
Abstract
Vulvovaginal candidiasis (VVC) is a prevalent condition affecting a significant portion of women worldwide. Licochalcone A (LA), a natural compound with diverse biological activities, holds promise as a protective agent against Candida albicans (C. albicans) infection. This study aims to investigate the potential of LA to safeguard vaginal epithelial cells (VECs) from C. albicans infection and elucidate the underlying molecular mechanisms. To simulate VVC in vitro, VK2-E6E7 cells were infected with C. albicans. Candida albicans biofilm formation, C. albicans adhesion to VK2-E6E7 cells, and C. albicans-induced cell damage and inflammatory responses were assessed by XTT reduction assay, fluorescence assay, LDH assay, and ELISA. CCK-8 assay was performed to evaluate the cytotoxic effects of LA on VK2-E6E7 cells. Western blotting assay was performed to detect protein expression. LA dose-dependently hindered C. albicans biofilm formation and adhesion to VK2-E6E7 cells. Furthermore, LA mitigated cell damage, inhibited the Bax/Bcl-2 ratio, and attenuated the secretion of pro-inflammatory cytokines in C. albicans-induced VK2-E6E7 cells. The investigation into LA’s impact on the Toll-like receptor 4 (TLR4)/nuclear factor-kappa B (NF-κB) pathway revealed that LA downregulated TLR4 expression and inhibited NF-κB activation in C. albicans-infected VK2-E6E7 cells. Furthermore, TLR4 overexpression partially abated LA-mediated protection, further highlighting the role of the TLR4/NF-κB pathway. LA holds the potential to safeguard VECs against C. albicans infection, potentially offering therapeutic avenues for VVC management.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Vulvovaginal candidiasis (VVC), a common vaginal infection (Nicolau Costa et al., 2021), is characterized by symptoms such as itching, burning, and abnormal vaginal discharge, leading to discomfort and decreased quality of life for affected individuals (Jacobsen, 2023). This condition afflicts a significant number of women worldwide, with about 75% experiencing at least one episode of VVC during their reproductive years (Lopes & Lionakis, 2022).
The overgrowth of Candida albicans (C. albicans) accounts for approximately 80–90% of VVC cases (Gaziano et al., 2020). Candida albicans, a commensal organism present in the vaginal microbiota of many healthy women (Spaggiari et al., 2023), can transition from a commensal state to a pathogenic state under certain conditions, including alterations in the vaginal environment or compromised immune defenses, precipitating symptomatic VVC (Niu et al., 2017). Such a pathogenic transition involves the adhesion of C. albicans to vaginal epithelial cells (VECs), and subsequent invasion and colonization (Zhao et al., 2022). VECs serve as a critical physical barrier against C. albicans by impeding their penetration into the vaginal mucosa (Saha et al., 2017; Zhang et al., 2018). Additionally, VECs also actively participate in the immune response against C. albicans infection by producing antimicrobial peptides and pro-inflammatory cytokines that aid in clearing the invading pathogens (Helmy et al., 2021). Therefore, safeguarding VECs from C. albicans adhesion and subsequent cell damage holds promise for VVC treatment.
Licochalcone A (LA), a natural flavonoid compound derived from licorice root (Glycyrrhiza species), has garnered increasing attention due to its potent antimicrobial properties (Lv et al., 2019). While exhibiting inhibitory effects on a range of microorganisms, including bacteria, viruses, and fungi (El-Saber Batiha et al., 2020), LA exhibits minimal cytotoxicity towards mammalian cells (Wu et al., 2019), enhancing its potential for clinical application. Particularly noteworthy, several studies have highlighted the capacity of LA to impede C. albicans biofilm formation and adhesion (Messier & Grenier, 2011; Seleem et al., 2016), suggesting its therapeutic potential for VVC.
This study aimed to explore the therapeutic effects of LA on VVC. To mimic VVC in vitro, we established a cellular model by infecting VK2-E6E7 cells with C. albicans, a widely used approach for investigating molecular mechanisms and drug development in VVC (Luan et al., 2020; Tan et al., 2023). Herein, we evaluated the impact of LA on C. albicans biofilm formation, adhesion to VK2-E6E7 cells, as well as C. albicans-induced cell damage and inflammatory responses in VK2-E6E7 cells using XTT reduction assay (Chaillot et al., 2017), fluorescence assay (Mikamo et al., 2018), LDH assay (Chaillot et al., 2017), and ELISA (Gao et al., 2019). Furthermore, this study delved into the underlying molecular mechanisms, with a focus on the Toll-like receptor 4 (TLR4)/nuclear factor-kappa B (NF-κB) signaling pathway. This work provides insight into the development of potential therapeutic strategies for VVC.
