
Abstract
The present study analyses small mammal bone and tooth accumulations recovered in three open-air archaeological sites from northern Mendoza (Argentina) in the central Monte Desert, one of the most arid rangelands of South America. The sites, with radiocarbon dates between ca. 2100 and 400 years BP, are located on the margins of a now-extinct swamp that formed a more widespread wetland environment in the past. In order to recognize the agents responsible for such bone and tooth accumulations, a taphonomic analysis was conducted evaluating relative abundances of skeletal elements, breakage patterns, digestive corrosion, signs of anthropic activity and post-depositional processes. The taphonomic analysis allowed the detection of owls and humans as the agents responsible for small mammal accumulations. On the one hand, the low proportion and degree of digested diagnostic elements, among other taphonomic processes, suggest owl pellet-derived small mammal assemblages. On the other hand, the thermo-altered elements detected, some showing a differential burning pattern, the abundance of large-sized and gregarious small mammals and the identification of cut-marks on a caviid femur shaft are possibly due to human exploitation/consumption of small mammals.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Vertebrate bone remains recovered in archaeological contexts have a long history of research interest (e.g. Olsen 1971; Gifford-González 2018). Increasingly, zooarchaeological and taphonomic analyses have been established as valuable tools to understand the processes and agents involved in the formation of the archaeological record, the evolution of human and environmental systems through time and the dynamics of the human and animal world existing in the past (Efremov 1940; Behrensmeyer et al. 2000). However, small vertebrate remains recovered from archaeological sites, including small mammals (≤ 1 kg), have generally been overlooked in archaeological studies for various reasons. Some of them are difficulties related to taxonomic determination/ deficiencies in the recovery of fieldwork samples, the lack of a coherent theoretical-methodological model applied to the study of such remains at a regional perspective, the small size bone remains associated with low energy return and an often unfounded assumption that past human groups did not exploit small vertebrates or that such exploitation can hardly be demonstrated archaeologically (Simonetti and Cornejo 1991; Pardiñas 1999; Fernández et al. 2017a).
For the past decades, a considerable increase in the research of small mammal remains recovered from archaeological sites in different regions of the world has demonstrated the potential of these remains to strengthen the discussion and understanding of human systems in the past and their evolution over time (e.g. Simonetti and Cornejo 1991; Andrade 2015; Andrade and Fernández 2017; Fernández et al. 2017a; Fernández-García et al. 2018, 2020; López-García et al. 2018; Mignino et al. 2018; Rhodes et al. 2018, 2019). Most of the research carried out in South America has been focused on the Pampean, Patagonian and Northwestern regions of Argentina (Frontini and Escosteguy 2015; Fernández et al. 2017a, and references therein). Some works have assessed archaeological small mammal bone and tooth accumulations recovered in Central Western Argentina (CWA) from a zooarchaeological, taphonomical and palaeoenvironmental perspective. Most of them were conducted in the southern sector of the area—33°–34° S—(Gasco et al. 2006; Fernández et al. 2009, 2015a, 2015b; Fernández 2012; Fernández and Pardiñas 2018), whereas in the northern sector—32°–33° S—this type of research is still scarce (López et al. 2016; Castillo et al. 2018a; López and Chiavazza 2019, 2020). These research works have contributed to our knowledge and discussion of modern and past small mammal assemblages and their significance in palaeoecological terms, identifying landscapes more heterogeneous during the Late Holocene than in present times. In this regard, this growing body of research has generally been oriented towards the discussion of the biological/palaeoecological significance of small mammal remains recovered from regional archaeological sites. Comparatively, the distinction of taphonomic patterns of anthropic activity on small mammal remains and the discussion of human-resource relationships over time has been poorly explored (Fernández 2012; Fernández et al. 2017a; López et al. 2019a). A clear example of this trend can be found in the extensive actualistic taphonomic research carried out in CWA on the signals that several non-human predators generate on small mammal bone and tooth accumulations as a result of their predation (e.g. Montalvo et al. 2016a, 2016b; Montalvo and Fernández 2019; López et al. 2017a, b, 2018, 2019b; López 2020).
Few regional zooarchaeological studies have systematically and suitably analysed bone sets of “non-classical” fauna (small fauna, such as small mammals and fishes) following appropriate and specific methodologies (e.g. Fernández 2012; Fernández et al. 2009, 2015a, 2015b; López et al. 2016; Corbat et al. 2017; Castillo et al. 2018a; López and Chiavazza 2019). A lack of interest in systematic studies of small mammal remains has been one of the reasons why regional zooarchaeological research assumes, often erroneously, the cultural or intrusive entity of such remains recovered in archaeological sites by applying neither suitable nor specific methodologies (e.g. Chiavazza 2007, 2010; Ots et al. 2016). In this respect, a zooarchaeological and taphonomic analysis of small mammal bone and tooth accumulations can assess, discuss and broaden our understanding of the taphonomic patterns generated by possible accumulating agents in archaeological sites, including humans. At the same time, it can also contribute to the study of the subsistence of human groups in the past and its variations over time, as shown in other regions of southern South America, such as central Chile, Pampa and Patagonia (e.g. Simonetti and Cornejo 1991; Pardiñas 1999; Alcaráz 2015; Santiago et al. 2016; Fernández et al. 2017a).
In recent times, appropriate recovery techniques have been incorporated in field and laboratory work in the arid zones of South America, particularly the drylands of CWA. Specific and systematic taphonomic analyses of zooarchaeological remains corresponding to lower-ranked resources have also grown in number and with a more cultural perspective. These include the study of small fishes and small mammals, apparently captured without the necessary technologies to enable mass capture (Corbat et al. 2017; López et al. 2019a). This new framework was triggered by growing interest in the ability of theoretical concepts such as Human Behavioural Ecology (HBE) to address problems related to the imbalance between resources and demography, including resource depression and intensification processes (sensu Broughton 1994; Broughton et al. 2011; Morgan 2015), which have been little explored in CWA (Neme 2007; Otaola 2013; Neme et al. 2015; López et al. 2019a).
Archaeological research conducted in CWA and guided by HBE has detected a broader dietary breadth within human groups since ca. 2000 years BP, associated with the incorporation of agriculture, pottery and the occupation of new areas related with a rapid human population growth and an increasing imbalance between humans and resources (e.g. Cortegoso 2006; Chiavazza 2013; Gil et al. 2014; Ots et al. 2016; López et al. 2019a). Analyses of archaeological contexts from northern Mendoza after ca. 2000 BP indicate a more diversified economy than in previous periods. This has been connected with the incorporation of lower-ranked resources in the human diet as a result of high-ranked resource depression after ca. 2000 BP (Corbat et al. 2017; López et al. 2019a). The most profitable regional prey items for these human groups were guanaco (Lama guanicoe) and armadillos—mainly Zaedyus pichiy, Chaetophractus villosus and Chaetophractus vellerosus—(Corbat et al. 2017).
The present study analyses small mammal bone and tooth accumulations recovered from three archaeological sites in northern Mendoza, the northern sector of Central Western Argentina. It aims to assess the possible accumulating agents of such bone sets and carry out a taphonomic analysis to distinguish natural and anthropic agents in each zooarchaeological assemblage, including the evaluation of post-depositional processes. Research results are discussed in the framework of a possible resource depression and intensification process (declining foraging efficiency) after ca. 2000 BP in northern Mendoza (López et al. 2019a).
Study area and archaeological contexts
Central Western Argentina is an area with significant environmental, archaeological and ethnographic diversity. It has been considered a peripheral and transitional region between cultural areas from North Western Argentina to the north (NWA) and Patagonia to the south. The modern climate is arid to semi-arid, with annual rainfalls ranging between 100 and 350 mm, occurring mainly in summer. The region also experiences highly variable temperatures and considerable seasonal climate differences (Rundel et al. 2007). Few rivers cross the area from west (Andean cordillera) to east (lowlands plains). Water availability in the region depends on the melting of glaciers and snow in the Andean cordillera. The study area is located within the South American Arid Diagonal (Abraham et al. 2020).
The Monte Desert, the most arid rangeland of Argentina, where the archaeological sites of the present study are located, is a phytogeographic unit that covers more than 50% of the territory of Central Western Argentina, located east of the cordilleran area. It is an arid to semi-arid area characterized by shrubby steppes of Larrea spp. and Bulnesia retama species, edaphic steppes of halophytes and arboreum stratums dominated by Prosopis flexuosa and Geoffroea decorticans (Rundel et al. 2007; Abraham et al. 2009). One of the most distinctive features of the Monte is the varied composition of its mammal fauna, particularly small mammals (Ojeda et al. 2002).
