actualistic taphonomic research to strengthen and systematise the knowledge on taphonomic signals generated by each predator on its prey.
The taxonomic structure of small mammal assemblages from archaeological contexts differed from the modern small mammal assemblages present in the area. Thus, richness, diversity and evenness were generally higher in the archaeological sets than in the current ones. Frequencies of the integrating species of archaeological assemblages were also different in comparison with the modern ones.
An extinct species, H. lagigliai, was detected in an archaeological site in a lagoon context. A little-known species highly adapted to desert environments was also detected in several archaeological sites (T. barrerae). Both the reported extinct species and the taxonomic composition of archaeological assemblages in comparison with the current ones suggest a scenario of greater environmental heterogeneity during the late Holocene than in the present. This situation may have allowed the coexistence of species with current allopatric distributions and the presence of H. lagigliai. The general impoverishment of modern small mammal assemblages in comparison with past ones could be related to the deep impact caused by the anthropic perturbations developed by economic activities on soils, vegetation and water availability since the arrival of European conquerors ca. 500 years BP and intensified during the last 150 years. The taxonomic and palaeoenvironmental implications of these processes has also been identified and discussed by research from other regions of southern South America, like Pampa and Patagonia (e.g. Fernández 2014; Teta et al. 2014). At the same time, Pardiñas and Teta (2013) have pointed a stability (despite minor variations) in the taxonomic structure of Holocenic small mammal assemblages from Patagonia, pointing out the strong changes when modern communities (last 150 years) are compared with past ones. This could be due both to Holocenic climate fluctuations that were not strong enough to produce deep changes in the taxonomic structure of small mammal communities and to the resilience of small mammal species to minor or moderate environmental fluctuations. This fact gives an idea of the impact generated by recent urban and economic activities on native landscapes, and particularly on small mammals (e.g. Tabeni and Ojeda 2003, 2005). As evidence of the effect of these changes, this study documents the difference between past and present small mammal assemblages.
More heterogeneous environments in the past could have supported greater diversity and resource availability around the landscape than in the present. This must be a factor to consider in regional archaeological research, considering the broader diet breath detected for human populations after ca. 2000 BP in the region. For this reason, new analytic variables are required to distinguish human and environmental factors to explain, discuss and understand the variability of the archaeological record and the cultural variability and evolution of the region. Despite differences from a taphonomic and palaeoenvironmental perspective, in general terms, small mammal bone accumulations from open-air archaeological sites have similar explanatory potentials to those reported for small mammal bone accumulations recovered in rockshelter sites. The former, however, are in worse conservation conditions and have lower integrity and resolution than the latter. Integrating small mammal bone accumulations from different site types in Central Western Argentina will certainly contribute to fuller, more detailed interpretations of past environments and the evolution of the human systems through time. At the same time, we expect that the taphonomic trends detected in open-air archaeological sites from the Monte Desert in Central Western Argentina contribute to the discussion of the generation, accumulation, recovery, potential and importance of small mammal bone and tooth accumulations recovered in open-air archaeological sites around the world.
References
Abraham E, Prieto MR (1981) Enfoque diacrónico de los cambios ecológicos y de las adaptaciones humanas en el NE árido mendocino. Cuadernos del CEIFAR 8:109–139