Materials and Methods
Microbial Strain
The C. albicans strain SC5314, obtained from the American Type Culture Collection (ATCC), was utilized in this study. Candida albicans were routinely cultivated in YPD liquid medium (1% yeast extract, 2% peptone, and 2% dextrose) at 30 °C overnight and washed twice with PBS before subsequent cellular experiments.
Cell Culture and Treatment
The vaginal epithelial cell line (VK2-E6E7) (ATCC) was cultured in Keratinocyte Serum Free Medium (Invitrogen) within a humidified incubator (37 °C; 5% CO2).
To replicate VVC in vitro, VK2-E6E7 cells were seeded into 96-well plates and grown to 95% confluency before infection with C. albicans at a concentration of 5 × 105 colony-forming units (CFU)/ml. LA powder (#68783; Sigma-Aldrich) was dissolved in dimethyl sulfoxide (DMSO; #D2650; Sigma-Aldrich) to prepare a 50 mM stock which was subsequently diluted to the desired concentrations. Prior to C. albicans infection, VK2-E6E7 cells were treated with LA for 2 h.
Cell Transfection
TLR4-specific pcDNA overexpression vector (TLR4-OE) and pcDNA3.1 empty vector (Vector) were purchased from Genechem. Transfection of these plasmids into VK2-E6E7 cells was performed using Lipofectamine®2000 reagent (Invitrogen), as previously described (Zhao et al., 2023). Transfected cells were incubated for 48 h prior to subsequent experiments.
Cytotoxicity Assay
The cytotoxic effect of LA on VK2-E6E7 cells was assessed using a CCK-8 assay, as previously described (Li et al., 2023). VK2-E6E7 cells were seeded into a 96-well plate (5 × 103 cells/well) and cultured with varying doses of LA (5, 10, 20, 40, 80, and 160 μM) for 24 h or 48 h. Then, CCK-8 reagent (#96,992, Sigma-Aldrich) was added (10 μl/well) and VK2-E6E7 cells were incubated for another 2 h in darkness. The absorbance value at 450 nm was measured using a microplate reader (BioTek).
XTT Reduction Assay
Biofilm formation of C. albicans was evaluated by measuring the metabolic activity using a colorimetric XTT reduction assay, as previously outlined (Alfian et al., 2022). Candida albicans (2 × 105 CFU) were seeded into 96-well plates and incubated with LA (62.5 μM or 150 μM) at 37 °C for 24 h. At the end of incubation, XTT solution was added (100 µl/well), and OD values were measured at 492 nm using a microplate reader.
Adherence of C. Albicans
Candida albicans adherence to VK2-E6E7 cells was assessed using a fluorescence assay. Candida albicans were labeled with fluorescein isothiocyanate (FITC; #46,950, Sigma-Aldrich) as previously described (Mikamo et al., 2018). VK2-E6E7 cell monolayers were cultured with FITC-labeled C. albicans, and fluorescence intensity was measured using a fluorescence microplate reader.
LDH Assay
VK2-E6E7 cell damage induced by C. albicans infection was assessed by detecting LDH activity in culture supernatants using the CytoTox 96 Non-Radioactive Cytotoxicity Assay Kit (#G1780; Promega) (Gao et al., 2019). VK2-E6E7 cell monolayers were cultured with C. albicans for 24 h. Then, CytoTox 96® reagent (50 μl/well) and stop solution (50 μl/well) were added. The absorbance values were observed at 490 nm.
ELISA
After treatments, pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6) in culture supernatants were quantified using commercially available antibodies (Pharmingen) (Steele & Fidel, 2002).
