
Abstract
Most descriptions of the extensor mechanism of the knee do not take into account its complexity and variability. The quadriceps femoris insertion into the patella is said to be through a common tendon with a three-layered arrangement: rectus femoris (RF) most superficially, vastus medialis (VM) and lateralis (VL) in the intermediate layer, and vastus intermedius (VI) most deeply. We dissected 20 limbs from 17 cadavers to provide a more detailed description of the anterior components of the knee: the tendon, the patellar retinacula, and the patellofemoral ligaments. Only three of the 20 specimens exhibited the typically described quadriceps pattern. The remainder had bilaminar and even more complex trilaminar and tetralaminar fiber arrangements. We found an oblique head of the vastus lateralis (VLO), separated from the longitudinal head by a layer of fat or fascia, in 60% of the specimens. However, we found no distinct oblique head of the vastus medialis (VMO) in any specimen. The medial patellofemoral ligament (MPFL) was more common than the lateral (LPFL), supporting its suggested role as the principal passive medial stabilizer of the patella. Because the quadriceps muscle group plays a direct role in patellofemoral joint function, investigation into the clinical applications of its highly variable anatomy may be worthwhile with respect to joint dysfunction and failures of TKAs.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The failure of many surgical procedures (eg, TKA) on the knee is believed to originate frequently in disorders of the mechanics of the joint. Patellar instability, including subluxation and maltracking, relates to the imbalance of the soft tissues. With its incidence being reported as much as 29% after TKAs, patellar instability is one of the most common problems after these procedures [26]. Atrophy [31], weakness [12], and a more proximal attachment of the VMO [13] and overdevelopment of the VL [20] have been suggested as causes of patellofemoral dysfunction. However, there are other components of the extensor mechanism that also could be implicated.
Farahmand et al. [12] noted the lack of data that may be used for the design or assessment of surgical procedures involving the patellofemoral joint stemming from a shortage of studies relating to joint structure and the balance among the quadriceps muscle components. Currently, much of the literature describes the quadriceps femoris insertion into the patella as a common tendon with a three-layered arrangement: RF being the most superficial, VMO and VL forming the intermediate, and VI being the deep layer. The true insertion of the quadriceps femoris is into the tibial tubercle, but convention considers it as inserting into the patella (Fig. 1) through the quadriceps tendon or the suprapatellar tendon, and the part distal to the sesamoid bone is known as the patellar ligament (ligamentum patellae), patellar tendon, or infrapatellar tendon.

A schematic is shown of the various patterns of the layers of the quadriceps tendon. (A) A sagittal section of the patella and its relation to the quadriceps and patellar tendons is illustrated. (B) Bilaminar, (C) trilaminar, and (D) quadrilaminar patterns of the quadriceps tendon are shown. R = Rectus femoris, L = Vastus lateralis, M = Vastus medialis, I = Vastus intermedius.
Improvement of surgical techniques and prosthetic designs in procedures such as the TKA requires more detailed analyses of the extensor mechanism. Such analyses must stem from refined and realistic assumptions that are based on foundational documentation of variations in quadriceps muscle forces and directionality. There is considerable evidence that alterations in the forces generated by the quadriceps bellies can affect patellofemoral joint function [1]. Parameters such as the physiologic cross-sectional area, a proxy measure for potential muscle force, and the angles formed by the fibers of the VMO and VLO and the longitudinal axis had significant variation among individuals in a cadaver study [12]. In addition, the quality of the static structures of the knee such as the medial and lateral retinacula and the closely associated patellofemoral ligaments appear to have a significant role in joint stability [2, 13]. If this is true with the large variations studied so far, it is reasonable to hypothesize that smaller variations would have similar, if more subtle, effects. However, to the best of our knowledge, this has not been investigated even though anatomic variations in the tendinous insertions of the quadriceps heads on and around the patella, and associated soft tissues, might affect the mechanics of the joint.
We examined the normal variability of several anatomic structures that may influence knee mechanics: (1) the presence or absence of distinct oblique portions of the VM and VL; (2) the anteroposterior (AP) relationships of the four quadriceps tendon components, reflected in their laminar patterns; and (3) the presence or absence of MPFL and LPFL.
The following is an overview and our collation of numerous published descriptions of the quadriceps insertion on and around the patella.