Abraham E, Del Valle HF, Roig F, Torres L, Ares JO, Coronato F, Godagnone YR (2009) Overview of the geography of the Monte Desert biome (Argentina). J Arid Environ 73:144–153
Albanese MS (2010) Ecología de la marmosa pálida Thylamys pallidior (Marsupialia: Didelphidae) en el Desierto de Monte central. Unpublished Ph.D. thesis, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires
Andrews P (1990) Owls, caves and fossils. Predation, preservation, and accumulation of small mammal bones in caves, with an analysis of the Pleistocene cave faunas from Westbury—sub-Mendip, Somerset, U.K. University of Chicago Press, Chicago
Andrews P (1995) Mammals as palaeoecological indicators. Acta Zool Cracov 38:59–72
Andrews P, Armour-Chelu M (1998) Taphonomic observations on a surface bone assemblage in a temperate environment. Bull. Soc. Géol Fr 169:433–442
Ballari SA, Anderson CB, Valenzuela AE (2016) Understanding trends in biological invasions by introduced mammals in southern South America: a review of research and management. Mammal Rev 46:229–240
Braun JK, Mares MA (2002) Systematics of the Abrocoma cinerea species complex (Rodentia: Abrocomidae), with a description of a new species of Abrocoma. J Mammal 83(1):1–19
Bruniard E (1982) La diagonal árida argentina: un límite climático real. Revista Geográfica IPGH 95:5–20
Cahiza P, Ots MJ (2014) Análisis distribucional de sociedades de pequeña escala en el Centro Oeste Argentino. Discusión de las estrategias metodológicas y las interpretaciones en el marco de los estudios regionales. In: Falabella FG, Sanhueza LR, Cornejo LB, Correa I (eds) Distribución espacial en sociedades no aldeanas: del registro arqueológico a la interpretación social, Serie Monográfica 4, Ediciones on demand edn. Sociedad Chilena de Antropología, Chile, pp 199–219
Castillo L, López JM, Chiavazza H (2018) El registro de micromamíferos en el Arroyo Tulumaya: punto arqueológico 70 (noreste de Mendoza). Aspectos tafonómicos e implicancias ambientales para los últimos 500 años. Revista de Arqueología Histórica Latinoamericana y Argentina 10:454–478
Chiavazza H (2007) Cambios ambientales y sistemas de asentamiento en el árido Normendocino. Arqueología en los paleocauces del Río Mendoza. Unpublished PhD thesis, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Argentina
Chiavazza H (2009) Garganta seca y arena en las botas: prospectanto antecedentes arqueológicos de las tierras áridas del noreste mendocino (Centro Oeste Argentino). Arqueología Iberoamericana 1:41–77
Chiavazza H (2012) Ocupaciones humanas en la planicie árida noreste de Mendoza: estudios arqueológicos en el Paleocauce Central (PC4). Relaciones de la Sociedad Argentina de Antropología 37:299–327
Chiavazza H (2014) Tendencias regionales del registro arqueológico de sociedades sin estructura aldeana del noreste de Mendoza (Argentina). In: Falabella FG, Sanhueza LR, Cornejo LB, Correa I (eds) Distribución espacial en sociedades no aldeanas: del registro arqueológico a la interpretación social, Serie Monográfica 4, Ediciones on demand. Sociedad Chilena de Antropología, Chile, pp 221–249
Chiavazza H (2016) Vivir y moverse en el desierto: la ocupación humana en entornos áridos (Noreste de Mendoza, Argentina). Sociedades de Paisajes Áridos y Semi-áridos 9:8–34
Chiavazza H, Acosta A, Quiroga M, Castillo L, Tobar V, Frías C, Castañar K, Puebla F, Anzorena J, Puebla L, Giannotti S, Guajardo V, Romera A, Hernández F (2010) Investigaciones arqueológicas en la Reserva Natural Villavicencio (RNV): Ambiente y Cultura en la transición del piedemonte precordillerano de Mendoza. Primeros Resultados. In: Bárcena JR, Chiavazza H (eds) Arqueología Argentina en el Bicentenario de la Revolución de Mayo. Zeta, Mendoza, pp 2071–2076
Cirignoli S, Teta P, Pardiñas UFJ, D’Elía G (2006) Tribu Oryzomyini. In: Barquez RM, Díaz M, Ojeda RA (eds) Mamíferos de Argentina. Sistemática y distribución. Sociedad Argentina para el Estudio de los Mamíferos (SAREM). Argentina, pp 166–175
Contreras JR, Rosi MI (1980) Comportamiento territorial y fidelidad al hábitat en una población de roedores del centro de la Provincia de Mendoza. Ecología Argentina 5:17–29
Corbalán V (2004) Uso de hábitat y ecología poblacional de pequeños mamíferos del Desierto del Monte Central, Mendoza, Argentina. Unpublished PhD thesis, Universidad Nacional de La Plata. 199 pp.