Western Blotting
Western blotting analysis was performed as previously described (Shroff et al., 2017). Protein lysates were extracted from VK2-E6E7 cells by RIPA buffer. After concentration quantification using a BCA Protein Assay Kit (#23,250, Thermo Fisher Scientific), equal amounts of protein lysates were separated by 10% SDS-PAGE and transferred onto PVDF membranes. Then, the membranes were blocked with 5% skim milk and probed with specific primary and secondary antibodies. Protein bands were visualized with an ECL kit and quantified using ImageJ software.
Statistical Analysis
All experiments were performed at least three times. Data are expressed as mean ± SD. Statistical analyses were performed using SPSS 21.0 software by Student’s t-test or one-way analysis of variance (ANOVA) with significance set at P < 0.05. Graphs were generated using GraphPad 6.0 software.
Results
Determination of Cytotoxic Effects of LA on VK2-E6E7 Cells
The chemical structure of LA is depicted in Fig. 1A. As previously reported (Seleem et al., 2016), the minimum inhibitory concentration (MIC) and minimum fungicidal concentration (MFC) values of LA against C. albicans were determined to be 62.5 μM and 150 μM, respectively. While the MIC of LA against C. albicans appears high, both in vitro and in vivo experiments have confirmed its efficacy against C. albicans with minimal toxicity observed against oral fibroblast cells and tongue tissue, suggesting the potential of LA as a novel antifungal agent for C. albicans-associated diseases, including VVC.

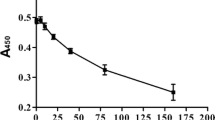
Determination of cytotoxic effects of LA on VK2-E6E7 cells. (A) The chemical structure of LA. (B and C) VK2-E6E7 cells were treated with LA at different doses (5, 10, 20, 40, 80, and 160 μM) for 24 and 48 h. The cytotoxic effect of LA on VK2-E6E7 cells was evaluated by CCK-8 assay. *P < 0.05; **P < 0.01
To determine the optimal concentration of LA for subsequent experiments, the cytotoxic effects of LA on VK2-E6E7 cells were assessed within a concentration range of 0–160 μM. CCK-8 results demonstrated that even at a concentration of 160 μM, LA did not induce noticeable cytotoxic effects on VK2-E6E7 cells after 24 or 48 h of incubation (Fig. 1B and C). Consequently, concentrations of 62.5 μM and 150 μM were chosen for subsequent experiments.
LA inhibits C. Albicans Biofilm Formation and Adhesion to VK2-E6E7 Cells
To examine the impact of LA on C. albicans biofilm formation, VK2-E6E7 cells were pretreated with LA (62.5 μM or 150 μM) 2 h prior to C. albicans infection. As illustrated in Fig. 2A, LA exhibited a dose-dependent inhibition of C. albicans biofilm formation. Additionally, the effects of LA on the adhesion of C. albicans to VK2-E6E7 cells were assessed. VK2-E6E7 cells were categorized into DMSO, C. albicans, C. albicans + LA (62.5 μM), and C. albicans + LA (150 μM) groups. Analysis of fluorescence emitted by FITC-labeled C. albicans demonstrated that LA decreased the adhesion of C. albicans to VK2-E6E7 cells in a dose-dependent manner (Fig. 2B). Thus, LA effectively suppresses both C. albicans biofilm formation and its adhesion to VK2-E6E7 cells.

LA inhibits C. albicans biofilm formation and adhesion to VK2-E6E7 cells. (A) The biofilm formation of C. albicans treated with LA at 0, 62.5 μM, or 150 μM groups was assessed by XTT reduction assay. (B) VK2-E6E7 cells were pretreated with LA at 62.5 μM or 150 μM and then infected with FITC-labeled C. albicans for 24 h. Candida albicans adhesion to VK2-E6E7 cells was assessed by measuring fluorescence intensity under a fluorescence microplate reader. *P < 0.05; **P < 0.01
LA Mitigates C. albicans-Induced Cell Damage and Inflammatory Responses in VK2-E6E7 Cells
Using an LDH assay, C. albicans-induced damage to VK2-E6E7 cells after 24 h of infection was measured. LA demonstrated a concentration-dependent ability to prevent C. albicans-induced damage to VK2-E6E7 cells (Fig. 3A). Furthermore, LA attenuated the C. albicans-induced increase in Bax protein levels and decrease in Bcl-2 protein levels in VK2-E6E7 cells (Fig. 3B). Subsequently, ELISA was employed to quantify the levels of pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6) secreted by VK2-E6E7 cells. Results depicted in Fig. 3C revealed that C. albicans infection significantly elevated the production of TNF-α, IL-1β, and IL-6 by VK2-E6E7 cells, whereas LA dose-dependently downregulated these levels. Therefore, LA exhibits a dose-dependent inhibition of cell damage and inflammatory responses induced by C. albicans in VK2-E6E7 cells.