The Superficial Layer of the Quadriceps Tendon
The RF, the most superficial muscle of the quadriceps group, inserts into the anterior portion of the base and the superior third of the anterior surface of the patella [3, 13, 40] in one of three possible ways. One description is the RF tendon attaches in part to the patella and the remainder continues superficial to the patella to join the ligamentum patellae [10]. A second interpretation is the RF tendon fuses with the tendon of VI to form a unilaminar suprapatellar tendon, either receiving some fibers from the VL and VM [18] or separately from the VL and VM tendons’ attachment to the patella [21]. A third description of the superficial quadriceps tendon insertion is it joins the tendons of the other quadriceps muscles to form a trilaminar tendon [9, 13]. The superficial fibers of this common tendinous attachment (unilaminar or trilaminar) continue over the patella and become continuous with the patellar ligament [9, 12, 13, 21, 28, 37, 38, 40].
The Intermediate Layer of the Quadriceps Tendon
Centrally, the VL and VM unite to form a continuous aponeurosis that inserts into the base of the patella, just posterior to the insertion of RF, and also continues laterally and medially to insert into the sides of the patella [13].
More laterally, the VL ends in an aponeurosis that blends with the lateral side of the suprapatellar [21] or RF [18] tendon and sends an expansion distally to the superolateral side of the patella [3, 4, 12, 18, 37]. Other fibers cross superficial to the patella and attach to the medial condyle of the tibia [9]. The lateral expansion of VL then blends with the capsule of the knee [21], thereby forming part of the lateral patellar retinaculum [3, 4, 18, 37].
Most fibers of the VM end in an aponeurosis that blends with the medial side of the suprapatellar tendon [21] or the RF tendon [13, 31]. The most distal fibers attach directly to the medial edge of the patella and extend more distally than fibers originating from any other part of the quadriceps group [10, 37]. From the distal edge of the VM, a tendinous expansion passes along the medial side of the patella. Deep fibers of this expansion reinforce the joint capsule as part of the medial patellar retinaculum [3, 4, 12, 13, 18, 46]. The obliquely oriented fibers derived from the more superficial VMO contribute to medial patellar stability through their contribution to the superficial layer of the MPFL [28]. Other tendinous fibers pass obliquely across the patella to attach to the lateral tibial condyle [9].
The most distal muscle fibers of the VM often are referred to as the VMO [3, 12, 13, 19, 20, 31, 44]. Unlike the situation with the VLO, it often is difficult to discriminate between the oblique and longitudinal heads of the VM because of the variability in the presence of a division such as a thin layer of fat or fascia or a nerve branch between the two heads [13, 18, 31, 44]. There is controversy regarding whether the VMO is a separate muscle with separate innervation because some anatomic studies fail to support such an idea [19, 22, 31]. Andrikoula et al. [3] claim the presence of a fascial plane mentioned in other studies is the result of a distal-proximal dissection that does not allow one to distinguish between a fascial separation and the lateral portion of a tunnel surrounding the sartorius.
Although surgeons and anatomists often accept the presence of VMO, little attention is paid to the VLO [13]. This is an anatomically distinct group of VL fibers, which is separated from the main belly of the VL by either a thin layer of fat or loose fascia [13, 16] or by a distinct change in fiber orientation [12]. Some describe this muscle group as originating from the lateral intermuscular septum [13], whereas others describe its origin as being from the iliotibial tract (ITT) [12]. These fibers are believed to be important because they provide a more direct lateral pull on the extensor mechanism [13], because they have a more lateral insertion on the patella than the longitudinal head of the VL [12, 44].
The Deep Layer of the Quadriceps Tendon
The VI has an intimate origin with VL proximally and the lateral intermuscular septum distally. It inserts through a broad, thin tendon into the base of the patella posterior to the VL and VM [13, 18, 40] or merged with them [10]. Jones [21] stated the VI also may join with the RF to form the suprapatellar tendon, which inserts on the base of the patella. Medially and laterally, this insertion reinforces the patellofemoral ligaments [13].
Quadriceps Tendon as a Whole
Figure 2 is a scheme illustrating the approximate areas of attachment of the quadriceps tendon components onto the base and sides of the patella. These areas are derived from previously published descriptions by Brizon and Castaing [9] and Segal and Jacob [35]. Fibers that run across the anterior surface of the patella to join the patellar ligament, or which cross the anterior surface diagonally, are not shown.