Corbalán V (2006) Microhabitat selection by murid rodents in the Monte desert of Argentina. J Arid Environ 65:102–110
Corbalán V, Debandi G (2005) Microhabitat use by Eligmodontia typus (Rodentia: muridae) in the Monte Desert (Argentina). Mamm Biol 71:124–127
Cramer MJ, Willig MR (2002) Habitat heterogeneity, habitat associations, and rodent species diversity in a sand shinnery oak landscape. J Mammal 83:743–753
Creighton GK, Gardner AL (2007) Genus Thylamys. In: Gardner AL (ed) Mammals of South America, Marsupials, Xenarthrans, shrews, and bats, vol 1. The University of Chicago Press, Chicago, pp 107–117
De Santis LJM, Roig VG, Justo ER (1991) La anatomía cráneo-dentaria de Tympanoctomys barrerae (Lawrence). Comparación con Octomys mimax y consideraciones acerca de su estado taxonómico (Rodentia: Octodontidae). Neotropica 37(98):113–122
Denys C, Dauphine Y, Fernández-Jalvo Y (1997) Apports biostratigraphiques et paléoécologiques de l’étude taphonomique des assemblages de micromammiféres. Geobios 20:197–206
Díaz MM, Barquez R, Verzi DH (2015) Genus Tympanoctomys. In: Patton JL, Pardiñas UFJ, D’Elía G (eds) Mammals of South America: vol. 2. Rodents. University of Chicago Press, Chicago, pp 1043–1048
Dunnum JL (2015) Family Caviidae. In: Patton JL, Pardiñas UFJ, D’Elía G (eds) Mammals of South America: vol. 2. Rodents. University of Chicago Press, Chicago, pp 690–725
Fernández FJ (2012) Microvertebrados del Holoceno de sitios arqueológicos en el sur de Mendoza (República Argentina): aspectos tafonómicos y sus implicancias en la subsistencia humana. PhD thesis, Universidad Nacional La Plata, 520 pp.
Fernández FJ (2014) Micromamíferos como indicadores de deterioro ambiental (antrópico) en el sur de Mendoza. In: Hucailuk C, Mercado I (Comps) E-ICES 10. Comisión Nacional de Energía Atómica, Buenos Aires, pp 282–293
Fernández FJ, Pardiñas UFJ (2018) Small mammals taphonomy and environmental evolution during Late Pleistocene-Holocene in Monte Desert: the evidence of Gruta del Indio (central West Argentina). J S Am Earth Sci 84:266–275
Fernández FJ, Moreira G, Neme G, De Santis LJM (2009) Microvertebrados exhumados del sitio arqueológico “Cueva Arroyo Colorado” (Mendoza, Argentina): aspectos tafonómicos y significación paleoambiental. Archaeofauna 18:99–118
Fernández FJ, del Papa LM, Moreira G, Prates L, De Santis LJM (2011a) Small mammal remains recovered from two archaeological sites in the middle and lower Negro River valley (Late Holocene, Argentina): Taphonomic issues and paleoenvironmental implications. Quat Int 245:136–147
Fernández FJ, Ballejo F, Moreira G, Tonni E, De Santis LJM (2011b) Roedores cricétidos de la provincia de Mendoza. Guía cráneo-dentaria orientada para su aplicación en estudios zooarqueológicos. Sociedad Argentina de Antropología-Universitas Sarmiento, Buenos Aires 115 pp
Fernández FJ, Neme G, De Santis LJM (2015a) Micromammals, taphonomy, paleoenvironments and human occupation during the Holocene in Arroyo Malo-3 (west-central Argentina). Archaeofauna 24:27–52
Fernández FJ, Rudzik SM, Neme G, De Santis LJM (2015b) Micromamíferos, tafonomía y ambientes durante los últimos 3.800 años A.P. en el Valle Medio del Río Grande, Mendoza, Argentina. Chungara 47:267–285
Fernández FJ, Teta P, Pardiñas UFJ (2017a) Evidencias arqueológicas de explotación antrópica de micromamíferos en el extremo austral de América del Sur. Anales de Arqueología y Etnología 72(1):9–32
Fernández FJ, Montalvo CI, Fernández-Jalvo Y, Andrews P, López JM (2017b) A re-evaluation of the taphonomic methodology for the study of small mammal fossil assemblages of South America. Quat Sci Rev 155:37–49
Fernández FJ, Torres J, Tammone M, López JM, Pardiñas UFJ (2017c) New data on the endemic cricetid rodent Holochilus lagigliai from central western Argentina: fossil record and potential distribution. Mammalia 81:621–625
Fernández-Jalvo Y, Andrews P (1992) Small mammal taphonomy of gran Dolina, Atapuerca (Burgos), Spain. J Archaeol Sci 19:407–428