LA mitigates C. albicans-induced cell damage and inflammatory responses in VK2-E6E7 cells. VK2-E6E7 cells were pretreated with LA at 62.5 μM or 150 μM and then infected with C. albicans for 24 h. (A) VK2-E6E7 cell damage was determined using LDH assay. (B) Bax and Bcl-2 protein levels in VK2-E6E7 cells were detected by western blotting. (C) TNF-α, IL-1β, and IL-6 levels in the culture supernatant were determined using ELISA. *P < 0.05; **P < 0.01
LA Inhibits the Activation of the TLR4/NF-κB Signaling Pathway in C. albicans-Infected VK2-E6E7 Cells
The toll-like receptor 2/4 (TLR2/4)-NF-κB signaling pathway has been implicated in VVC pathogenesis (Feng et al., 2023). Notably, LA has been shown to inhibit TLR4 expression and NF-κB signaling in LPS-induced C57BL/6 mice and macrophages (Cai et al., 2023). Thus, the levels of TLR2, TLR4, and NF-κB-related proteins (NF-κB p65 and p-NF-κB p65) were assessed in VK2-E6E7 cells from the DMSO, C. albicans, C. albicans + LA (62.5 μM), and C. albicans + LA (150 μM) groups. Western blotting analysis revealed that C. albicans infection significantly increased TLR2, TLR4, and p-NF-κB p65 protein levels; however, LA treatment dose-dependently reversed the increase in TLR4 and p-NF-κBp 65 protein levels, while barely affecting TLR2 protein levels (Fig. 4A). These results suggest that LA may downregulate TLR4 expression and inhibit NF-κB activation in C. albicans-infected VK2-E6E7 cells.

LA inhibits the activation of the TLR4/NF-κB signaling pathway in C. albicans-infected VK2-E6E7 cells. (A) VK2-E6E7 cells were pretreated with LA at 62.5 μM or 150 μM and then infected with C. albicans for 24 h. TLR2, TLR4, p-NF-κB p65, and NF-κB p65 protein levels in VK2-E6E7 cells were detected by western blotting. (B) VK2-E6E7 cells were transfected with Vector and TLR4-OE. TLR4 protein levels in transfected VK2-E6E7 cells were determined by western blotting. (C) VK2-E6E7 cells were transfected with TLR4-OE, pretreated with LA at 150 μM, and infected with C. albicans for 24 h. TLR4, p-NF-κB p65, and NF-κB p65 protein levels in VK2-E6E7 cells were determined by western blotting. *P < 0.05; **P < 0.01
To further verify whether LA inhibits the TLR4/NF-κB signaling pathway in VK2-E6E7 cells infected by C. albicans, TLR4 was first overexpressed in VK2-E6E7 cells. The efficiency of transfection was confirmed by western blotting (Fig. 4B). Subsequently, cells were assigned to DMSO, C. albicans, C. albicans + LA (150 μM), C. albicans + TLR4-OE, and C. albicans + LA (150 μM) + TLR4-OE groups. As depicted in Fig. 4C, TLR4 overexpression augmented the C. albicans-induced increase in TLR4 and p-NF-κB protein expression and significantly reversed LA-mediated inhibition of these proteins. Therefore, the TLR4/NF-κB signaling pathway may play a role in LA-meditated protection against C. albicans infection.
TLR4 Overexpression Abates LA-Mediated NF-κB Deactivation and Protection Against C. albicans Infection in VK2-E6E7 Cells
Next, we investigated whether the TLR4/NF-κB signaling pathway is involved in the protective effects of LA. As shown in Fig. 5A, TLR4 overexpression exacerbated C. albicans adhesion to VK2-E6E7 cells and partially mitigated the LA-mediated reduction of C. albicans adhesion to VK2-E6E7 cells. Moreover, TLR4 overexpression also exacerbated cell damage and inflammatory responses induced by C. albicans in VK2-E6E7 cells and abrogated LA-mediated inhibition of these responses (Fig. 5B–D). In summary, LA may protect VECs against C. albicans infection by inhibiting the TLR4/NF-κB signaling pathway.