The approximate areas of attachment of the quadriceps tendon components onto the base and sides of the patella are superimposed on photographs of the bone. RF = Rectus femoris, VL = Vastus lateralis, VM = Vastus medialis, VI = Vastus intermedius, L = Lateral, S = Superior, M = Medial.
The four muscular elements of the quadriceps fuse to form the quadriceps tendon [3, 4, 9, 12, 13, 18, 37, 39, 40] 2 cm proximal to the patella [16]. Some authors suggest the three layers of the quadriceps tendon remain distinct to their insertion into the patella [13, 47]. In addition, MRI has shown the tendon has more lamination in its midline and medial aspects than its lateral aspects [47]. The quadriceps tendon includes contributions from the RF superficially, the VM and VL intermediately, and the VI deeply [3, 4, 9, 37]. Zeiss et al. [47] described the superficial layer of the quadriceps tendon as originating from the thickening of the deep fascia posterior to the RF, the deep layer from the anterior thickening of the deep fascia anterior to the VI, and the middle layers originating from the thickening of the fascia posterior to the VM and VL. The tendinous fibers of the three layers crisscross and meet along their edges to insert into the proximal part of the patella [9]. However, Standring [39] did not describe the quadriceps tendon as having any laminations, but rather said the components come together to form one structure attached to the sides and base of the patella.
Although the usual description of the quadriceps tendon includes three layers, a MRI study [47] revealed only 56% of the subjects presented with a trilaminar appearance. Thirty percent presented with only two different fiber planes, 5% presented with a four-layered quadriceps tendon, and 7% were classified as having one layer because no fascial boundaries could be distinguished. In addition, Zeiss et al. [47] reported each layer of the quadriceps tendon is composed of fibers from more than one of the quadriceps muscles. For example, the intermediate layer usually is described as receiving fibers from the tendons of the VL and VM, but according to Zeiss et al. [47], the intermediate layer also received contributions from the RF and VI. Moreover, Zeiss et al. [47] noted the thickness of the fascial layers inserting into the patella correlated to the number of fiber layers joining to form each one. However, because the MRI slices were 3 or 5 mm thick, partial volume effects could have obscured some fascial planes and made it appear there were fewer layers than actually existed.
Fibrous Layers Superficial to the Quadriceps Tendon and Related Structures
The fibrous layers that cover the knee anteriorly consist of three separate, distinguishable layers [3, 8]. The deep anterior fibrous layer, which is actually medial and lateral to the patella, is made up of the patellofemoral [40] and meniscopatellar ligaments [8]. The MPFL is larger and stronger than the LPFL [8, 40].
The intermediate layer is subdivided into deep and superficial midline layers. The deep midline layer is comprised of the fibers creating the quadriceps and patellar tendons and the crossed fibers of the VM and VL traveling to the tibial condyles of the opposite side (Figs. 3, 4) [3, 12, 40]. Laterally, the superficial midline layer is a thick extension of the ITT. Medially, the superficial midline layer is a thin expansion from the sartorius that connects to the fascia lata [8].

Tendon fibers from the VL run inferomedially across the patella (*) toward the medial tibial condyle. S = Superior, M = Medial.

Tendon fibers from the VM (arrows) run inferolaterally across the patella toward the lateral tibial condyle. A = Anterior, I = Inferior.
The superficial aponeurotic layer is a continuation of the fascia lata proximally and the crural fascia distally [8].
Because casual examinations have led us to believe formation of the quadriceps tendon often is quite different from the way it usually is described, we performed an anatomic study to describe variations in the quadriceps insertion on the patella and to describe other soft tissues of the extensor mechanism.
Materials and Methods
We selected 20 lower limbs from 17 individuals from among cadavers that previously had been dissected by students in the anatomy laboratory at Drexel University College of Medicine; in no case did the dissections affect the anterior tissues superficial to the knee. The cadaveric specimens were from 11 females and six males; the mean age was 86.3 ± 10.3 years; 11 specimens came from the right limb and nine specimens came from the left limb. The specimens had been embalmed with a solution containing formalin, phenol, glycerin, and ethanol. All were inspected visually to ensure the extensor mechanism of the knee was intact and not disturbed. A section of the limb including the knee was collected by cutting through it at midthigh and approximately 15 cm distal to the tibial tubercle. All bodies had been donated for education and research through the Humanity Gifts Registry of the Commonwealth of Pennsylvania (formerly the Anatomical Board), and this study was approved by the Drexel University Institutional Review Board.