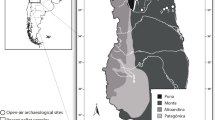
The small mammal bone and tooth accumulations analysed in the present study were recovered during fieldwork from open-air archaeological sites in the Monte Desert lowlands of northern Mendoza, using a 1.5-mm screen mesh. The sites are located on the margins of a recently drained large swamp that covered 58.000 ha, part of an extensive wetland environment that occupied the east of Mendoza until the beginnings of the twentieth century. This wetland environment was quite different from the present landscape, which is characterized by general aridity with scarce water sources, urban development, massive implantation of exotic flora (mainly trees) and an intensive agricultural system of wine production since the mid-nineteenth century (Chiavazza 2011; Prieto and Chiavazza 2006; Prieto and Rojas 2012; Mafferra 2017, 2018; López and Chiavazza 2019). The taxonomic composition of the small mammal assemblages in the present study reflects that wetland environment, indicating a considerable recent loss of biodiversity (extirpation and extinction of small mammals) (López 2018; López and Chiavazza, ms). The sites have 14C chronologies dating the deposits between ca. 2100 and 400 years BP (Fig. 1; Table 1). Two of the sites are located in the low piedmont of the Andean cordillera, in Mendoza city, Ruinas de San Francisco sector Pilastra Noroeste (SF-PNO) and Memorial de la Bandera (MB), whereas the site Punto Arqueológico 13.1 (PA13.1) is located in a sector of eastern plains associated to the Mendoza river course, in the locality of San José (Lavalle). Human occupation of the piedmont and plains of the Monte Desert from CWA occurred after ca. 2000 BP and can be considered a marginal area in comparison with the Andean precordillera to the west, which was occupied since ca. 11,000 BP and associated with the exploitation of wild camelids, the highest-ranked resources of the region (Chiavazza 2007; Corbat et al. 2017).

Top: Study area in phytogeographic perspective and archaeological sites of the present study: 1, Memorial de la Bandera (MB); 2, Ruinas de San Francisco sector Pilastra Noroeste (SF-PNO); 3, Punto Arqueológico 13.1 (PA13.1). Small mammal bone accumulations previously studied in Central Western Argentina: 4, Agua de la Tinaja (López et al. 2020); 5, Rincón de los Helados; 6, Vaquería Gruta 1 (López et al. 2016); 7, Vaquería Gruta 2 (López and Chiavazza 2020); 8, Agua de los Pajaritos; 9, PA52; 10, PA14.7; 11, PA24.2; 12, PA46.4; 13, PA68.1 (López and Chiavazza 2019); 14, Laguna el Sosneado; 15, Arroyo Malo 3; 16, Cueva Palulo; 17, Cueva Arroyo Colorado (Fernández 2012); 18, Gruta del Indio (Fernández and Pardiñas 2018); 19, Rincón del Atuel 1; 20, Agua de los Caballos; 21, Agua de la Mula; 22, Cueva de Luna; 23, La Peligrosa (Fernández 2012). Area of Bermejo swamp was recovered from Prieto and Rojas (2012). Down: Schematic profile of the relief in the study area (northern Mendoza) from a biogeographic and geomorphologic perspective
SF-PNO (32° 52′ S; 68° 49′ W; ca. 738 m a.s.l.) is a sector of a colonial Franciscan church, located a few metres west of the Cacique Guaymallén irrigation canal (existing since prehistoric times and still extant). Despite historical connotations of this site, thermoluminiscence dating corresponding to a pottery fragment resulted in 530 ± 50 BP (UCTL 1971)—290–300 cm depth—. In other sector of the same temple, but at the same depth, a charcoal sample was radiocarbonic dated to 440 ± 40 BP (URU 0279), pointing to human occupations previous to historical times. An area of 8 m2 (4 × 2 m) was excavated in arbitrary layers of 5 cm until a depth of 410 cm, distinguishing silty to clayey soils, mainly (Prieto Olavarría and Chiavazza 2009; Chiavazza et al. 2010). Small mammal remains were mainly recovered under 100 cm, in temporal contexts before the eighteen century. Medium-sized mammals, armadillos, small fishes and camelids (Castillo et al. 2018b) dominated the zooarchaeological record. Both MB and SF-PNO are close to one another, separated by only 1.5 km between them, in the same urban area and sedimentary context. This fact is interesting, taking into account their different chronology, which also allow evaluating them diachronically.
MB (32° 53′ S; 68° 50′ W; ca. 780 m a.s.l.) was detected in archaeological rescue works and was excavated to a depth of 460 cm, divided into 1-m2 grids, which became one of the largest archaeological excavations conducted in Central Western Argentina (100 m2). It is located near to an alluvial channel called Zanjón Frías. The soils were relatively homogenous with slight inclinations, varying from silty to clayey. Four radiocarbon samples of charcoal were reported at 370 ± 50 BP (LP2645) at 200 cm depth, 1230 ± 60 BP (LP2644) at 400 cm depth (Chiavazza 2015), 1910 ± 80 (LP3180) at 450 cm depth and 2100 ± 80 years BP (LP3195) at 460 cm depth (Prieto-Olavarría et al. 2019). Combustion structures, hearths and pit-houses were detected. Small and medium-sized birds, small fishes, reptiles, armadillos and camelids (Chiavazza 2015; López 2018) composed the zooarchaeological record. Small mammal remains were only identified in pre-hispanic contexts (ca. 1200–2100 BP).
PA13.1 (32° 23′ S, 68° 14′ W; ca. 565 m a.s.l.) corresponds to a surface excavation (0–10 cm; 16 m2) on sandy soils (sand dune). The ceramic material recovered provides a relative chronology (1600–200 BP), whereas a carbon sample from an adjacent excavation in the same site resulted in 1330 ± 70 BP (URU 0281) (Chiavazza 2014). Armadillos and small fishes (≤ 1 kg) stand out among the fauna detected (Chiavazza 2007).
Materials and methods
The small mammal bone and tooth accumulation recovered from each site was analysed as a pool. This decision was made considering both the temporal range of the sites (last two millennia at most) and the need to obtain reliable taphonomic results taking into account sample sizes.
A total of 251 small mammal bone and tooth remains was recovered in MB (all detected in pre-hispanic contexts), of which 146 corresponded to cranial remains. A total of 464 remains was recovered in SF-PNO, of which 73 were cranial remains. Lastly, 253 remains were recovered in PA13.1, of which 72 corresponded to cranial elements.
Bone and tooth remains were examined using a stereomicroscope (× 8–× 50). Taxonomic identifications were made to the highest possible resolution level (species level, when possible), based on cranial and tooth remains by comparison with current specimens from the Vertebrate Collection of IADIZA (CCT CONICET Mendoza) and with specialized literature (Fernández et al. 2011a, 2017b; Pardiñas et al. 2013; Patton et al. 2015). Some rodent species are indistinguishable from cranial or dental remains allowing identifications at genus level only, such as Ctenomys and Eligmodontia. The number of identified specimens per taxon (NISP), minimal number of individuals (MNI) and minimal number of elements (MNE) were calculated (Lyman 2008). MNI was calculated on the basis of the best represented homologous cranial element.
A taphonomic analysis was conducted to distinguish patterns generated by human and non-human predators on small mammal bone and tooth accumulations. Generally, each type of predator generates taphonomic patterns that allow to distinguish the action among types of predators, at least (e.g. owls, eagles, carnivores, humans). The taphonomic methods proposed by Andrews (1990), Fernández-Jalvo and Andrews (1992), Fernández-Jalvo et al. (2016) and Fernández et al. (2017b) were followed to evaluate the action of natural predators on small mammal bone and tooth accumulations. This methodology is based on three main variables: relative abundance of skeletal elements, breakage patterns and digestion. Each predator produces enough taphonomic signals and trends to be recognized and distinguished (Andrews 1990). Relative abundances of skeletal elements were calculated considering the representation of each element in the whole sample (MNEi) based on the expected number of each element in one individual (Ei) and on MNI by using the following formula: MNEi / (Ei x MNI) × 100 (Dodson and Wexlar 1979). In order to evaluate relationships between cranial and postcranial elements, two indexes were calculated: postcrania/crania (pc/c) and femora+humeri/mandibles+maxillae (Andrews 1990). Values lower than 1 indicate better representation of cranial elements, whereas values higher than 1 show a better representation of postcranial remains (Andrews 1990). To evaluate relationships between the representation of distal and proximal parts of the skeleton, the following index was used: tibiae+radii or ulnae/femora+humeri (Andrews 1990). Values lower than 1 mean a better representation of proximal elements, whereas results higher than 1 indicate a better representation of distal elements. Two calculations were used to measure tooth loss: empty mandibular and maxillary alveoli/molars and empty mandibular and premaxillary alveoli/incisors (Andrews 1990). Values higher than 1 indicate loss of molars and incisors, whereas results lower than 1 reflect loss of mandibles and maxillae/premaxillae. Owls, which generally swallow their prey whole, generate little bone loss and a more equitable representation between skeletal parts than other predators, like diurnal birds of prey, mammalian carnivores and humans (Andrews 1990).