Fernández-Jalvo Y, Scott L, Andrews P (2011) Taphonomy in palaeocological interpretations. Quat Sci Rev 30:1296–1302
Fernández-Jalvo Y, Andrews P, Sevilla P, Requejo V (2014) Digestion versus abrasión features in rodent bones. Lethaia 47:323–336
García Llorca J, Cahiza P (2007) Aprovechamiento de Recursos Faunísticos en las Lagunas de Guanacache (Mendoza, Argentina). Análisis zooarqueológico de La Empozada y Altos de Melién II. Chungara 39:117–133
Garvey R, Bettinger R (2018) A regional approach to prehistoric landscape use in west-central Argentina. J Archaeol Sci Rep 19:846–855
Gómez GN (2005) Analysis of bone modifications of Bubo virginianus pellets from Argentina. Journal of Taphonomy 3:1–16
Gonçalves P, Teta P, Bonvicino C (2015) Genus Holochilus. In: Patton JL, Pardiñas UFJ, D’Elía G (eds) Mammals of South America: vol. 2. Rodents. University of Chicago Press, Chicago, pp 325–334
Gonnet JM, Ojeda RA (1998) Habitat use by small mammals in the arid Andean foothills of the Monte Desert of Mendoza, Argentina. J Arid Environ 38:349–357
Hammer Ø, Harper D, Ryan P (2001) PAST: Paleontological Statistics software package for education and data analysis. Paleontología Electrónica 4:1–9
Krebs CJ (1989) Ecological methodology. Harper and Row Publishers, New York
Labraga JC, Villalba R (2009) Climate in the Monte Desert: past trends, present conditions, and future projections. J Arid Environ 73:154–163
López JM (2018). Restos óseos de micromamíferos recuperados en sitios arqueológicos del norte de Mendoza para contextos correspondientes al Holoceno tardío: un enfoque zooarqueológico, tafonómico y paleoambiental. Unpublished PhD thesis, Facultad de Filosofía y Letras, Universidad Nacional de Cuyo
López JM, Chiavazza H, Rosi MI (2016) Small mammal remains recovered from two archaeological sites in northwestern Mendoza (late Holocene, Argentina): taxonomic composition, taphonomic issues, and paleoenvironmental implications. Quat Int 391:26–37
López JM, Rosi MI, Tabeni S, Bender B, Chiavazza H (2017a) Taphonomic analysis of small mammal bone remains preyed upon by wildcats (Carnivora: Felidae) from the Central Monte Desert. Boreas 46:282–293
López JM, Fernández FJ, Montalvo CI, Chiavazza H, De Santis LJM (2017b) The role of the Accipitriformes Geranoaetus melanoleucus and Geranoaetus polyosoma as small mammal bones accumulators in modern and archaeological sites from Central Western Argentina. Journal of Taphonomy 15(1–3):91–108
López JM, Fernández FJ, Castillo L, Pereyra-Lobos R (2018) Taphonomy of small mammal bone accumulations generated by the Chaco owl (Strix chacoensis, Strigiformes) from the Monte Desert (Mendoza, Argentina): a contribution to South American archaeological and palaeontological studies. Boreas 47:780–791
Lyman RL (2008) Quantitative Paleozoology. Cambridge University Press, Cambridge
Mafferra LE (2017) Los paisajes forestales en torno a la ciudad colonial de Mendoza, con base en el registro antracológico. Intersecciones Antro 18:43–53
Medina ME, Teta P, Rivero D (2012) Burning damage and small-mammal human consumption in Quebrada del Real 1 (Cordoba, Argentina): an experimental approach. J Archaeol Sci 39 (3):737–743
Montalvo CI, Tejerina P (2009) Análisis tafonómico de los huesos de anfibios y roedores depredados por Athene cunicularia (Strigiformes, Strigidae) en La Pampa, Argentina. In: Berón M, Luna L, Bonomo M, Montalvo CI, Aranda C, Carrera Aizpitarte M (eds) Mamül Mapu: pasado y presente desde la arqueología pampeana. Editorial Libros del Espinillo, Buenos Aires, pp 323–334
Montalvo CI, Fernández FJ, Tallade PO (2016) The role of Bubo virginianus magellanicus as rodent bone accumulator in archaeological sites: a case study for the Atuel River (Mendoza, Argentina). Int J Osteoarchaeol 26:974–986
Navarro D, Rojo L, De Francesco C, Hassan GS (2012) Paleoecología y reconstrucciones paleoambientales en Mendoza durante el Holoceno. In: Gil AF, Neme G (eds) Paleoecología humana en el sur de Mendoza. Sociedad Argentina de Antropología, Buenos Aires, pp 17–56