All remaining skin and superficial fascia was stripped away until the fascia lata and crural fascia were visible. These fascial layers then were dissected to reveal the fascia contributing to the extensor mechanism. The two sublayers of the intermediate fascial layer could not be identified in some specimens, but fibers that could be distinguished were noted.
Once the epimysium of the muscles was bluntly dissected away, the muscle fiber orientations were noted and documented as were the presence of distinct oblique and longitudinal heads of VM and VL. The RF then was bisected longitudinally using a pair of scissors to the point where it joined the common tendon. The fused VM and VL tendons were bisected using scissors to the point where they reached the common tendon of the quadriceps femoris. The laminar appearance, composition, and order were observed and recorded.
On the medial and lateral aspects of the thigh, the deep fibrous layer was bluntly dissected to search for the MPFL and LPFL. They were noted as being present when there was a distinct horizontal thickening in the deep fibrous layer that was not easily destroyed with blunt dissection.
Once all fibrous layers were described and recorded, we bisected the quadriceps tendon to the base of the patella and completely bisected the patella vertically by cutting through it with a razor saw. The two halves were spread so the composition of the fibrous layers and their insertion into the patella could be described and recorded.
Results
We observed no discernible fat or fascial plane separating the oblique and longitudinal heads of the VM in any of the specimens, although a progressive change in fiber orientation led to a distinct difference between the proximal and distal fibers. Twelve of the 20 specimens had an independent and identifiable group of obliquely oriented muscle fibers originating from the lateral intermuscular septum in the distal thigh forming the VLO (Table 1; Fig. 5). These fibers were separated from the main body of VL by a thin plane of fatty tissue. In 11 of those specimens, these oblique fibers blended with the common tendon of the quadriceps femoris at the superolateral corner of the patella. In the twelfth, the muscle fibers blended with the tendinous part of VL to ultimately insert in that same area.

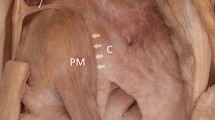
The LPFL is lifted by the hemostat. The curved arrows indicate the fascial plane separating the oblique and longitudinal heads of the VL. A = Anterior, I = Inferior.
Of the 20 cadaveric specimens, six had a bilaminar quadriceps tendon (Table 1; Figs. 1, 6) rather than a trilaminar tendon. There were three distinct lamination patterns in the bilaminar quadriceps tendons. In four of the specimens, the VM and VL joined the RF to create the superficial layer, whereas the VI alone contributed to the deep layer. In a fifth specimen, all four muscle groups sent contributing fibers to the superficial lamina, whereas the VI was the sole contributor to the deep lamina. The sixth bilaminar quadriceps tendon was characterized by the VM contributing to the superficial and deep layers.

This is an example of a two-layered quadriceps tendon whose superficial (S) and deep (D) layers fuse proximal to the patella. Articularis genus (lower curved arrow) and fibers from the VM (upper curved arrow) can be seen inserting into the suprapatellar bursa (*). A = Anterior, S = Superior.
Eleven specimens had a trilaminar quadriceps tendon, but only three of those tendons had the traditionally described three-layered arrangement [9–11, 13, 34, 35, 40] with the superficial layer consisting of RF, intermediate layer of the VM and VL, and deep layer of the VI (Table 1; Figs. 1, 7, 8). The other eight had a more complex organization of muscle fibers (Table 1; Fig. 1). One specimen had a fiber arrangement in which the VL was the only muscle contributing to the intermediate layer, because the fibers of the VM joined those of the RF to create the superficial layer. In four other specimens, the tendinous layers from the VM and VL contributed to the superficial and intermediate layers of the quadriceps tendon. One specimen had a lamination pattern in which the RF contributed to the superficial and intermediate quadriceps layers. The superficial layer of two of the quadriceps tendons contained fibers from the RF and VM. In one of these specimens, the VI split and joined the VM and VL to create the intermediate layer.

In this three-layered quadriceps tendon, the layers remain distinct until they come together and fuse into one tendon proximal to the patella. The superficial (S), intermediate (I), and deep (D) layers are indicated. Su = Superior.

In this specimen with a three-layered quadriceps tendon, the layers fuse into a single tendon proximal to the patella, but a bundle of fibers (unlabeled short arrow) first passes from the VI to the intermediate layer joining the VL and VM. The superficial (S), intermediate (I), and deep (D) layers are indicated. Su = Superior.