Breakage patterns on cranial (skulls, mandibles, teeth) and postcranial bones (femora, humeri, tibiae, ulnae) were analysed considering the degree of completeness of each element, in which the percentage of breakage was also calculated considering complete and fractured elements separately. Four categories were used for skull breakage: complete, maxillary with zygomatic, maxillary without zygomatic and minor fragments. Four degrees were used for mandible breakage: complete, ascendant ramus broken, without ascendant ramus and without ascendant ramus and inferior edge broken. Breakage of diagnostic long bones (femora, humeri, tibiae and ulnae) was recorded as complete, proximal, shaft and distal parts (Andrews 1990).
Modifications by digestion were evaluated on molars, incisors, femora and humeri distinguishing four degrees of digestive corrosion (light, moderate, heavy and extreme). Digestion is one of the most reliable taphonomic variables to identify accumulating agents of small mammal bone and tooth remains from archaeological and palaeontological sites. In fact, its clear taphonomic signals can be differentiated from other taphonomic alterations, and its traces are not modified by post-depositional processes (Andrews 1990; Fernández-Jalvo et al. 2014; Fernández et al. 2017b). Recently, Fernández et al. (2017b) developed a new methodology following Andrew’s book (Andrews 1990), but applied to South American small mammals, which generally have different tooth morphologies from those reported by that pioneer book and, indeed, different ways in which teeth are modified by digestion (Fernández et al. 2017b).
In order to strengthen the interpretation of possible accumulating agents, predictability and size indexes were calculated (Pardiñas 1999). The first was calculated to evaluate relationships between proportions of grouped and non-grouped small mammals, using the following formula: (MNI colonials + MNI gregarious + MNI grouped) / (MNI cryptic + MNI solitaries). Values higher than 1 could indicate accumulations generated by humans, carnivores or diurnal raptors, whereas values lower than 1 could be a signal of the action of nocturnal avian raptors. The second index allows the distinction between small mammal accumulations generated by nocturnal raptors (mainly strigiformes) and those accumulated by other predators (including humans) and was calculated through the following formula: (MNI small-sized [≤ 50 g] + MNI medium-sized [50 to 150 g] / MNI large-sized [≥ 150 g]). Values higher than 1 indicate sets generated by nocturnal raptors. Considering that humans, in contrast with the great majority of avian raptors, generally prey on large-sized small mammals, values lower than 1 could indicate anthropic exploitation, if and when, they could be connected with other taphonomic variables. In this sense, it would be more likely to detect anthropic action/exploitation in grouped/gregarious, large-sized small mammal species, which are conspicuous and predictable in landscape, rather than in owls, for instance, which generally feed on cryptic, solitary and smaller small mammals than eagles, carnivores and humans (Pardiñas 1999).
Anthropic signals were evaluated by taking into account relative abundance of skeletal elements, proportions of large-sized/gregarious or grouped small mammals and thermal alterations and cut-marks (Pardiñas 1999; Quintana 2005; Medina et al. 2012; Braun et al. 2016; Romaniuk et al. 2016). Thermal alterations in elements were evaluated with a chromatic scale (unburnt bones = white-cream; burnt bones = yellow-reddish-brown-black; calcined bones = blue-greyish) (Shipman et al. 1984; Stiner 1994) and by distinguishing between partial and completely thermo-altered bones (Medina et al. 2016; Romaniuk et al. 2016). The last distinction was also made in order to differentiate between natural/human-indirect fires (generally burnt or charred and calcined bones on the whole surface, respectively) (Rhodes et al. 2016) and anthropic burning patterns from exposing small mammals directly to fire (partially/heterogeneously/differentially burnt bones) (Medina et al. 2012). In general, cut-marks generated by humans are lineal on the bone surface and of variable width, showing linear marks with a V-shaped cross-section generally restricted to the cortical area and showing parallel striations, an internal surface with longitudinal micro-striations and shallow micro-striations transversal to the long axis (Blumenschine et al. 1996). Marks mimicking cut-marks generated by post-depositional processes, like trampling or water transport, differ from those produced by humans (mimic marks are generally less deep, shorter and much more frequent) (Fernández-Jalvo and Andrews 2016).
The most profitable, highest-ranked faunal resources for CWA pre-hispanic human groups were L. guanicoe (guanaco) and armadillos (Corbat et al. 2017). Complementarily for the present study and exploring future and possible applications from the analysis of archaeological small mammal remains—extensible to low-ranked faunal remains—, a regional high-ranked resources index was calculated (HRI = ∑NISP Artiodactyls + ratio Armadillos/∑NISP total; after Corbat 2015 and Corbat et al. 2017). This index (with values ranging from 0 to 1) allows assessing abundance and dependence of human groups on faunal resources. Decreasing values in this index have proven to be a good predictor of resource depression/diversification processes (López et al. 2019a). Regarding small mammals, only remains of large-sized/and gregarious or grouped taxa were considered in this calculation.
Post-depositional processes were also analysed, such as weathering, trampling, the action of root traces and abrasion (Andrews 1990; Fernández-Jalvo et al. 2014). To distinguish between pre- and post-depositional breakage, rough and angled broken edges (by trampling) were differentiated from breakage by predation (rounded and digested broken edges) (Andrews 1990).
In order to explore sample ordination in a multivariate space, a principal components analysis (PCA) was additionally conducted against previously reported modern and archaeological small mammal samples using the software PAST (Hammer et al. 2001). The PCA worked over a data matrix composed of digestion percentage on isolated incisors, isolated molars, incisors in situ, molars in situ, femora and humeri; percentage of breakage of skulls, mandibles, femora and humeri; calculated indexes of relative abundance and total average of relative abundance of skeletal elements.
Results
A total of 968 bone and tooth specimens was recovered and analysed from which 248 corresponded to cranial and tooth elements identifiable taxonomically (119 from MB, 58 from SF-PNO and 71 from PA13.1). A total MNI of 72 was calculated (25 from MB, 23 from SF-PNO and 24 from PA13.1). A high proportion of large-sized/gregarious small mammals was detected in MB (68%) and PA13.1 (75%), whereas in SF-PNO, it was about 20%. The Predictability Index (PI) resulted in values greater than 1 in MB and PA13.1, whereas the Size Index (SI) was lower than 1 in such sets. A contrary trend was detected for SF-PNO, where PI and SI resulted in values lower and higher than 1, respectively (Table 2).
Femora from SF-PNO and humeri from PA13.1 were the most abundant elements, with relative abundances close to 100%, followed by mandibles (MB and SF-PNO) and femora and astragali from PA13.1 with relative abundances between 60 and 70% each. Humeri, tibiae and pelvises from SF-PNO represented between 50 and 55% each, whereas a relative abundance close to 50% was calculated for tibiae recovered from MB. A low proportion (≤ 15% each) of isolated molars, ribs, vertebrae, scapulae, radii, metapodials and phalanges was detected in the three sites (Fig. 2). Average values of relative abundance for the total sample per site did not exceed 28% (Tables 1 and 3).

Comparison of relative abundance of skeletal elements. Top: Archaeological sites from Northern Mendoza (present study) and archaeological accumulations where humans generated small mammal bone accumulations (always mixed with the participation of different kind of predators). MB, Memorial de la Bandera; SF-PNO, Ruinas de San Francisco Sector Pilastra Noroeste; PA13.1, Punto Arqueológico 13.1; A1, Angostura 1 (Fernández et al. 2011b); KV502 (Dewar and Jerardino 2007). Down: Archaeological sites from northern Mendoza (present study) and small mammal bone accumulations generated by avian raptors (Montalvo and Fernández 2019)
The indexes calculated to evaluate relationships between cranial and postcranial elements indicated a better representation of postcranial elements than cranial ones in SF-PNO and PA13.1, whereas in MB, cranial elements were better represented than postcranial ones (Table 1). In relation to the index calculated to measure relationships between distal/proximal parts of the skeleton, in MB, distal elements were more abundant than proximal ones, whereas in SF-PNO and PA13.1, proximal elements were considerable better represented than distal ones. Alv/i index showed loss of mandibles and premaxillae in MB and SF-PNO, whereas loss of incisors was detected in PA13.1. Alv/m index showed an equal representation of empty alveoli and isolated molars in MB, a considerable loss of molars in SF-PNO and loss of mandibles and maxillae in PA13.1.