TLR4 overexpression abates LA-mediated NF-κB deactivation and protection against C. albicans infection in VK2-E6E7 cells. VK2-E6E7 cells were transfected with TLR4-OE, pretreated with LA at 150 μM, and infected with C. albicans for 24 h. (A) C. albicans adhesion to VK2-E6E7 cells was assessed by measuring fluorescence intensity under a fluorescence microplate reader. (B) VK2-E6E7 cell damage was determined using LDH assay. (C) Bax and Bcl-2 protein levels in VK2-E6E7 cells were detected by western blotting. (D) TNF-α, IL-1β, and IL-6 levels in the culture supernatant were determined using ELISA. *P < 0.05; **P < 0.01
Discussion
VVC presents a prevalent concern affecting a substantial portion of women globally. In the pursuit of effective therapeutic interventions, this study delves into the potential of LA as a protective agent for VECs (VK2-E6E7 cells) against C. albicans infection and elucidates its underlying mechanisms, particularly focusing on the TLR4/NF-κB signaling pathway.
The interaction between C. albicans and VECs involves processes of adhesion, invasion, and subsequent cellular damage (Wachtler et al., 2011). Notably, prior research has shown that LA inhibits C. albicans growth in epithelial cells (Messier & Grenier, 2011) and suppresses C. albicans biofilm formation and adhesion to oral fibroblast cells (Seleem et al., 2016). Consistent with these findings, our study unveils LA’s efficacy in inhibiting C. albicans biofilm formation and adhesion to VK2-E6E7 cells in a dose-dependent manner, indicating that LA exerts protective effects on VECs by impeding C. albicans biofilm formation and adhesion.
In the context of VVC, a protective immune response often leads to concurrent tissue inflammation (Yu et al., 2018). Recent studies have demonstrated elevated production of pro-inflammatory factors by VVC mice and C. albicans-infected VK2-E6E7 cells (Chen et al., 2021; Zhang et al., 2022). LA has been found to exert anti-inflammatory effects by suppressing the secretion of pro-inflammatory cytokines (Chu et al., 2012; Li et al., 2021; Shu et al., 2022) and protecting against cell damage and subsequent tissue injury (Hou et al., 2019; Zhu et al., 2023). This is further supported by our findings that LA can mitigate C. albicans-induced cell damage and inflammatory responses in VK2-E6E7 cells, evidenced by concentration-dependent inhibition of C. albicans-induced increase in LDH activity, Bax/Bcl-2 ration, and the production of TNF-α, IL-1β, and IL-6. These findings underscore LA’s potential to alleviate tissue damage and inflammation in VVC.
In response to C. albicans infection, the TLRs/NF-κB signaling pathway is generally triggered and activated (Liu et al., 2018; Wang et al., 2013), playing a significant role in VVC pathogenesis (Feng et al., 2023). Given that LA has been reported to inhibit the TLR4/NF-κB signaling pathway in LPS/GalN-induced acute liver injury (Lv et al., 2019), we investigated its regulatory effects on the TLRs/NF-κB signaling pathway in C. albicans-infected VK2-E6E7 cells. Our results unequivocally demonstrate that LA downregulates TLR4 expression and inhibits NF-κB activation in C. albicans-infected VK2-E6E7 cells, highlighting LA’s modulatory impact on the TLR4/NF-κB signaling pathway. Furthermore, overexpression of TLR4 partially reverses LA-mediated inhibitory effects on C. albicans-induced NF-κB activation and LA’s protective effects on VK2-E6E7 cells against C. albicans infection, emphasizing the pivotal role of the TLR4/NF-κB signaling pathway in LA’s capacity to safeguard VECs from C. albicans infection. These results also corroborate the findings of previous studies on LA, which support that the anti-inflammatory properties of LA manifest through the inhibition of the NF-κB signaling pathway (Furusawa et al., 2009; Hu & Liu, 2016).