Novillo A, Ojeda RA (2008) The exotic mammals of Argentina. Biol Invasions 10:1333–1344
Novillo A, Cuevas MF, Ojeda AA, Ovejero RJA, Torres M, Eugenia M, Ojeda RA (2017) Habitat selection and coexistence in small mammals of the southern Andean foothills (Argentina). Mammal Research 62(3):219–227
Ojeda RA (2006) Familia Caviidae. In: Barquez R, Díaz M, Ojeda RA (eds) Mamíferos de Argentina. Sistemática y Distribución. Sociedad Argentina para el estudio de los Mamíferos, Argentina, pp 206–210
Ojeda RA, Mares MA (1982) Conservation of South American mammals: Argentina as a paradigm. In: Mares MA, Genoways H (eds) Mammalian biology in South America. The University of Pittsburgh Press, Pittsburgh, pp 505–521
Ojeda RA, Borghi CE, Roig VG (2002) Mamíferos de Argentina. In: Ceballos G, Simonetti JA (eds) Biodiversidad y conservación de mamíferos neotropicales. CONABIO, México, pp 23–63
Ojeda AA, Gallardo MH, Mondaca F, Ojeda RA (2007) Nuevos registros de Tympanoctomys barrerae (Rodentia, Octodontidae). Mastozoología Neotropical 14(2):267–270
Ortíz PE, Madozzo Jaén M, Jayat P (2012) Micromammals and paleoenvironments: climatic oscillations in the Monte desert of Catamarca (Argentina) during the last two millenia. J Arid Environ 77:103–109
Ots MJ, García Llorca J, Cahiza P (2016) Recursos y estrategias de consumo en el centro de Mendoza entre los siglos X-XVI AD. Intersecciones Antro 17:375–387
Pardiñas UFJ (1999) Tafonomía de microvertebrados en yacimientos arqueológicos de Patagonia (Argentina). Arqueología 9:265–340
Pardiñas UFJ, Teta P (2013) Holocene stability and recent dramatic changes in micromammalian communities of northwestern Patagonia. Quat Int 305:127–140
Patton JL, Pardiñas UFJ, D’elía G (2015) Mammals of South America, vol 2. 1336 pp. University of Chicago Press, Chicago
Prieto MR, Abraham E (2000) Caminos y comercio como factores de cambio ambiental en las planicies áridas de Mendoza (Argentina) entre los siglos XVII y XIX. Revista Theomai 2 15pp
Prieto MR, Chiavazza H (2006) Aportes de la historia ambiental para el análisis del patrón de asentamiento huarpe en el oasis norte de Mendoza. Anales de Arqueología y Etnología 58–59:163–195
Prieto Olavarría C, Chiavazza H, Porta V, Bontorno E (2016) Variabilidad alfarera y procesos ambientales en un sitio de límite cultural. Estilos tecnológicos de la cerámica del río Desaguadero (PA68) (Mendoza, Centro Oeste Argentino). Relaciones de la Sociedad Argentina de Antropología 41:131–150
Prieto MR, Rojas F (2012) Documentary evidence for changing climatic and anthropogenic influences on the Bermejo Wetland in Mendoza, Argentina, during the 16th–20th century. Clim Past 8:951–961
Prieto MR, Villagra PE, Lana NB, Abraham E (2003) Utilización de documentos históricos en la reconstrucción de la vegetación de la Llanura de la Travesía (Argentina) a principios del siglo XIX. Rev Chil Hist Nat 76:613–622
Prieto MR, Abraham E, Dussel P (2008) Transformaciones de un ecosistema palustre. La gran ciénaga del Bermejo-Mendoza, siglos XVIII y XIX. Multequina 17(2):147–164
Prieto MR, Rojas F, Castrillejo T, Hernández F (2012) Procesos ambientales y construcción del territorio a partir de un estudio de caso: la ciénaga del Bermejo, oasis Norte de Mendoza, 1810-1930. Revista de Historia Americana y Argentina 47:1–29
Quintana CA (2016) Tafonomía de los microvertebrados del sitio arqueológico Cueva Tixi (Pleistoceno tardío-Holoceno tardío), Tandilia Oriental (provincia de Buenos Aires, Argentina). Online publication of Asociación Paleontológica Argentina (PEAPA) 16:14–51
Rodríguez D, Ojeda RA (2013) Scaling coexistence and assemblage patterns of desert small mammals. Mamm Biol 78(5):313–321
Roig VG (1991) Desertification and distribution of mammals in the southern cone of South America. In: Mares MA, Schmidly D (eds) Latin American Mammalogy. University of Oklahoma Press, Norman, pp 239–279