Three of the specimens had a quadriceps tendon comprised of four distinct fascial layers (Table 1; Fig. 1). Two of these specimens had a similar arrangement with the RF and the superficial VM fibers combining to create the superficial layer, the VL alone creating a second layer, the remaining part of the VM constituting the third layer, and the VI becoming the deep layer. The third specimen with a tetralaminar quadriceps arrangement had the RF forming the superficial layer and the VM in the intermediate layer. The VL, however, split to create two intermediate layers, one of which blended with the VM in the midline and one of which was deeper. The VI formed the deepest layer.
In all specimens, the quadriceps tendon was most laminated proximally and in the midline. The tendons of the quadriceps muscles first began to fuse several centimeters proximal to the patella on the medial and lateral sides; and as the tendinous layers continued distally, they fused closer to and then in the midline. Some variability was observed at the quadriceps tendon’s insertion into the patella. At its insertion, it always is composed of one common tendon (Figs. 6–8), except for two specimens in which distinct fascial planes separated the laminae. There also was variability in the distance from the patella at which the fascial planes fused into one bundle in the midline. These distances ranged from less than 1 cm to approximately 5.5 cm. Once the layers fused to form a common tendon, it generally remained intact all the way to its insertion on and around the patella. Nevertheless, in the common tendon, there could be some variability in the fascial planes. There was one specimen in which a group of fibers from the intermediate layer joined the deep layer. Another specimen with a bilaminar proximal arrangement had a trilaminar arrangement distally because a loose fascial plane separated the deep layer into two distinguishable planes, but before they reached the patella, all the layers had merged.
The regions of the MPFL and LPFL in 18 of the 20 specimens studied were suitable for determining the presence and nature of the ligaments. Of those excluded, one was from a subject who had a TKA, and in the other, the deep fibrous layer was completely dissected before this study. Twelve of the 18 specimens had intact, observable MPFLs (Table 1; Figs. 9–11), but only one had an intact and observable LPFL (Table 1; Fig. 5). The MPFL and LPFL contributed to the deepest fiber layer of the knee and appeared fused to the fibrous layer of the joint capsule. Each MPFL ran from the medial epicondylar region of the femur near the adductor tubercle to, with one exception, the deep aspect of the VMO and the patella (Fig. 9). In the one exception, all of the MPFL fibers attached directly to the patella. The patellar attachment was, in all specimens, on the deep-medial border of the bone. In the specimen with an MPFL and LPFL present, the MPFL was visibly the larger and stronger of the two.

The medial patellofemoral ligament (MPFL) runs between the medial femoral epicondyle and the superolateral corner of the patella (*). It is partially covered by VM, which sends a fascial expansion to the ligament. A = Anterior, S = Superior.

In this specimen the MPFL is a very wide band attaching to the medial femoral epicondyle anteroinferior to the adductor tubercle (+). VM = vastus medialis, A = Anterior, S = Superior.

The MPFL (arrow) has an extensive connection to the VM before it reaches the patella. + = Adductor tubercle, A = Anterior, S = Superior.
Discussion
This study was done to gather data on the anatomic variability of the extensor apparatus to provide a basis for further investigation of the roles such variations might play in complications of TKA and other procedures. Specifically we explored: (1) the presence or absence of distinct oblique portions of the VM and VL; (2) the AP relationships of the four quadriceps tendon components, reflected in their laminar patterns; and (3) the presence or absence of the MPFL and LPFL.
We acknowledge several limitations. First, connective tissue properties are altered by the fixation process in embalming specimens. This can affect strength of fasciae, but there is no evidence that it alters the visible relationships. Second, this is an elderly subject population. It is possible that the features described could change with time and that different results might be obtained in a young group of knees.