Breakage of cranial elements was considerably high in the three sets (≥ 95% each) (Tables 1 and 4). The proportions of broken postcranial elements were also high (≥ 76% in each set), although lower than in cranial elements. The great majority of broken bones (≥ 95 in each set) showed rough and/or angled edges, probably as a consequence of trampling.
Alterations by digestion, mainly light, were observed on diagnostic elements from the three small mammal bone and tooth accumulations (Tables 1 and 5). Long bones were the most modified elements by digestive corrosion (between 25 and 54% per set). Molars and incisors were affected in lesser proportions (≤ 22 % and ≤ 18%, respectively). Isolated teeth were generally more affected than those detected in situ, with the exception of SF-PNO, where the two isolated molars recovered not were modified by digestion. In MB, digested teeth and postcranial elements were also recovered from inside combustion structures.
Thermal alterations were detected in cranial and postcranial elements recovered from MB and PA13.1 (10.35% and 27.66%, respectively) (Fig. 3, Tables 1 and 6), whereas no thermo-altered elements were found in SF-PNO. All thermo-altered elements (n = 26) in MB were only detected from inside the combustion structures or hearths associated with pit-houses on bones corresponding to large-sized small mammals (caviomorph and cricetid rodents). On the one hand, about 60% of thermo-altered elements from this site (n = 15) presented a distinctive burning pattern (distal, differential or heterogeneous burning) on the distal extremities of long bones, on one vertebrae and on one hemimandible corresponding to the extinct cricetid rodent Holochilus lagigliai. In turn, the surface of the remaining thermo-altered elements was completely burnt or calcined. On the other hand, 70 elements from PA13.1 showed thermal alterations, all corresponding to large-sized rodents (caviomorphs). Of them, 18.6% (n = 13) was differentially, partially or heterogeneously burnt, 65.7% (n = 46) was completely burnt on the whole surface, whereas 15.7% (n = 11) was calcined. No thermo-altered elements showed evidence of digestion. Only two short and shallow oblique/transversal cut-marks were detected on the shaft of a complete caviid femur recovered from SF-PNO, showing a V-shaped cross-section restricted to the cortical area and parallel striations (Fig. 3). Digested elements were also recovered mixed in the same layer where such femur was recovered.

Taphonomic modifications on small mammal bone and tooth accumulations recovered in archaeological sites from northern Mendoza: Taphonomic signals possibly consequence of anthropic activity: a–g Burning on the distal extremities. a–d Burnt bones from PA 13.1 (a, humerus; b, tibia; c, astragalus; d, tibia—caviomorph rodents—); e–g Burnt bones from Memorial de la Bandera (e and g Hemimandible of Holochilus lagigliai; f humerus); h Cut-marks possibly made by humans on a femur shaft. Taphonomic signals of digestion generated by owls. i Light digestion on cricetid femur; j Light digestion on cricetid humerus
PCA was performed considering the taphonomic values of archaeological samples from the present study, archaeological small mammal accumulations probably generated (partial or completely) by humans and modern small mammal bone accumulations generated by several avian raptors, where components 1 and 2 accounted for 85.19% of inertia (Fig. 4; Electronic supplementary material S1 and S2). The Regional High-Ranked Resources Index yielded low values: 0.05 for MB, 0.068 for SF-PNO and 0.32 for PA13.1, indicating low dependence on high-ranked resources (Fig. 5).

Principal component analysis (based on taphonomic variables) of small mammal bone accumulations recovered in three archaeological sites from northern Mendoza (MB, Memorial de la Bandera; SF-PNO, Ruinas de San Francisco sector Pilastra Noroeste; PA13.1, Punto Arqueológico 13.1) and comparison both with small mammal accumulations from archaeological sites probably accumulated by human beings and with modern small mammal bone samples generated by several raptors. Archaeological sites: A1 (Angostura 1; Fernández et al. 2011a) and KV502 (Dewar and Jerardino 2007). Raptors: Athene cunicularia (Montalvo et al. 2020); Tyto furcata 1(Gómez 2007); Tyto furcata 2 (Iglesias 2009); Puma concolor (Montalvo et al. 2007); Wildcats (López et al. 2017a); Bubo magellanicus (Montalvo et al. 2016a); Geranoaetus melanoleucus and Geranoaetus polyosoma (López et al. 2017b); Strix chacoensis (López et al. 2018)

High-ranked resources index (HRI = Σ NISP artiodactyls + armadillos/ Σ NISP total; after Corbat 2015; Corbat et al. 2017; López et al. 2020) for zooarchaeological contexts from northern Mendoza, based on averaged and calibrated ages (after López et al. 2019). The figure includes the new results from Punto Arqueológico 13.1 (PA13.1), Memorial de la Bandera (MB) and Ruinas de San Francisco sector Pilastra Noroeste (SF-PNO), including in the calculation NISP of large-sized small mammals from these sites
Weathering was recorded in 29.1% (n = 73) of elements from MB, in 16.6% (n = 77) of elements from SF-PNO and in 80% (n = 201) of elements recovered from PA13.1 (Table 1). In MB and SF-PNO, only category 1 weathering was observed (slight cracking), whereas in PA13.1, most of the weathered bones presented slight cracking (n = 191), although category 2 weathering (more extensive cracking) was detected in 10 elements. Root traces were detected infrequently (≤ 5.9% per sample, at most). Traces of abrasion by water transport were not detected.
Discussion
The results obtained from the taphonomic analysis of small mammal bone and tooth remains recovered in MB, SF-PNO and PA13.1 archaeological sites suggest that more than one type of accumulating agent generated each accumulation. The small sample sizes have probably influenced the results obtained, but the taphonomic signals assignable to the action of owls and humans could be distinguished in each accumulation. López and Chiavazza (in press) detected a considerable effect of sample size on the species richness observed in these three accumulations when discussing the palaeoecological implications of these assemblages.
The low average values of relative abundance of skeletal elements and the high proportions of broken bones with rough and angled edges suggest the action of destructive post-depositional processes, like trampling, and consequent bone loss in small mammal accumulations (Andrews 1990). The presence of complete burnt bones may have biased the original taphonomic features of each bone set, modifying breakage patterns and relative abundances, mainly (Fig. 2).
Weathering was considerably high in PA13.1 which, added to the effects of trampling, reflect a low preservation of bones and suggest a bone assemblage probably never incorporated into the sedimentary matrix of the site, as is the case of subsurface archaeological sites in the sandy soils of plains from the Monte Desert in southern South America (Fernández 2012; López and Chiavazza 2019). The lowest proportions of weathered and broken elements in MB and SF-PNO indicate a better preservation and faster incorporation into the sedimentary matrix of such sites.
Owls as accumulating agents
Small mammals (≤ 1 kg) (rodents, marsupials and bats) represent the most important component of owls diet from southern South America, but rodents are particularly the most abundant prey of these raptors (e.g. Bellocq 2000; Nabte et al. 2006; Montalvo et al. 2020). In fact, rodents were the most common small mammal remains found in the present study.
The low proportion of diagnostic elements affected by digestion and the main digestion degree detected (light) suggest the action of avian raptors, particularly owls, in the accumulation of small mammal remains. Generally, owls swallow their prey whole, generating low bone breakage in the regurgitated pellets. They also generate a relative equal representation among skeletal parts, with loss of elements from the autopodium, and mainly, a low proportion and degree of digestive corrosion on bones and teeth (Andrews 1990). Despite the fact that post-depositional processes could have biased some taphonomic signals generated by predators, both the degree and proportion of digested elements suggest the action of an owl more destructive than the barn owl (Tyto furcata, López 2020; Tyto alba, Andrews 1990), such as the Great horned owl Bubo virginianus/Bubo magellanicusor the burrowing owl Athene cunicularia. These raptors generate higher frequencies of elements modified by digestion, more bone loss and more disproportions between elements and skeletal parts than the barn owl, albeit always within a general taphonomic owl pattern (modification categories 2–3 at most, sensu Andrews 1990) (Gómez 2005; Montalvo et al. 2016a, 2020; Montalvo and Fernández 2019). Avian raptors, and specifically owls, are the most common accumulating agents of small mammal remains identified in archaeological sites from CWA (Fernández et al. 2009, 2015a, 2015b; Fernández 2012; López et al. 2016; Fernández and Pardiñas 2018; López and Chiavazza 2019, 2020).