Rosi MI, Cona M, Roig VG (2002) Estado actual del conocimiento del roedor fosorial Ctenomys mendocinus Philippi 1869 (Rodentia: Ctenomyidae). Mastozoología Neotropical 9:277–295
Rosi MI, Cona M, Roig VG, Massarini AI, Verzi D (2005) Ctenomys mendocinus. Mamm Species 777:1–6
Rundel PW, Villagra PE, Dillon MO, Roig-Juñent S, Debandi G (2007) Arid and semi-arid ecosystems. In: Veblen T, Young K, Orme A (eds) The physical geography of South America. Oxford University Press, Oxford, pp 158–183
Sassi P, Chiappero P, Borgui C, Gardenal C (2011) High genetic differentiation among populations of the small cavy Microcavia australis occupying different habitats. J Exp Zool 315:337–348
Scheifler N, Teta P, Pardiñas UFJ (2012) Small mammals (Didelphimorphia and Rodentia) of the archaeological site Calera (Pampean region, Buenos Aires Province, Argentina): Taphonomic history and Late Holocene environments. Quat Int 278:32–44
Stahl PW (1996) The recovery and interpretation of microvertebrate bone assemblages from archaeological contexts. J Archaeol Method Th 3:31–75
Tabeni S, Ojeda RA (2003) Assessing mammal responses to perturbations in temperate aridlans of Argentina. J Arid Environ 55:715–726
Tabeni S, Ojeda RA (2005) Ecology of the Monte Desert small mammals in disturbed and undisturbed habitats. J Arid Environ 63:244–255
Taraborelli P, Moreno P, Sassi P, Dacar M, Ojeda RA (2011) New eco-morphological-behavioural approach of the chinchilla rats in the pre-Andean foothills of the Monte Desert (Argentina). J Nat Hist 45:25–28
Teta P, Pardiñas UFJ, Silveira M, Aldazabal V, Eugenio E (2013) Roedores sigmodontinos del sitio arqueológico “El Divisadero Monte 6” (Holoceno tardío, Buenos Aires, Argentina): taxonomía y reconstrucción ambiental. Mastozoología Neotropical 20(1):171–177
Teta P, Formoso A, Tammone M, de Tommaso DC, Fernández FJ, Torres J, Pardiñas UFJ (2014) Micromamíferos, cambio climático e impacto antrópico: ¿Cuánto han cambiado las comunidades del sur de América del Sur en los últimos 500 años? Therya 5:7–38
Verzi DH, Tonni E, Scaglia OA, San Cristóbal JO (2002) The fossil record of the desert-adapted South American rodent Tympanoctomys (Rodentia, Octodontidae). Paleoenvironmental and biogeographic significance. Palaeogeogr Palaeocl 179:149–158
Villagra PE, Defossé GE, del Valle HF, Tabeni S, Rostagno M, Cesca E, Abraham E (2009) Land use and disturbance effects on the dynamics of natural ecosystems of the Monte Desert: implications for their management. J Arid Environ 73:202–211
Weksler M, Bonvicino C (2015) Genus Oligoryzomys. In: Patton JL, Pardiñas UFJ, D’Elía G (eds) Mammals of South America: vol. 2. Rodents. The University of Chicago Press, Chicago, pp 417–437
Acknowledgements
We wish to thank two anonymous reviewers for their accurate, clear and useful suggestions and comments that greatly improved our manuscript. This research was carried out in the framework of the following projects: Project 06/G724 (SECTyP, UNCuyo, Argentina): "Ocupaciones humanas en el Monte Árido III (OHMA_III): variabilidad arqueológica en escenarios diferenciados del Monte entre los 2000 y 300 años AP"; Project 05-G (Facultad de Ciencias Exactas y Naturales, UNLPam, Argentina): "Análisis tafonómico de huesos de vertebrados depredados por aves rapaces y mamíferos carnívoros".
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interests
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
ESM 1
Brief description of general features of the open-air archaeological sites from the Monte Desert and its archaeological record. (DOCX 16.7 kb)
Rights and permissions
About this article

Cite this article
López, J.M., Chiavazza, H. Amidst wind, sand and raptors. Small mammal bone remains recovered in open-air archaeological sites from the Monte Desert in Central Western Argentina: taphonomic and palaeoenvironmental implications. Archaeol Anthropol Sci 11, 5149–5169 (2019). https://doi.org/10.1007/s12520-019-00846-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12520-019-00846-z