The observations of this study are consistent with those in studies that deny the existence of an observable, anatomically distinct VMO [13, 18, 22, 31]. Nevertheless, the fiber orientation in the distal part of the muscle is noticeably more horizontal than in the proximal part, and there is considerable evidence the distal part has a role in maintaining patellar alignment [24]. The distal part, which can be considered the functional equivalent of the VMO, has its own motor point, is a relatively weak extensor, inserts into the medial border of the patella [23], is more active at 90° flexion than the rest of quadriceps [22], and pulls the patella medially [15, 17, 25]. Although Peeler et al. [31] said there was no sign of a separate nerve entering the VMO, numerous authors disagree [16, 17, 20, 41, 44]. Also, a large number of anatomic atlases and texts have included illustrations that show numerous twigs from the nerve to the VM entering the muscle along its length [7, 29, 33–36, 39, 43]. Therefore, it is reasonable to conclude, even in the absence of a separate, anatomically identifiable muscle which could be called the VMO, that the distal portion of the VM functions differently from the rest and can be thought of as the VMO in a practical context. Our observations support reports of two heads of the VL in some subjects [5, 12, 13, 16]. Although a clear distinction between the heads often can be made, we found no evidence they have separate innervation. The VL is served by one branch of the femoral nerve, which divides and subdivides so four twigs enter the muscle [7, 30]. All of them enter in the upper or middle part, ramifying within the fleshy substance. There was no sign of a separate nerve to the VLO, although one or more of the twigs could have innervated that part. Hollinshead [18] made a similar observation of the VL innervation, although he said there could be as many as three separate branches of the femoral nerve, all entering in the upper third. With electromyography, the VLO, like the VMO, was particularly active at 90° flexion indicating that the two muscles work together to maintain patellar stability [6].
This and previous studies differ in the observation of the variation and complexity of the quadriceps tendon arrangements. The only previous study that mentions variability in the laminar arrangement of the quadriceps tendon was that of Zeiss et al. [47], but although they described similar quadriceps fiber arrangements, the question arises regarding whether these two studies actually are describing the same objects. It is possible that in MRI studies imaging issues such as voxel size and partial volume effects cause other factors to look like the formation of anatomic layers. This can be answered by a study in which knees are imaged and then dissected. Evaluation of the clinical consequences of these anatomic differences may be worth investigating, because differences in the laminations and insertions of the quadriceps into the patella could affect patellar motion through differences in the angle and magnitude of the forces acting on it and thus be a predisposing factor for PFJ dysfunction such as patellar maltracking and TKA failure.
The MPLF could be identified in 66.7% (12 of 18) of our limbs.Various reports, as reviewed by Amis et al. [2], give differing frequencies of its presence for reasons that are not clear. When it is present, it exhibits considerable strength and contributes approximately 55% of the passive soft tissue resistance to lateral patellar subluxation [1]. The MPFL’s intimate relationship with the VMO as observed in this and other studies, its tensile strength, and its prevalence continue to support its role as a strong medial static stabilizer of the patella [3, 13, 16, 28]. In agreement with previous descriptions, the MPFL also was larger and more easily defined during dissection than the LPFL [8]. Other authors imply the LPFL is present in approximately two-thirds of knees [27, 32], therefore we were surprised to see it in only one specimen in our study. One possible explanation is we did not consider it present simply when horizontal fibers were seen in that region; we also used a crude test of strength. Others therefore might have concluded it was more common in our sample. Another possible explanation for the discrepancy could relate to the varying descriptions of its location. Fulkerson and Gossling [14], for example, stated the LPFL is the deep layer of the lateral patellar retinaculum distinct from the fibrous capsular layer of the knee. However, others state it is part of the knee capsule [27, 32, 42]. The LPFL we describe was extracapsular but at least partially fused with the fibrous layer.
TKA designs and surgical techniques have been based largely on assumptions regarding the sizes and proportions of the bony surfaces and a generalized model of extensor mechanism structure and function. Our study showed the extensor mechanism should be redefined in light of the considerable variations we observed. If the retinacular structures, patellofemoral ligaments, and the quadriceps muscles have major variability in bulk, directionality, and resultant force, these factors may have a direct influence on the surgical outcome. The lack of a reliable classification system of extensor mechanism anatomy and its relation to function has prevented testing of this hypothesis. Ultimately, the clinical importance of any of these structures or relationships, alone or in combination, can be assessed only by developing valid noninvasive imaging tools for use in large populations. Willan et al. [45] called for combining current anatomic data sets with advanced imaging techniques such as three-dimensional imaging. Such data, based on imaging of each individual’s structures, could provide a basis for prospective clinical trials of clinical protocols and devices directed at adapting to subtle variations.
References
Amis AA. Current concepts on anatomy and biomechanics of patellar stability. Sports Med Arthrosc. 2007;15:48–56.