Humans as accumulating agents
Both the detection of thermal alterations in MB and PA13.1 and the presence of cut-marks on a caviid femur shaft from SF-PNO might suggest anthropic activity in the generation of the small mammal bone and tooth assemblages of these three sets. PI and SI indexes indicated a greater representation of the grouped/colonial/gregarious and the largest species in MB and PA13.1 than the smallest and cryptic/solitary ones, whereas the latter were more abundant only in SF-PNO. The high representation of grouped/colonial/gregarious and large-sized small mammals could also be identified among the signals of anthropic activity.
The presence of partial, differential or heterogeneously burnt bones in MB and PA13.1 (Fig. 3, Table 6)—mainly burnt on their extremities or distal parts—can be associated with human activity and, particularly, with a cooking method that consists of exposing these small prey directly on fire (Medina et al. 2012; Fernández et al. 2017a). High temperature produces a retraction of the muscles and meat of the axial and appendicular distal portions, which can be directly exposed to fire, burning the exposed unmeaty portions of the bones (e.g. Pardiñas 1999; Medina et al. 2012). In this sense, small mammals could have been burnt or cooked while they were still in a partial or complete articulate state with some of their tissues intact (sensu Romaniuk et al. 2016). The high representation of the largest small mammals (≥ 150 g) in MB and PA13.1 does not suggest the action of nocturnal avian raptors (Strigiformes), which generally feed on smaller prey (≤ 150 g) (e.g. Andrews 1990; Bellocq 2000; Montalvo et al. 2016a; López et al. 2018). The detection of completely burnt bones could also be due to natural/post-depositional fires (e.g. Rhodes et al. 2016).
The detection of cut-marks only on the shaft of a complete caviid femur (large-sized rodent, ≥150 g) recovered from SF-PNO suggests that this accumulation was partially generated by humans. First, it is not common to find cut-marks on archaeological small mammal bones accumulated by humans (Fernández et al. 2017a). The detection of cut-marks in this type of assemblage is scarce because pre-cooking processing does not require difficult tasks or the intensive use of cutting tools (Fernández-Jalvo et al. 1999; Fernández et al. 2017a; Martínez-Polanco 2017). The scarcity of this type of evidence in SF-PNO, the greater abundance of cryptic/solitary/smallest species, the absence of thermo-altered bones and the presence of a considerable proportion of bones with evidence of light digestion (probably generated by owls) could support the idea that Quintana (2005) defined as isolated butchery activities. This interpretation is related with the scarce representativeness of all the body parts in relation to marks and traces connected with anthropic processing of small mammal bones. Second, the absence of burnt bones in SF-PNO could also be associated with another cooking technique. Thus, zooarchaeological analysis of the largest fauna (≥ 1 kg)—which presented a low proportion of thermo-altered bones—, added to the analysis of pottery from SF-PNO, has suggested that in SF-PNO food could have been boiled (Castillo et al. 2018b). The absence of burnt bones in archaeological small vertebrate assemblages could be due to the fact that, under certain circumstances, the defleshed meat was cooked separately from the bone, the muscle mass was eaten uncooked-undercooked or that the elements were cooked, but the fire did not affect the bone (e.g. Blasco and Fernández Peris 2009; Medina et al. 2012). Considering the frequency and intensity of affected elements and the taphonomic signals detected, the human contribution is probably quite small compared with elements accumulated by avian raptors in the three sites but, particularly, in SF-PNO.
So far, the detection of anthropic signals on small mammal accumulations from archaeological sites in CWA has been scarce, in one site from the southern transitional region with North Patagonia, corresponding to isolated and weak taphonomic signals of anthropic activity (Fernández et al. 2009, 2017a; Fernández 2012; Fig. 1, sites 17, 19 and 23). These studies interpreted the incorporation of small mammals to the human diet as casual. Only in one archaeological site from an intermountain valley in the highlands of northern Mendoza (100 km west of the present study area) did the considerable number of thermo-altered elements of large-sized small mammals (many of them presenting a partial, differential or heterogeneous burning pattern) suggest that humans could have been partially responsible for such small mammal accumulation (López et al. 2020; Fig. 1, site 4). Simonetti and Cornejo (1991) reported frequent human exploitation on largest-sized (≥ 150 g) and colonial small mammals due to the detection of burnt bones belonging to caviomorph rodents in two late Holocene sites from the Andean Cordillera in central Chile. Based on the zooarchaeological information available in CWA, López et al. (2019a) interpreted that the exploitation of different faunal resources after ca. 2000 BP in northern Mendoza could reflect a process of resource diversification as a consequence of high-ranked resource depression. A recent study identified a broader diet after ca. 2000 BP as ever before for prehistoric human groups from northern Mendoza, associating it with an intensification process probably due to a declining foraging efficiency as a consequence of an increase in human demography and occupation of new and marginal areas of piedmont and plains from CWA (López et al. 2019a). Low-ranked resources identified after that date are generally small mammals and fishes detected without specific technology suggesting mass capture (Chiavazza 2013; Corbat et al. 2017; López et al. 2019a). The results obtained in the present study contribute to supporting this intensification hypothesis.
The low HRI values calculated for the zooarchaeological accumulations of the present study reflect a low dependence on high-ranked resources (Fig. 5; López et al. 2019a; López et al. 2020). However, in the remaining zooarchaeological studies conducted in CWA, in the great majority of sites (see Fig. 1), other non-human predators—mainly owls—were the accumulating agents of small mammal bone and tooth remains ( Fernández 2012; López et al. 2016; Castillo et al. 2018a; Fernández and Pardiñas 2018; López 2018; López and Chiavazza 2019).
The prevalence of the largest-sized small mammals (≥ 150 g) in PA13.1—a subsurface site on sandy soils strongly affected by trampling and weathering—could be the consequence of differential preservation and should not be attributable to the dietary preferences of certain predators. Thus, small mammal bones corresponding to larger species could be better preserved than bones from smaller species due to their differences in size and density (Denys et al. 1997). In this site, however, burnt bones with a pattern assignable to human consumption were also detected.
The higher availability of patch resources evidenced in the past by a greater environmental heterogeneity than at present—inferred from ethnohistorical research and archaeological small mammal assemblages (e.g. Prieto and Chiavazza 2006; Prieto and Rojas 2012; López and Chiavazza in press)—could have influenced the broader human diet detected after ca. 2000 BP, hinting at an ecological rather than cultural pattern. However, based on small mammal assemblages as ecological indicators, these environmental heterogeneity conditions would have been stable in Central Western Argentina, at least, during the last 4000 years (Fernández 2012; Fernández et al. 2015a, b, 2017c; López et al. 2016; Castillo et al. 2018a; López 2018; López and Chiavazza 2019). Since the occupation of CWA by human groups (ca. 11,000 BP), the human diet always depended almost completely on high-ranked resources, mainly wild camelids—L. guanicoe—(López et al. 2019a). Only after ca. 2000 BP is a diet breath expansion detected as well as a decrease in high-ranked resource dependence. For that reason, the changes detected in the human diet after ca. 2000 BP could be due to cultural factors.
The detection of more than one accumulating agent in the small mammal bone sets (humans and owls) probably precludes a clear discrimination of the specific accumulating agent of each bone set. However, various types of predators are often responsible for the small mammal bone accumulations recovered in archaeological sites from different parts of the world, generating mixed assemblages both by different kinds of natural predators and by human and non-human predators (e.g. Matthews 2006; Fernández et al. 2011b, 2017a, 2019; López et al. 2017a).
A complementary PCA yielded values grouping the archaeological samples of MB, SF-PNO and PA13.1 together and with two archaeological samples where humans were accumulating agents, separating them from samples generated by Strigiformes and Accipitriformes (Fig. 4; Fig. S1 and Table S2). However, as noticed, the PCA usually isolates modern from archaeological or palaeontological samples due to the influence of post-depositional effects caused by trampling, weathering and burning. The fact that the modern samples appear grouped and separated from archaeological samples does not necessarily mean that humans are involved in the generation of the latter, but that post-depositional processes and time averaging may be acting as well. For example, the former may affect the archaeological record increasing element loss and breakage and the mixture of taphonomic signals (Andrews 1990), while the latter may influence the increase of species richness and the representation of each species (Terry 2010), among other factors.