Conclusion
This study sheds light on the potential of LA in safeguarding VECs against C. albicans infection. The demonstrated impacts on C. albicans biofilm formation, adhesion to VECs, as well as the mitigation of C. albicans-induced cellular damage and inflammatory responses in VECs, alongside its regulation of the TLR4/NF-κB pathway, position LA as a promising therapeutic agent for VVC. Future research endeavors should delve deeper into exploring the effects of LA on the yeast-to-hyphal morphological transition of C. albicans and its efficacy in treating VVC in vivo.
Data Availability
Data are available with reasonable requirements.
References
Alfian, A. R., Watchaputi, K., Sooklim, C., & Soontorngun, N. (2022). Production of new antimicrobial palm oil-derived sophorolipids by the yeast Starmerella riodocensis sp. nov. against Candida albicans hyphal and biofilm formation. Microbial Cell Factories, 21, 163.
Cai, M., Xu, Y. C., Deng, B., Chen, J. B., Chen, T. F., Zeng, K. F., Chen, S., Deng, S., Tan, Z., Ding, W., et al. (2023). Radix Glycyrrhizae extract and licochalcone a exert an anti-inflammatory action by direct suppression of toll like receptor 4. Journal of Ethnopharmacology, 302, 115869.
Chaillot, J., Tebbji, F., García, C., Wurtele, H., Pelletier, R., & Sellam, A. (2017). pH-dependant antifungal activity of valproic acid against the human fungal pathogen Candida albicans. Frontiers in Microbiology, 8, 1956.
Chen, Z., Luo, T., Huang, F., Yang, F., Luo, W., Chen, G., Cao, M., Wang, F., & Zhang, J. (2021). Kangbainian lotion ameliorates vulvovaginal candidiasis in mice by inhibiting the growth of fluconazole-resistant Candida albicans and the dectin-1 signaling pathway activation. Frontiers in Pharmacology, 12, 816290.
Chu, X., Ci, X., Wei, M., Yang, X., Cao, Q., Guan, M., Li, H., Deng, Y., Feng, H., & Deng, X. (2012). Licochalcone a inhibits lipopolysaccharide-induced inflammatory response in vitro and in vivo. Journal of Agricultural and Food Chemistry, 60, 3947–3954.
El-Saber Batiha, G., MagdyBeshbishy, A., El-Mleeh, A., Abdel-Daim, M. M., & Prasad Devkota, H. (2020). Traditional uses, bioactive chemical constituents, and pharmacological and toxicological activities of Glycyrrhiza glabra L. (Fabaceae). Biomolecules, 10, 352.
Feng, X., Zhang, H., Hu, K., Shi, G., Wu, D., Shao, J., Wang, T., & Wang, C. (2023). Longdan Xiegan decoction ameliorates vulvovaginal candidiasis by inhibiting the NLRP3 inflammasome via the Toll-like receptor /MyD88 pathway. Journal of Ethnopharmacology, 318, 116869.
Furusawa, J., Funakoshi-Tago, M., Mashino, T., Tago, K., Inoue, H., Sonoda, Y., & Kasahara, T. (2009). Glycyrrhiza inflata-derived chalcones, Licochalcone A, Licochalcone B and Licochalcone D, inhibit phosphorylation of NF-κB p65 in LPS signaling pathway. International Immunopharmacology, 9, 499–507.
Gao, Y., Liang, G., Wang, Q., She, X., Shi, D., Shen, Y., Su, X., Wang, X., Wang, W., Li, D., et al. (2019). Different host immunological response to C. albicans by human oral and vaginal epithelial cells. Mycopathologia, 184, 1–12.
Gaziano, R., Sabbatini, S., Roselletti, E., Perito, S., & Monari, C. (2020). Saccharomyces cerevisiae-based probiotics as novel antimicrobial agents to prevent and treat vaginal infections. Frontiers in Microbiology, 11, 718.
Helmy, Y. A., Kassem, I. I., & Rajashekara, G. (2021). Immuno-modulatory effect of probiotic E. coli Nissle 1917 in polarized human colonic cells against Campylobacter jejuni infection. Gut Microbes, 13, 1857514.
Hou, X., Yang, S., & Zheng, Y. (2019). Licochalcone A attenuates abdominal aortic aneurysm induced by angiotensin II via regulating the miR-181b/SIRT1/HO-1 signaling. Journal of Cellular Physiology, 234, 7560–7568.