Amis AA, Firer P, Mountney J, Senavongse W, Thomas NP. Anatomy and biomechanics of the medial patellofemoral ligament. Knee. 2003;10:215–220.
Andrikoula S, Tokis A, Vasiliadis HS, Georgoulis A. The extensor mechanism of the knee joint: an anatomical study. Knee Surg Sports Traumatol Arthrosc. 2006;14:214–220.
Bencardino JT, Rosenberg ZS, Brown RR, Hassankhani A, Lustrin ES, Beltran J. Traumatic musculotendinous injuries of the knee: diagnosis with MR imaging. Radiographics. 2000;20:S103–S120.
Bevilaqua-Grossi D, Monteiro-Pedro V, Bérzin F. Functional analysis of the patellar stabilizers. Acta Ortop Bras. 2004;12:99–104.
Bevilaqua-Grossi D, Monteiro-Pedro V, Sousa GC, Silva Z, Bérzin F. Contribution to the anatomical study of the oblique portion of the vastus lateralis muscle. Braz J Morphol Sci. 2004;21:47–52.
Brash JC. Neuro-Vascular Hila of Limb Muscles. Edinburgh, United Kingdom: E & S Livingstone Ltd; 1955.
Brizon J, Castaing J. [The Anatomy Sheets, Bundle III, Arthrology of the Limbs] [in French]. Paris, France: Librairie Maloine SA; 1953.
Brizon J, Castaing J. [The Anatomy Sheets, Bundle V, Muscles of the Lower Limb] [in French]. Paris, France: Librairie Maloine SA; 1953.
Bryce TH. Quain’s Elements of Anatomy. Vol IV, Part II, Myology. London, United Kingdom: Longmans Green and Co; 1923.
Cailliet R. Knee Pain and Disability. Philadelphia, PA: FA Davis Company; 1973.
Farahmand F, Senavongse W, Amis AA. Quantitative study of the quadriceps muscles and trochlear groove geometry related to instability of the patellofemoral joint. J Orthop Res. 1998;16:136–143.
Fulkerson JP. Disorders of the Patellofemoral Joint. Ed 4. Philadelphia, PA: Lippincott Williams & Wilkins; 2004.
Fulkerson JP, Gossling HR. Anatomy of the knee joint lateral retinaculum. Clin Orthop Relat Res. 1980;153:183–188.
Goh JCH, Leed PYC, Bose K. A cadaver study of the function of the oblique part of vastus medialis. J Bone Joint Surg Br. 1995;77:225–231.
Grelsamer RP, McConnell J. The Patella—A Team Approach. Gaithersburg, MD: Aspen Publishers; 1998.
Günal I, Araç Ş, Şahinoğlu K, Birvar K. The innervation of vastus medialis obliquus. J Bone Joint Surg Br. 1992;74:624.
Hollinshead H. Anatomy for Surgeons, Vol 3: Back and Limbs. Ed 2. New York, NY: Harper and Row Publishers; 1969.
Hubbard JK, Sampson HW, Elledge JR. Prevalence and morphology of the vastus medialis oblique muscle in human cadavers. Anat Rec. 1997;249:135–142.
Hughston J, Walsh MM, Puddu G. Patellar Subluxation and Dislocation. Philadelphia, PA: WB Saunders; 1984.
Jones FW. Buchanan’s Manual of Anatomy. Ed 8. London, United Kingdom: Baillière Tindall and Cox; 1949.
Lefebvre R, Leroux A, Poumarat G, Galtier B, Guillot M, Vanneuville G, Boucher JP. Vastus medialis: anatomical and functional considerations and implications based upon human and cadaveric studies. J Manipulative Physiol Ther. 2006;29:139–144.
Leroux A, Boucher JP, Poumarat G. Investigation of quadriceps femoris function through electrical stimulation. J Athl Train. 1997;32:115–118.
Lieb FJ, Perry J. Quadriceps function: an anatomical and mechanical study using amputated limbs. J Bone Joint Surg Am. 1968;50:1535–1548.
Lin F, Wang G, Koh JL, Hendrix RW, Zhang LQ. In vivo and noninvasive three-dimensional patellar tracking induced by individual heads of quadriceps. Med Sci Sports Exerc. 2004;36:93–101.
Malek MM. Knee Surgery: Complications, Pitfalls, and Salvage. New York, NY: Springer; 2001.