Conclusions
The taphonomic analysis of small mammal bone and tooth accumulations recovered in three archaeological sites from central northern Mendoza with chronologies after ca. 2000 BP indicates that humans and avian raptors were probably the accumulating agents responsible for such bone sets. Traces of digestion, mainly light, were observed on teeth and postcranial elements which, added to the relative abundance of skeletal elements and breakage patterns, suggest the action of owls. A prejudiced taphonomic view only addressed to distinguishing non-human predators of archaeological small mammal accumulations can bias our interpretations and analysis. Indeed, taphonomic traces probably assignable to human activity, like thermal alterations (MB and PA13.1), cut-marks (SF-PNO) and/or higher proportions of gregarious/grouped and large-sized small mammals (MB and PA13.1) were detected in the three archaeological small mammal bone sets from northern Mendoza analysed in the present study. Humans would have exploited small mammals as a consequence of high-ranked resource depression. Archaeological small mammal accumulations from northern Mendoza that present taphonomic traces assignable to human predatory activity have only been detected in Monte Desert sites. From a biogeographic point of view, this is interesting for future archaeological and palaeoecological discussions.
The mixture of predators could account for the originality detected in the taxonomic structure of archaeological small mammal assemblages, considering the differences in the dietary preferences of owls and humans. Therefore, the participation of more than one accumulating agent with different habits and feeding behaviours could have influenced the abundance and presence of certain species and taphonomic patterns.
The present study aims to develop new and bigger regional research of small mammal accumulations recovered in archaeological sites from a zooarchaeological, taphonomic and palaeoenvironmental perspective to strengthen our understanding of human and environmental systems through space and time. At the same time, further zooarchaeological studies based on complete faunal assemblages from northern Mendoza are required to strengthen our understanding of changes in human subsistence of regional pre-hispanic societies.
Data availability
Material is stored in the Laboratorio de Arqueología Histórica y Etnohistoria (Facultad de Filosofía y Letras, Universidad Nacional de Cuyo, Argentina) and in the Centro de Investigaciones Ruinas de San Francisco (Área Fundacional de Mendoza, Argentina). Data available on request from the authors.
References
Abraham EM, Del Valle HF, Roig F, Torres L, Ares JO, Coronato F, Godagnone YR (2009) Overview of the geography of the Monte Desert biome (Argentina). J Arid Environ 73:144–153
Abraham EM, Rodríguez MD, Rubio MC, Guida-Johnson B, Gómez L, Rubio C (2020) Disentangling the concept of “South American Arid Diagonal”. J Arid Environ 175:104089
Alcaráz AP (2015) Aprovechamiento antrópico de fauna menor en el curso inferior del río Colorado (transición pampeano-patagónica oriental): el sitio El Tigre como caso de estudio. Intersecciones Antro 16:115–129
Andrade A (2015) Distinguishing between cultural and natural depositional agents: micromammal taphonomy from the archaeological site Cueva y Paredón Loncomán (Patagonia, Argentina). J Archaeol Sci Rep 3:122–131
Andrade A, Fernández PM (2017) Rodent consumption by hunter-gatherers in north Patagonian Andean forests (Argentina): insights from the small vertebrate taphonomic analysis of two late Holocene archaeological sites. J Archaeol Sci Rep 11:390–399
Andrews P (1990) Owls, caves and fossils. Predation, preservation and accumulation of small mammal bones in caves, with an analysis of the Pleistocene Cave Faunas from Westbury-Sub-Mendip, Somerset, U.K. University of Chicago Press, Chicago
Behrensmeyer AK, Kidwell SM, Gastaldo RA (2000) Taphonomy and paleobiology. Paleobiology 26:103–147
Bellocq MI (2000) A review of the trophic ecology of the Barn Owl in Argentina. J Raptor Res 34:108–119
Blasco R, Fernández Peris J (2009) Middle Pleistocene bird consumption at Level XI of Bolomor Cave (Valencia, Spain). J Archaeol Sci 36:2213–2223
Blumenschine RJ, Marean CW, Capaldo SD (1996) Blind tests of inter-analyst correspondence and accuracy in the identification of cut marks, percussion marks and carnivore tooth marks on bone surfaces. J Archaeol Sci 23:493–507
Braun DR, Pante M, Archer W (2016) Cut marks on bone surfaces: influences on variation in the form of traces of ancient behaviour. Interface Focus 6:20160006
Broughton J (1994) Late Holocene resource intensification in the Sacramento Valley: the vertebrate evidence. J Archaeol Sci 21:501–514
Broughton J, Cannon M, Bayham F, Byers D (2011) Prey body size and ranking in zooarchaeology: theory, empirical evidence and applications from the Northern Great Basin. Am Antiq 76:403–428
Castillo L, López JM, Chiavazza H (2018a) El registro de micromamíferos en el Arroyo Tulumaya: punto arqueológico 70 (noreste de Mendoza). Aspectos tafonómicos e implicancias ambientales para los últimos 500 años. Revista de Arqueología Histórica Argentina y Latinoamericana 12:454–478
Castillo L, Araujo E, Chiavazza H, Prieto Olavarría C (2018b) Cocinar y alimentarse en tiempos de conquista. Reconstruyendo paquetes culinarios a partir de análisis cerámicos y arqueofaunísticos (Mendoza, siglos XV-XVII). Arqueología 24:109–132
Chiavazza H (2007) Cambios ambientales y sistemas de asentamiento en el árido Normendocino. Arqueología en los paleocauces del Río Mendoza. Unpublished PhD thesis. Facultad de Ciencias Naturales y Museo. Universidad Nacional de La Plata. Argentina
Chiavazza H (2011) Procesos sociales y ambientales en el sector urbano de Mendoza entre los siglos XV-XVIII: arqueología urbana e historia ambiental. Comechingonia virtual 4:227–253
Chiavazza H (2013) “No tan simples”: pesca y horticultura entre grupos originarios del norte de Mendoza. Comechingonia Virtual 7:27–45
Chiavazza H (2014) Tendencias regionales del registro arqueológico de sociedades sin estructura aldeana del noreste de Mendoza (Argentina). In: Falabella, FG, Sanhueza LR, Cornejo LB, Correa I (eds), Distribución espacial en sociedades no aldeanas: del registro arqueológico a la interpretación social. Serie Monográfica de la Sociedad Chilena de Antropología 4. Ediciones on demand. Chile. pp, 221–249
Chiavazza H (2015) Pescadores y horticultores ceramistas del valle de Mendoza. In: Bárcena JR (ed.), Arqueología y Etnohistoria del Centro Oeste Argentino. Xama Serie Monografías 5. Zeta. Mendoza, pp. 45–62
Chiavazza H, Prieto Olavarría C, Zorrilla V (2010). Arqueología de una formación urbana. Los registros en una ciudad de desarrollo continuo: Mendoza en el siglo XVI. In: Chiavazza H, Cerutti C (eds.), Arqueología de ciudades Americanas del siglo XVI. Facultad de Filosofía y Letras, Universidad Nacional de Cuyo. Mendoza, pp. 317-342
Corbat M, Zangrando A, Gil AF, Chiavazza H (2017) Explotación de peces e intensificación en ambientes áridos: comparando el registro en humedales del centro-occidento de Argentina. Lat Am Antiq 28:196–212
Cortegoso V (2006) Comunidades agrícolas en el Valle de Potrerillos (NO de Mendoza) durante el Holoceno tardío: organización de la tecnología y la vivienda. Intersecciones Antro 7:77–94
Denys C, Dauphine Y, Fernández-Jalvo Y (1997) Apports biostratigraphiques et paléoécologiques de l’étude taphonomique des assemblages de micromammiféres. Geobios 20:197–206
Dewar G, Jerardino A (2007) Micromammals: when humans are the hunters. Journal of Taphonomy 5:1–14
Dodson P, Wexlar D (1979) Taphonomic investigations of owl pellets. Paleobiology 5:275–284
Efremov JA (1940) Taphonomy: new branch of paleontology. Pan-Am Geol 74:81–93
Fernández FJ (2012) Microvertebrados del Holoceno de sitios arqueológicos en el sur de Mendoza (República Argentina): aspectos tafonómicos y sus implicancias en la subsistencia humana. PhD thesis. Universidad Nacional La Plata. 520 pp.
Fernández FJ, Pardiñas UFJ (2018) Small mammals taphonomy and environmental evolution during Late Pleistocene-Holocene in Monte Desert: the evidence of Gruta del Indio (central west Argentina). J S Am Earth Sci 84:266–275
Fernández FJ, Moreira G, Neme G, De Santis LJM (2009) Microvertebrados exhumados del sitio arqueológico “Cueva Arroyo Colorado” (Mendoza, Argentina): aspectos tafonómicos y significación paleoambiental. Archaeofauna 18:99–118
Fernández FJ, Ballejo F, Moreira G, Tonni E, De Santis LJM (2011a) Roedores cricétidos de la provincia de Mendoza. Guía cráneo-dentaria orientada para su aplicación en estudios zooarqueológicos. Sociedad Argentina de Antropología-Universitas Sarmiento. Buenos Aires. 115 pp.