Hu, J., & Liu, J. (2016). Licochalcone A attenuates lipopolysaccharide-induced acute kidney injury by inhibiting NF-κB activation. Inflammation, 39, 569–574.
Jacobsen, I. D. (2023). The role of host and fungal factors in the commensal-to-pathogen transition of Candida albicans. Current Clinical Microbiology Reports, 10, 55–65.
Li, P., Yu, C., Zeng, F. S., Fu, X., Yuan, X. J., Wang, Q., Fan, C., Sun, B. L., & Sun, Q. S. (2021). Licochalcone A attenuates chronic neuropathic pain in rats by inhibiting microglia activation and inflammation. Neurochemical Research, 46, 1112–1118.
Li, D., Zhang, T., Yang, H., Yang, W., Zhang, C., & Gao, G. (2023). Effect of vitamin D on the proliferation and barrier of atrophic vaginal epithelial cells. Molecules, 28, 6605.
Liu, J., Geng, F., Sun, H., Wang, X., Zhang, H., Yang, Q., & Zhang, J. (2018). Candida albicans induces TLR2/MyD88/ NF-κB signaling and inflammation in oral lichen planus-derived keratinocytes. Journal of Infection in Developing Countries, 12, 780–786.
Lopes, J. P., & Lionakis, M. S. (2022). Pathogenesis and virulence of Candida albicans. Virulence, 13, 89–121.
Luan, T., Liu, X., Mao, P., Wang, X., Rui, C., Yan, L., Wang, Y., Fan, C., Li, P., & Zeng, X. (2020). The role of 17β-estrogen in Candida albicans adhesion on human vaginal epithelial cells via FAK phosphorylation. Mycopathologia, 185, 425–438.
Lv, H., Yang, H., Wang, Z., Feng, H., Deng, X., Cheng, G., & Ci, X. (2019). Nrf2 signaling and autophagy are complementary in protecting lipopolysaccharide/d-galactosamine-induced acute liver injury by licochalcone A. Cell Death & Disease, 10, 313.
Messier, C., & Grenier, D. (2011). Effect of licorice compounds licochalcone A, glabridin and glycyrrhizic acid on growth and virulence properties of Candida albicans. Mycoses, 54, e801–e806.
Mikamo, H., Yamagishi, Y., Sugiyama, H., Sadakata, H., Miyazaki, S., Sano, T., & Tomita, T. (2018). High glucose-mediated overexpression of ICAM-1 in human vaginal epithelial cells increases adhesion of Candida albicans. Journal of Obstetrics and Gynaecology, 38, 226–230.
Nicolau Costa, K. M., Sato, M. R., Barbosa, T. L. A., Rodrigues, M. G. F., Medeiros, A. C. D., Damasceno, B., & Oshiro-Junior, J. A. (2021). Curcumin-loaded micelles dispersed in ureasil-polyether materials for a novel sustained-release formulation. Pharmaceutics, 13, 675.
Niu, X. X., Li, T., Zhang, X., Wang, S. X., & Liu, Z. H. (2017). Lactobacillus crispatus modulates vaginal epithelial cell innate response to Candida albicans. Chinese Medical Journal, 130, 273–279.
Saha, D., Koli, S., Patgaonkar, M., & Reddy, K. V. (2017). Expression of hemoglobin-α and β subunits in human vaginal epithelial cells and their functional significance. PLoS ONE, 12, e0171084.
Seleem, D., Benso, B., Noguti, J., Pardi, V., & Murata, R. M. (2016). In vitro and in vivo antifungal activity of lichochalcone-A against Candida albicans biofilms. PLoS ONE, 11, e0157188.
Shroff, A., Sequeira, R., & Reddy, K. V. R. (2017). Human vaginal epithelial cells augment autophagy marker genes in response to Candida albicans infection. American Journal of Reproductive Immunology, 77, e12639.
Shu, J., Cui, X., Liu, X., Yu, W., Zhang, W., Huo, X., & Lu, C. (2022). Licochalcone A inhibits IgE-mediated allergic reaction through PLC/ERK/STAT3 pathway. International Journal of Immunopathology and Pharmacology, 36. https://doi.org/10.1177/03946320221135462
Spaggiari, L., Squartini Ramos, G. B., Squartini Ramos, C. A., Ardizzoni, A., Pedretti, N., Blasi, E., De Seta, F., & Pericolini, E. (2023). Anti-Candida and anti-inflammatory properties of a vaginal gel formulation: Novel data concerning vaginal infection and dysbiosis. Microorganisms, 11, 1551.