Merican AM, Amis AA. Anatomy of the lateral retinaculum of the knee. J Bone Joint Surg Br. 2008;90:527–534.
Motamedi K, Seeger LL, Hame SL. Imaging of postoperative knee extensor mechanism. Eur J Radiol. 2005;54:199–205.
Netter FH. Atlas of Human Anatomy. Ed 4. Philadelphia, PA: Saunders Elsevier; 2003.
Patil S, Grigoris P, Shaw-Dunn J, Reece AT. Innervation of vastus lateralis muscle. Clin Anat. 2007;20:556–559.
Peeler J, Cooper J, Porter MM, Thliveris JA, Anderson JE. Structural parameters of the vastus medialis muscle. Clin Anat. 2005;18:281–289.
Reider B, Marshall JL, Koslin B, Ring B, Girgis FG. The anterior aspect of the knee joint: an anatomical study. J Bone Joint Surg Am. 1981;63:351–356.
Romanes GJ. Cunningham’s Textbook of Anatomy. Ed 11. London, United Kingdom: Oxford University Press; 1972.
Schaeffer JP. Morris’ Human Anatomy. Ed 11. New York, NY: The Blackstone Company; 1953.
Segal P, Jacob M. The Knee. Chicago, IL: Year Book Medical Publishers, Inc.; 1983.
Sobotta J. [Atlas of Descriptive Anatomy of Man, Section III. The Nervous and Circulatory Systems and the Sensory Organs of Man] [in German]. München, Germany: JF Lehmann’s Verlag; 1920.
Sonin AH, Fitzgerald SW, Bresler ME, Kirsch MD, Hoff FL, Friedman H. MR imaging appearance of the extensor mechanism of the knee: functional anatomy and injury patterns. Radiographics. 1995;15:367–382.
Staeubli HU, Bollmann C, Kreutz R, Becker W, Rauschning W. Quantification of intact quadriceps tendon, quadriceps tendon insertion, and suprapatellar fat pad: MR arthrography, anatomy, and cryosections in the sagittal plane. AJR Am J Roentgenol. 1999;173:691–698.
Standring S. Gray’s Anatomy. Ed 39. Edinburgh, United Kingdom: Elsevier Churchill Livingstone; 2005.
Testut L, Jacob O. [Treatise of Topographic Anatomy With Medicosurgical Applications] [in French]. Vol II. Paris, France: Octave Doin et Fils Éditeurs; 1914.
Thiranagama R. Nerve supply of the human vastus medialis muscle. J Anat. 1990;170:193–198.
Vieira EL, Vieira EA, da Silva RT, Berlfein PA, Abdalla RJ, Cohen M. An anatomic study of the iliotibial tract. Arthroscopy. 2007;23:269–274.
von Lanz T, Wachsmuth W. [Practical Anatomy, First Volume/Fourth Part] [in German]. Berlin, Germany: Julius Springer; 1938.
Weinstabl R, Scharf W, Firbas W. The extensor apparatus of the knee joint and its peripheral vasti: anatomic investigation and clinical relevance. Surg Radiol Anat. 1989;11:17–22.
Willan PL, Ransome JA, Mahon M. Variability in human quadriceps muscles: quantitative study and review of clinical literature. Clin Anat. 2002;15:116–128.
Yu JS, Petersilge C, Sartoris DJ, Pathria MN, Resnick D. MR imaging of injuries of the extensor mechanism of the knee. Radiographics. 1994;14:541–551.
Zeiss J, Saddemi SR, Ebraheim NA. MR imaging of the quadriceps tendon: normal layered configuration and its importance in cases of tendon rupture. AJR Am J Roentgenol. 1992;159:1031–1034.
Author information
Authors and Affiliations
Corresponding author
Additional information
Each author certifies that he or she has no commercial associations (eg, consultancies, stock ownership, equity interest, patent/licensing arrangements, etc) that might pose a conflict of interest in connection with the submitted article.
Each author certifies that his or her institution has approved the human protocol for this investigation and that all investigations were conducted in conformity with ethical principles of research.
About this article
Cite this article
Waligora, A.C., Johanson, N.A. & Hirsch, B.E. Clinical Anatomy of the Quadriceps Femoris and Extensor Apparatus of the Knee. Clin Orthop Relat Res 467, 3297–3306 (2009). https://doi.org/10.1007/s11999-009-1052-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11999-009-1052-y