Fernández FJ, del Papa LM, Moreira G, Prates L, De Santis LJM (2011b) Small mammal remains recovered from two archaeological sites in the middle and lower Negro River valley (Late Holocene, Argentina): Taphonomic issues and paleoenvironmental implications. Quat Int 245:136–147
Fernández FJ, Neme G, De Santis LJM (2015a) Micromammals, Taphonomy, paleoenvironments and human occupation during the Holocene in Arroyo Malo-3 (west-central Argentina). Archaeofauna 24:27–52
Fernández FJ, Rudzik SM, Neme G, De Santis LJM (2015b) Micromamíferos, tafonomía y ambientes durante los últimos 3.800 años A.P. en el Valle Medio del Río Grande, Mendoza. Argentina. Chungara 47:267–285
Fernández FJ, Teta P, Pardiñas UFJ (2017a) Evidencias arqueológicas de explotación antrópica de micromamíferos en el extremo austral de América del Sur. Anales de Arqueología y Etnología 72:9–32
Fernández FJ, Montalvo CI, Fernández-Jalvo Y, Andrews P, López JM (2017b) A re-evaluation of the taphonomic methodology for the study of small mammal fossil assemblages of South America. Quat Sci Rev 155:37–49
Fernández FJ, Torres J, Tammone M, López JM, Pardiñas UFJ (2017c) New data on the endemic cricetid rodent Holochilus lagigliai from central western Argentina: fossil record and potential distribution. Mammalia 81:621–625
Fernández FJ, Hadler P, Cherem JJ, Saldanha JP, Stutz NS, Dias AS, Pardiñas UFJ (2019) Holocene small mammals hunted by owls and humans in southern Brazil: taphonomic evidence and biological significance. Boreas 48:953–965
Fernández-García M, López-García JM, Bennàsar M, Gabucio MJ, Bargalló A, Chacón MG, Saladié P, Vallverdú J, Vaquero M, Carbonell E (2018) Paleoenvironmental context of Neanderthal occupations in northeastern Iberia: the small-mammal assemblage from Abric Romaní (Capellades, Barcelona, Spain). Palaeogeogr Palaeocl 506:154–167
Fernández-García M, López-García JM, Royer A, Lécuyer C, Allué E, Burjachs F, Chacón MG, Saladié P, Vallverdú J, Carbonell E (2020) Combined palaeoecological methods using small-mammal assemblages to decipher environmental context of a long-term Neanderthal settlement in northeastern Iberia. Quat Sci Rev 228:106072
Fernández-Jalvo Y, Andrews P (1992) Small mammal taphonomy of Gran Dolina, Atapuerca (Burgos), Spain. J Archaeol Sci 19:407–428
Fernández-Jalvo Y, Andrews P (2016) Atlas of taphonomic identifications. Springer, Dodrecht
Fernández-Jalvo Y, Andrews P, Denys C (1999) Cut marks on small mammals at Olduvai Gorge Bed-1. J Hum Evol 36:587–589
Fernández-Jalvo Y, Andrews P, Sevilla P, Requejo V (2014) Digestion versus abrasion features in rodent bones. Lethaia 47:323–336
Fernandez-Jalvo Y, Andrews P, Denys C, Sese C, Stoetzel E, Marin-Monfort D, Pesquero D (2016) Taphonomy for taxonomists, implications in small mammal studies. Quat Sci Rev 139:138–157
Frontini R, Escosteguy P (2015) El rol de los pequeños animales en los estudios arqueofaunísticos de Argentina. Archaeofauna 24:67–85
Gasco A, Rosi MI, Durán V (2006) Análisis arqueofaunístico de microvertebrados en «Caverna de las Brujas» (Malargüe-Mendoza-Argentina). Anales de Arqueología y Etnología, Special Volume 61:135–162
Gifford-González D (2018) An introduction to Zooarchaeology. Springer International Publishing, New York
Gil AF, Giardina M, Neme G, Ugan A (2014) Demografía humana e incorporación del cultígenos en el centro occidente argentino: explorando tendencias en las fechas radiocarbónicas. Revista Española de Antropología Americana 44:523–553
Gómez GN (2005) Analysis of bone modifications of Bubo virginianus pellets from Argentina. Journal of Taphonomy 3:1–16
Gómez GN (2007) Predators categorizations based on taphonomic analysis of micromammals bones: a comparison to proposed models. In: Gutiérrez MA, Miotti L, Barrientos G, Mengoni Goñalons G, Salemme M (eds) Taphonomy and Zooarqueology in Argentina. BAR International Series, Oxford, pp 89–103
Hammer Ø, Harper D, Ryan P (2001) PAST: Paleontological Statistics software package for education and data analysis. Paleontología Electrónica 4:1–9
Iglesias AC (2009) Tafonomía de pequeños vertebrados depredados por Buteo polyosoma (Aves, Falconiformes). Unpublished Licentiate thesis, Facultad de Ciencias Exactas y Naturales, Universidad Nacional de La Pampa
López JM (2018) Restos óseos de micromamíferos recuperados en sitios arqueológicos del norte de Mendoza para contextos correspondientes al Holoceno tardío: un enfoque zooarqueológico, tafonómico y paleoambiental. Unpublished PhD thesis. Facultad de Filosofía y Letras. Universidad Nacional de Cuyo
López JM (2020) Actualistic taphonomy of barn owl pellet‐derived small mammal bone accumulations in arid environments of South America. J Quat Sci 35:1057–1069. https://doi.org/10.1002/jqs.3251
López JM, Chiavazza H (2019) Amidst wind, sand and raptors. Small mammal bone remains recovered in open-air archaeological sites from the Monte Desert in Central Western Argentina: taphonomic and palaeoenvironmental implications. Archaeol Anthropol Sci 11:5149–5169
López JM, Chiavazza H (2020) Micromamíferos recuperados en sitios arqueológicos de precordillera en los andes del centro occidente argentino (Holoceno tardío): aspectos tafonómicos y paleoambientales. Lat Am Antiq 31:409–429. https://doi.org/10.1017/laq.2020.13
López JM, Chiavazza H, Rosi MI (2016) Small mammal remains recovered from two archaeological sites in northwestern Mendoza (late Holocene, Argentina): taxonomic composition, taphonomic issues and paleoenvironmental implications. Quat Int 391:26–37
López JM, Rosi MI, Tabeni S, Bender B, Chiavazza H (2017a) Taphonomic analysis of small mammal bone remains preyed upon by wildcats (Carnivora: Felidae) from the central Monte Desert. Boreas 46:282–293
López JM, Fernández FJ, Montalvo CI, Chiavazza H, De Santis LJM (2017b) The role of the Accipitriformes Geranoaetus melanoleucus and Geranoaetus polyosoma as small mammal bones accumulators in modern and archaeological sites from Central Western Argentina. Journal of Taphonomy 15(1–3):91–108
López JM, Fernández FJ, Castillo L, Pereyra-Lobos R (2018) Taphonomy of small mammal bone accumulations generated by the Chaco owl (Strix chacoensis, Strigiformes) from the Monte Desert (Mendoza, Argentina): a contribution to South American archaeological and palaeontological studies. Boreas 47:780–791
López JM, Neme G, Gil AF (2019a) Resource intensification and zooarchaeological record in the southern margins of pre-hispanic Andean Agriculture. Archaeol Antrhopol Sci 11:5287–5300
López JM, Fernández FJ, Giardina M (2019b) In the stomach of Geranoaetus melanoleucus: Taphonomy of small mammal bone remains and implications for South American archaeological and palaeontological studies. J Archaeol Sci Rep 26:101932
López JM, Fernández FJ, Teta P, Rosi MI, Sironi O, Bárcena JR (2020) Zooarchaeology, taphonomy and palaeoenvironments: small mammal remains from an intermountain valley of the southern Andean cordillera (Mendoza, Argentina). J Archaeol Sci Rep 31:102365
López-García JM, Livraghi A, Romandini M, Peresani M (2018) The De Nadale cave (Zovencedo, Berici Hills, northeastern Italy): a small-mammal fauna from near the onset of marine isotope stage 4 and its palaeoclimatic implications. Palaeogeogr Palaeocl 506:196–201
Lyman RL (2008) Quantitative paleozoology. Cambridge University Press, Cambridge
Mafferra LE (2017) Los paisajes forestales en torno a la ciudad colonial de Mendoza, con base en el registro antracológico. Intersecciones Antro 18:43–53