Steele, C., & Fidel, P. L., Jr. (2002). Cytokine and chemokine production by human oral and vaginal epithelial cells in response to Candida albicans. Infection and Immunity, 70, 577–583.
Tan, Y., Lin, Q., Yao, J., Zhang, G., Peng, X., & Tian, J. (2023). In vitro outcomes of quercetin on Candida albicans planktonic and biofilm cells and in vivo effects on vulvovaginal candidiasis. Evidences of its mechanisms of action. Phytomedicine, 114, 154800.
Wachtler, B., Wilson, D., & Hube, B. (2011). Candida albicans adhesion to and invasion and damage of vaginal epithelial cells: Stage-specific inhibition by clotrimazole and bifonazole. Antimicrobial Agents and Chemotherapy, 55, 4436–4439.
Wang, M., Wang, F., Yang, J., Zhao, D., Wang, H., Shao, F., Wang, W., Sun, R., Ling, M., Zhai, J., & Song, S. (2013). Mannan-binding lectin inhibits Candida albicans-induced cellular responses in PMA-activated THP-1 cells through Toll-like receptor 2 and Toll-like receptor 4. PLoS ONE, 8, e83517.
Wu, S. C., Yang, Z. Q., Liu, F., Peng, W. J., Qu, S. Q., Li, Q., Song, X. B., Zhu, K., & Shen, J. Z. (2019). Antibacterial effect and mode of action of flavonoids from licorice against methicillin-resistant Staphylococcus aureus. Frontiers in Microbiology, 10, 2489.
Yu, X. Y., Fu, F., Kong, W. N., Xuan, Q. K., Wen, D. H., Chen, X. Q., He, Y. M., He, L. H., Guo, J., Zhou, A. P., et al. (2018). Streptococcus agalactiae inhibits Candida albicans hyphal development and diminishes host vaginal mucosal TH17 response. Frontiers in Microbiology, 9, 198.
Zhang, X., Li, T., Chen, X., Wang, S., & Liu, Z. (2018). Nystatin enhances the immune response against Candida albicans and protects the ultrastructure of the vaginal epithelium in a rat model of vulvovaginal candidiasis. BMC Microbiology, 18, 166.
Zhang, J., Peng, J., Li, D., Mei, H., Yu, Y., Li, X., She, X., & Liu, W. (2022). Divergent EGFR/MAPK-mediated immune responses to clinical Candida pathogens in vulvovaginal candidiasis. Frontiers in Immunology, 13, 894069.
Zhao, T., Zhang, K., Shi, G., Ma, K., Wang, B., Shao, J., Wang, T., & Wang, C. (2022). Berberine inhibits the adhesion of Candida albicans to vaginal epithelial cells. Frontiers in Pharmacology, 13, 814883.
Zhao, J., Ma, M., Li, L., & Fang, G. (2023). Oxysophoridine protects against cerebral ischemia/reperfusion injury via inhibition of TLR4/p38MAPK-mediated ferroptosis. Molecular Medicine Reports, 27, 44.
Zhu, W., Wang, M., Jin, L., Yang, B., Bai, B., Mutsinze, R. N., Zuo, W., Chattipakorn, N., Huh, J. Y., Liang, G., et al. (2023). Licochalcone A protects against LPS-induced inflammation and acute lung injury by directly binding with myeloid differentiation factor 2 (MD2). British Journal of Pharmacology, 180, 1114–1131.
Acknowledgements
This study is supported by Zhenjiang Key Research and Development Plan—Social Development (SH2023010) and Open-Fund Research Topic of Zhenjiang Clinical Research Center for Obstetrics and Gynecology (SS2022003-KFA02).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors have no conflict of interest to report.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Li, W., Yin, Y., Li, T. et al. Licochalcone A Protects Vaginal Epithelial Cells Against Candida albicans Infection Via the TLR4/NF-κB Signaling Pathway. J Microbiol. 62, 525–533 (2024). https://doi.org/10.1007/s12275-024-00134-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12275-024-00134-z