Mafferra LE (2018) Arqueología de los paisajes forestales del norte de Mendoza, centro-oeste Argentina (siglos VIII-XIX). British Archaeological Reports BAR International series, Oxford
Martínez-Polanco MF (2017) El Cuy (Cavia sp.), un recurso alimenticio clave en Aguazuque, un sitio arqueológico de la sabana de Bogotá, Colombia. Lat Am Antiq 27:512–526
Matthews T (2006) Taphonomic characteristics of micromammals predated by small mammalian carnivores in South Africa: application to fossil accumulations. Journal of Taphonomy 4:143–161
Medina M, Teta P, Rivero D (2012) Burning damage and small-mammal human consumption in Quebrada del Real 1 (Cordoba, Argentina): an experimental approach. J Archaeol Sci 39:737–743
Mignino J, Izeta A, Cattáneo R (2018) Modern and archaeological owl pellets as paleoenvironmental and taphonomic markers in human occupation contexts in the Ongamira Valley, Córdoba, Argentina. J Archaeol Sci Rep 18:65–77
Montalvo CI, Fernández FJ (2019) Review of the actualistic taphonomy of small mammals ingested by South American predators. Its importance in the interpretation of the fossil record Publicación Electrónica de la Asociación Paleontológica Argentina 19: 18–46
Montalvo CI, Pessino M, González V (2007) Taphonomic analysis of remains of mammals eaten by pumas (Puma concolor, Carnivora Felidae) in central Argentina. J Archaeol Sci 34:2151–2160
Montalvo CI, Fernández FJ, Tallade PO (2016a) The role of Bubo virginianus magellanicus as rodent bone accumulator in archaeological sites: a case study for the Atuel River (Mendoza, Argentina). Int J Osteoarchaeol 26:974–986
Montalvo CI, Fernández FJ, Galmes MA, Santillán MA, Cereghetti G (2016b) Crowned solitary eagle (Buteogallus coronatus) as accumulator of armadillo osteoderms in the archaeological record? An actualistic taphonomic study for central Argentina. Quat Int 391:90–99
Montalvo CI, Fernández FJ, Tomassini RL, Mignino J, Kin MS, Santillán MA (2020) Spatial and temporal taphonomic study of bone accumulations of the burrowing owl (Athene cunicularia) in central Argentina. J Archaeol Sci Rep 30:102197
Morgan C (2015) Is it intensification yet? Current archaeological perspectives on the evolution of hunter-gatherer economies. J Archaeol Res 23:163–213
Nabte MJ, Saba SL, Pardiñas UFJ (2006) Dieta del Búho magallánico (Bubo magellanicus) en el desierto del Monte y la Patagonia Argentina. Ornitología Neotropical 17:27–38
Neme G (2007) Cazadores-recolectores de altura en los Andes Meridionales. British Archaeological Reports (BAR), Oxford
Neme G, Gil AF, Otaola C, Giardina M (2015) Resource exploitation and human mobility: trends in the archaeofaunal and isotopic record from Central Western Argentina. Int J Osteoarchaeol 25:866–876
Ojeda RA, Borghi CE, Roig VG (2002) Mamíferos de Argentina. In: Ceballos G, Simonetti JA (eds) Biodiversidad y conservación de mamíferos neotropicales. CONABIO, México, pp 23–63
Olsen S (1971) Zooarchaeology: animal bones in archaeology and their interpretation. Addison-Wesley Pub Co, Reading
Otaola C (2013) ¿Intensificación en la cordillera del sur de Mendoza? Un enfoque tafonómico. Magallania 41:159–176
Ots MJ, García Llorca J, Cahiza P (2016) Recursos y estrategias de consumo en el centro de Mendoza entre los siglos X-XVI AD. Intersecciones Antro 17:375–387
Pardiñas UFJ (1999) Tafonomía de microvertebrados en yacimientos arqueológicos de Patagonia Argentina. Arqueología 9:265–340
Pardiñas UFJ, Teta P, Voglino D, Fernández FJ (2013) Enlarging rodent diversity in west-central Argentina: a new species of the genus Holochilus (Cricetidae, sigmodontinae). J Mammal 94:231–240
Patton JL, Pardiñas UFJ, D’elía G (2015) Mammals of South America, vol 2 Rodents. University of Chicago Press, Chicago and London
Prieto M del R, Chiavazza H (2006) Aportes de la historia ambiental para el análisis del patrón de asentamiento huarpe en el oasis norte de Mendoza. Anales de Arqueología y Etnología 58-59:163–195
Prieto M del R, Rojas F (2012) Documentary evidence for changing climatic and anthropogenic influences on the Bermejo wetland in Mendoza, Argentina, during the 16th-20th century. Clim Past 8:951–961
Prieto Olavarría C, Chiavazza H (2009) El ocaso de los pueblos y el inicio de la urbe. Las cerámicas indígenas del valle de Güentata (Mendoza). In: Oliva F, de Grandis N, Rodríguez J (eds.), Arqueología Argentina en los inicios de un nuevo siglo, II. Laborde Editor. Rosario, pp. 1–14
Prieto-Olavarría C, Chiavazza H, Musaubach MG (2019) Microrrestos de vegetales cultivados y silvestres en fragmentos cerámicos. Primeras evidencias en el Centro Occidente Argentino. Arqueología 25:221–231
Quintana CA (2005) Despiece de microroedores en el Holoceno Tardío de las Sierras de Tandilia (Argentina). Archaeofauna 14:227–241
Rhodes SE, Walker MJ, López-Jiménez A, López-Martínez M, Haber-Uriarte M, Fernández-Jalvo Y, Chazan M (2016) Fire in the Early Palaeolithic: evidence from burnt small mammal bones at Cueva Negra del Estrecho del Río Quípar, Murcia, Spain. J Archaeol Sci Rep 9:427–436
Rhodes SE, Ziegler R, Starkovich BM, Conard NJ (2018) Small mammal taxonomy, taphonomy, and the paleoenvironmental record during the Middle and Upper Paleolithic at Geißenklösterle cave (Ach Valley, southwestern Germany). Quat Sci Rev 185:199–221
Rhodes SE, Starkovich BM, Conard NJ (2019) Did climate determine Late Pleistocene settlement dynamics in the Ach Valley, SW Germany? PLoS One 14:e0215172
Romaniuk AA, Shepherd AN, Clarke DV, Sheridan AJ, Fraser S, Bartoziewicz L, Herman JS (2016) Rodents: food or pests in Neolithic Orkney. R Soc Open Sci 3:160514
Rundel PW, Villagra PE, Dillon MO, Roig-Juñent S, Debandi G (2007) Arid and semi-arid ecosystems. In: Veblen T, Young K, Orme A (eds) The physical geography of South America. Oxford University Press, Oxford, pp 158–183
Santiago F, Salemme M, Pardiñas UFJ (2016) Análisis de restos de roedores del sitio arqueológico Las Vueltas 1, Tierra del Fuego, Argentina. Arqueología 22 (Dossier): 211-230
Shipman P, Fosterb G, Schoeninger M (1984) Burnt bones and teeth: an experimental study of color, morphology, crystal structure and shrinkage. J Archaeol Sci 11:301–325
Simonetti JA, Cornejo LE (1991) Archaeological evidence of rodent consumption in Central Chile. Lat Am Antiq 2:92–96
Stiner M (1994) Honor among thieves. A Zooarchaeological Study of Neandertal Ecology. Princeton University Press, Princeton
Acknowledgements
We thank Fernando J. Fernández for his help and advices. Clara Otaola and J. Tyler Faith provided valuable comments on an early version of the manuscript. Comments and suggestions made by Juan Manuel López-García and one anonymous reviewer improved the first version of the manuscript. This work was made in the framework of the following projects: 06/G710, 06/G653, 06/G805, 06/G792 (Facultad de Filosofía y Letras, Universidad Nacional de Cuyo, Argentina), PICT 2013/0190 (Agencia Nacional de Promoción Científica y Tecnológica, Argentina) and 05G (Facultad de Ciencias Exactas y Naturales, Universidad Nacional de La Pampa, Argentina). Juan Pablo Aguilar helped us with the map used in Fig. 1. We thank Silvia Barbuzza (CETI, Facultad de Filosofía y Letras, Universidad Nacional de Cuyo) for her English editing and proofreading work.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
López, J.M., Chiavazza, H. From owl prey to human food: taphonomy of archaeological small mammal remains from the late Holocene wetlands of arid environments in Central Western Argentina. Archaeol Anthropol Sci 12, 276 (2020). https://doi.org/10.1007/s12520-020-01213-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-020-01213-z